

Loss of the Novel Mitochondrial Membrane Protein FAM210B Is Associated with Hepatocellular Carcinoma

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Retrieval and Analysis
2.2. Tissue Sample Collection
2.3. Survival Analysis
2.4. Cell Culture
2.5. RNA Extraction and Real-Time Quantitative Reverse Transcription PCR Detection
2.6. Construction of Stable Cell Lines
2.7. Cell Counting Kit-8 (CCK-8) Assay
2.8. Plate Cloning Experiments
2.9. Soft-Agar Clone-Forming Assay
2.10. Migration and Invasion Assays
2.11. Western Blot
2.12. Immunohistochemistry (IHC) Staining
2.13. Tumor Xenograft Model
2.14. Statistical Analysis
3. Results
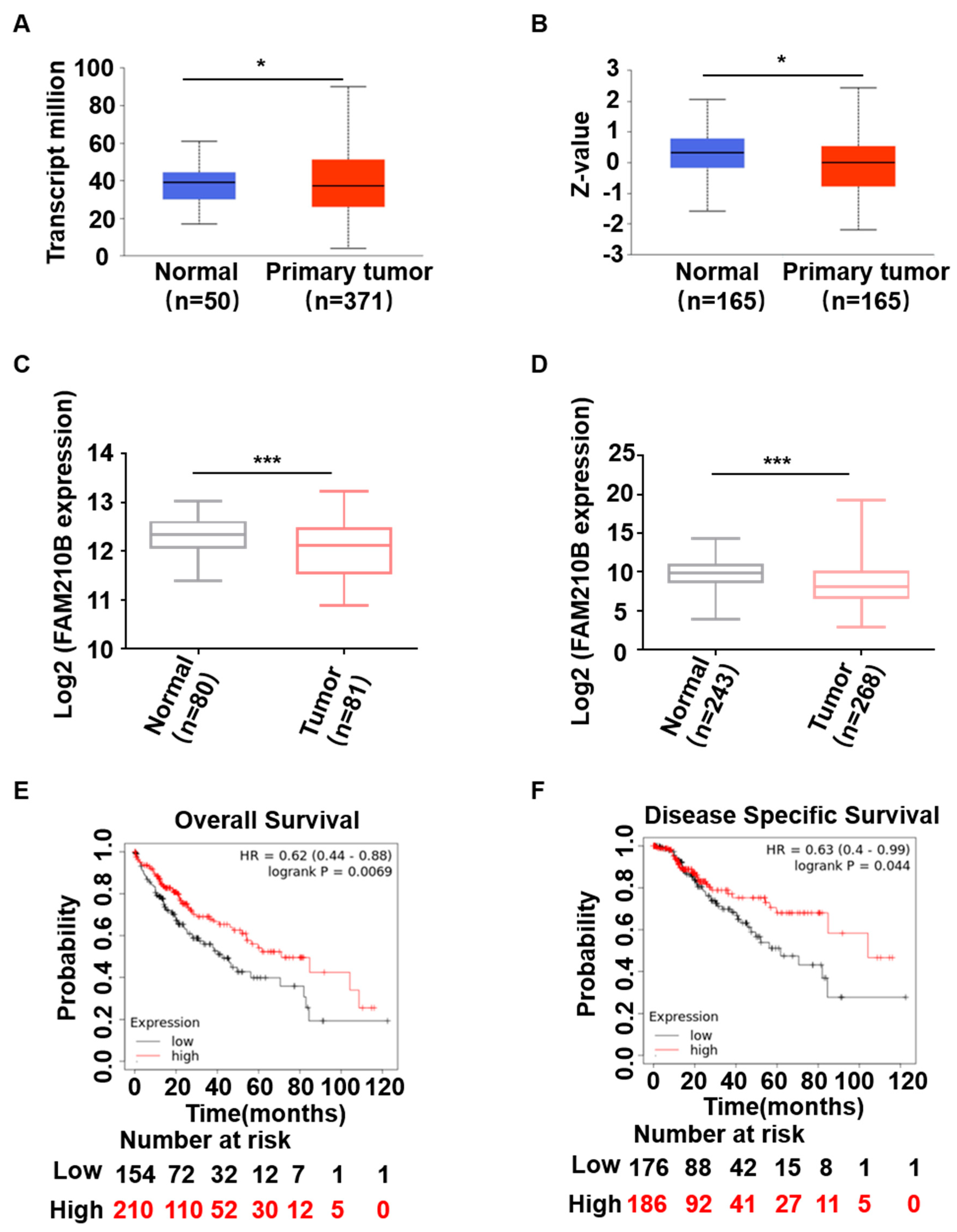
3.1. Patterns and Prognostic Implications of FAM210B Expression in HCC Patients
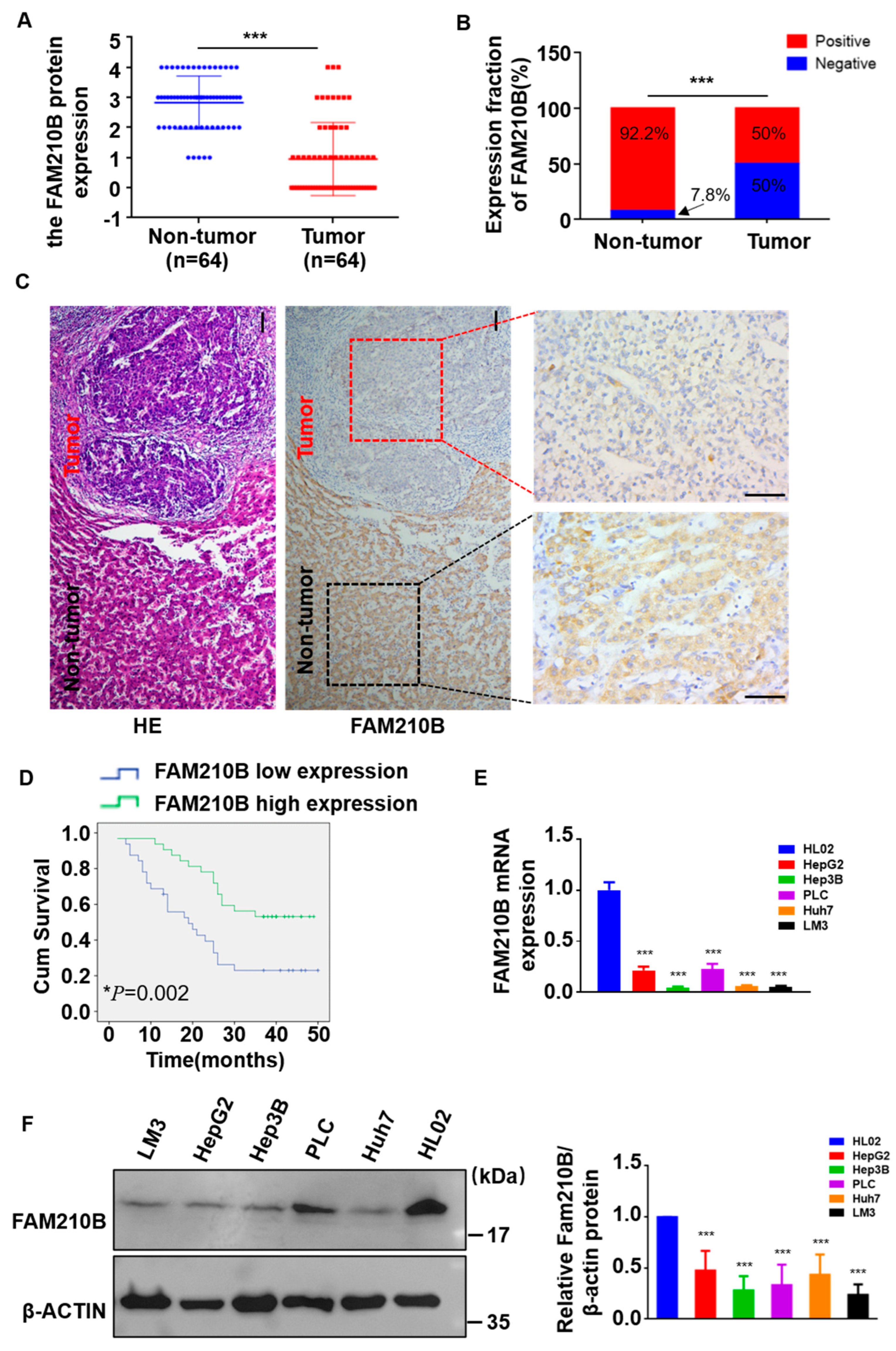
3.2. The Expression Level of FAM210B Was Reduced in Clinical Tissues of HCC
3.3. Low-Expression Levels of FAM210B Are Linked to Advancement of HCC and Poor Patient Prognosis
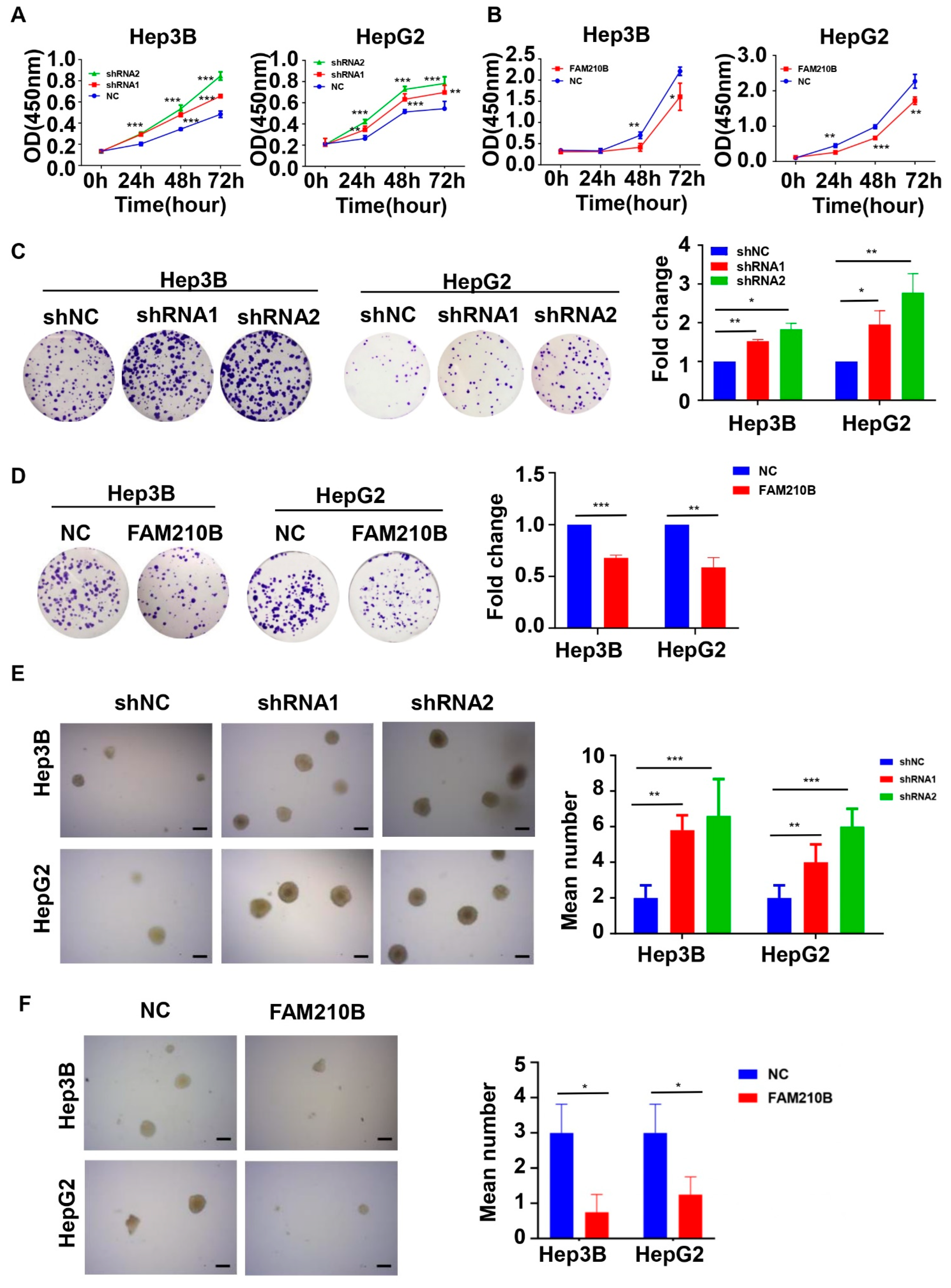
3.4. FAM210B Overexpression Inhibited HCC Cell Proliferation In Vitro
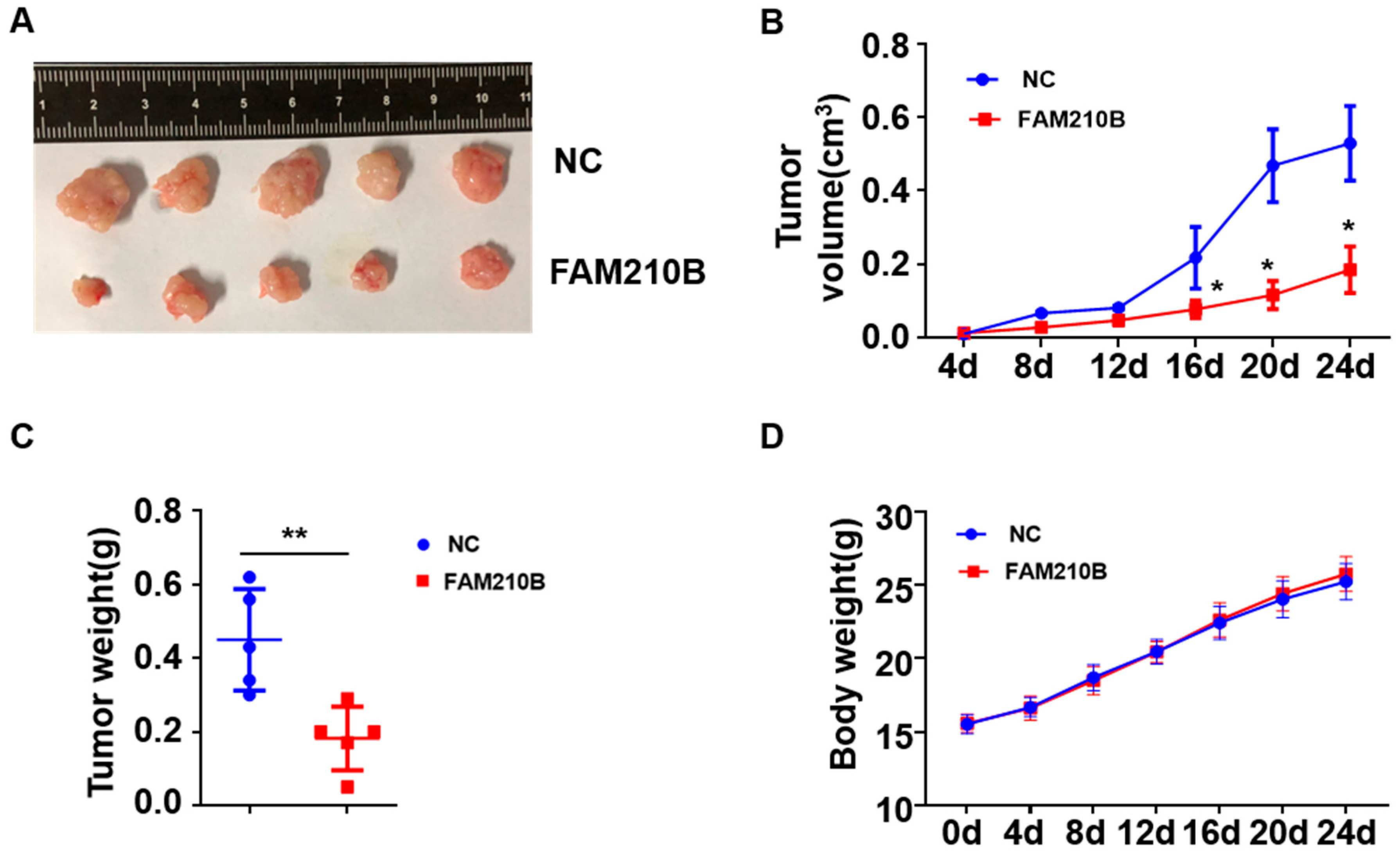
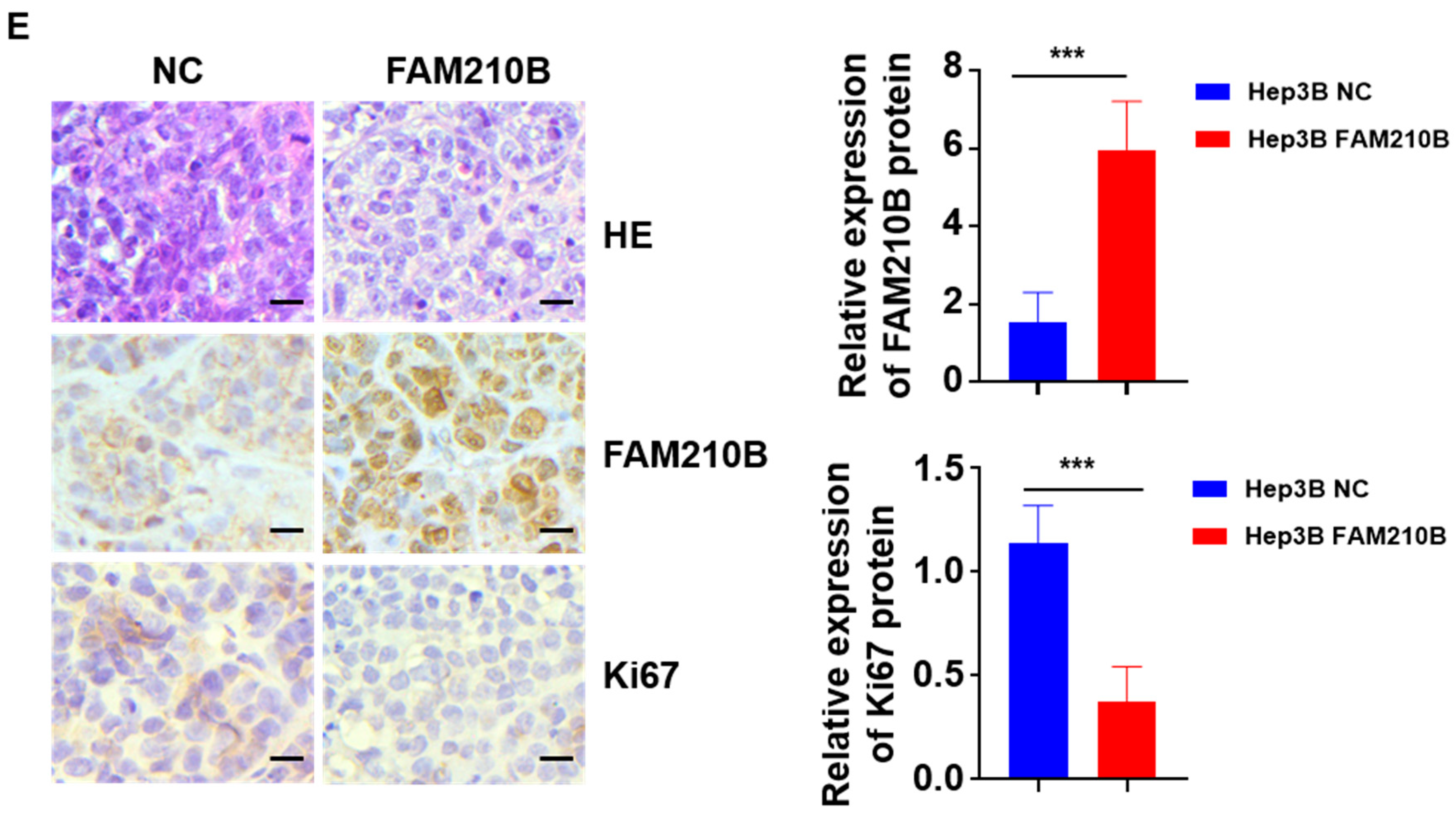
3.5. FAM210B Overexpression Inhibited Tumor Growth In Vivo
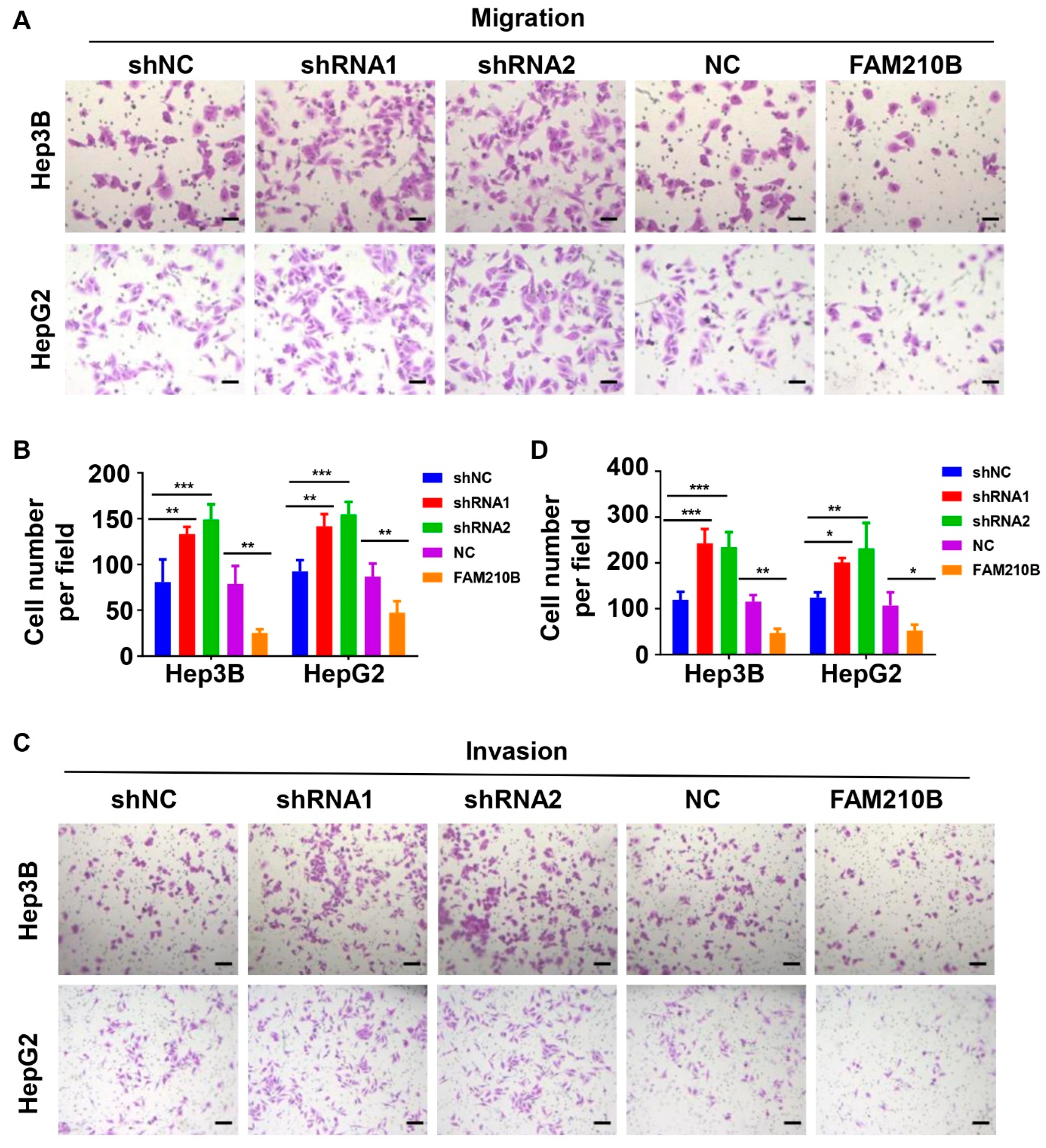
3.6. FAM210B Overexpression Inhibited the Migratory and Invasive Capacities of Hepatoma Cells
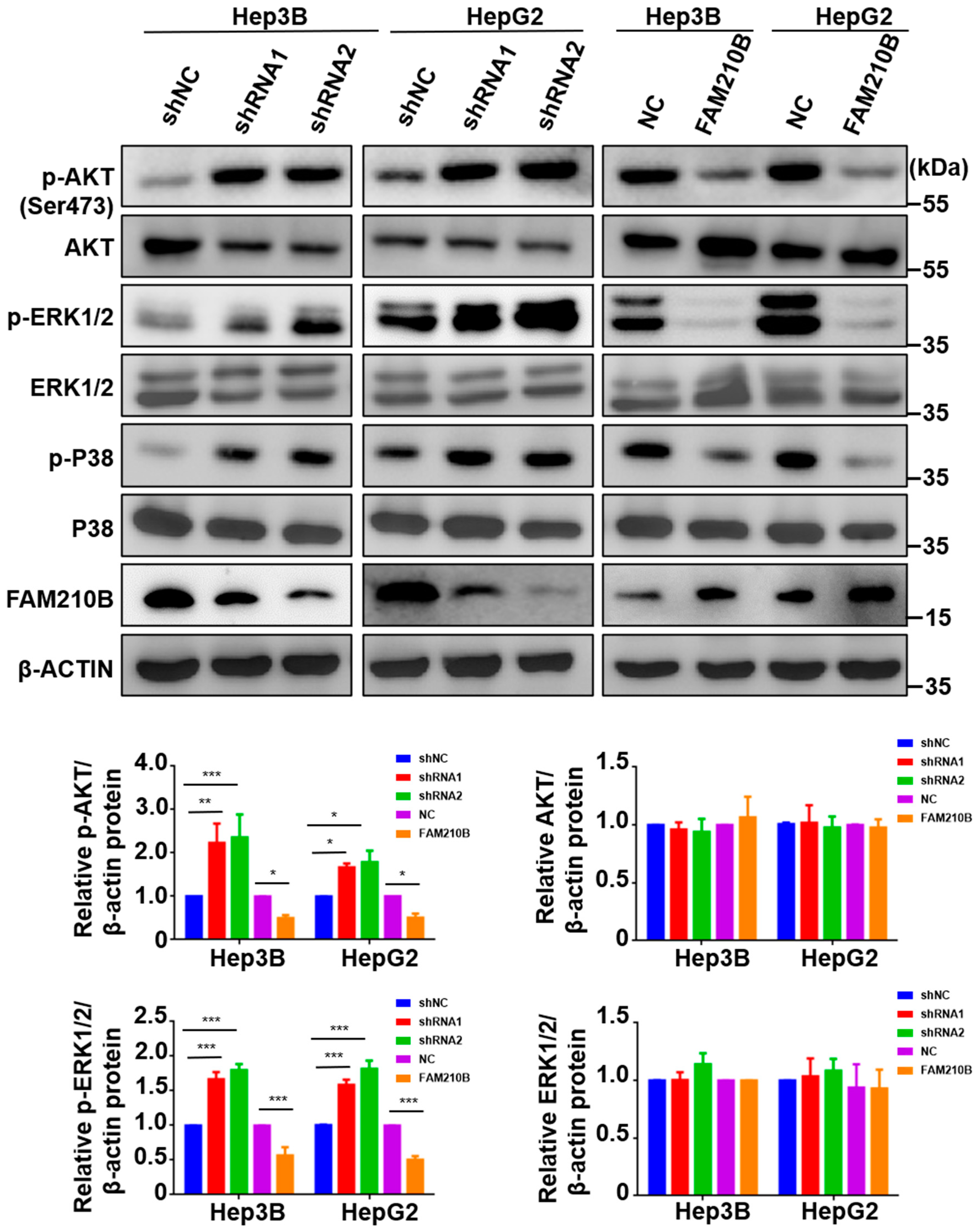
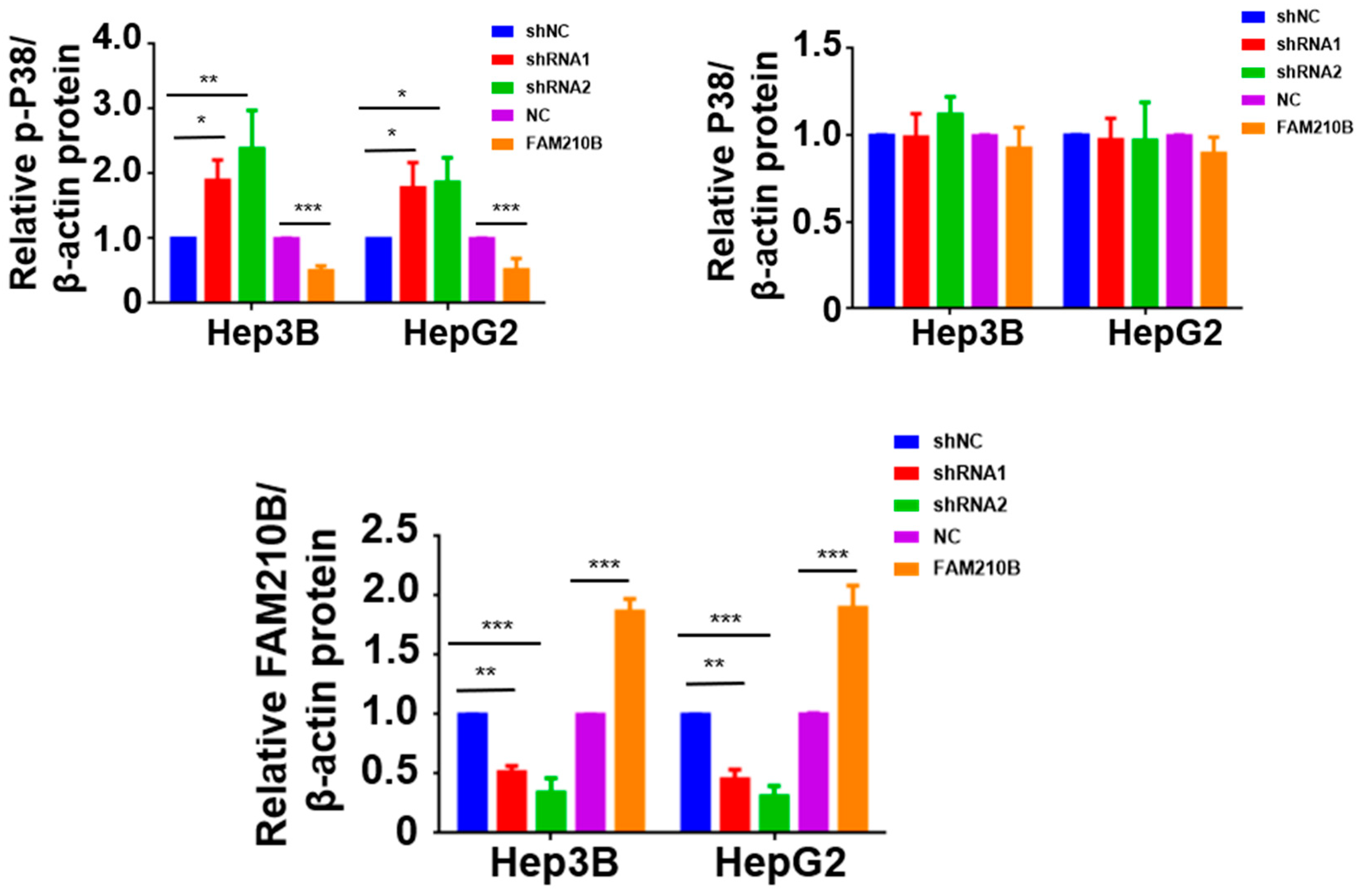
3.7. FAM210B Regulated ERK-AKT Signaling Pathways
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Llovet, J.M.; Kelley, R.K.; Villanueva, A.; Singal, A.G.; Pikarsky, E.; Roayaie, S. Hepatocellular carcinoma. Nat. Rev. Dis. Primers. 2021, 7, 6. [Google Scholar] [CrossRef] [PubMed]
