
Reversal of Azole Resistance in Candida albicans by Human Neutrophil Peptide



Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Strains
2.2. Antifungal Drug Susceptibility Testing
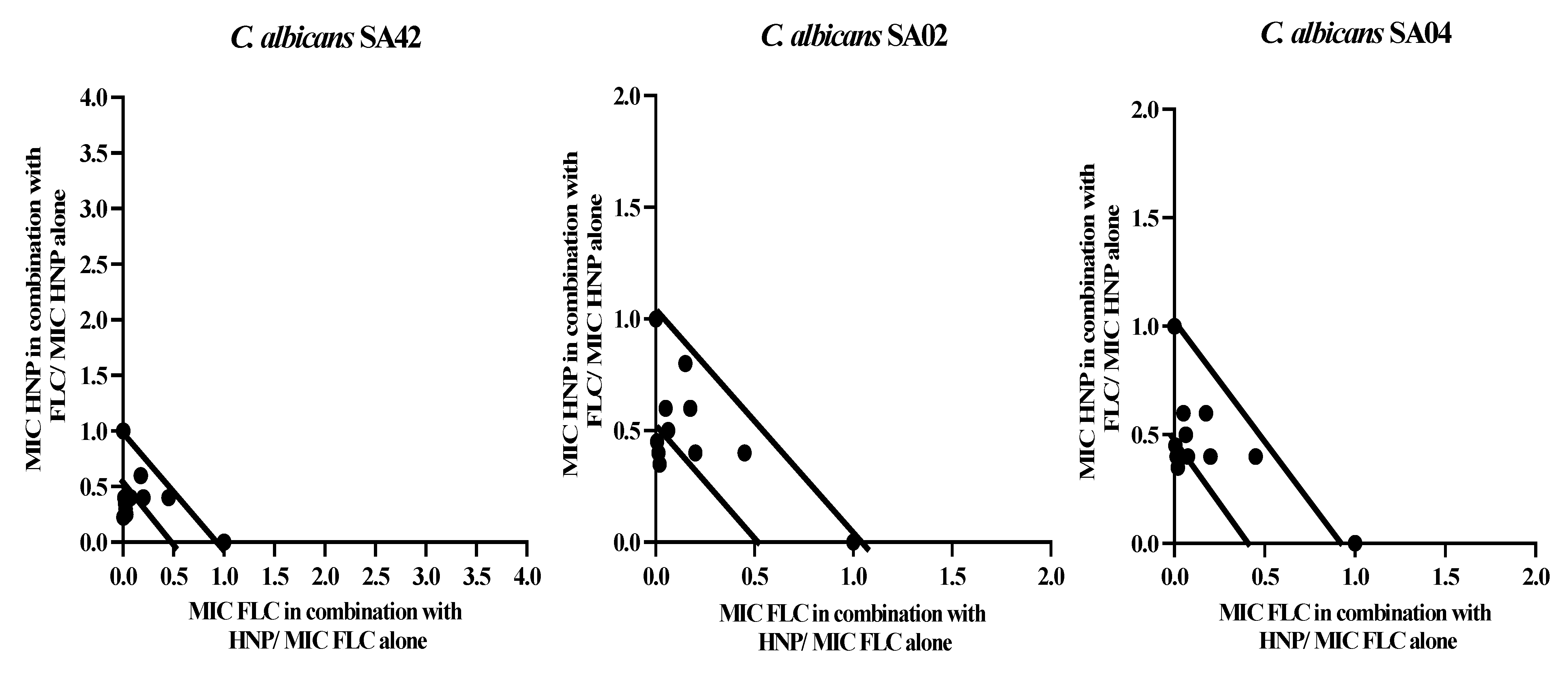
2.3. Combination Interactions and Isobolograms
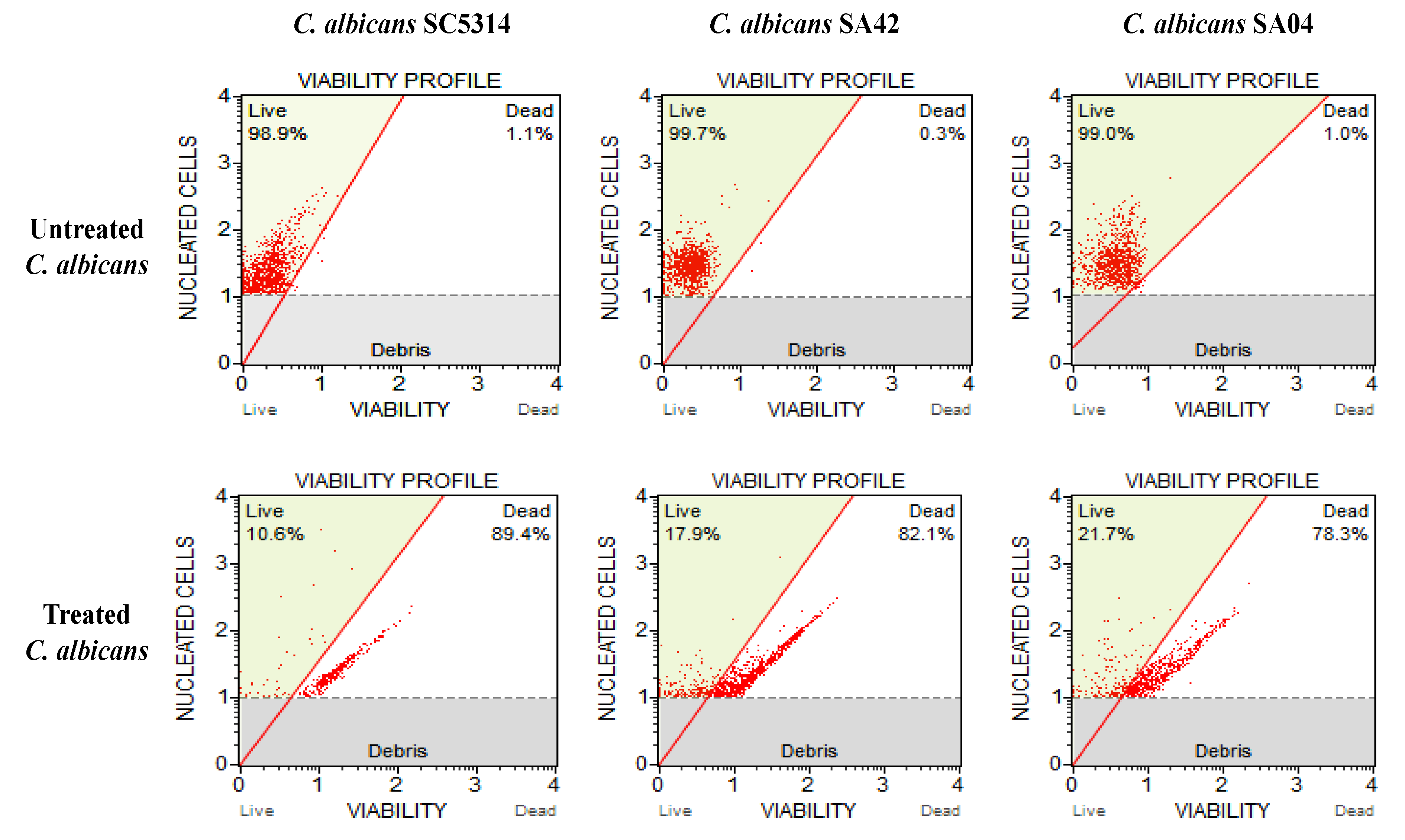
2.4. MUSE Viability Assay
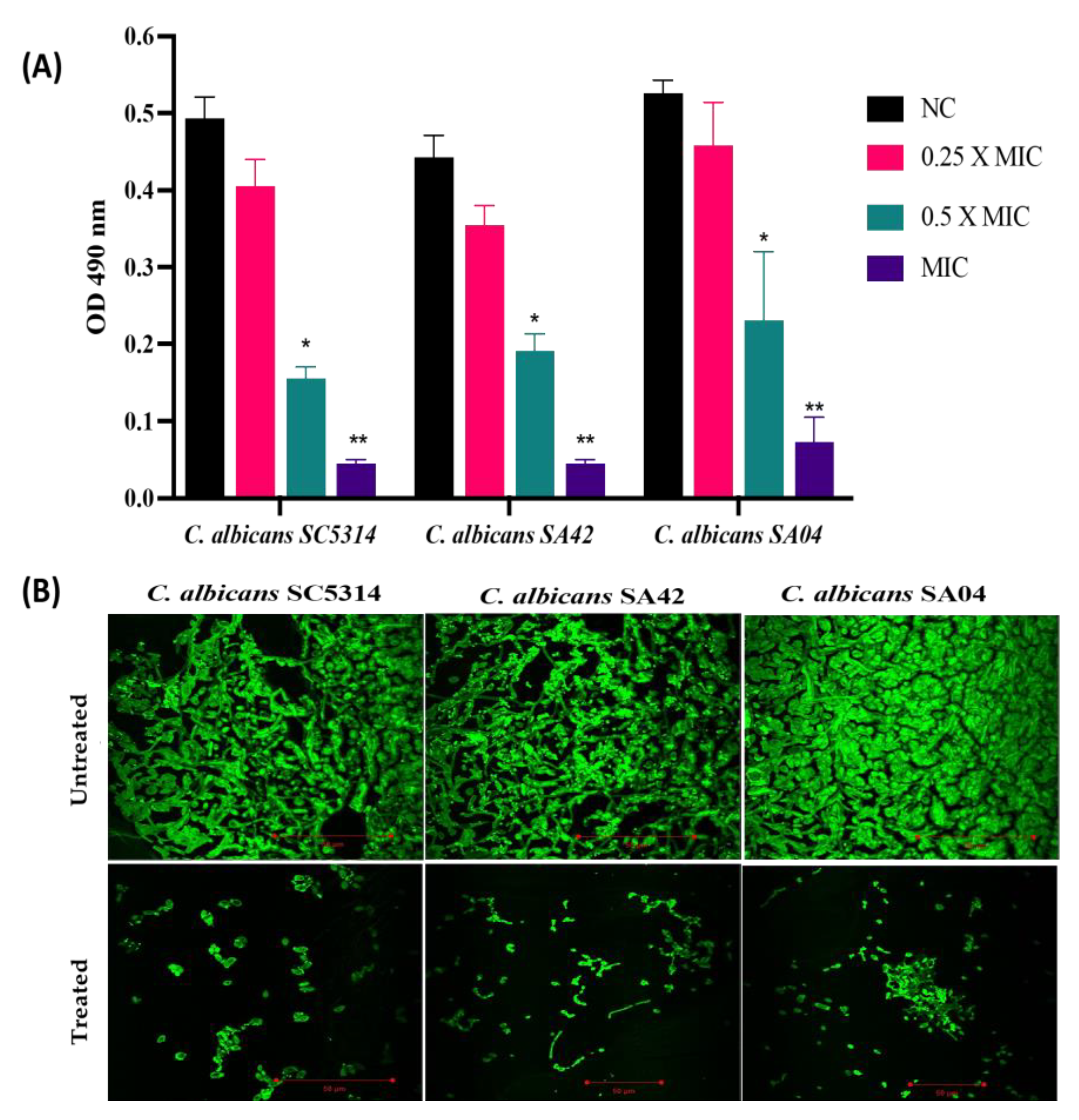
2.5. Biofilm Assay
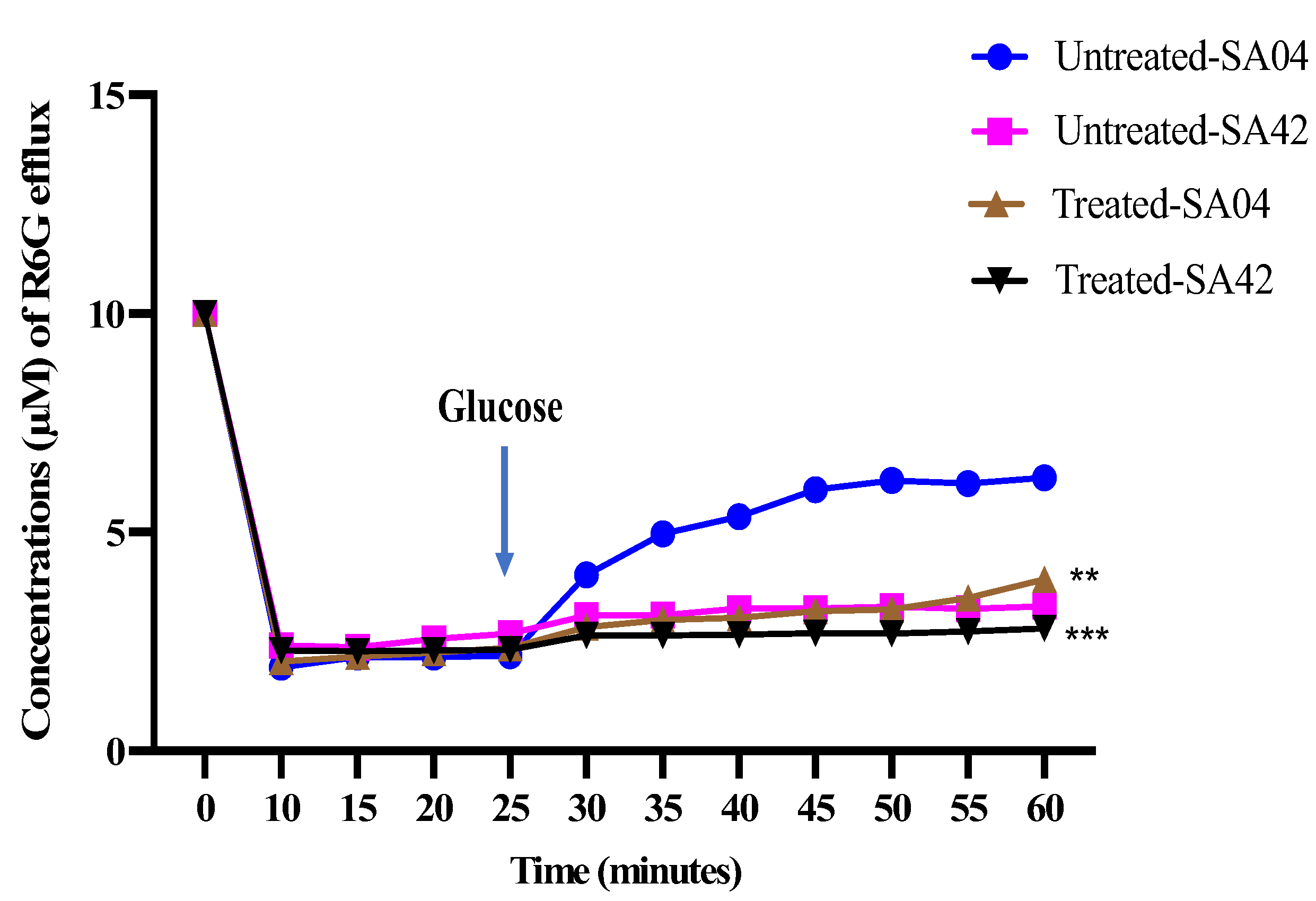
2.6. Efflux Pumps Inhibitory Activity
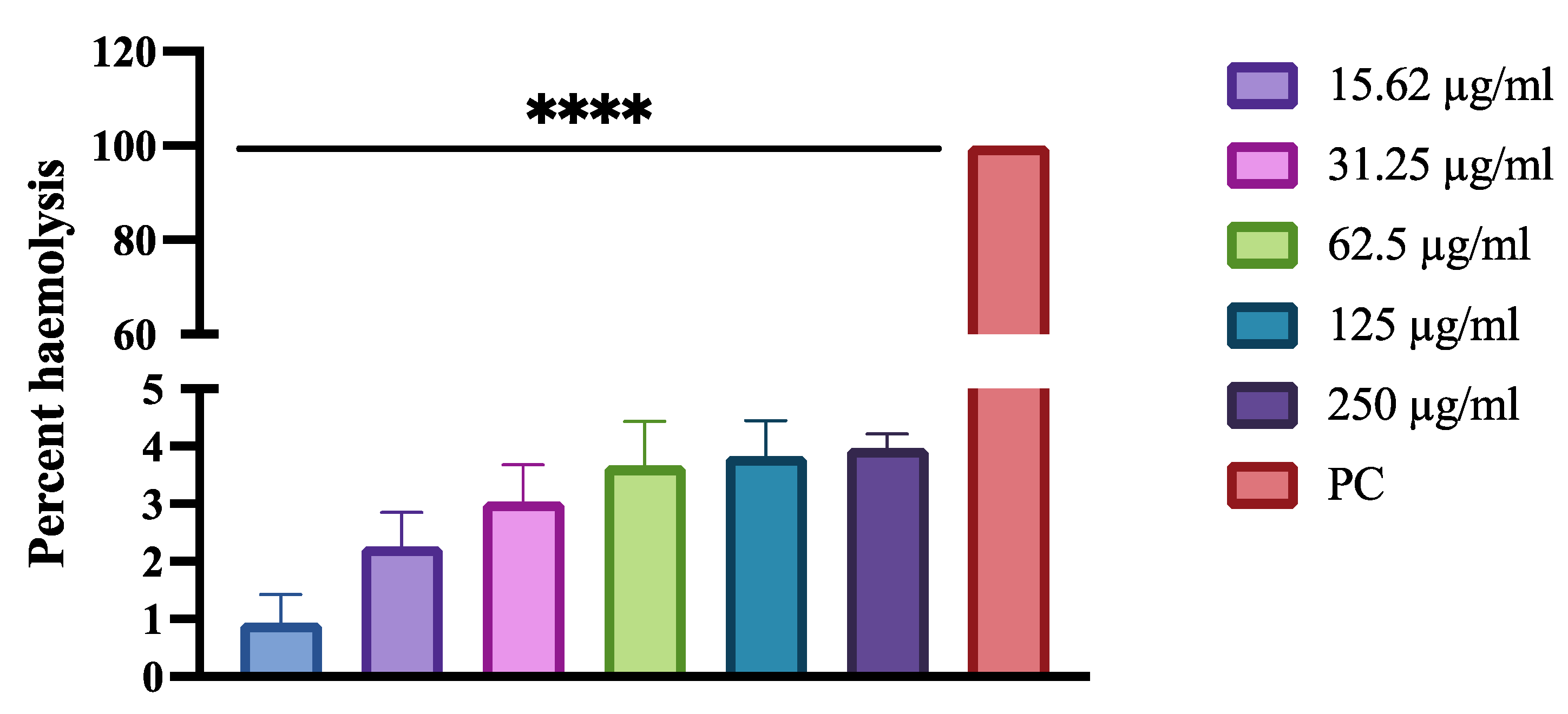
2.7. Cytotoxicity Studies
2.8. Statistical Analysis
3. Results
3.1. Antifungal Susceptibility
3.2. Combinations of HNP with FLC
Combinations in 1:1 Ratio
3.3. Combinations in Varied Ratios
3.4. MUSE Viability Testing
3.5. Biofilm Inhibition Property of HNP
3.6. Efflux Pumps Inhibitory Activity of HNP
3.7. Cytotoxicity Effects
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaplan, J.E.; Hanson, D.; Dworkin, M.S.; Frederick, T.; Bertolli, J.; Lindegren, M.L.; Holmberg, S.; Jones, J.L. Epidemiology of Human Immunodeficiency Virus-Associated Opportunistic Infections in the United States in the Era of Highly Active Antiretroviral Therapy. Clin. Infect. Dis. 2000, 30, S5–S14. [Google Scholar] [CrossRef]
- Pappas, P.G.; Kauffman, C.A.; Andes, D.R.; Clancy, C.J.; Marr, K.A.; Ostrosky-Zeichner, L.; Reboli, A.C.; Schuster, M.G.; Vazquez, J.A.; Walsh, T.J.; et al. Clinical Practice Guideline for the Management of Candidiasis: 2016 Update by the Infectious Diseases Society of America. Clin. Infect. Dis. 2015, 62. [Google Scholar]
