

A Comprehensive Review on Collagen Type I Development of Biomaterials for Tissue Engineering: From Biosynthesis to Bioscaffold

 , ,
, ,  , ,
, , 

Abstract
:1. Introduction
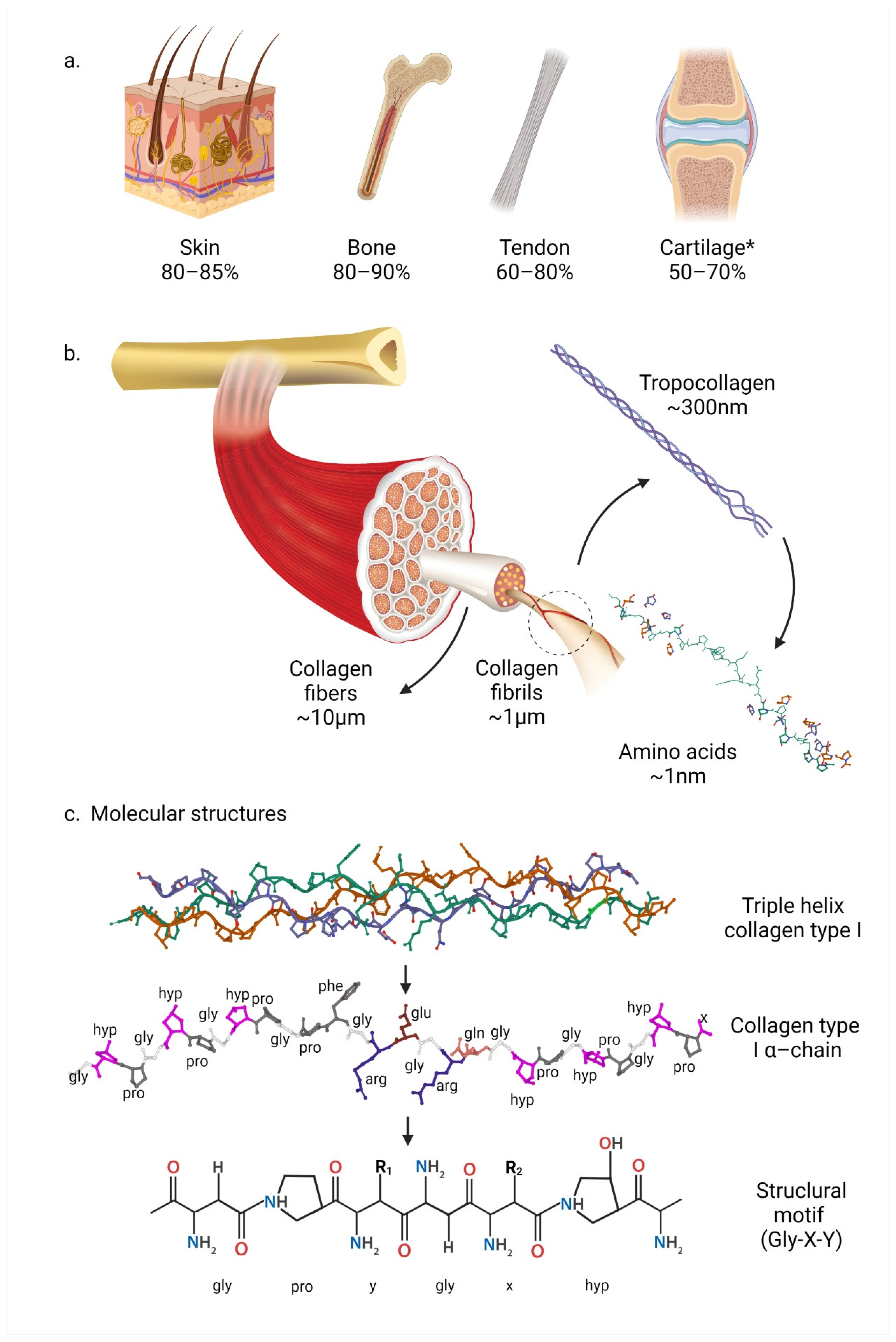
2. Molecular Structure and Biosynthesis of Collagen Type I
2.1. Types of Collagens
2.2. Collagen Type I Supramolecular Structure
2.3. Collagen Type I Molecular Structure and Content
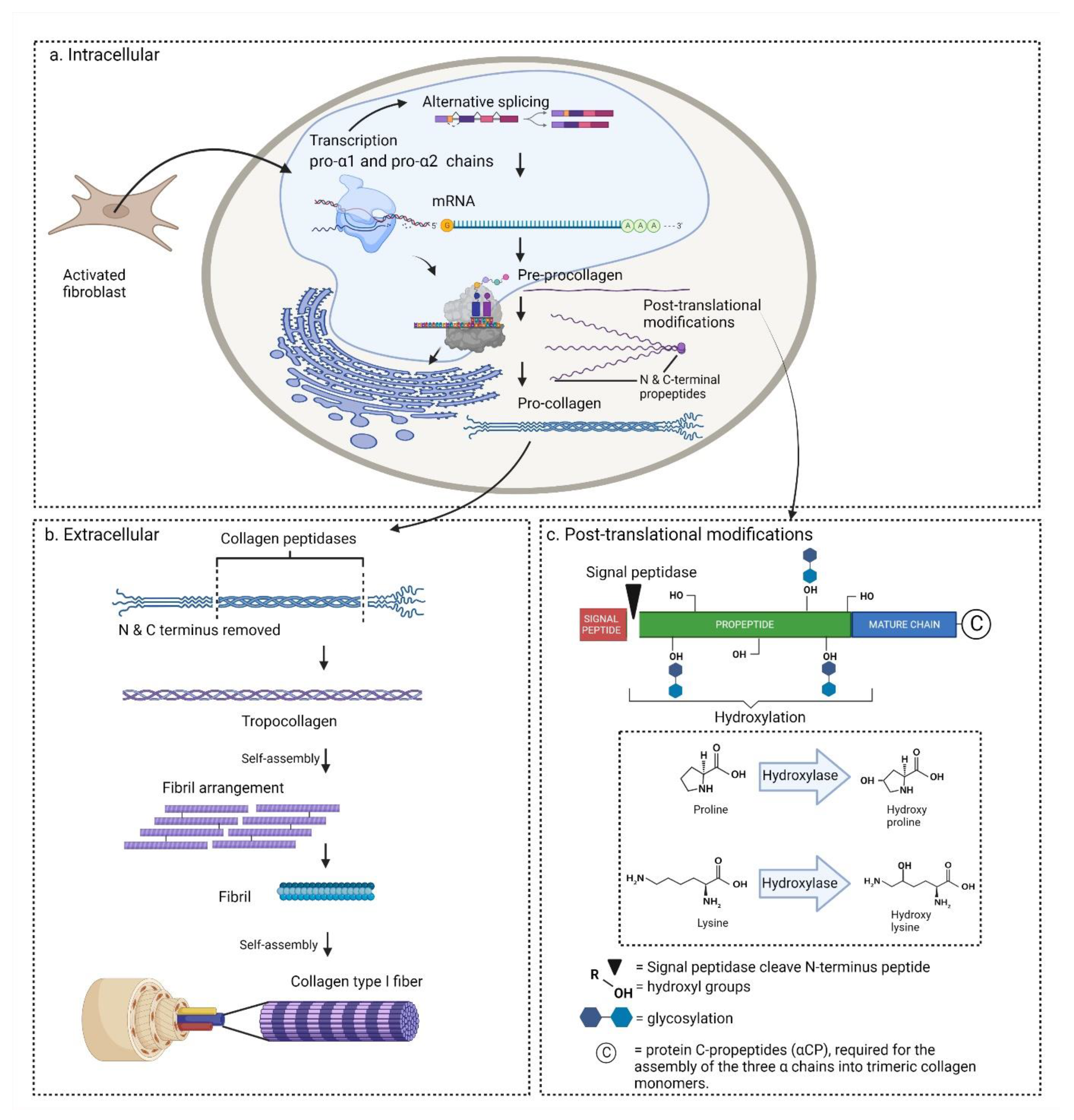
2.4. Biosynthesis
2.4.1. Transcription and Translation
2.4.2. Post-Translational Modifications
2.4.3. Collagen Secretion
2.4.4. Clinical Significance of Biosynthesis for Tissue Engineering
3. Sources of Collagen Type I
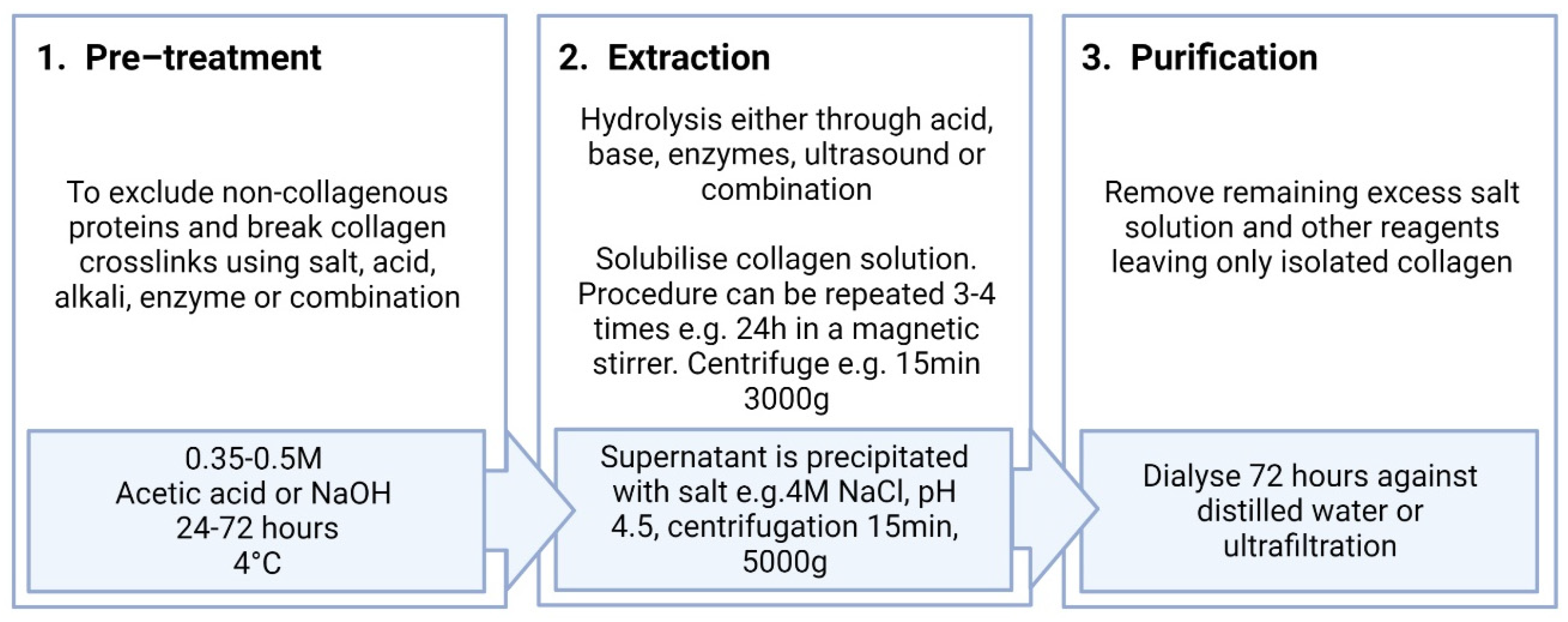
4. Extraction of Collagen Type I
4.1. Salting-Out Method
4.2. Acid Extraction Method
4.3. Alkali Extraction Method
4.4. Enzymatic Extraction Method
4.5. Ultrasound Extraction Method
4.6. Collagen Extraction Process
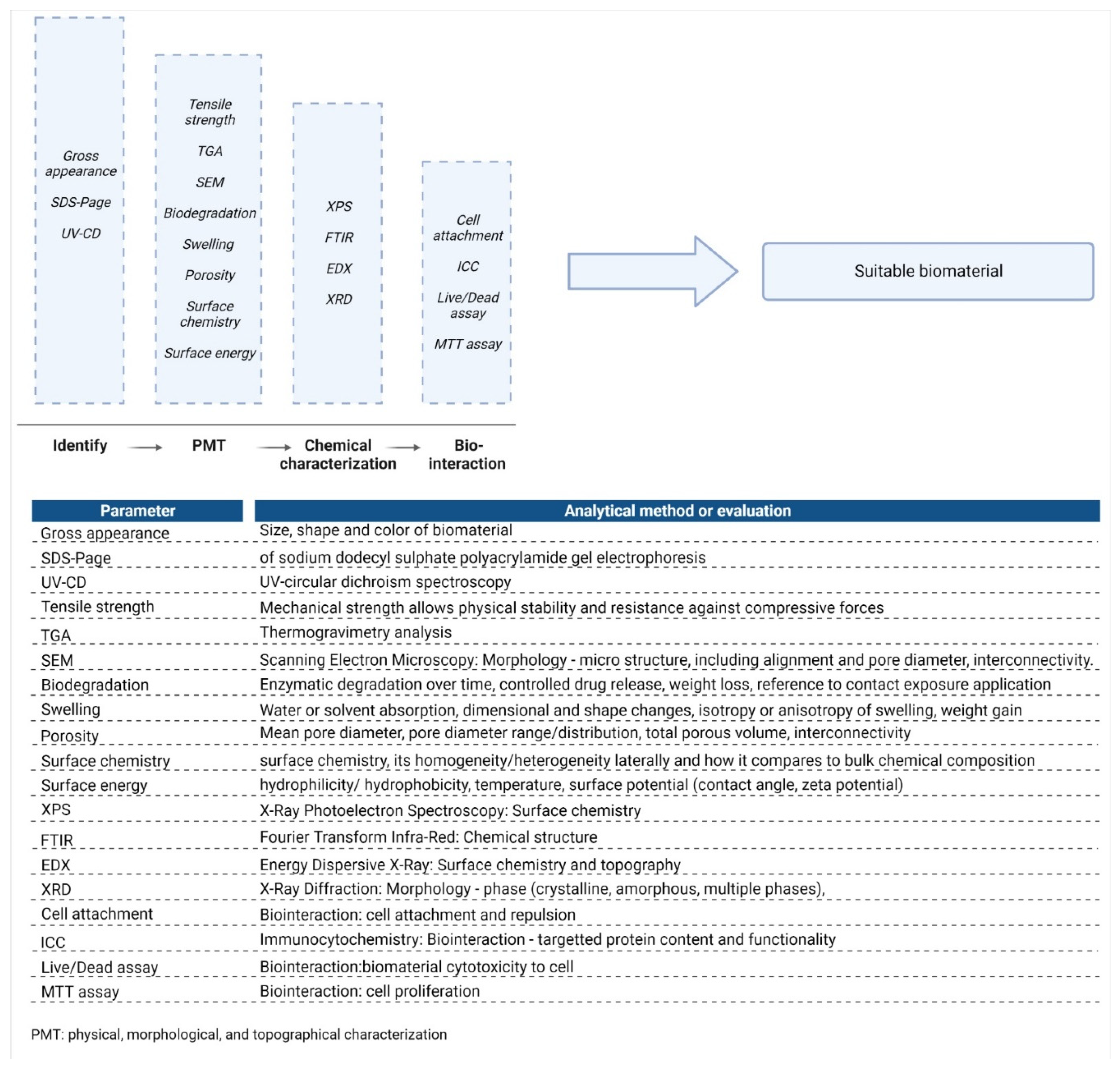
5. Physicochemical Characterisation of Collagen Type I
5.1. Parameters and Test Methodologies for Biomaterial Analysis
5.2. Characterisation of Purified Collagen Type I
5.3. Biocompatibility and Immunogenicity Properties of Collagen Type I
5.4. Three-Dimensional Stability
5.4.1. Mechanical Strength
5.4.2. Thermal Stability: Denaturing Temperature
5.4.3. Porosity and Pore Size
5.4.4. Biodegradation
5.5. Swelling Ratio
5.6. Water Vapour Transmission Rate
5.7. Surface Characteristics
5.8. Chemical Characteristics of Collagen Type I
5.8.1. X-ray Photoelectron Spectroscopy
5.8.2. Fourier Transform Infrared
5.8.3. Energy Dispersive X-ray
5.8.4. X-ray Diffraction
6. Current Insights and Conclusion
6.1. Current Insights over the Last Five Years
6.2. Trends and Future Perspectives
6.3. Conclusions
Author Contributions
Funding

Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sharma, P.; Kumar, P.; Sharma, R.; Bhatt, V.D.; Dhot, P.S. Tissue Engineering; Current Status & Futuristic Scope. J. Med. Life 2019, 12, 225–229. [Google Scholar]
- Cen, L.; Liu, W.; Cui, L.; Zhang, W.; Cao, Y. Collagen tissue engineering: Development of novel biomaterials and applications. Pediatr. Res. 2008, 63, 492–496. [Google Scholar] [CrossRef] [PubMed]
- Wolf, K.; Alexander, S.; Schacht, V.; Coussens, L.M.; von Andrian, U.H.; van Rheenen, J.; Deryugina, E.; Friedl, P. Collagen-based cell migration models in vitro and in vivo. Semin. Cell Dev. Biol. 2009, 20, 931–941. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M. Processing of collagen based biomaterials and the resulting materials properties. Biomed. Eng. Online 2019, 18, 24. [Google Scholar] [CrossRef]
- Ricard-Blum, S. The Collagen Family. Cold Spring Harb. Perspect. Biol. 2011, 3, 1–19. [Google Scholar] [CrossRef]
- Henriksen, K.; Karsdal, M.A. Type I Collagen. In Biochemistry of Collagens, Laminins and Elastin: Structure, Function and Biomarkers; Elsevier Inc.: Amsterdam, The Netherlands, 2016; pp. 1–11. ISBN 9780128098998. [Google Scholar]
- Davison-Kotler, E.; Marshall, W.S.; García-Gareta, E. Sources of collagen for biomaterials in skin wound healing. Bioengineering 2019, 6, 56. [Google Scholar] [CrossRef] [PubMed]
- Fedarko, N.S. Osteoblast/Osteoclast Development and Function in Osteogenesis Imperfecta; Elsevier Inc.: Amsterdam, The Netherlands, 2013; ISBN 9780123971654. [Google Scholar]
- Tresoldi, I.; Oliva, F.; Benvenuto, M.; Fantini, M.; Masuelli, L.; Bei, R.; Modesti, A. Tendon’s ultrastructure. Muscles Ligaments Tendons J. 2013, 3, 2. [Google Scholar] [CrossRef]
- Sophia Fox, A.J.; Bedi, A.; Rodeo, S.A. The basic science of articular cartilage: Structure, composition, and function. Sports Health 2009, 1, 461–468. [Google Scholar] [CrossRef]
- Irawan, V.; Sung, T.C.; Higuchi, A.; Ikoma, T. Collagen Scaffolds in Cartilage Tissue Engineering and Relevant Approaches for Future Development. Tissue Eng. Regen. Med. 2018, 15, 673–697. [Google Scholar] [CrossRef]
- Chen, S.; Mienaltowski, M.J.; Birk, D.E. Regulation of corneal stroma extracellular matrix assembly. Exp. Eye Res. 2015, 133, 69–80. [Google Scholar] [CrossRef]
- Walters, B.D.; Stegemann, J.P. Strategies for directing the structure and function of three-dimensional collagen biomaterials across length scales. Acta Biomater. 2014, 10, 1488–1501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Busra, M.F.B.M.; Chowdhury, S.R.; Bin Ismail, F.; Bin Saim, A.; Idrus, R.B.H. Tissue-engineered skin substitute enhances wound healing after radiation therapy. Adv. Ski. Wound Care 2016, 29, 120–129. [Google Scholar] [CrossRef] [PubMed]
- Bettini, S.; Bonfrate, V.; Syrgiannis, Z.; Sannino, A.; Salvatore, L.; Madaghiele, M.; Valli, L.; Giancane, G. Biocompatible Collagen Paramagnetic Scaffold for Controlled Drug Release. Biomacromolecules 2015, 16, 2599–2608. [Google Scholar] [CrossRef]
- Helary, C.; Abed, A.; Mosser, G.; Louedec, L.; Letourneur, D.; Coradin, T.; Giraud-Guille, M.M.; Meddahi-Pellé, A. Evaluation of dense collagen matrices as medicated wound dressing for the treatment of cutaneous chronic wounds. Biomater. Sci. 2015, 3, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Abou Neel, E.A.; Bozec, L.; Knowles, J.C.; Syed, O.; Mudera, V.; Day, R.; Hyun, J.K. Collagen-Emerging collagen based therapies hit the patient. Adv. Drug Deliv. Rev. 2013, 65, 429–456. [Google Scholar] [CrossRef]
- Chai, H.J.; Li, J.H.; Huang, H.N.; Li, T.L.; Chan, Y.L.; Shiau, C.Y.; Wu, C.J. Effects of sizes and conformations of fish-scale Collagen peptides on facial skin qualities and transdermal penetration efficiency. J. Biomed. Biotechnol. 2010, 2010, 757301. [Google Scholar] [CrossRef]
- Rico-Llanos, G.A.; Borrego-González, S.; Moncayo-Donoso, M.; Becerra, J.; Visser, R. Collagen Type I Biomaterials as Scaffolds for Bone Tissue Engineering. Polymers 2021, 13, 599. [Google Scholar] [CrossRef]
- Antoine, E.E.; Vlachos, P.P.; Rylander, M.N. Review of collagen i hydrogels for bioengineered tissue microenvironments: Characterization of mechanics, structure, and transport. Tissue Eng.-Part B Rev. 2014, 20, 683–696. [Google Scholar] [CrossRef]
- Silvipriya, K.S.; Kumar, K.K.; Bhat, A.R.; Dinesh Kumar, B.; John, A.; James, S. Collagen: Animal Sources and Biomedical Application. J. Appl. Pharm. Sci. 2015, 5, 123–127. [Google Scholar] [CrossRef]
- Lucaciu, O.; Apostu, D.; Mester, A.; Campian, R.S.; Gheban, D.; Miron, R.J. Atelo-collagen type I bovine bone substitute and membrane in guided bone regeneration: A series of clinical cases and histopathological assessments. Histol. Histopathol. 2019, 34, 1061–1071. [Google Scholar] [CrossRef]
- Zhu, B.Z.; Cao, D.S.; Xie, J.; Li, H.H.; Chen, Z.H.; Bao, Q. Clinical experience of the use of Integra in combination with negative pressure wound therapy: An alternative method for the management of wounds with exposed bone or tendon. J. Plast. Surg. Hand Surg. 2021, 55, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Bella, J.; Eaton, M.; Brodsky, B.; Berman, H.M. Crystal and molecular structure of a collagen-like peptide at 1.9 A resolution. Science. 1994, 266, 75–81. [Google Scholar] [CrossRef] [PubMed]
- Engel, J.; Bächinger, H.P. Structure, stability and folding of the collagen triple helix. Top. Curr. Chem. 2005, 247, 7–33. [Google Scholar] [CrossRef]
- Cui, F.Z.; Li, Y.; Ge, J. Self-assembly of mineralized collagen composites. Mater. Sci. Eng. R Reports 2007, 57, 1–27. [Google Scholar] [CrossRef]
- Xu, Y.F.; Nudelman, F.; Eren, E.D.; Wirix, M.J.M.; Cantaert, B.; Nijhuis, W.H.; Hermida-Merino, D.; Portale, G.; Bomans, P.H.H.; Ottmann, C.; et al. Intermolecular channels direct crystal orientation in mineralized collagen. Nat. Commun. 2020, 11, 5068. [Google Scholar] [CrossRef] [PubMed]
- Bou-Gharios, G.; Abraham, D.; Crombrugghe, B. de Type I collagen structure, synthesis, and regulation. In Principles of Bone Biology; Elsevier: Alpharetta, GA, USA, 2020; pp. 295–337. [Google Scholar]
- Chapman, J.A.; Tzaphlidou, M.; Meek, K.M.; Kadler, K.E. The collagen fibril—A model system for studying the staining and fixation of a protein. Electron Microsc. Rev. 1990, 3, 143–182. [Google Scholar] [CrossRef]
- Schmitt, F.O.; Hall, C.E.; Jakus, M.A. Electron microscope investigations of the structure of collagen. J. Cell. Comp. Physiol. 1942, 20, 11–33. [Google Scholar] [CrossRef]
- Bigi, A.; Cojazzi, G.; Roveri, N.; Koch, M.H.J. Differential scanning calorimetry and X-ray diffraction study of tendon collagen thermal denaturation. Int. J. Biol. Macromol. 1987, 9, 363–367. [Google Scholar] [CrossRef]
- Erickson, B.; Fang, M.; Wallace, J.M.; Orr, B.G.; Les, C.M.; Banaszak Holl, M.M. Nanoscale Structure of Type I Collagen Fibrils: Quantitative Measurement of D-spacing. Biotechnol. J. 2013, 8, 117. [Google Scholar] [CrossRef]
- Vidal, B.D.C.; Mello, M.L.S. Collagen type I amide I band infrared spectroscopy. Micron 2011, 42, 283–289. [Google Scholar] [CrossRef]
- Ueda, M.; Saito, S.; Murata, T.; Hirano, T.; Bise, R.; Kabashima, K.; Suzuki, S. Combined multiphoton imaging and biaxial tissue extension for quantitative analysis of geometric fiber organization in human reticular dermis. Sci. Rep. 2019, 9, 10644. [Google Scholar] [CrossRef] [Green Version]
- Gautieri, A.; Vesentini, S.; Redaelli, A.; Buehler, M.J. Hierarchical structure and nanomechanics of collagen microfibrils from the atomistic scale up. Nano Lett. 2011, 11, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Freeman, R.; Boekhoven, J.; Dickerson, M.B.; Naik, R.R.; Stupp, S.I. Biopolymers and supramolecular polymers as biomaterials for biomedical applications. MRS Bull. 2015, 40, 1089–1100. [Google Scholar] [CrossRef]