- Rumgay, H.; Arnold, M.; Ferlay, J.; Lesi, O.; Cabasag, C.J.; Vignat, J.; Laversanne, M.; McGlynn, K.A.; Soerjomataram, I. Global burden of primary liver cancer in 2020 and predictions to 2040. J. Hepatol. 2022, 77, 1598–1606. [Google Scholar] [CrossRef] [PubMed]
- Altayar, O.; Shah, R.; Chang, C.Y.; Falck-Ytter, Y.; Muir, A.J. AGA Technical Review on Systemic Therapies for Hepatocellular Carcinoma. Gastroenterology 2022, 162, 937–951. [Google Scholar] [CrossRef]
- Cheng, A.L. Pursuing efficacious systemic therapy for hepatocellular carcinoma. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 95–96. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.D.; Heimbach, J.K. New advances in the diagnosis and management of hepatocellular carcinoma. BMJ 2020, 371, m3544. [Google Scholar] [CrossRef]
- Kanwal, F.; Khaderi, S.; Singal, A.G.; Marrero, J.A.; Loo, N.; Asrani, S.K.; Amos, C.I.; Thrift, A.P.; Gu, X.; Luster, M.; et al. Risk factors for HCC in contemporary cohorts of patients with cirrhosis. Hepatology 2023, 77, 997–1005. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, B. Hepatocellular carcinoma: Molecular mechanism, targeted therapy, and biomarkers. Cancer Metastasis Rev. 2023. [Google Scholar] [CrossRef]
- Yang, Y.; Ouyang, J.; Zhou, Y.; Zhou, J.; Zhao, H. The CRAFITY score: A promising prognostic predictor for patients with hepatocellular carcinoma treated with tyrosine kinase inhibitor and immunotherapy combinations. J. Hepatol. 2022, 77, 574–576. [Google Scholar] [CrossRef]
- Johnson, P.; Zhou, Q.; Dao, D.Y.; Lo, Y. Circulating biomarkers in the diagnosis and management of hepatocellular carcinoma. Nat. Rev. Gastroenterol. Hepatol. 2022, 19, 670–681. [Google Scholar] [CrossRef]
- Santarsiero, A.; Pappalardo, I.; Rosa, G.M.; Pisano, I.; Superchi, S.; Convertini, P.; Todisco, S.; Scafato, P.; Infantino, V. Mitochondrial Role in Intrinsic Apoptosis Induced by a New Synthesized Chalcone in Hepatocellular Carcinoma Cells. Biomedicines 2022, 10, 3120. [Google Scholar] [CrossRef]