- Pfaller, M.A.; Diekema, D.J.; Procop, G.W.; Rinaldi, M.G. Multicenter Comparison of the VITEK 2 Antifungal Susceptibility Test with the CLSI Broth Microdilution Reference Method for Testing Amphotericin B, Flucytosine, and Voriconazole against Candida spp. J. Clin. Microbiol. 2007, 45, 3522–3528. [Google Scholar] [CrossRef] [PubMed]
- Robbins, N.; Wright, G.D.; Cowen, L.E. Antifungal Drugs: The Current Armamentarium and Development of New Agents. In The Fungal Kingdom; ASM Press: Washington, DC, USA, 2017. [Google Scholar]
- Rather, I.A.; Sabir, J.S.M.; Asseri, A.H.; Ali, S. Antifungal Activity of Human Cathelicidin LL-37, a Membrane Disrupting Peptide, by Triggering Oxidative Stress and Cell Cycle Arrest in Candida Auris. J. Fungi 2022, 8, 204. [Google Scholar] [CrossRef]
- Marichal, P.; Koymans, L.; Willemsens, S.; Bellens, D.; Verhasselt, P.; Luyten, W.; Borgers, M.; Ramaekers, F.C.S.; Odds, F.C.; Bossche, H.V. Contribution of mutations in the cytochrome P450 14α-demethylase (Erg11p, Cyp51p) to azole resistance in Candida albicans. Microbiology 1999, 145, 2701–2713. [Google Scholar] [CrossRef]