- Grant, M.E. From collagen chemistry towards cell therapy—A personal journey. Int. J. Exp. Pathol. 2007, 88, 203. [Google Scholar] [CrossRef] [PubMed]
- Brodsky, B.; Persikov, A.V. Molecular structure of the collagen triple helix. Adv. Protein Chem. 2005, 70, 301–339. [Google Scholar] [CrossRef] [PubMed]
- Van Der Rest, M.; Garrone, R. Collagen family of proteins. FASEB J. 1991, 5, 2814–2823. [Google Scholar] [CrossRef]
- Hashim, P.; Mohd Ridzwan, M.S.; Bakar, J.; Mat Hashim, D. Collagen in food and beverage industries. Int. Food Res. J. 2015, 22, 1–8. [Google Scholar]
- Gomez-Guillen, M.C.; Gimenez, B.; Lopez-Caballero, M.E.; Montero, M.P. Functional and bioactive properties of collagen and gelatin from alternative sources: A review. Food Hydrocoll. 2011, 25, 1813–1827. [Google Scholar] [CrossRef]
- Wu, M.; Cronin, K.; Crane, J.S. Biochemistry, Collagen Synthesis; StatPearls: Tampa, FL, USA, 2021. [Google Scholar]
- Stefanovic, B. RNA protein interactions governing expression of the most abundant protein in human body, type I collagen. Wiley Interdiscip. Rev. RNA 2013, 4, 535–545. [Google Scholar] [CrossRef]
- Gelse, K.; Pöschl, E.; Aigner, T. Collagens-Structure, function, and biosynthesis. Adv. Drug Deliv. Rev. 2003, 55, 1531–1546. [Google Scholar] [CrossRef]
- Ramachandran, G. Biochemistry of Collagen; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013; ISBN 9781475746020. [Google Scholar]
- Makareeva, E.; Leikin, S. Collagen Structure, Folding and Function. In Osteogenesis Imperfecta: A Translational Approach to Brittle Bone Disease; Elsevier Inc.: Alpharetta, GA, USA, 2013; pp. 71–84. ISBN 9780123971654. [Google Scholar]
- Yamauchi, M.; Sricholpech, M. Lysine post-translational modifications of collagen. Essays Biochem. 2012, 52, 113–133. [Google Scholar] [CrossRef] [PubMed]
- Kirkness, M.W.; Lehmann, K.; Forde, N.R. Mechanics and structural stability of the collagen triple helix. Curr. Opin. Chem. Biol. 2019, 53, 98–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudson, D.M.; Eyre, D.R. Collagen Prolyl 3-Hydroxylation: A Major Role for a Minor Post-Translational Modification; Taylor & Francis: Abingdon, UK, 2013; Volume 54, pp. 245–251. [Google Scholar]
- Shoulders, M.D.; Raines, R.T. Collagen Structure and Stability. Annu. Rev. Biochem. 2009, 78, 929–958. [Google Scholar] [CrossRef] [PubMed]
- Calonje, E.; Brenn, T.; Lazar, A.; Billings, S.D. McKee’s Pathology of the Skin with Clinical Correlations; Elsevier: Amsterdam, The Netherlands, 2020; p. 1917. [Google Scholar]
- Shaker, J.L.; Albert, C.; Fritz, J.; Harris, G. Recent developments in osteogenesis imperfecta. F1000Research 2015, 4, 681. [Google Scholar] [CrossRef]
- Mortier, G.R.; Cohn, D.H.; Cormier-Daire, V.; Hall, C.; Krakow, D.; Mundlos, S.; Nishimura, G.; Robertson, S.; Sangiorgi, L.; Savarirayan, R.; et al. Nosology and classification of genetic skeletal disorders: 2019 revision. Am. J. Med. Genet. A 2019, 179, 2393–2419. [Google Scholar] [CrossRef]
- Semler, O.; Rehberg, M.; Mehdiani, N.; Jackels, M.; Hoyer-Kuhn, H. Current and Emerging Therapeutic Options for the Management of Rare Skeletal Diseases. Paediatr. Drugs 2019, 21, 95–106. [Google Scholar] [CrossRef]
- Hosseinpour, S.; Rad, M.R.; Khojasteh, A.; Zadeh, H.H. Antibody Administration for Bone Tissue Engineering: A Systematic Review. Curr. Stem Cell Res. Ther. 2018, 13, 292–315. [Google Scholar] [CrossRef]
- Grafe, I.; Yang, T.; Alexander, S.; Homan, E.P.; Lietman, C.; Jiang, M.M.; Bertin, T.; Munivez, E.; Chen, Y.; Dawson, B.; et al. Excessive transforming growth factor-β signaling is a common mechanism in osteogenesis imperfecta. Nat. Med. 2014, 20, 670–675. [Google Scholar] [CrossRef]
- Kołodziejska, B.; Kaflak, A.; Kolmas, J. Biologically inspired collagen/apatite composite biomaterials for potential use in bone tissue regeneration-A review. Materials 2020, 13, 1748. [Google Scholar] [CrossRef]
- Horwitz, E.M.; Gordon, P.L.; Koo, W.K.K.; Marx, J.C.; Neel, M.D.; McNall, R.Y.; Muul, L.; Hofmann, T. Isolated allogeneic bone marrow-derived mesenchymal cells engraft and stimulate growth in children with osteogenesis imperfecta: Implications for cell therapy of bone. Proc. Natl. Acad. Sci. USA 2002, 99, 8932–8937. [Google Scholar] [CrossRef]
- Horwitz, E.M.; Prockop, D.J.; Fitzpatrick, L.A.; Koo, W.W.K.; Gordon, P.L.; Neel, M.; Sussman, M.; Orchard, P.; Marx, J.C.; Pyeritz, R.E.; et al. Transplantability and therapeutic effects of bone marrow-derived mesenchymal cells in children with osteogenesis imperfecta. Nat. Med. 1999, 5, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Götherström, C.; Walther-Jallow, L. Stem Cell Therapy as a Treatment for Osteogenesis Imperfecta. Curr. Osteoporos. Rep. 2020, 18, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Götherström, C.; David, A.L.; Walther-Jallow, L.; Åström, E.; Westgren, M. Mesenchymal Stem Cell Therapy for Osteogenesis Imperfecta. Clin. Obstet. Gynecol. 2021, 64, 898–903. [Google Scholar] [CrossRef] [PubMed]
- Chitty, L.S.; David, A.L.; Gottschalk, I.; Oepkes, D.; Westgren, M.; Götherström, C.; Consortium, O. EP21.04: BOOSTB4: A clinical study to determine safety and efficacy of pre- and/or postnatal stem cell transplantation for treatment of osteogenesis imperfecta. Ultrasound Obstet. Gynecol. 2016, 48 (Suppl. 1), 356. [Google Scholar] [CrossRef]
- Ritelli, M.; Colombi, M. Molecular Genetics and Pathogenesis of Ehlers–Danlos Syndrome and Related Connective Tissue Disorders. Genes 2020, 11, 547. [Google Scholar] [CrossRef]
- Walters, M.J. Birds, Pigs, and People: The Rise of Pandemic Flus. In Seven Modern Plagues; Chapter 7; Island Press: Washington, DC, USA, 2014; pp. 151–173. [Google Scholar] [CrossRef]
- Silva, T.H.; Moreira-Silva, J.; Marques, A.L.P.; Domingues, A.; Bayon, Y.; Reis, R.L. Marine origin collagens and its potential applications. Mar. Drugs 2014, 12, 5881–5901. [Google Scholar] [CrossRef]
- King’ori, A.M. A Review of the uses of poultry eggshells and shell membranes. Int. J. Poult. Sci. 2011, 10, 908–912. [Google Scholar] [CrossRef]
- de Araújo Cordeiro, A.R.R.; Bezerra, T.K.A.; de Queiroz, A.L.M.; de Sousa Galvão, M.; Cavalcanti, M.T.; Pacheco, M.T.B.; Madruga, M.S. Collagen production from chicken keel bone using acid and enzymatic treatment at a temperature of 30 °C. Food Sci. Technol. 2019, 40, 491–497. [Google Scholar] [CrossRef]
- Cliche, S.; Amiot, J.; Avezard, C.; Gariépy, C. Extraction and characterization of collagen with or without telopeptides from chicken skin. Poult. Sci. 2003, 82, 503–509. [Google Scholar] [CrossRef]
- Theng, C.H.; Huda, N.; Muhammad, N.A.N.; Wariyah, C.; Hashim, H. Physicochemical properties of duck feet collagen with different soaking time and its application in surimi. Int. J. Adv. Sci. Eng. Inf. Technol. 2018, 8, 832–841. [Google Scholar] [CrossRef]
- Salvatore, L.; Gallo, N.; Aiello, D.; Lunetti, P.; Barca, A.; Blasi, L.; Madaghiele, M.; Bettini, S.; Giancane, G.; Hasan, M.; et al. An insight on type I collagen from horse tendon for the manufacture of implantable devices. Int. J. Biol. Macromol. 2020, 154, 291–306. [Google Scholar] [CrossRef] [PubMed]
- Fauzi, M.B.; Lokanathan, Y.; Aminuddin, B.S.; Ruszymah, B.H.I.; Chowdhury, S.R. Ovine tendon collagen: Extraction, characterisation and fabrication of thin films for tissue engineering applications. Mater. Sci. Eng. C 2016, 68, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Fessler, J.H. Some properties of neutral-salt-soluble collagen. 2. Biochem. J. 1960, 76, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Mocan, E.; Tagadiuc, O.; Nacu, V. Aspects of collagen isolation procedure. Clin. Res. Stud. 2011, 2, 8–10. [Google Scholar]
- Zhang, J.; Duan, R. Characterisation of acid-soluble and pepsin-solubilised collagen from frog (Rana nigromaculata) skin. Int. J. Biol. Macromol. 2017, 101, 638–642. [Google Scholar] [CrossRef]
- Noorzai, S.; Verbeek, C.J.R. Collagen: From Waste to Gold. In Biotechnological Applications of Biomass; Chapter 12; IntechOpen: London, UK, 2020; pp. 1–36. [Google Scholar] [CrossRef]
- Skopinska-Wisniewska, J.; Olszewski, K.; Bajek, A.; Rynkiewicz, A.; Sionkowska, A. Dialysis as a method of obtaining neutral collagen gels. Mater. Sci. Eng. C 2014, 40, 65–70. [Google Scholar] [CrossRef]
- Lin, X.; Chen, Y.; Jin, H.; Zhao, Q.; Liu, C.; Li, R.; Yu, F.; Chen, Y.; Huang, F.; Yang, Z.; et al. Collagen Extracted from Bigeye Tuna (Thunnus obesus) Skin by Isoelectric Precipitation: Physicochemical Properties, Proliferation, and Migration Activities. Mar. Drugs 2019, 17, 261. [Google Scholar] [CrossRef]
- Wang, L.; Yang, B.; Du, X.; Yang, Y.; Liu, J. Optimization of conditions for extraction of acid-soluble collagen from grass carp (Ctenopharyngodon idella) by response surface methodology. Innov. Food Sci. Emerg. Technol. 2008, 9, 604–607. [Google Scholar] [CrossRef]
- Liu, D.; Wei, G.; Li, T.; Hu, J.; Lu, N.; Regenstein, J.M.; Zhou, P. Effects of alkaline pretreatments and acid extraction conditions on the acid-soluble collagen from grass carp (Ctenopharyngodon idella) skin. Food Chem. 2015, 172, 836–843. [Google Scholar] [CrossRef]
- Schmidt, M.M.; Dornelles, R.C.P.P.; Mello, R.O.; Kubota, E.H.; Mazutti, M.A.; Kempka, A.P.; Demiate, I.M. Collagen extraction process. Int. Food Res. J. 2016, 23, 913–922. [Google Scholar]