- Weber, F.; Utpatel, K.; Evert, K.; Treeck, O.; Buechler, C. Chemerin and Chemokine-like Receptor 1 Expression Are Associated with Hepatocellular Carcinoma Progression in European Patients. Biomedicines 2023, 11, 737. [Google Scholar] [CrossRef] [PubMed]
- Kondo, A.; Fujiwara, T.; Okitsu, Y.; Fukuhara, N.; Onishi, Y.; Nakamura, Y.; Sawada, K.; Harigae, H. Identification of a novel putative mitochondrial protein FAM210B associated with erythroid differentiation. Int. J. Hematol. 2016, 103, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Yien, Y.Y.; Shi, J.; Chen, C.; Cheung, J.T.; Grillo, A.S.; Shrestha, R.; Li, L.; Zhang, X.; Kafina, M.D.; Kingsley, P.D.; et al. FAM210B is an erythropoietin target and regulates erythroid heme synthesis by controlling mitochondrial iron import and ferrochelatase activity. J. Biol. Chem. 2018, 293, 19797–19811. [Google Scholar] [CrossRef]
- Sun, S.; Liu, J.; Zhao, M.; Han, Y.; Chen, P.; Mo, Q.; Wang, B.; Chen, G.; Fang, Y.; Tian, Y.; et al. Loss of the novel mitochondrial protein FAM210B promotes metastasis via PDK4-dependent metabolic reprogramming. Cell Death Dis. 2017, 8, e2870. [Google Scholar] [CrossRef]
- He, G.; Fu, S.; Li, Y.; Li, T.; Mei, P.; Feng, L.; Cai, L.; Cheng, Y.; Zhou, C.; Tang, Y.; et al. TCGA and ESTIMATE data mining to identify potential prognostic biomarkers in HCC patients. Aging 2020, 12, 21544–21558. [Google Scholar] [CrossRef]
- Villa, E.; Critelli, R.; Lei, B.; Marzocchi, G.; Cammà, C.; Giannelli, G.; Pontisso, P.; Cabibbo, G.; Enea, M.; Colopi, S.; et al. Neoangiogenesis-related genes are hallmarks of fast-growing hepatocellular carcinomas and worst survival. Results from a prospective study. Gut 2016, 65, 861–869. [Google Scholar] [CrossRef]