- Franz, R.; Kelly, S.L.; Lamb, D.C.; Kelly, D.E.; Ruhnke, M.; Morschhäuser, J. Multiple Molecular Mechanisms Contribute to a Stepwise Development of Fluconazole Resistance in Clinical Candida albicans Strains. Antimicrob. Agents Chemother. 1998, 42, 3065–3072. [Google Scholar] [CrossRef] [PubMed]
- Sanglard, D.; Ischer, F.; Monod, M.; Bille, J. Cloning of Candida albicans genes conferring resistance to azole antifungal agents: Characterization of CDR2, a new multidrug ABC transporter gene. Microbiology 1997, 143, 405–416. [Google Scholar] [CrossRef] [PubMed]
- Cannon, R.D.; Lamping, E.; Holmes, A.R.; Niimi, K.; Baret, P.; Keniya, M.V.; Tanabe, K.; Niimi, M.; Goffeau, A.; Monk, B.C. Efflux-Mediated Antifungal Drug Resistance. Clin. Microbiol. Rev. 2009, 22, 291–321. [Google Scholar] [CrossRef] [PubMed]
- Ahmad Rather, I.; Seo, B.J.; Rejish Kumar, V.J.; Choi, U.-H.; Choi, K.-H.; Lim, J.H.; Park, Y.-H. Isolation and characterization of a proteinaceous antifungal compound from Lactobacillus plantarum YML007 and its application as a food preservative. Lett. Appl. Microbiol. 2013, 57, 69–76. [Google Scholar] [CrossRef]
- Ahmad, A.; Khan, A.; Manzoor, N. Reversal of efflux mediated antifungal resistance underlies synergistic activity of two monoterpenes with fluconazole. Eur. J. Pharm. Sci. 2013, 48, 80–86. [Google Scholar] [CrossRef]
- Ahmad, A.; Wani, M.Y.; Khan, A.; Manzoor, N.; Molepo, J. Synergistic Interactions of Eugenol-Tosylate and Its Congeners with Fluconazole against Candida Albicans. PLoS ONE 2015, 10, e0145053. [Google Scholar] [CrossRef]