- Chen, J.; Li, L.; Yi, R.; Xu, N.; Gao, R.; Hong, B. Extraction and characterization of acid-soluble collagen from scales and skin of tilapia (Oreochromis niloticus). LWT-Food Sci. Technol. 2016, 66, 453–459. [Google Scholar] [CrossRef]
- Yang, H.; Shu, Z. The extraction of collagen protein from pig skin. J. Chem. Pharm. Res. 2014, 6, 683–687. [Google Scholar]
- de Almeida, P.F.; de Araújo, M.G.O.; Santana, J.C.C. Obtenção de colágeno a partir de tarsos de frangos para produção de geleias. Acta Sci.-Technol. 2012, 34, 345–351. [Google Scholar] [CrossRef] [Green Version]
- Hattori, S.; Adachi, E.; Ebihara, T.; Shirai, T.; Someki, I.; Irie, S. Alkali-treated collagen retained the triple helical conformation and the ligand activity for the cell adhesion via α2β1 integrin. J. Biochem. 1999, 125, 676–684. [Google Scholar] [CrossRef]
- Wang, L.; Liang, Q.; Chen, T.; Wang, Z.; Xu, J.; Ma, H. Characterization of collagen from the skin of Amur sturgeon (Acipenser schrenckii). Food Hydrocoll. 2014, 38, 104–109. [Google Scholar] [CrossRef]
- Hashemi, S.M.B.; Mousavi Khaneghah, A.; Koubaa, M.; Barba, F.J.; Abedi, E.; Niakousari, M.; Tavakoli, J. Extraction of essential oil from Aloysia citriodora Palau leaves using continuous and pulsed ultrasound: Kinetics, antioxidant activity and antimicrobial properties. Process Biochem. 2018, 65, 197–204. [Google Scholar] [CrossRef]
- Zou, Y.; Yang, H.; Zhang, X.; Xu, P.; Jiang, D.; Zhang, M.; Xu, W.; Wang, D. Effect of ultrasound power on extraction kinetic model, and physicochemical and structural characteristics of collagen from chicken lung. Food Prod. Process. Nutr. 2020, 2, 3. [Google Scholar] [CrossRef]
- Akram, A.N.; Zhang, C. Extraction of collagen-II with pepsin and ultrasound treatment from chicken sternal cartilage; physicochemical and functional properties. Ultrason. Sonochem. 2020, 64, 105053. [Google Scholar] [CrossRef]
- Petcharat, T.; Benjakul, S.; Karnjanapratum, S.; Nalinanon, S. Ultrasound-assisted extraction of collagen from clown featherback (Chitala ornata) skin: Yield and molecular characteristics. J. Sci. Food Agric. 2021, 101, 648–658. [Google Scholar] [CrossRef]
- Ran, X.G.; Wang, L.Y. Use of ultrasonic and pepsin treatment in tandem for collagen extraction from meat industry by-products. J. Sci. Food Agric. 2014, 94, 585–590. [Google Scholar] [CrossRef]
- Karami, A.; Tebyanian, H.; Soufdoost, R.S.; Motavallian, E.; Barkhordari, A.; Nourani, M.R. Extraction and Characterization of Collagen with Cost-Effective Method from Human Placenta for Biomedical Applications. World J. Plast. Surg. 2019, 8, 352. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.H.; Silva, R.M.; Dumont, V.C.; Neves, J.S.; Mansur, H.S.; Heneine, L.G.D. Extraction and characterization of highly purified collagen from bovine pericardium for potential bioengineering applications. Mater. Sci. Eng. C 2013, 33, 790–800. [Google Scholar] [CrossRef] [PubMed]
- Techatanawat, S.; Surarit, R.; Suddhasthira, T.; Khovidhunkit, S.O.P. Type I collagen extracted from rat-tail and bovine Achilles tendon for dental application: A comparative study. Asian Biomed. 2011, 5, 787–798. [Google Scholar] [CrossRef]
- Banerjee, I.; Mishra, D.; Das, T.; Maiti, S.; Maiti, T.K. Caprine (Goat) collagen: A potential biomaterial for skin tissue engineering. J. Biomater. Sci. Polym. Ed. 2012, 23, 355–373. [Google Scholar] [CrossRef]
- Pati, F.; Datta, P.; Adhikari, B.; Dhara, S.; Ghosh, K.; Mohapatra, P.K. Das Collagen scaffolds derived from fresh water fish origin and their biocompatibility. J. Biomed. Mater. Res.-Part A 2012, 100A, 1068–1079. [Google Scholar] [CrossRef]
- Liu, D.; Liang, L.; Regenstein, J.M.; Zhou, P. Extraction and characterisation of pepsin-solubilised collagen from fins, scales, skins, bones and swim bladders of bighead carp (Hypophthalmichthys nobilis). Food Chem. 2012, 133, 1441–1448. [Google Scholar] [CrossRef]
- Wang, J.; Pei, X.; Liu, H.; Zhou, D. Extraction and characterization of acid-soluble and pepsin-soluble collagen from skin of loach (Misgurnus anguillicaudatus). Int. J. Biol. Macromol. 2018, 106, 544–550. [Google Scholar] [CrossRef]
- Uriarte-Montoya, M.H.; Arias, J.; Plascencia-Jatomea, M.; Santacruz, H.; Rouzaud-Sández, O.; Cardenas-Lopez, J.L.; Rios, E.M.; Brauer, J.M.E. Jumbo squid (Dosidicus gigas) mantle collagen: Extraction, characterization, and potential application in the preparation of chitosan-collagen biofilms. Bioresour. Technol. 2010, 101, 4212–4219. [Google Scholar] [CrossRef]
- Addad, S.; Exposito, J.-Y.; Faye, C.; Ricard-Blum, S.; Lethias, C. Isolation, characterization and biological evaluation of jellyfish collagen for use in biomedical applications. Mar. Drugs 2011, 9, 967–983. [Google Scholar] [CrossRef]
- Jankangram, W.; Chooluck, S.; Pomthong, B. Comparison of the properties of collagen extracted from dried jellyfish and dried squid. African J. Biotechnol. 2016, 15, 642–648. [Google Scholar] [CrossRef]
- Wang, T.; Lin, S.; Shen, Y.; Liu, S.; Wang-McCall, T.; Chin, M.; Lin, T.; Yang, C.; Wu, W.; Yang, C. Type I collagen from bullfrog (Rana catesbeiana) fallopian tube. African J. Biotechnol. 2011, 10, 8414–8420. [Google Scholar] [CrossRef]
- Peng, Y.Y.; Glattauer, V.; Ramshaw, J.A.M.; Werkmeister, J.A. Evaluation of the immunogenicity and cell compatibility of avian collagen for biomedical applications. J. Biomed. Mater. Res.-Part A 2010, 93, 1235–1244. [Google Scholar] [CrossRef]
- Parenteau-Bareil, R.; Gauvin, R.; Cliche, S.; Gariépy, C.; Germain, L.; Berthod, F. Comparative study of bovine, porcine and avian collagens for the production of a tissue engineered dermis. Acta Biomater. 2011, 7, 3757–3765. [Google Scholar] [CrossRef] [PubMed]
- Fauzi, M.B.; Lokanathan, Y.; Nadzir, M.M.; Aminuddin, S.; Ruszymah, B.H.I.; Chowdhury, S.R.; Ramanauskiene, K.; Stelmakiene, A.; Briedis, V.; Ivanauskas, L.; et al. Attachment, proliferation, and morphological properties of human dermal fibroblasts on ovine tendon collagen scaffolds: A comparative study. Malaysian J. Med. Sci. 2017, 24, 33–43. [Google Scholar] [CrossRef]
- Thievessen, I.; Fakhri, N.; Steinwachs, J.; Kraus, V.; McIsaac, R.S.; Gao, L.; Chen, B.C.; Baird, M.A.; Davidson, M.W.; Betzig, E.; et al. Vinculin is required for cell polarization, migration, and extracellular matrix remodeling in 3D collagen. FASEB J. 2015, 29, 4555–4567. [Google Scholar] [CrossRef] [PubMed]
- Parenteau-Bareil, R.; Gauvin, R.; Berthod, F. Collagen-based biomaterials for tissue engineering applications. Materials 2010, 3, 1863–1887. [Google Scholar] [CrossRef]
- Inanc, S.; Keles, D.; Oktay, G. An improved collagen zymography approach for evaluating the collagenases MMP-1, MMP-8, and MMP-13. Biotechniques 2017, 63, 174–180. [Google Scholar] [CrossRef]
- Carvalho, A.M.; Marques, A.P.; Silva, T.H.; Reis, R.L. Evaluation of the potential of collagen from codfish skin as a biomaterial for biomedical applications. Mar. Drugs 2018, 16, 495. [Google Scholar] [CrossRef]
- Amri, M.A.; Firdaus, M.A.B.; Fauzi, M.B.; Chowdhury, S.R.; Fadilah, N.R.; Wan Hamirul, W.K.; Reusmaazran, M.Y.; Aminuddin, B.S.; Ruszymah, B.H.I. Cytotoxic evaluation of biomechanically improved crosslinked ovine collagen on human dermal fibroblasts. Biomed. Mater. Eng. 2014, 24, 1715–1724. [Google Scholar] [CrossRef]
- Teramoto, N.; Hayashi, A.; Yamanaka, K.; Sakiyama, A.; Nakano, A.; Shibata, M. Preparation and mechanical properties of photo-crosslinked fish gelatin/imogolite nanofiber composite hydrogel. Materials 2012, 5, 2573–2585. [Google Scholar] [CrossRef]
- Miles, C.A.; Bailey, A.J. Thermal Denaturation of Collagen Revisited. Proc. Indian Acad. Sci. Chem. Sci. 1999, 111, 71–80. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, S.; Shen, L.; Li, G. Factors affecting thermal stability of collagen from the aspects of extraction, processing and modification. J. Leather Sci. Eng. 2020, 2, 19. [Google Scholar] [CrossRef]
- Subhan, F.; Ikram, M.; Shehzad, A.; Ghafoor, A. Marine Collagen: An Emerging Player in Biomedical applications. J. Food Sci. Technol. 2015, 52, 4703–4707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bozec, L.; Odlyha, M. Thermal Denaturation Studies of Collagen by Microthermal Analysis and Atomic Force Microscopy. Biophys. J. 2011, 101, 228. [Google Scholar] [CrossRef]
- Ghodbane, S.A.; Dunn, M.G. Physical and mechanical properties of cross-linked type I collagen scaffolds derived from bovine, porcine, and ovine tendons. J. Biomed. Mater. Res.-Part A 2016, 104, 2685–2692. [Google Scholar] [CrossRef]
- Mh Busra, F.; Rajab, N.F.; Tabata, Y.; Saim, A.B.; Idrus, R.B.H.; Chowdhury, S.R. Rapid treatment of full-thickness skin loss using ovine tendon collagen type I scaffold with skin cells. J. Tissue Eng. Regen. Med. 2019, 13, 874–891. [Google Scholar] [CrossRef]
- Salleh, A.; Mustafa, N.; Teow, Y.H.; Fatimah, M.N.; Khairudin, F.A.; Ahmad, I.; Fauzi, M.B. Dual-Layered Approach of Ovine Collagen-Gelatin/Cellulose Hybrid Biomatrix Containing Graphene Oxide-Silver Nanoparticles for Cutaneous Wound Healing: Fabrication, Physicochemical, Cytotoxicity and Antibacterial Characterisation. Biomedicines 2022, 10, 816. [Google Scholar] [CrossRef]
- Xu, R.; Xia, H.; He, W.; Li, Z.; Zhao, J.; Liu, B.; Wang, Y.; Lei, Q.; Kong, Y.; Bai, Y.; et al. Controlled water vapor transmission rate promotes wound-healing via wound re-epithelialization and contraction enhancement. Sci. Rep. 2016, 6, 24596. [Google Scholar] [CrossRef]
- Chen, J.; Li, J.; Li, Z.; Yi, R.; Shi, S.; Wu, K.; Li, Y.; Wu, S. Physicochemical and functional properties of type I collagens in Red Stingray (Dasyatis akajei) Skin. Mar. Drugs 2019, 17, 558. [Google Scholar] [CrossRef]