- Hao, K.; Luk, J.M.; Lee, N.P.Y.; Mao, M.; Zhang, C.; Ferguson, M.D.; Lamb, J.; Dai, H.; Ng, I.O.; Sham, P.C.; et al. Predicting prognosis in hepatocellular carcinoma after curative surgery with common clinicopathologic parameters. BMC Cancer 2009, 9, 389. [Google Scholar] [CrossRef] [PubMed]
- Weiss, F.; Lauffenburger, D.; Friedl, P. Towards targeting of shared mechanisms of cancer metastasis and therapy resistance. Nat. Rev. Cancer 2022, 22, 157–173. [Google Scholar] [CrossRef]
- Han, Q.W.; Shao, Q.H.; Wang, X.T.; Ma, K.L.; Chen, N.H.; Yuan, Y.H. CB2 receptor activation inhibits the phagocytic function of microglia through activating ERK/AKT-Nurr1 signal pathways. Acta Pharmacol. Sin. 2022, 43, 2253–2266. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Zhang, X.; Huang, G.; Cheng, L.; Lv, J.; Zheng, D.; Lin, S.; Wang, S.; Wu, Q.; Long, Y.; et al. Myeloid-derived suppressor cells promote lung cancer metastasis by CCL11 to activate ERK and AKT signaling and induce epithelial-mesenchymal transition in tumor cells. Oncogene 2021, 40, 1476–1489. [Google Scholar] [CrossRef]
- Deng, R.; Liu, S.; Shen, S.; Guo, H.; Sun, J. Circulating hepatitis B virus RNA: From biology to clinical applications. Hepatology 2022, 76, 1520–1530. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhao, M.; Zhao, L.; Geng, Y.; Li, G.; Chen, L.; Yu, J.; Yuan, H.; Zhang, H.; Yun, H.; et al. HBx-induced HSPA8 stimulates HBV replication and suppresses ferroptosis to support liver cancer progression. Cancer Res. 2023, 83, 1048–1061. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Zhang, W.; Ben, Y. Identification of Key Regulators of Hepatitis C Virus-Induced Hepatocellular Carcinoma by Integrating Whole-Genome and Transcriptome Sequencing Data. Front. Genet. 2021, 12, 741608. [Google Scholar] [CrossRef] [PubMed]