- Bapat, P.; Singh, G.; Nobile, C. Visible Lights Combined with Photosensitizing Compounds Are Effective against Candida albicans Biofilms. Microorganisms 2021, 9, 500. [Google Scholar] [CrossRef]
- Pierce, C.G.; Uppuluri, P.; Tristan, A.R.; Wormley, F.L.; Mowat, E.; Ramage, G.; Lopez-Ribot, J.L. A simple and reproducible 96-well plate-based method for the formation of fungal biofilms and its application to antifungal susceptibility testing. Nat. Protoc. 2008, 3, 1494–1500. [Google Scholar] [CrossRef] [PubMed]
- Zohra, M.; Fawzia, A. Hemolytic Activity of Different Herbal Extracts Used in Algeria. Int. J. Pharm. Sci. Res. 2014, 5, 495–500. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 27th ed; CLSI supplement M100; International journal of STD & AIDS; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2017; p. 18. [Google Scholar]
- Becher, R.; Wirsel, S.G.R. Fungal cytochrome P450 sterol 14α-demethylase (CYP51) and azole resistance in plant and human pathogens. Appl. Microbiol. Biotechnol. 2012, 95, 825–840. [Google Scholar] [CrossRef]
- Leonart, L.; Tonin, F.; Ferreira, V.L.; Penteado, S.T.D.S.; Motta, F.D.A.; Pontarolo, R. Fluconazole Doses Used for Prophylaxis of Invasive Fungal Infection in Neonatal Intensive Care Units: A Network Meta-Analysis. J. Pediatr. 2017, 185, 129–135.e6. [Google Scholar] [CrossRef] [PubMed]