- Baer, D.R.; Artyushkova, K.; Richard Brundle, C.; Castle, J.E.; Engelhard, M.H.; Gaskell, K.J.; Grant, J.T.; Haasch, R.T.; Linford, M.R.; Powell, C.J.; et al. Practical guides for x-ray photoelectron spectroscopy: First steps in planning, conducting, and reporting XPS measurements. J. Vac. Sci. Technol. A 2019, 37, 03140111. [Google Scholar] [CrossRef]
- Sasmal, P.; Begam, H. Extraction of type-I collagen from sea fish and synthesis of Hap/collagen composite. Procedia Mater. Sci. 2014, 5, 1136–1140. [Google Scholar] [CrossRef]
- Fauzi, M.; Chowdhury, S.R.; Idrus, R.; Aminuddin, B. Fabrication of collagen type I scaffold for skin tissue engineering. Regen. Res. 2014, 3, 60–61. [Google Scholar]
- Zhang, F.; Wang, A.; Li, Z.; He, S.; Shao, L. Preparation and Characterisation of Collagen from Freshwater Fish Scales. Food Nutr. Sci. 2011, 02, 818–823. [Google Scholar] [CrossRef]
- León-Mancilla, B.H.; Araiza-Téllez, M.A.; Flores-Flores, J.O.; Piña-Barba, M.C. Physico-chemical characterization of collagen scaffolds for tissue engineering. J. Appl. Res. Technol. 2016, 14, 77–85. [Google Scholar] [CrossRef]
- Yang, Y.L.; Motte, S.; Kaufman, L.J. Pore size variable type I collagen gels and their interaction with glioma cells. Biomaterials 2010, 31, 5678–5688. [Google Scholar] [CrossRef] [PubMed]
- Rabotyagova, O.S.; Cebe, P.; Kaplan, D.L. Collagen Structural Hierarchy and Susceptibility to Degradation by Ultraviolet Radiation. Mater. Sci. Eng. C. Mater. Biol. Appl. 2008, 28, 1420. [Google Scholar] [CrossRef]
- Ratner, B.D. The biocompatibility manifesto: Biocompatibility for the twenty-first century. J. Cardiovasc. Transl. Res. 2011, 4, 523–527. [Google Scholar] [CrossRef]
- Lotfi, M.; Nejib, M.; Naceur, M. Cell Adhesion to Biomaterials: Concept of Biocompatibility. In Advances in Biomaterials Science and Biomedical Applications; InTech: London, UK, 2013. [Google Scholar]
- Yu, Q.; Zhao, L.; Guo, C.; Yan, B.; Su, G. Regulating Protein Corona Formation and Dynamic Protein Exchange by Controlling Nanoparticle Hydrophobicity. Front. Bioeng. Biotechnol. 2020, 8, 210. [Google Scholar] [CrossRef]
- Bai, X.; Wang, J.; Mu, Q.; Su, G. In vivo Protein Corona Formation: Characterizations, Effects on Engineered Nanoparticles’ Biobehaviors, and Applications. Front. Bioeng. Biotechnol. 2021, 9, 646708. [Google Scholar] [CrossRef]
- Hirsh, S.L.; McKenzie, D.R.; Nosworthy, N.J.; Denman, J.A.; Sezerman, O.U.; Bilek, M.M.M. The Vroman effect: Competitive protein exchange with dynamic multilayer protein aggregates. Colloids Surfaces B Biointerfaces 2013, 103, 395–404. [Google Scholar] [CrossRef]
- Hu, W.J.; Eaton, J.W.; Tang, L. Molecular basis of biomaterial-mediated foreign body reactions. Blood 2001, 98, 1231–1238. [Google Scholar] [CrossRef] [PubMed]
- Serpooshan, V.; Mahmoudi, M.; Zhao, M.; Wei, K.; Sivanesan, S.; Motamedchaboki, K.; Malkovskiy, A.V.; Goldstone, A.B.; Cohen, J.E.; Yang, P.C.; et al. Protein Corona Influences Cell-Biomaterial Interactions in Nanostructured Tissue Engineering Scaffolds. Adv. Funct. Mater. 2015, 25, 4379–4389. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.M.; Rodriguez, A.; Chang, D.T. Foreign body reaction to biomaterials. Semin. Immunol. 2008, 20, 86. [Google Scholar] [CrossRef] [PubMed]
- Li, J.J.; Zreiqat, H. Tissue Response to Biomaterials; Elsevier Inc.: Amsterdam, The Netherlands, 2019; Volume 1–3, ISBN 9780128051443. [Google Scholar]
- Noskovicova, N.; Hinz, B.; Pakshir, P. Implant Fibrosis and the Underappreciated Role of Myofibroblasts in the Foreign Body Reaction. Cells 2021, 10, 1794. [Google Scholar] [CrossRef]
- Wadhwa, M.; Knezevic, I.; Kang, H.-N.; Thorpe, R. Immunogenicity assessment of biotherapeutic products: An overview of assays and their utility. Biologicals 2015, 43, 298–306. [Google Scholar] [CrossRef]
- Velasco, M.A.; Narváez-Tovar, C.A.; Garzón-Alvarado, D.A. Design, Materials, and Mechanobiology of Biodegradable Scaffolds for Bone Tissue Engineering. Biomed Res. Int. 2015, 2015, 729076. [Google Scholar] [CrossRef]
- Frolova, A.; Aksenova, N.; Novikov, I.; Maslakova, A.; Gafarova, E.; Efremov, Y.; Bikmulina, P.; Elagin, V.; Istranova, E.; Kurkov, A.; et al. A collagen basketweave from the giant squid mantle as a robust scaffold for tissue engineering. Mar. Drugs 2021, 19, 679. [Google Scholar] [CrossRef]
- El-Sherbiny, I.M.; Yacoub, M.H. Hydrogel scaffolds for tissue engineering: Progress and challenges. Glob. Cardiol. Sci. Pract. 2013, 3, 316–342. [Google Scholar] [CrossRef]
- Zhu, J.; Marchant, R.E. Design properties of hydrogel tissue-engineering scaffolds. Expert Rev. Med. Devices 2011, 8, 607–626. [Google Scholar] [CrossRef]
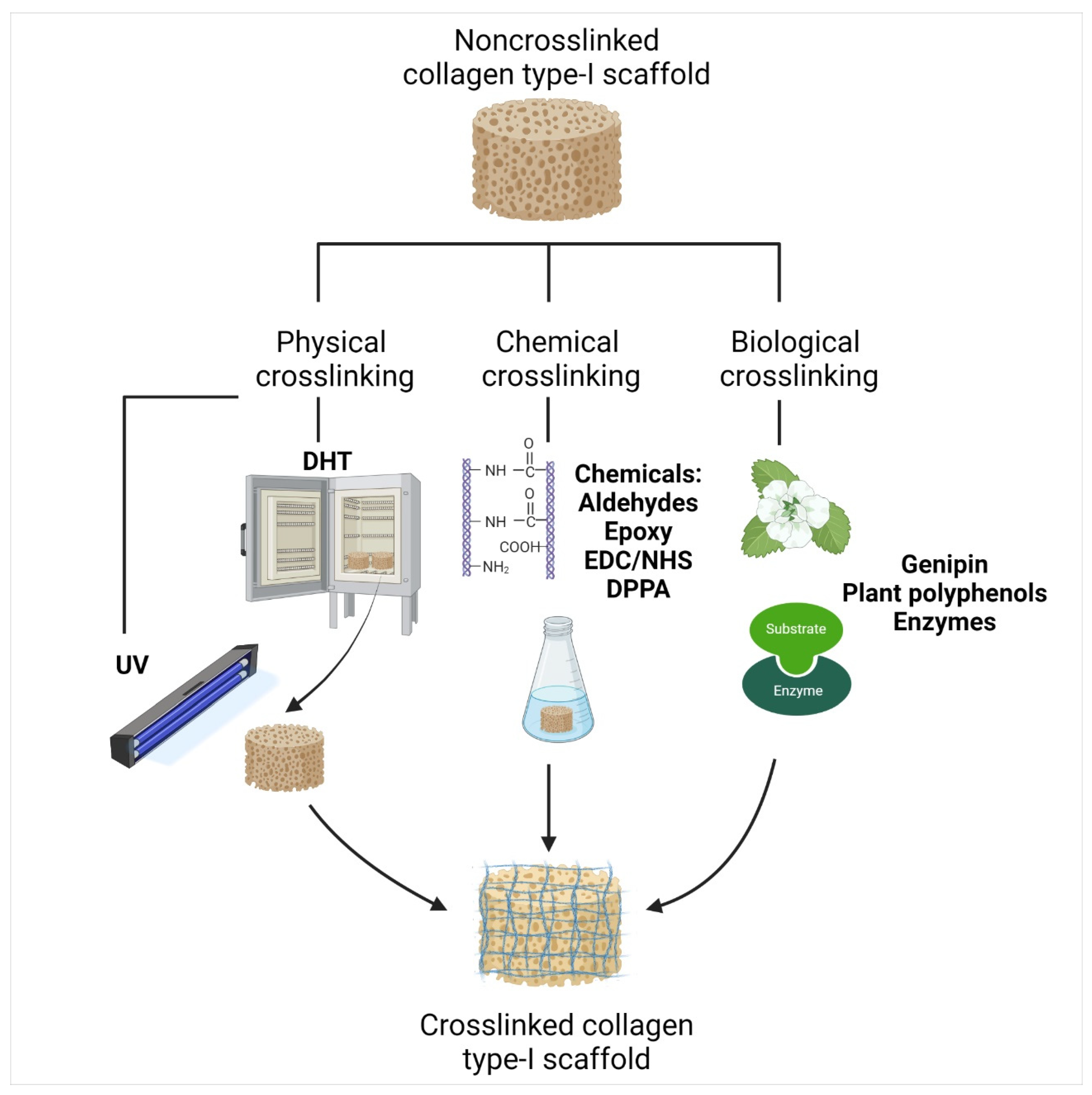
- Jiang, Y.-H.; Lou, Y.-Y.; Li, T.-H.; Liu, B.-Z.; Chen, K.; Zhang, D.; Li, T. Cross-linking methods of type I collagen-based scaffolds for cartilage tissue engineering. Am. J. Transl. Res. 2022, 14, 1146. [Google Scholar]
- Adamiak, K.; Sionkowska, A. Current methods of collagen cross-linking: Review. Int. J. Biol. Macromol. 2020, 161, 550–560. [Google Scholar] [CrossRef] [PubMed]
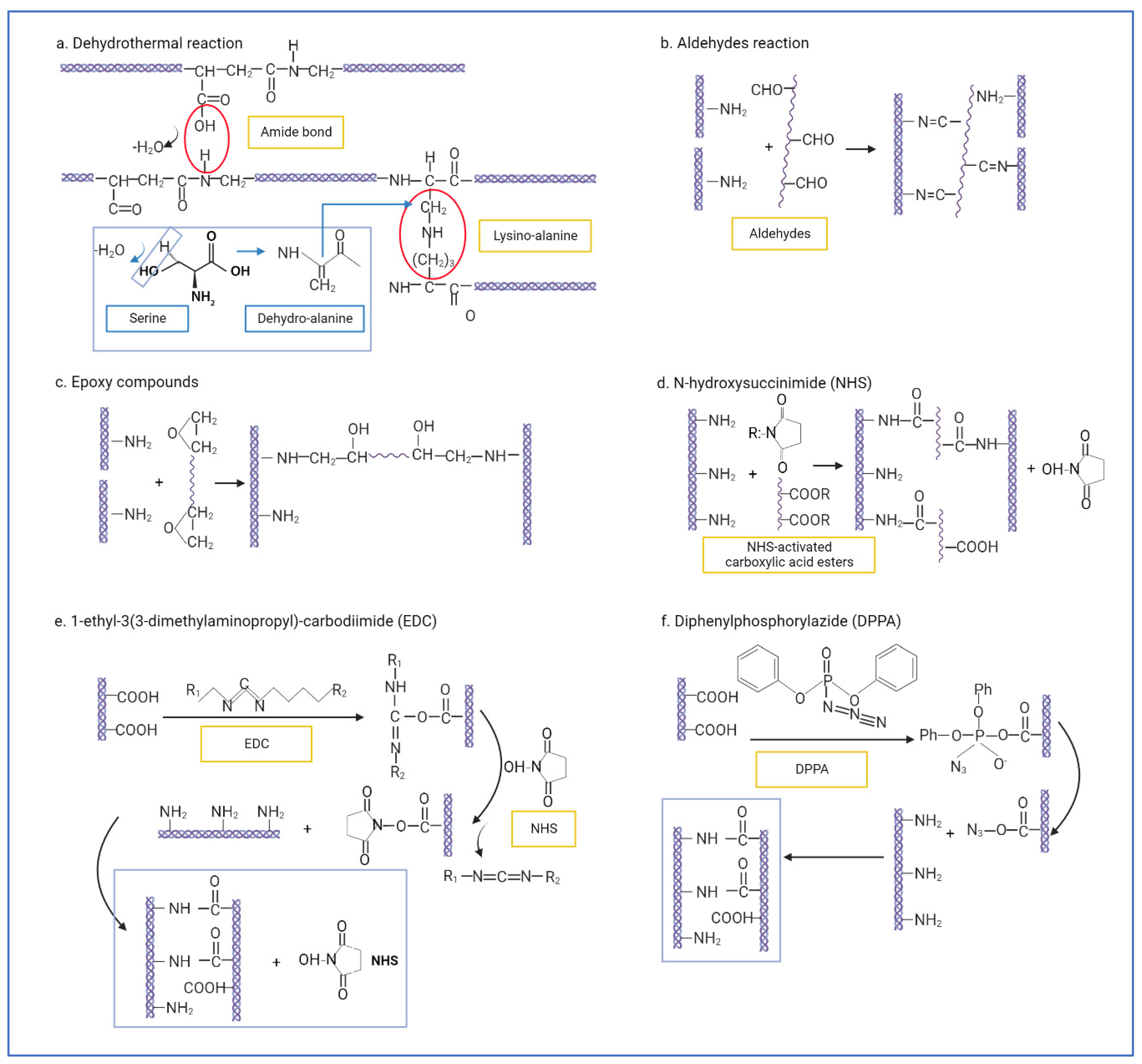
- Gorham, S.D.; Light, N.D.; Diamond, A.M.; Willins, M.J.; Bailey, A.J.; Wess, T.J.; Leslie, N.J. Effect of chemical modifications on the susceptibility of collagen to proteolysis. II. Dehydrothermal crosslinking. Int. J. Biol. Macromol. 1992, 14, 129–138. [Google Scholar] [CrossRef]
- Chen, X.; Zhou, L.; Xu, H.; Yamamoto, M.; Shinoda, M.; Kishimoto, M.; Tanaka, T.; Yamane, H. Effect of the Application of a Dehydrothermal Treatment on the Structure and the Mechanical Properties of Collagen Film. Materials 2020, 13, 377. [Google Scholar] [CrossRef] [PubMed]
- Sionkowska, A. Thermal denaturation of UV-irradiated wet rat tail tendon collagen. Int. J. Biol. Macromol. 2005, 35, 145–149. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, M.; Liu, W.; Li, G. Properties of alkali-solubilized collagen solution crosslinked by N-hydroxysuccinimide activated adipic acid. KARJ 2011, 23, 41–48. [Google Scholar] [CrossRef]
- Petite, H.; Frei, V.; Huc, A.; Herbage, D. Use of diphenylphosphorylazide for cross-linking collagen-based biomaterials. J. Biomed. Mater. Res. 1994, 28, 159–165. [Google Scholar] [CrossRef]
- Roche, S.; Ronzière, M.C.; Herbage, D.; Freyria, A.M. Native and DPPA cross-linked collagen sponges seeded with fetal bovine epiphyseal chondrocytes used for cartilage tissue engineering. Biomaterials 2000, 22, 9–18. [Google Scholar] [CrossRef]
- Sarikaya, B.; Aydin, H.M. Collagen/Beta-Tricalcium Phosphate Based Synthetic Bone Grafts via Dehydrothermal Processing. Biomed Res. Int. 2015, 2015, 576532. [Google Scholar] [CrossRef]
- Fagerholm, P.; Lagali, N.S.; Merrett, K.; Jackson, W.B.; Munger, R.; Liu, Y.; Polarek, J.W.; Söderqvist, M.; Griffith, M. A biosynthetic alternative to human donor tissue for inducing corneal regeneration: 24-month follow-up of a phase 1 clinical study. Sci. Transl. Med. 2010, 2, 46ra61. [Google Scholar] [CrossRef]
- Koh, L.B.; Islam, M.M.; Mitra, D.; Noel, C.W.; Merrett, K.; Odorcic, S.; Fagerholm, P.; Jackson, W.B.; Liedberg, B.; Phopase, J.; et al. Epoxy Cross-Linked Collagen and Collagen-Laminin Peptide Hydrogels as Corneal Substitutes. J. Funct. Biomater. 2013, 4, 162. [Google Scholar] [CrossRef]
- Kozłowska, J.; Sionkowska, A. Effects of different crosslinking methods on the properties of collagen-calcium phosphate composite materials. Int. J. Biol. Macromol. 2015, 74, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Chu, C.; Rung, S.; Yang, R.; Man, Y.; Qu, Y. Application of EGCG Modified EDC/NHS Cross-Linked Extracellular Matrix to Promote Macrophage Adhesion. bioRxiv 2020. [Google Scholar] [CrossRef]
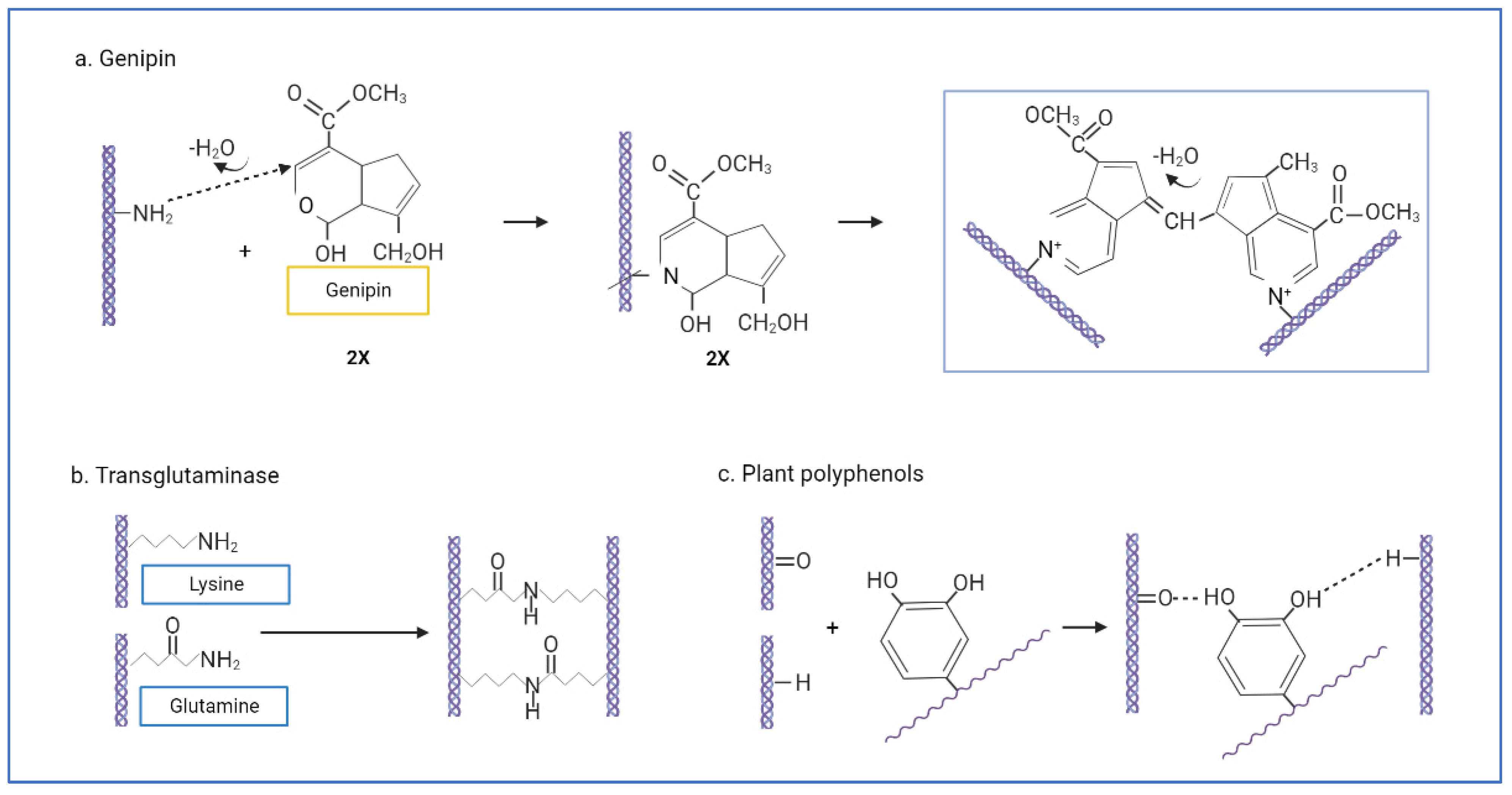
- Yoo, J.S.; Kim, Y.J.; Kim, S.H.; Choi, S.H. Study on Genipin: A New Alternative Natural Crosslinking Agent for Fixing Heterograft Tissue. Korean J. Thorac. Cardiovasc. Surg. 2011, 44, 197. [Google Scholar] [CrossRef] [PubMed]
- Bi, L.; Cao, Z.; Hu, Y.; Song, Y.; Yu, L.; Yang, B.; Mu, J.; Huang, Z.; Han, Y. Effects of different cross-linking conditions on the properties of genipin-cross-linked chitosan/collagen scaffolds for cartilage tissue engineering. J. Mater. Sci. Mater. Med. 2011, 22, 51–62. [Google Scholar] [CrossRef]
- Sung, H.W.; Huang, R.N.; Huang, L.L.H.; Tsai, C.C.; Chiu, C.T. Feasibility study of a natural crosslinking reagent for biological tissue fixation. J. Biomed. Mater. Res. 1998, 42, 560–567. [Google Scholar] [CrossRef]
- Griffin, M.; Casadio, R.; Bergamini, C.M. Transglutaminases: Nature’s biological glues. Biochem. J. 2002, 368, 377–396. [Google Scholar] [CrossRef]
- Nonaka, M.; Tanaka, H.; Okiyama, A.; Motoki, M.; Ando, H.; Umeda, K.; Matsuura, A. Polymerization of Several Proteins by Ca2+-Independent Transglutaminase Derived from Microorganisms. Agric. Biol. Chem. 1989, 53, 2619–2623. [Google Scholar] [CrossRef]
- Fang, C.-H.; Sun, C.-K.; Lin, Y.-W.; Hung, M.-C.; Lin, H.-Y.; Li, C.-H.; Lin, I.-P.; Chang, H.-C.; Sun, J.-S.; Chang, J.Z.-C. Metformin-Incorporated Gelatin/Nano-Hydroxyapatite Scaffolds Promotes Bone Regeneration in Critical Size Rat Alveolar Bone Defect Model. Int. J. Mol. Sci. 2022, 23, 558. [Google Scholar] [CrossRef]
- Kilic Bektas, C.; Kimiz, I.; Sendemir, A.; Hasirci, V.; Hasirci, N. A bilayer scaffold prepared from collagen and carboxymethyl cellulose for skin tissue engineering applications. J. Biomater. Sci. Polym. Ed. 2018, 29, 1764–1784. [Google Scholar] [CrossRef]