- Cui, F.; Blach, S.; Mingiedi, C.M.; Gonzalez, M.A.; Alaama, A.S.; Mozalevskis, A.; Séguy, N.; Rewari, B.B.; Chan, P.-L.; Le, L.-V.; et al. Global reporting of progress towards elimination of hepatitis B and hepatitis C. Lancet Gastroenterol. Hepatol. 2023, 8, 332–342. [Google Scholar] [CrossRef]
- Claeys, L.; Romano, C.; De Ruyck, K.; Wilson, H.; Fervers, B.; Korenjak, M.; Zavadil, J.; Gunter, M.J.; De Saeger, S.; De Boevre, M.; et al. Mycotoxin exposure and human cancer risk: A systematic review of epidemiological studies. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1449–1464. [Google Scholar] [CrossRef]
- He, Y.; Su, Y.; Duan, C.; Wang, S.; He, W.; Zhang, Y.; An, X.; He, M. Emerging role of aging in the progression of NAFLD to HCC. Ageing Res. Rev. 2023, 84, 101833. [Google Scholar] [CrossRef]
- Mak, L.; Hui, R.W.; Lee, C.; Mao, X.; Cheung, K.; Wong, D.K.; Lui, D.T.; Fung, J.; Yuen, M.; Seto, W. Glycemic burden and the risk of adverse hepatic outcomes in patients with chronic hepatitis B with type 2 diabetes. Hepatology 2023, 77, 606–618. [Google Scholar] [CrossRef]
- Marti-Aguado, D.; Clemente-Sanchez, A.; Bataller, R. Cigarette smoking and liver diseases. J. Hepatol. 2022, 77, 191–205. [Google Scholar] [CrossRef]
- Pea, A.; Jamieson, N.B.; Braconi, C. Biology and Clinical Application of Regulatory RNAs in Hepatocellular Carcinoma. Hepatology 2021, 73, 38–48. [Google Scholar] [CrossRef]
- Craig, A.J.; von Felden, J.; Garcia-Lezana, T.; Sarcognato, S.; Villanueva, A. Tumour evolution in hepatocellular carcinoma. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 139–152. [Google Scholar] [CrossRef]