- Adedokun, E.O.; Rather, I.A.; Bajpai, V.K.; Choi, K.-H.; Park, Y.-H. Isolation and Characterization of Lactic Acid Bacteria from Nigerian Fermented Foods and Their Antimicrobial Activity. J. Pure Appl. Microbiol. 2014, 8, 3411–3420. [Google Scholar]
- Rather, I.A.; Kim, B.-C.; Bajpai, V.K.; Park, Y.-H. Self-medication and antibiotic resistance: Crisis, current challenges, and prevention. Saudi J. Biol. Sci. 2017, 24, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Quinn, K.; Henriques, M.; Parker, T.; Slutsky, A.S.; Zhang, H. Human neutrophil peptides: A novel potential mediator of inflammatory cardiovascular diseases. Am. J. Physiol. Circ. Physiol. 2008, 295, H1817–H1824. [Google Scholar] [CrossRef]
- Ehmann, D.; Koeninger, L.; Wendler, J.; Malek, N.P.; Stange, E.F.; Wehkamp, J.; Jensen, B.A.H. Fragmentation of Human Neutrophil α-Defensin 4 to Combat Multidrug Resistant Bacteria. Front. Microbiol. 2020, 11, 1147. [Google Scholar] [CrossRef]
- Delattin, N.; Cammue, B.P.; Thevissen, K. Reactive oxygen species-inducing antifungal agents and their activity against fungal biofilms. Futur. Med. Chem. 2014, 6, 77–90. [Google Scholar] [CrossRef]
- Desai, J.; Mitchell, A.P.; Andes, D. Fungal Biofilms, Drug Resistance, and Recurrent Infection. Cold Spring Harb. Perspect. Med. 2014, 4, a019729. [Google Scholar] [CrossRef]
- Ramage, G.; Rajendran, R.; Sherry, L.; Williams, C. Fungal Biofilm Resistance. Int. J. Microbiol. 2012, 2012, 1–14. [Google Scholar] [CrossRef]