- Han, B.; Jaurequi, J.; Tang, B.W.; Nimni, M.E. Proanthocyanidin: A natural crosslinking reagent for stabilizing collagen matrices. J. Biomed. Mater. Res. A 2003, 65, 118–124. [Google Scholar] [CrossRef]
- Yu, H.L.; Feng, Z.Q.; Zhang, J.J.; Wang, Y.H.; Ding, D.J.; Gao, Y.Y.; Zhang, W.F. The Evaluation of Proanthocyanidins/Chitosan/Lecithin Microspheres as Sustained Drug Delivery System. Biomed Res. Int. 2018, 2018, 9073420. [Google Scholar] [CrossRef] [PubMed]
- Metreveli, N.O.; Namicheishvili, L.O.; Dzhariashvili, K.K.; Chikvaidze, E.N.; Mrevlishvili, G.M. A microcalorimetric and electron spin resonance study of the influence of UV radiation on collagen. Biophysics. 2006, 51, 29–32. [Google Scholar] [CrossRef]
- Hawkins, C.L.; Davies, M.J. Generation and propagation of radical reactions on proteins. Biochim. Biophys. Acta-Bioenerg. 2001, 1504, 196–219. [Google Scholar] [CrossRef]
- Veelaert, S. Properties and applications of dialdehyde starch. Żywność Technol. Jakość 1996, 2, 75–83. [Google Scholar]
- Zeeman, R.; Dijkstra, P.J.; Van Wachem, P.B.; Van Luyn, M.J.A.; Hendriks, M.; Cahalan, P.T.; Feijen, J. Crosslinking and modification of dermal sheep collagen using 1,4- butanediol diglycidyl ether. J. Biomed. Mater. Res. 1999, 46, 424–433. [Google Scholar] [CrossRef]
- Sung, H.W.; Hsu, H.L.; Shih, C.C.; Lin, D.S. Cross-linking characteristics of biological tissues fixed with monofunctional or multifunctional epoxy compounds. Biomaterials 1996, 17, 1405–1410. [Google Scholar] [CrossRef]
- Olde Damink, L.H.H.; Dijkstra, P.J.; Van Luyn, M.J.A.; Van Wachem, P.B.; Nieuwenhuis, P.; Feijen, J. Cross-linking of dermal sheep collagen using a water-soluble carbodiimide. Biomaterials 1996, 17, 765–773. [Google Scholar] [CrossRef]
- Grabarek, Z.; Gergely, J. Zero-length crosslinking procedure with the use of active esters. Anal. Biochem. 1990, 185, 131–135. [Google Scholar] [CrossRef]
- Gauza-Włodarczyk, M.; Kubisz, L.; Mielcarek, S.; Włodarczyk, D. Comparison of thermal properties of fish collagen and bovine collagen in the temperature range 298–670 K. Mater. Sci. Eng. C 2017, 80, 468–471. [Google Scholar] [CrossRef]
- Fujii, K.K.; Taga, Y.; Takagi, Y.K.; Masuda, R.; Hattori, S.; Koide, T. The Thermal Stability of the Collagen Triple Helix Is Tuned According to the Environmental Temperature. Int. J. Mol. Sci. 2022, 23, 2040. [Google Scholar] [CrossRef]
- Nagai, T. Collagen from diamondback squid (Thysanoteuthis rhombus) outer skin. Zeitschrift fur Naturforsch.-Sect. C J. Biosci. 2004, 59, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Ikoma, T.; Kobayashi, H.; Tanaka, J.; Walsh, D.; Mann, S. Physical properties of type I collagen extracted from fish scales of Pagrus major and Oreochromis niloticas. Int. J. Biol. Macromol. 2003, 32, 199–204. [Google Scholar] [CrossRef]
- Gaill, F.; Mann, K.; Wiedemann, H.; Engel, J.; Timpl, R. Structural comparison of cuticle and interstitial collagens from annelids living in shallow sea-water and at deep-sea hydrothermal vents. J. Mol. Biol. 1995, 246, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Leikina, E.; Mertts, M.V.; Kuznetsova, N.; Leikin, S. Type I collagen is thermally unstable at body temperature. Proc. Natl. Acad. Sci. USA 2002, 99, 1314–1318. [Google Scholar] [CrossRef]
- Gevorkian, S.G.; Allahverdyan, A.E.; Gevorgyan, D.S.; Simonian, A.L.; Hu, C.K. Stabilization and anomalous hydration of collagen fibril under heating. PLoS ONE 2013, 8, e78526. [Google Scholar] [CrossRef]
- Yamamoto, K.; Yoshizawa, Y.; Yanagiguchi, K.; Ikeda, T.; Yamada, S.; Hayashi, Y. The Characterization of Fish (Tilapia) Collagen Sponge as a Biomaterial. Int. J. Polym. Sci. 2015, 2015, 957385. [Google Scholar] [CrossRef]
- Pati, F.; Adhikari, B.; Dhara, S. Isolation and characterization of fish scale collagen of higher thermal stability. Bioresour. Technol. 2010, 101, 3737–3742. [Google Scholar] [CrossRef]
- Safandowska, M.; Pietrucha, K. Effect of fish collagen modification on its thermal and rheological properties. Int. J. Biol. Macromol. 2013, 53, 32–37. [Google Scholar] [CrossRef]
- Sallehuddin, N.; Md Fadilah, N.I.; Hwei, N.M.; Wen, A.P.Y.; Yusop, S.M.; Rajab, N.F.; Hiraoka, Y.; Tabata, Y.; Fauzi, M.B. Characterization and Cytocompatibility of Collagen–Gelatin–Elastin (CollaGee) Acellular Skin Substitute towards Human Dermal Fibroblasts: In Vitro Assessment. Biomedicines 2022, 10, 1327. [Google Scholar] [CrossRef]
- Loh, Q.L.; Choong, C. Three-dimensional scaffolds for tissue engineering applications: Role of porosity and pore size. Tissue Eng.-Part B Rev. 2013, 19, 485–502. [Google Scholar] [CrossRef]
- Polo-Corrales, L.; Latorre-Esteves, M.; Ramirez-Vick, J.E. Scaffold design for bone regeneration. J. Nanosci. Nanotechnol. 2014, 14, 15–56. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lu, H.; Kawazoe, N.; Chen, G. Pore size effect of collagen scaffolds on cartilage regeneration. Acta Biomater. 2014, 10, 2005–2013. [Google Scholar] [CrossRef] [PubMed]
- Sionkowska, A.; Kozłowska, J. Properties and modification of porous 3-D collagen/hydroxyapatite composites. Int. J. Biol. Macromol. 2013, 52, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lu, H.; Kawazoe, N.; Chen, G. Preparation of collagen scaffolds with controlled pore structures and improved mechanical property for cartilage tissue engineering. J. Bioact. Compat. Polym. 2013, 28, 426–438. [Google Scholar] [CrossRef]
- Xu, C.; Thiruvadi, V.S.; Whitmore, R.; Liu, H. Delivery systems for biomedical applications: Basic introduction, research frontiers and clinical translations. In Biomaterials in Translational Medicine; Chapter 5; Academic Press: Cambridge, MA, USA, 2019; pp. 93–116. [Google Scholar] [CrossRef]
- Annabi, N.; Nichol, J.W.; Zhong, X.; Ji, C.; Koshy, S.; Khademhosseini, A.; Dehghani, F. Controlling the porosity and microarchitecture of hydrogels for tissue engineering. Tissue Eng.-Part B Rev. 2010, 16, 371–383. [Google Scholar] [CrossRef]
- Jurczyk, M.U.; Żurawski, J.; Wirstlein, P.K.; Kowalski, K.; Jurczyk, M. Response of inflammatory cells to biodegradable ultra-fine grained Mg-based composites. Micron 2020, 129, 102796. [Google Scholar] [CrossRef]
- Rothamel, D.; Benner, M.; Fienitz, T.; Happe, A.; Kreppel, M.; Nickenig, H.J.; Zöller, J.E. Biodegradation pattern and tissue integration of native and cross-linked porcine collagen soft tissue augmentation matrices—An experimental study in the rat. Head Face Med. 2014, 10, 10. [Google Scholar] [CrossRef]
- Chattopadhyay, S.; Raines, R.T. Collagen-based biomaterials for wound healing. Biopolymers 2014, 101, 821–833. [Google Scholar] [CrossRef]
- Valenzuela-Rojo, D.R.; López-Cervantes, J.; Sánchez-Machado, D.I. Tilapia (Oreochromis aureus) Collagen for Medical Biomaterials. In Seaweed Biomaterials; Chapter 4; Maiti, S., Ed.; IntechOpen: London, UK, 2018; ISBN 978-1-78984-847-2. [Google Scholar] [CrossRef]
- Song, X.; Zhu, C.; Fan, D.; Mi, Y.; Li, X.; Fu, R.Z.; Duan, Z.; Wang, Y.; Feng, R.R. A novel human-like collagen hydrogel scaffold with porous structure and sponge-like properties. Polymers 2017, 9, 638. [Google Scholar] [CrossRef]
- Takallu, S.; Mirzaei, E.; Azadi, A.; Karimizade, A.; Tavakol, S. Plate-shape carbonated hydroxyapatite/collagen nanocomposite hydrogel via in situ mineralization of hydroxyapatite concurrent with gelation of collagen at pH = 7.4 and 37 °C. J. Biomed. Mater. Res.-Part B Appl. Biomater. 2018, 107, 1920–1929. [Google Scholar] [CrossRef]
- Ghica, M.V.; Kaya, M.G.A.; Dinu-Pîrvu, C.E.; Lupuleasa, D.; Udeanu, D.I. Development, optimization and in vitro/in vivo characterization of collagen-dextran spongious wound dressings loaded with flufenamic acid. Molecules 2017, 22, 1552–1574. [Google Scholar] [CrossRef]
- Tronci, G.; Grant, C.A.; Thomson, N.H.; Russell, S.J.; Wood, D.J. Multi-scale mechanical characterization of highly swollen photo-activated collagen hydrogels. J. R. Soc. Interface 2015, 12, 1–13. [Google Scholar] [CrossRef]
- Shi, Y.; Zhang, H.; Zhang, X.; Chen, Z.; Zhao, D.; Ma, J. A comparative study of two porous sponge scaffolds prepared by collagen derived from porcine skin and fish scales as burn wound dressings in a rabbit model. Regen. Biomater. 2020, 7, 63–70. [Google Scholar] [CrossRef]
- Amirrah, I.N.; Razip Wee, M.F.M.; Tabata, Y.; Idrus, R.B.H.; Nordin, A.; Fauzi, M.B. Antibacterial-integrated collagen wound dressing for diabetes-related foot ulcers: An evidence-based review of clinical studies. Polymers 2020, 12, 2168. [Google Scholar] [CrossRef]
- Hussain, Z.; Thu, H.E.; Shuid, A.N.; Katas, H.; Hussain, F. Recent Advances in Polymer-based Wound Dressings for the Treatment of Diabetic Foot Ulcer: An Overview of State-of-the-art. Curr. Drug Targets 2017, 19, 527–550. [Google Scholar] [CrossRef]
- Gentleman, M.M.; Gentleman, E. The role of surface free energy in osteoblast-biomaterial interactions. Int. Mater. Rev. 2014, 59, 417–429. [Google Scholar] [CrossRef]
- Chang, H.-I.; Wang, Y. Cell Responses to Surface and Architecture of Tissue Engineering Scaffolds. In Regenerative Medicine and Tissue Engineering-Cells and Biomaterials; Eberli, D., Ed.; IntechOpen: Taiwan, China, 2011; pp. 569–588. ISBN 978-953-307-663-8. [Google Scholar]
- Marín-Pareja, N.; Cantini, M.; González-García, C.; Salvagni, E.; Salmerón-Sánchez, M.; Ginebra, M.-P. Different Organization of Type I Collagen Immobilized on Silanized and Nonsilanized Titanium Surfaces Affects Fibroblast Adhesion and Fibronectin Secretion. ACS Appl. Mater. Interfaces 2015, 7, 20667–20677. [Google Scholar] [CrossRef]