- Nishida, N. Role of Oncogenic Pathways on the Cancer Immunosuppressive Microenvironment and Its Clinical Implications in Hepatocellular Carcinoma. Cancers 2021, 13, 3666. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Pan, J.; Liu, T.; Yin, W.; Miao, Q.; Zhao, Z.; Gao, Y.; Zheng, W.; Li, H.; Deng, R.; et al. Novel TCF21(high) pericyte subpopulation promotes colorectal cancer metastasis by remodelling perivascular matrix. Gut 2023, 72, 710–721. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Lv, P.; Ma, J.; Chen, N.; Guo, H.; Chen, Y.; Gan, X.; Wang, R.; Liu, X.; Fan, S.; et al. Antrodia cinnamomea exerts an anti-hepatoma effect by targeting PI3K/AKT-mediated cell cycle progression in vitro and in vivo. Acta Pharm. Sin. B 2022, 12, 890–906. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Liu, Y.; Liu, L.; Si, C.; Xu, Y.; Wu, X.; Wang, C.; Sun, Z.; Kang, Q. Exosomal circTUBGCP4 promotes vascular endothelial cell tipping and colorectal cancer metastasis by activating Akt signaling pathway. J. Exp. Clin. Cancer Res. 2023, 42, 46. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.; Ro, S.W. MAPK/ERK Signaling Pathway in Hepatocellular Carcinoma. Cancers 2021, 13, 3026. [Google Scholar] [CrossRef]
- Koga, Y.; Tsurumaki, H.; Aoki-Saito, H.; Sato, M.; Yatomi, M.; Takehara, K.; Hisada, T. Roles of Cyclic AMP Response Element Binding Activation in the ERK1/2 and p38 MAPK Signalling Pathway in Central Nervous System, Cardiovascular System, Osteoclast Differentiation and Mucin and Cytokine Production. Int. J. Mol. Sci. 2019, 20, 1346. [Google Scholar] [CrossRef]
- Chan, L.-K.; Ho, D.W.-H.; Kam, C.S.; Chiu, E.Y.-T.; Lo, I.L.-O.; Yau, D.T.-W.; Cheung, E.T.-Y.; Tang, C.-N.; Tang, V.W.-L.; Lee, T.K.-W.; et al. RSK2-inactivating mutations potentiate MAPK signaling and support cholesterol metabolism in hepatocellular carcinoma. J. Hepatol. 2021, 74, 360–371. [Google Scholar] [CrossRef] [PubMed]