- Sherry, L.; Rajendran, R.; Lappin, D.F.; Borghi, E.; Perdoni, F.; Falleni, M.; Tosi, D.; Smith, K.; Williams, C.; Jones, B.; et al. Biofilms formed by Candida albicans bloodstream isolates display phenotypic and transcriptional heterogeneity that are associated with resistance and pathogenicity. BMC Microbiol. 2014, 14, 182. [Google Scholar] [CrossRef]
- Aguiar, F.; Santos, N.; Cavalcante, C.D.P.; Andreu, D.; Baptista, G.; Gonçalves, S. Antibiofilm Activity on Candida albicans and Mechanism of Action on Biomembrane Models of the Antimicrobial Peptide Ctn[15–34]. Int. J. Mol. Sci. 2020, 21, 8339. [Google Scholar] [CrossRef]
- Chairatana, P.; Chiang, I.-L.; Nolan, E.M. Human α-Defensin 6 Self-Assembly Prevents Adhesion and Suppresses Virulence Traits of Candida albicans. Biochemistry 2017, 56, 1033–1041. [Google Scholar] [CrossRef]
- Oshiro, K.G.N.; Rodrigues, G.; Monges, B.E.D.; Cardoso, M.H.; Franco, O.L. Bioactive Peptides Against Fungal Biofilms. Front. Microbiol. 2019, 10, 2169. [Google Scholar] [CrossRef] [PubMed]
- Perez-Rodriguez, A.; Eraso, E.; Quindós, G.; Mateo, E. Antimicrobial Peptides with Anti-Candida Activity. Int. J. Mol. Sci. 2022, 23, 9264. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.S.A.; Malik, A.; Ahmad, I. Anti-candidal activity of essential oils alone and in combination with amphotericin B or fluconazole against multi-drug resistant isolates of Candida albicans. Med Mycol. 