- Adamczak, M.I.; Genet, M.J.; Scislowska-Czarnecka, A.; Dupont, C.; Pamula, E. Surface characterization, collagen adsorption and cell behaviour on poly(L-lactide-co-glycolide). Acta Bioeng. Biomech. 2011, 13, 63–75. [Google Scholar]
- Popa, L.; Ghica, M.V.; Albu, M.G.; Ortan, A.; Dinu-Pîrvu, C.-E. Hysteresis of contact angle. Dynamic wettability studies of collagen and doxycycline porous matrices crosslinked with tannic acid. Dig. J. Nanomater. Biostructures 2013, 8, 937–943. [Google Scholar]
- Mateos-Timoneda, M.A.; Castano, O.; Planell, J.A.; Engel, E. Effect of structure, topography and chemistry on fibroblast adhesion and morphology. J. Mater. Sci. Mater. Med. 2014, 25, 1781–1787. [Google Scholar] [CrossRef]
- Zhou, N.; Ma, X.; Hu, W.; Ren, P.; Zhao, Y.; Zhang, T. Effect of RGD content in poly(ethylene glycol)-crosslinked poly(methyl vinyl ether-alt-maleic acid) hydrogels on the expansion of ovarian cancer stem-like cells. Mater. Sci. Eng. C. Mater. Biol. Appl. 2021, 118, 111477. [Google Scholar] [CrossRef]
- Huang, C.-C.; Kang, M.; Shirazi, S.; Lu, Y.; Cooper, L.F.; Gajendrareddy, P.; Ravindran, S. 3D Encapsulation and tethering of functionally engineered extracellular vesicles to hydrogels. Acta Biomater. 2021, 126, 199–210. [Google Scholar] [CrossRef]
- Le Saux, G.; Magenau, A.; Böcking, T.; Gaus, K.; Gooding, J.J. The Relative Importance of Topography and RGD Ligand Density for Endothelial Cell Adhesion. PLoS ONE 2011, 6, e21869. [Google Scholar] [CrossRef]
- Liu, C.; Shen, S.Z.; Han, Z. Surface wettability and chemistry of ozone perfusion processed porous collagen scaffold. J. Bionic Eng. 2011, 8, 223–233. [Google Scholar] [CrossRef]
- Scarano, A.; Lorusso, F.; Orsini, T.; Morra, M.; Iviglia, G.; Valbonetti, L. Biomimetic surfaces coated with covalently immobilized collagen type I: An x-ray photoelectron spectroscopy, atomic force microscopy, micro-CT and histomorphometrical study in rabbits. Int. J. Mol. Sci. 2019, 20, 724. [Google Scholar] [CrossRef]
- Rumian, L.; Wojak, I.; Scharnweber, D.; Pamuła, E. Resorbable scaffolds modified with collagen type I or hydroxyapatite: In vitro studies on human mesenchymal stem cells. Acta Bioeng. Biomech. 2013, 15, 61–67. [Google Scholar] [CrossRef]
- Sanjurjo-Rodríguez, C.; Castro-Viñuelas, R.; Hermida-Gómez, T.; Fernández-Vázquez, T.; Fuentes-Boquete, I.M.; de Toro-Santos, F.J.; Díaz-Prado, S.M.; Blanco-García, F.J. Ovine mesenchymal stromal cells: Morphologic, phenotypic and functional characterization for osteochondral tissue engineering. PLoS ONE 2017, 12, e0171231. [Google Scholar] [CrossRef]
- Strauss, K.; Chmielewski, J. Advances in the design and higher-order assembly of collagen mimetic peptides for regenerative medicine. Curr. Opin. Biotechnol. 2017, 46, 34–41. [Google Scholar] [CrossRef]
- Driver, V.R.; Lavery, L.A.; Reyzelman, A.M.; Dutra, T.G.; Dove, C.R.; Kotsis, S.V.; Kim, H.M.; Chung, K.C. A clinical trial of Integra Template for diabetic foot ulcer treatment. Wound Repair Regen. 2015, 23, 891–900. [Google Scholar] [CrossRef]
- Aubert, L.; Dubus, M.; Rammal, H.; Bour, C.; Mongaret, C.; Boulagnon-Rombi, C.; Garnotel, R.; Schneider, C.; Rahouadj, R.; Laurent, C.; et al. Collagen-Based Medical Device as a Stem Cell Carrier for Regenerative Medicine. Int. J. Mol. Sci. 2017, 18, 2210. [Google Scholar] [CrossRef]
- Hefka Blahnová, V.; Vojtová, L.; Pavliňáková, V.; Muchová, J.; Filová, E. Calcined Hydroxyapatite with Collagen I Foam Promotes Human MSC Osteogenic Differentiation. Int. J. Mol. Sci. 2022, 23, 4236. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Yang, R.; Chu, C.; Hu, C.; Zou, Q.; Li, Y.; Zuo, Y.; Man, Y.; Li, J. Topological structure of electrospun membrane regulates immune response, angiogenesis and bone regeneration. Acta Biomater. 2021, 129, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Xu, Y.; Xia, J.; Zhou, Z.; Fang, Y.; Zhang, L.; Sun, W. Multi-scale hierarchical scaffolds with aligned micro-fibers for promoting cell alignment. Biomed. Mater. 2021, 16, 045047. [Google Scholar] [CrossRef]
- Chun, Y.H.; Park, S.-K.; Kim, E.J.; Lee, H.J.; Kim, H.; Koh, W.-G.; Cunha, G.F.; Myung, D.; Na, K.-S. In vivo biocompatibility evaluation of in situ-forming polyethylene glycol-collagen hydrogels in corneal defects. Sci. Rep. 2021, 11, 23913. [Google Scholar] [CrossRef] [PubMed]
- Djavid, G.E.; Tabaie, S.M.; Tajali, S.B.; Totounchi, M.; Farhoud, A.; Fateh, M.; Ghafghazi, M.; Koosha, M.; Taghizadeh, S. Application of a collagen matrix dressing on a neuropathic diabetic foot ulcer: A randomised control trial. J. Wound Care 2020, 29, S13–S18. [Google Scholar] [CrossRef]
- Anselmo, A.C.; Mitragotri, S. Nanoparticles in the clinic. Bioeng. Transl. Med. 2016, 1, 10–29. [Google Scholar] [CrossRef]
- Naghizadeh, Z.; Karkhaneh, A.; Nokhbatolfoghahaei, H.; Farzad-Mohajeri, S.; Rezai-Rad, M.; Dehghan, M.M.; Aminishakib, P.; Khojasteh, A. Cartilage regeneration with dual-drug-releasing injectable hydrogel/microparticle system: In vitro and in vivo study. J. Cell. Physiol. 2021, 236, 2194–2204. [Google Scholar] [CrossRef]
- Coentro, J.Q.; di Nubila, A.; May, U.; Prince, S.; Zwaagstra, J.; Järvinen, T.A.H.; Zeugolis, D.I. Dual drug delivery collagen vehicles for modulation of skin fibrosisin vitro. Biomed. Mater. 2022, 17, 025017. [Google Scholar] [CrossRef]
- Chen, Q.C.; Wu, J.; Liu, Y.; Li, Y.Q.; Zhang, C.Q.; Qi, W.C.; Yeung, K.W.K.; Wong, T.M.; Zhao, X.L.; Pan, H.B. Electrospun chitosan/PVA/bioglass Nanofibrous membrane with spatially designed structure for accelerating chronic wound healing. Mater. Sci. Eng. C-Materials Biol. Appl. 2019, 105, 110083. [Google Scholar] [CrossRef]
- Dhinasekaran, D.; Vimalraj, S.; Rajendran, A.R.; Saravanan, S.; Purushothaman, B.; Subramaniam, B. Bio-inspired multifunctional collagen/electrospun bioactive glass membranes for bone tissue engineering applications. Mater. Sci. Eng. C 2021, 126, 111856. [Google Scholar] [CrossRef]
- Pinho, L.C.; Alves, M.M.; Colaço, B.; Fernandes, M.H.; Santos, C. Green-Synthesized Magnesium Hydroxide Nanoparticles Induced Osteoblastic Differentiation in Bone Co-Cultured Cells. Pharmaceuticals 2021, 14, 1281. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Kong, B.; Xie, H.; Zhang, J.; Liu, W.; Liu, S.; Zhang, Y.; Yang, F.; Xiao, J.; Mi, S.; et al. PCL scaffold combined with rat tail collagen type I to reduce keratocyte differentiation and prevent corneal stroma fibrosis after injury. Exp. Eye Res. 2022, 217, 108936. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yaron, J.R.; Guo, Q.; Kilbourne, J.; Awo, E.A.; Burgin, M.; Schutz, L.N.; Wallace, S.E.; Lowe, K.M.; Lucas, A.R. Topical Application of Virus-Derived Immunomodulating Proteins and Peptides to Promote Wound Healing in Mouse Models. Methods Mol. Biol. 2021, 2225, 217–226. [Google Scholar] [CrossRef]
- Marín-Payá, J.C.; Díaz-Benito, B.; Martins, L.A.; Trujillo, S.C.; Cordón, L.; Lanceros-Méndez, S.; Gallego Ferrer, G.; Sempere, A.; Gómez Ribelles, J.L. Biomimetic 3D Environment Based on Microgels as a Model for the Generation of Drug Resistance in Multiple Myeloma. Materials 2021, 14, 7121. [Google Scholar] [CrossRef] [PubMed]
- Motta, A.; Barone, R.; Macaluso, F.; Giambalvo, F.; Pecoraro, F.; Di Marco, P.; Cassata, G.; Puleio, R.; Migliaresi, C.; Guercio, A.; et al. Silk-based matrices and c-Kit positive cardiac progenitor cells for a cellularized silk fibroin scaffold: Study of an in vivo model. Cells. Tissues. Organs 2022, 1, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Oyama, T.G.; Oyama, K.; Kimura, A.; Yoshida, F.; Ishida, R.; Yamazaki, M.; Miyoshi, H.; Taguchi, M. Collagen hydrogels with controllable combined cues of elasticity and topography to regulate cellular processes. Biomed. Mater. 2021, 16, 045037. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.M.N.; Moreira, A.P.D.; Carvalho, C.W.P.; Luchese, R.; Ribeiro, E.; McGuinness, G.B.; Mendes, M.F.; Oliveira, R.N. Physically cross-linked gels of PVA with natural polymers as matrices for manuka honey release in wound-care applications. Materials 2019, 12, 559. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Liang, Z.; Yang, L.; Du, W.; Yu, T.; Tang, H.; Li, C.; Qiu, H. The role of natural polymers in bone tissue engineering. J. Control. Release 2021, 338, 571–582. [Google Scholar] [CrossRef]
- Lokanathan, Y.; Fauzi, M.B.; Man, R.C.; Rashidbenam, Z.; Saim, A.B.; Idrus, R.B.H.; Yunus, M.H.M. Article preliminary study on the development of in vitro human respiratory epithelium using collagen type i scaffold as a potential model for future tracheal tissue engineering. Appl. Sci. 2021, 11, 1787. [Google Scholar] [CrossRef]
- Wang, J.K.; Çimenoğlu, Ç.; Cheam, N.M.J.; Hu, X.; Tay, C.Y. Sustainable aquaculture side-streams derived hybrid biocomposite for bone tissue engineering. Mater. Sci. Eng. C 2021, 126, 112104. [Google Scholar] [CrossRef]
- Antoniac, I.V.; Antoniac, A.; Vasile, E.; Tecu, C.; Fosca, M.; Yankova, V.G.; Rau, J.V. In vitro characterization of novel nanostructured collagen-hydroxyapatite composite scaffolds doped with magnesium with improved biodegradation rate for hard tissue regeneration. Bioact. Mater. 2021, 6, 3383–3395. [Google Scholar] [CrossRef] [PubMed]