- Sekar, D. KAP Letter to the Editor: H19 Promotes HCC Bone Metastasis by Reducing Osteoprotegerin Expression in a PPP1CA/p38MAPK-Dependent Manner and Sponging miR-200b-3p. Hepatology 2021, 74, 1713. [Google Scholar] [CrossRef]
- Jin, A.L.; Zhang, C.Y.; Zheng, W.J.; Xian, J.R.; Yang, W.J.; Liu, T.; Chen, W.; Li, T.; Wang, B.L.; Pan, B.S.; et al. CD155/SRC complex promotes hepatocellular carcinoma progression via inhibiting the p38 MAPK signalling pathway and correlates with poor prognosis. Clin. Transl. Med. 2022, 12, e794. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Features | Total Cases | FAM210B Levels | χ2 | ap Value | |
|---|---|---|---|---|---|
| Low | High | ||||
| Total number | 64 | 32 | 32 | ||
| Gender | 0.642 | 0.423 | |||
| Male | 57 | 27 (47.4%) | 30 (52.6%) | ||
| Female | 7 | 5 (71.4%) | 2 (28.6%) | ||
| Age | 0.567 | 0.451 | |||
| <60 | 35 | 19 (54.3%) | 16 (45.7%) | ||
| ≥60 | 29 | 13 (44.8%) | 16 (55.2%) | ||
| Tumor size (cm) | 2.400 | 0.121 | |||
| <5 | 40 | 17 (42.5%) | 23 (57.5%) | ||
| ≥5 | 24 | 15 (62.5%) | 9 (37.5%) | ||
| Histological grade | - a | 0.005 * | |||
| High | 9 | 3 (33.3%) | 6 (66.7%) | ||
| Middle | 24 | 7 (29.2%) | 17 (70.8%) | ||
| Low | 31 | 22 (71.0%) | 9 (29.0%) | ||
| Invasion depth | 0.066 | 0.798 | |||
| Tis-T1 | 25 | 12 (48.0%) | 13 (52.0%) | ||
| T2–T4 | 39 | 20 (51.3%) | 19 (48.7%) | ||
| Lymphatic metastasis | 4.655 | 0.031 * | |||
| with | 20 | 14 (70.0%) | 6 (30.0%) | ||
| without | 44 | 18 (40.9%) | 26 (59.1%) | ||
| TNM stage | 0.066 | 0.798 | |||
| I–II | 39 | 19 (48.7%) | 20 (51.3%) | ||
| III–IV | 25 | 13 (52.0%) | 12 (48.0%) | ||
| Survival status | 5.317 | 0.021 * | |||
| Live | 25 | 8 (32.0%) | 17 (68.0%) | ||
| Death | 39 | 24 (61.5%) | 15 (38.5%) | ||