2012, 50, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Gbelska, Y.; Krijger, J.-J.; Breunig, K.D. Evolution of gene families: The multidrug resistance transporter genes in five related yeast species. FEMS Yeast Res. 2006, 6, 345–355. [Google Scholar] [CrossRef]
- Tong, Y.; Liu, M.; Zhang, Y.; Liu, X.; Huang, R.; Song, F.; Dai, H.; Ren, B.; Sun, N.; Pei, G.; et al. Beauvericin counteracted multi-drug resistant Candida albicans by blocking ABC transporters. Synth. Syst. Biotechnol. 2016, 1, 158–168. [Google Scholar] [CrossRef] [PubMed]
- Holmes, A.R.; Cardno, T.S.; Strouse, J.J.; Ivnitski-Steele, I.; Keniya, M.V.; Lackovic, K.; Monk, B.C.; A Sklar, L.; Cannon, R.D. Targeting efflux pumps to overcome antifungal drug resistance. Future Med. Chem. 2016, 8, 1485–1501. [Google Scholar] [CrossRef] [PubMed]
- Syeda, F.; Liu, H.-Y.; Tullis, E.; Liu, M.; Slutsky, A.S.; Zhang, H. Differential signaling mechanisms of HNP-induced IL-8 production in human lung epithelial cells and monocytes. J. Cell. Physiol. 2008, 214, 820–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Isolates | HNB | FLC | ||
|---|---|---|---|---|
| MIC (µg/mL) | MFC (µg/mL) | MIC (µg/mL) | ||
| FLC susceptible | SC5314 | 7.813 | 15.625 | 0.5 |
| SA91 | 7.813 | 15.625 | 0.5 | |
| SA19 | 7.813 | 31.25 | 0.5 | |
| SA42 | 15.625 | 31.25 | 0.5 | |