- Ide, K.; Takahashi, S.; Sakai, K.; Taga, Y.; Ueno, T.; Dickens, D.; Jenkins, R.; Falciani, F.; Sasaki, T.; Ooi, K.; et al. The dipeptide prolyl-hydroxyproline promotes cellular homeostasis and lamellipodia-driven motility via active β1-integrin in adult tendon cells. J. Biol. Chem. 2021, 297, 100819. [Google Scholar] [CrossRef] [PubMed]
- Park, B.-H.; Jeong, E.-S.; Lee, S.; Jang, J.-H. Bio-functionalization and in-vitro evaluation of titanium surface with recombinant fibronectin and elastin fragment in human mesenchymal stem cell. PLoS ONE 2021, 16, e0260760. [Google Scholar] [CrossRef]
- Li, K.; Zhang, L.; Li, J.; Xue, Y.; Zhou, J.; Han, Y. pH-Responsive ECM Coating on Ti Implants for Antibiosis in Reinfected Models. ACS Appl. Bio. Mater. 2022, 5, 344–354. [Google Scholar] [CrossRef]
- Bilodeau, C.; Goltsis, O.; Rogers, I.M.; Post, M. Limitations of recellularized biological scaffolds for human transplantation. J. Tissue Eng. Regen. Med. 2020, 14, 521–538. [Google Scholar] [CrossRef] [PubMed]
- Atala, A.; Bauer, S.B.; Soker, S.; Yoo, J.J.; Retik, A.B. Tissue-engineered autologous bladders for patients needing cystoplasty. Lancet 2006, 367, 1241–1246. [Google Scholar] [CrossRef]
- Böer, U.; Lohrenz, A.; Klingenberg, M.; Pich, A.; Haverich, A.; Wilhelmi, M. The effect of detergent-based decellularization procedures on cellular proteins and immunogenicity in equine carotid artery grafts. Biomaterials 2011, 32, 9730–9737. [Google Scholar] [CrossRef]
- Bayrak, A.; Tyralla, M.; Ladhoff, J.; Schleicher, M.; Stock, U.A.; Volk, H.D.; Seifert, M. Human immune responses to porcine xenogeneic matrices and their extracellular matrix constituents in vitro. Biomaterials 2010, 31, 3793–3803. [Google Scholar] [CrossRef]
- Antunes, M.; Bonani, W.; Reis, R.L.; Migliaresi, C.; Ferreira, H.; Motta, A.; Neves, N.M. Development of alginate-based hydrogels for blood vessel engineering. Mater. Sci. Eng. C Mater. Biol. Appl. 2021, 134, 112588. [Google Scholar] [CrossRef]
- Lee, S.; Lee, H.S.; Chung, J.J.; Kim, S.H.; Park, J.W.; Lee, K.; Jung, Y. Enhanced Regeneration of Vascularized Adipose Tissue with Dual 3D-Printed Elastic Polymer/dECM Hydrogel Complex. Int. J. Mol. Sci. 2021, 22, 2886. [Google Scholar] [CrossRef]
- Terpstra, M.L.; Li, J.; Mensinga, A.; De Ruijter, M.; Van Rijen, M.H.P.; Androulidakis, C.; Galiotis, C.; Papantoniou, I.; Matsusaki, M.; Malda, J.; et al. Bioink with cartilage-derived extracellular matrix microfibers enables spatial control of vascular capillary formation in bioprinted constructs. Biofabrication 2022, 14, 034104. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Park, T.H.; Ryu, J.Y.; Kim, D.K.; Oh, E.J.; Kim, H.M.; Shim, J.-H.; Yun, W.-S.; Huh, J.B.; Moon, S.H.; et al. Osteogenesis of 3d-printed pcl/tcp/bdecm scaffold using adipose-derived stem cells aggregates; an experimental study in the canine mandible. Int. J. Mol. Sci. 2021, 22, 5409. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Pérez, J.; Madden, P.W.; Ahearne, M. Multilayered Fabrication Assembly Technique to Engineer a Corneal Stromal Equivalent. Bio-protocol 2021, 11, e3963. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wang, W.; Lin, S.; Yang, Y.; Song, L.; Jing, Y.; Chen, L.; He, Z.; Li, W.; Xiong, A.; et al. Fabrication of a bio-instructive scaffold conferred with a favorable microenvironment allowing for superior implant osseointegration and accelerated in situ vascularized bone regeneration via type H vessel formation. Bioact. Mater. 2022, 9, 491–507. [Google Scholar] [CrossRef]
- Ebrahim, N.; Badr, O.A.M.; Yousef, M.M.; Hassouna, A.; Sabry, D.; Farid, A.S.; Mostafa, O.; Al Saihati, H.A.; Seleem, Y.; Aziz, E.A.E.; et al. Functional recellularization of acellular rat liver scaffold by induced pluripotent stem cells: Molecular evidence for wnt/b-catenin upregulation. Cells 2021, 10, 2819. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Class | Source and Tissue Type | Pre-Treatment Method | Extraction and Purification Method | Scaffold Comparisons | Reference |
|---|---|---|---|---|---|
| Mammalian | Human-placenta | Alkaline; 0.1 M NaOH in 1:10 (w/v) ratio; 6 h stir | Acid; 0.06 M acetic acid 1:25 (w/v) overnight; NaCl 3 M pH 7 to collect supernatant; dialysis against distilled water 5 days | -Cost effective (human waste) -Safe and fast extraction method | Karami et al., 2019 [91] |
| Bovine skin, bone, pericardium | Alkaline; 300 mL of 0.1 M NaOH at 4 °C 48 h; filtered 3X | Acid–enzyme; 300 mL 10 mM HCl with pepsin to solution ratio of 1:20; pH 2.0 4 °C 12 h; urea and NaCl to collect supernatant; 1 M Tris for pepsin deactivation; dialysis against 1 M phosphate buffer 4 °C 16 h | -Collagen in the purified form -Can be a substitute/replaced from the commercial collagen -Simple extraction method | Santos et al., 2013 [92] | |
| Rat tail | Salt; 10% NaCl 4 °C 24 h | Acid–enzyme; 0.5 M HCl and pepsin 1:50 4 °C 24 h; NaCl to collect supernatant and washed with phosphate buffer pH 7.4, 0.02 M | -Biocompatible -Induce contraction effect in in vitro model -Excellent for dental application | Techatanawat et al., 2011 [93] | |
| Goat tendon | Acid; 1% (v/v) acetic acid 4 °C 72 h | Acid; 1% v/v after cotton mesh filtration and dialysis against 10 mM PBS 48 h, then further dialysed against 0.05 M Na2 HPO4 | -In vitro and in vivo study towards HUVEC cells showed prominent result -Good for future angiogenesis study | Banerjee et al., 2012 [94] | |
| Sheep tendon | Acid; 0.35 M acetic acid 4 °C 24–48 h | Acid; 0.35 M acetic acid; NaCl salting-out 4 °C 24–48 h; supernatant undergoes dialysis against Na2HPO4 and PBS 14 kDa 4 °C 72 h | -Biocompatible toward human dermal fibroblasts (HDFs) -Greater availability and wider acceptability (religious views) -Can be fabricated into porous scaffolds | Fauzi et al., 2016 [71] | |
| Fish | Freshwater fish scales | Salt; 1.0 M NaCl, 0.05 M Tris HCl, 20.0 mM EDTA 48 h (pH7.5); demineralisation 0.5 M EDTA 48 h (pH 7.4) | Acid; 0.5 M acetic acid (pH 2.5) 48 h; NaCl (0.9 M) salting-out 24 h; resolubilised supernatant undergoes dialysis against 0.1 M acetic acid and deionised water 24 h | -Cost effective -Alternative to other collagen sources -Highly biocompatible | Pati et al., 2012 [95] |
| Bighead carp fins, scales, skin, bones, and swim bladders | Alkaline; 0.1 M NaOH 1:10 (w/v) ratio; 4 °C 36 h | Acid–enzyme; 0.5 M acetic acid 0.1% (w/v) pepsin 1:10 (w/v days; NaCl to collect supernatant 2 M; dialysis against distilled water 7 kda | -Alternative to other sources of collagen -Pepsin-solubilized collagen (PSC) can be extracted | Liu et al., 2012 [96] | |
| Loach skin | Alkaline–salt; 10% NaCl 1:5 (w/v) 4 °C 1 h. 0.1 mol/L NaOH 1:10(w/v) 4 °C 24 h | Acid–enzyme; 0.5 mol/L acetic acid 1:30(w/v) 4 °C 24 h 5% (w/v) pepsin 2X; NaCl 0.9 mol/L to collect supernatant; dialysis against distilled water 8 kDa | -Alternative to other sources of collagen -Acid-soluble collagen (ASC) and pepsin-soluble collagen (PSC) can be extracted | Wang et al., 2018 [97] | |
| Clown featherback skin | Alkaline; 0.1 M NaOH 1:15 (w/v) 4 °C 8 h | Acid–ultrasound; 0.5 M acetic acid 4 °C 48 h; in 30 min ultrasonication (20–80% between 10–30 min); NaCl to collect supernatant to 2.6 M; dialysis against distilled water for 2 days | -Ultrasonic application improves the extraction efficiency -Higher yield production, alternative to other collagen sources | Petcharat et al., 2020 [89] | |
| Marine | Jumbo squid mantles | Acid; 6 M urea in sodium acetate (pH 6.8) and neutral buffer 24 h | Acid; 0.5 M acetic acid 24 h; supernatant collected for acid soluble collagen | -Acid-soluble collagen (ASC) can be extracted -Have some similar characteristics to bovine collagen | Uriarte-Montoya et al., 2010 [98] |
| Jellyfish tissues | Acid; 0.5 M acetic acid | Acid–enzyme; 0.5 M acetic acid centrifuged and added pepsin (1–15 mg/mg tissue) overnight; NaCl (0.9 M) used to collect supernatant and dialysed against 0.1 M acetic acid | -Alternative to other sources of collagen -Biocompatible in in vitro testing | Addad et al., 2011 [99] | |
| Jellyfish and squid | Acid; 0.5 M acetic acid 1:15 (w/v) 4 °C 3 days | Acid; 0.5 M acetic acid 1:15 (w/v) 4 °C 3 days after filtration through cheesecloth; NaCl (0.9 M) and 0.05 M Tris and used to collect supernatant and dialysed against 0.1 M acetic acid 3 days and distilled water another 3 days | -Alternative to other sources of collagen -Pepsin-soluble collagen (PSC) can be extracted | Jankangram et al., 2016 [100] | |
| Amphibian | Bullfrog fallopian tubes | Alkali; 0.1 M NaOH 4 °C 1 day | Acid–enzyme; 0.5 M acetic acid with 10% pepsin 4 °C 2 days; NaCl (0.7 M) to collect supernatant; resolubilised acid solution dialysed against 0.1 M acetic acid 1 day; distilled water 2 days | -Pepsin-soluble collagen (PSC) can be extracted -Potential alternative and supplements from other sources of collagen | Wang et al., 2011 [101] |
| Avian | Skin, skin dermis | Acid (acetic acid) | Acid-based extraction (acetic acid); enzymatic method (pepsin) | -Non-immunogenic and non-allergenic -Biocompatible -Can be fabricated into porous scaffolds | Peng et al., 2010, Parenteau-Bareil et al. 2011 [102,103] |
| Chicken cartilage | Acid (EDTA) | Salting-out (NaCl) + ultrasound | -Increased yield production through ultrasound -Pepsin-soluble collagen (PSC) can be extracted | Akram and Zhang, 2020 [88] | |
| Chicken lungs | Acid salt (sodium carbonate) | Acid extraction (acetic acid) + salting-out (NaCl) + enzymatic method (pepsin) + ultrasound | -Increased yield production through ultrasound -Alternative source to mammalian collagen -Higher thermal stability | Zou et al., 2020 [87] |