| Variables | Univariate Analysis | Multivariate Analysis | ||
|---|---|---|---|---|
| HR (95%CI) | p Value | HR (95%CI) | p Value | |
| Gender | 2.057 (0.801–5.283) | 0.134 | - | - |
| Age | 0.763 (0.403–1.445) | 0.407 | - | - |
| Tumor Size (cm) | 2.700 (1.435–5.082) | 0.002 * | 2.622 (1.164–5.908) | 0.020 * |
| Invasion depth | 5.680 (2.484–12.985) | <0.001 * | 2.925 (1.033–8.283) | 0.043 * |
| Lymphatic metastasis | 15.873 (6.919–36.414) | <0.001 * | 11.097 (3.693–33.346) | <0.001 * |
| TNM stage | 6.727 (3.331–13.585) | <0.001 * | 4.572 (1.763–11.856) | 0.002 * |
| FAM210B expression | 0.378 (0.197–0.726) | 0.003 * | 0.300 (0.125–0.722) | 0.007 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, Y.; Pan, X.; Liu, Y.; Li, X.; Lin, K.; Zhu, J.; Zhan, L.; Kan, C.; Zheng, H. Loss of the Novel Mitochondrial Membrane Protein FAM210B Is Associated with Hepatocellular Carcinoma. Biomedicines 2023, 11, 1232. https://doi.org/10.3390/biomedicines11041232
Zhou Y, Pan X, Liu Y, Li X, Lin K, Zhu J, Zhan L, Kan C, Zheng H. Loss of the Novel Mitochondrial Membrane Protein FAM210B Is Associated with Hepatocellular Carcinoma. Biomedicines. 2023; 11(4):1232. https://doi.org/10.3390/biomedicines11041232
Chicago/Turabian StyleZhou, Yuanqin, Xianzhu Pan, Yakun Liu, Xiaofei Li, Keqiong Lin, Jicheng Zhu, Li Zhan, Chen Kan, and Hong Zheng. 2023. "Loss of the Novel Mitochondrial Membrane Protein FAM210B Is Associated with Hepatocellular Carcinoma" Biomedicines 11, no. 4: 1232. https://doi.org/10.3390/biomedicines11041232
APA StyleZhou, Y., Pan, X., Liu, Y., Li, X., Lin, K., Zhu, J., Zhan, L., Kan, C., & Zheng, H. (2023). Loss of the Novel Mitochondrial Membrane Protein FAM210B Is Associated with Hepatocellular Carcinoma. Biomedicines, 11(4), 1232. https://doi.org/10.3390/biomedicines11041232