| FLC resistant | SA02 | 31.25 | 62.5 | 125 |
| SA61 | 15.625 | 62.5 | 125 | |
| SA04 | 62.5 | 250 | 250 | |
| Candida Isolates | FICA | FICB | ΣFIC | INT | |
|---|---|---|---|---|---|
| FLC susceptible | SC5314 | 0.250 | 0.500 | 0.750 | ADD |
| SA91 | 0.125 | 0.500 | 0.625 | ADD | |
| SA19 | 1.000 | 0.063 | 1.063 | IND | |
| SA42 | 0.031 | 0.250 | 0.281 | SYN | |
| FLC resistant | SA02 | 0.250 | 0.250 | 0.500 | SYN |
| SA61 | 0.500 | 0.063 | 0.563 | ADD | |
| SA04 | 0.031 | 0.125 | 0.156 | SYN | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, M.I.; Choudhry, H.; Jahan, S.; Rather, I.A. Reversal of Azole Resistance in Candida albicans by Human Neutrophil Peptide. Biomedicines 2023, 11, 513. https://doi.org/10.3390/biomedicines11020513
Khan MI, Choudhry H, Jahan S, Rather IA. Reversal of Azole Resistance in Candida albicans by Human Neutrophil Peptide. Biomedicines. 2023; 11(2):513. https://doi.org/10.3390/biomedicines11020513
Chicago/Turabian StyleKhan, Mohammad Imran, Hani Choudhry, Sadaf Jahan, and Irfan A. Rather. 2023. "Reversal of Azole Resistance in Candida albicans by Human Neutrophil Peptide" Biomedicines 11, no. 2: 513. https://doi.org/10.3390/biomedicines11020513
APA StyleKhan, M. I., Choudhry, H., Jahan, S., & Rather, I. A. (2023). Reversal of Azole Resistance in Candida albicans by Human Neutrophil Peptide. Biomedicines, 11(2), 513. https://doi.org/10.3390/biomedicines11020513