| Characterisation | Method | Expectation | Reference |
|---|---|---|---|
| Biocompatibility | |||
| Biocompatibility and immunogenicity | MTT assay Live/dead assay Cell attachment Immunocytochemistry (ICC) | Cell proliferation and growth; SEM; more than 80% cell adhesion after 24 h; integrin-related protein expression and cell attachment | Addad et al., 2011 [99], Fauzi et al., 2016 [71], Fauzi et al., 2017 [104], Thievessen et al., 2015 [105] |
| Physical, Morphological, and Topographical (PMT) Characterisation for Three-dimensional stability | |||
| Weight | SDS-PAGE | Type I collagen is composed of β (250 kDa), α1 (130 kDa), and α2 (115 kDa) | Peng et al., 2010 [102], Parenteau-Bareil et al., 2010 [106], Fauzi et al., 2016 [71] Inanc et al., 2017 [107] |
| Native conformation | UV-circular dichroism (CD) spectroscopy | CD with a positive maximum absorption band at around 222 nm | Carvalho et al., 2018 [108] |
| Mechanical strength | Tensile strength and Young’s modulus | No gold standard, but fish and reptile should be more fragile than mammalian material | Amri et al. 2014 [109], Teramoto et al. 2012 [110] |
| Thermal stability | Thermogravimetry analysis (TGA) Differential scanning calorimetry (DSC) | No gold standard for what Td of biomaterial should be, but it must be stable to use normal temperatures or higher; in general, comparable or higher than native rat tail tendon collagen fibre Td of 65 °C; Td of extracted collagen in solution are 37–40 °C, 26- °C, and 6–20 °C, for mammalians, fish, and deep-sea animals, respectively | Miles and Bailey 1999 [111], Zhang et al., 2020 [112], Subhan et al., 2015 [113], Bozec and Odlyha, 2011 [114] |
| Porosity and pore size | SEM | Pore size > 80 µm for fibrogenic and <20 µm for chondrogenic growth; the average mean size for ovine-, bovine-, and porcine-derived scaffolds are 73.05 ± 10.79 µm, 85.84 ± 9.51 µm, and 87.32 ± 10.69 µm, respectively | Ghodbane and Dunn 2016 [115] |
| Biodegradation | Enzymatic biodegradation method | Depends on application example within 14 days for cutaneous wound healing | Mh Busra et al., 2019 [116] Salleh et al., 2022 [117] |
| Swelling ratio | Swelling ratio protocol | 1000–2700% for biomaterials | Ghodbane and Dunn 2016 [115] |
| Water vapor transmission rate (WVTR) | WVTR method | Within range 2028.3 ± 237.8 g/m2/day to maintain a moist environment and enhance the normal healing phase | Xu et al., 2016 [118] |
| Surface and particle physicality | Contact angle zeta potential | Angle < 90° hydrophilic; isoelectric points (pIs) close to 6 at zero zeta potential | Chen et al., 2019 [119] |
| Chemical Characterisation | |||
| X-ray photoelectron spectroscopy (XPS) | XPS | Samples should show ≈0.1 atomic % as nominal sensitivity with an elemental sensitivity that may differ as much as ≈100; for the assessment of chemical components, a sample size larger than ≈10 μm will be convenient | Baer et al., 2019 [120] |
| Fourier transform infrared (FTIR) | FTIR | Collagen type I functional groups include amides I, II, and III; range of peak intensity between 1450 cm−1 and 1235 cm−1 commonly indicates the helical structure of collagen; amide A at the higher peak intensity of 3350 cm−1 can be attributed to collagen type I; At peak intensity of 1632 cm−1, this indicates the higher-order arrangement of the collagen structure, which refers to β-sheet and triple helix structure | Sasmal and Begam 2014 [121] Fauzi et al., 2016 [71] |
| Energy dispersive X-ray (EDX) | EDX | Major elements in collagen type 1 are oxygen, nitrogen, and carbon with a higher percentage of oxygen, followed by nitrogen and carbon | Fauzi et al., 2014 [122] |
| X-ray diffraction (XRD). | XRD | Collagen XRD generally consists of 2 clear peaks, where the first peak is sharper than the second peak; collagen type I from different sources of mammalian, avian, marine, fish, etc., via XRD has been proven closer to the amorphous phase rather than crystallinity | Zhang et al., 2011 [123], Fauzi et al., 2016 [71], León-Mancilla et al., 2016 [124] |
| Table | Mechanism | Main Characteristics | ||
|---|---|---|---|---|
| Pros | Cons | |||
| Physical | Dehydrothermal treatment (DHT) | Water is removed as amide bonds are formed between amine and carboxyl groups of collagens (inter-molecular). Lysino-alanine bonds are also formed between lysine residues and dehydro-alanine in collagen chains (intra-molecular) [112,144]. | Safe. Non-toxic to cells. Thermally stable. Sterilization. | May cause denaturation. Low collagenase resistance [153]. |
| Ultraviolet irradiation (UV) | Non-ionizing irradiation to induce collagen amino acids residues—Trp, Phe, and Tyr—to generate free radicals to cause inter-molecular crosslinks [146]. | Rapid process. Non-toxic to cells. Sterilization. | May cause denaturation. Poor crosslinking strength. | |
| Gamma ray/electron beams | Free radical mediation—hydroxyl radicals from amino acid residues—Phe, His, and Met—form intermolecular crosslinks [164]. Furthermore, two tyrosyl radicals can form dityrosine bonds and crosslink [165]. | |||
| Chemical | Glutaraldehyde (GA) | Aldehydes crosslink collagen by forming Schiff bases between the ε-amino groups from lysine or hydroxylysine residues and the aldehyde groups of aldehyde [112]. | Low cost. High reactivity. High water solubility. | Potential cytotoxicity to cells. |
| Dialdehyde starch | Low cytotoxicity. Biodegradable. Resistant to collagenase. | Costly. Sensitive to oxidation [166]. | ||
| Epoxy compounds | Epoxy groups form multipoint crosslinking with ε-amino groups of lysyl residues at alkali condition [167]. | Makes tissues more pliable [168]. Biocompatible with corneal cells [152]. | Affected by pH [167]. | |
| N-hydroxysuccinimide (NHS) | Carboxyl groups from polycarboxylic acids are turned into active ester groups, which then crosslink with free amino groups within different collagen chains [147]. | Highly resistant to collagenase [142]. | Requires thorough washing. | |
| 1-ethyl-3(3-dimethylaminopropyl)-carbodiimide (EDC) | Crosslinking via amido bond between carboxyl groups and amino groups in adjacent collagen chains. EDC activates carboxyl groups (Glu and Asp) into active O-acylisourea groups, which undergo nucleophilic attack by free amine groups (lys or hyl), forming amido bonds. Usually, NHS is added to catalyse the reaction (EDC/NHS crosslinking) [169,170]. | Stronger crosslinking effect. Zero-length crosslinker and intra-fibrillar. Low cytotoxicity. | Can cause excessive crosslinking and stiffness. | |
| Diphenylphosphorylazide (DPPA) | Converts carboxyl groups into acylazide groups, which react with the free amino groups of adjacent collagen chains (amido bonds) [148]. | Fast. Non-toxic. Resistant to collagenase. | Difficult to remove solvent. | |
| Biological | Genipin | Crosslinking by initiating a nucleophilic attack on amine with two amino groups. The genipin C3-atom undergoes nucleophilic attack from primary amine groups (lys, hyl, arg), which then reacts with another collagen amino group by replacing an oxygen on the genipin ring, forming a nitrogen–iridoid (heterocyclic compound). Two iridoids are linked through a radical reaction to form intramolecular and intermolecular crosslinks [155,156] | Natural resources, biocompatible with cells. Resistant to collagenase. | Excessive concentration will reduce scaffold mechanical strength. Blue reaction after crosslinking. Sensitive to light and oxidation. |
| Transglutaminase (TG) | Catalysed γ-carboxamide groups of gln residues become the acyl donor for acyl receptors, which are the ε-amino groups of lys residues or primary amino groups. Then, ε-(γ-glutamyl) lysine bonds form intra- and intermolecular bonds [158]. | Non-toxic, good biocompatibility. Similar to natural crosslinking. | Enzyme deregulation may cause harmful diseases. | |
| Plant polyphenols/procyanidin/proanthocyanidins (PA) | Multipoint hydrogen bonds formed between phenolic hydroxyl groups of polyphenols and hydroxyl, carboxyl, amino, or amide groups of collagen chains [162] | Very low cytotoxicity. Antioxidants. Anticancer, antimicrobial, antiangiogenic, and anti-inflammatory [163]. | Sensitivity to thermal treatment, light, and oxidation. | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amirrah, I.N.; Lokanathan, Y.; Zulkiflee, I.; Wee, M.F.M.R.; Motta, A.; Fauzi, M.B. A Comprehensive Review on Collagen Type I Development of Biomaterials for Tissue Engineering: From Biosynthesis to Bioscaffold. Biomedicines 2022, 10, 2307. https://doi.org/10.3390/biomedicines10092307
Amirrah IN, Lokanathan Y, Zulkiflee I, Wee MFMR, Motta A, Fauzi MB. A Comprehensive Review on Collagen Type I Development of Biomaterials for Tissue Engineering: From Biosynthesis to Bioscaffold. Biomedicines. 2022; 10(9):2307. https://doi.org/10.3390/biomedicines10092307
Chicago/Turabian StyleAmirrah, Ibrahim N., Yogeswaran Lokanathan, Izzat Zulkiflee, M. F. Mohd Razip Wee, Antonella Motta, and Mh Busra Fauzi. 2022. "A Comprehensive Review on Collagen Type I Development of Biomaterials for Tissue Engineering: From Biosynthesis to Bioscaffold" Biomedicines 10, no. 9: 2307. https://doi.org/10.3390/biomedicines10092307
APA StyleAmirrah, I. N., Lokanathan, Y., Zulkiflee, I., Wee, M. F. M. R., Motta, A., & Fauzi, M. B. (2022). A Comprehensive Review on Collagen Type I Development of Biomaterials for Tissue Engineering: From Biosynthesis to Bioscaffold. Biomedicines, 10(9), 2307. https://doi.org/10.3390/biomedicines10092307