
Periodic and Aperiodic EEG Features as Potential Markers of Developmental Dyslexia

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Reading Tasks
2.3. EEG Data Analyses
2.4. Statistical Analyses
3. Results
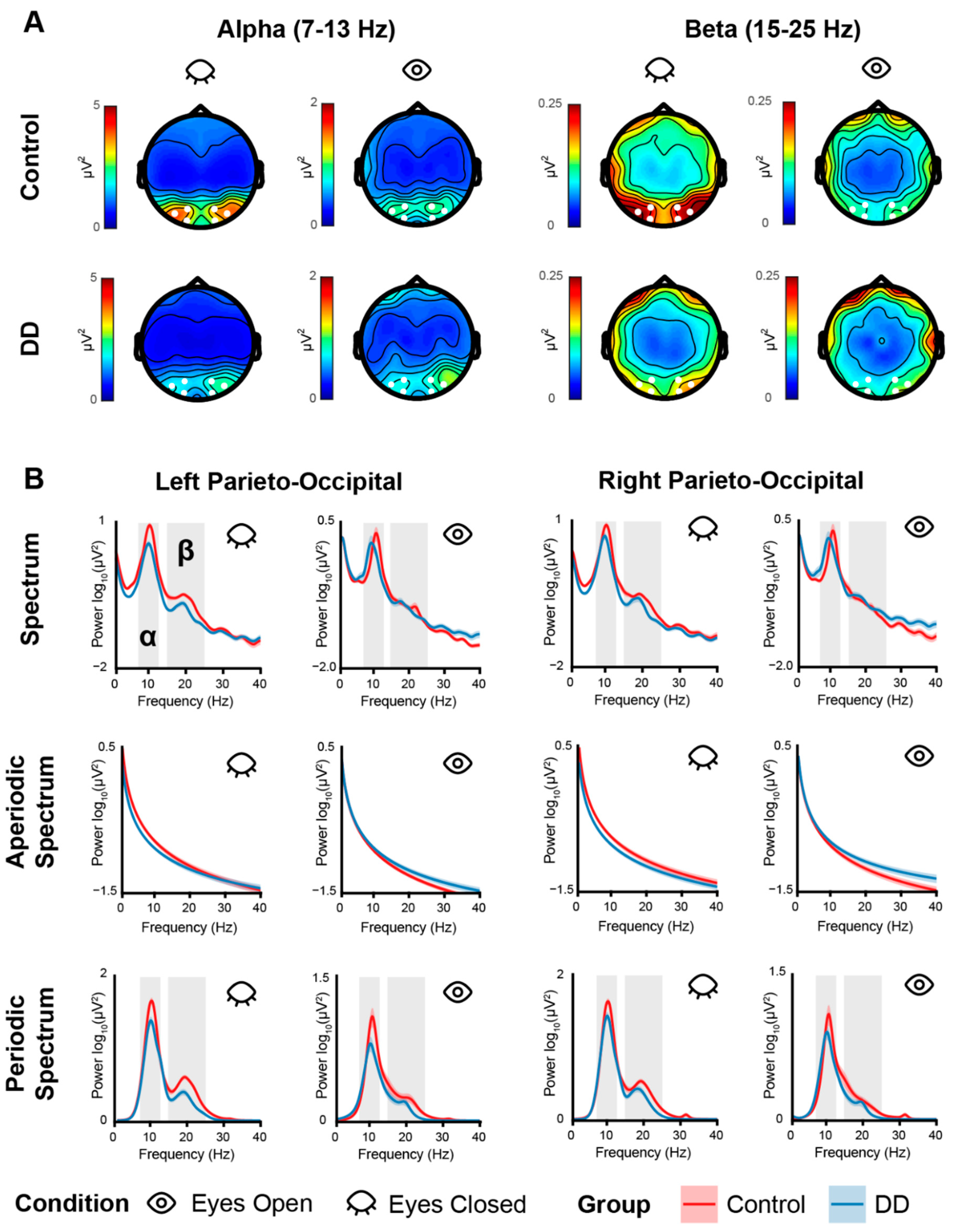
3.1. EEG Results—Aperiodic Activity
3.2. EEG Results—Periodic Activity
3.3. Linear Models and Correlation Results
3.4. Results Overview

4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association: Washington, DC, USA, 2013; ISBN 978-0-89042-555-8. [Google Scholar]
- Di Gregorio, F.; La Porta, F.; Petrone, V.; Battaglia, S.; Orlandi, S.; Ippolito, G.; Romei, V.; Piperno, R.; Lullini, G. Accuracy of EEG Biomarkers in the Detection of Clinical Outcome in Disorders of Consciousness after Severe Acquired Brain Injury: Preliminary Results of a Pilot Study Using a Machine Learning Approach. Biomedicines 2022, 10, 1897. [Google Scholar] [CrossRef] [PubMed]
- Liloia, D.; Crocetta, A.; Cauda, F.; Duca, S.; Costa, T.; Manuello, J. Seeking Overlapping Neuroanatomical Alterations between Dyslexia and Attention-Deficit/Hyperactivity Disorder: A Meta-Analytic Replication Study. Brain Sci. 2022, 12, 1367. [Google Scholar] [CrossRef] [PubMed]
- Pani, S.M.; Saba, L.; Fraschini, M. Clinical Applications of EEG Power Spectra Aperiodic Component Analysis: A Mini-Review. Clin. Neurophysiol. 2022, 143, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Karamacoska, D.; Barry, R.J.; Steiner, G.Z.; Coleman, E.P.; Wilson, E.J. Intrinsic EEG and Task-Related Changes in EEG Affect Go/NoGo Task Performance. Int. J. Psychophysiol. 2018, 125, 17–28. [Google Scholar] [CrossRef]
- Mahjoory, K.; Cesnaite, E.; Hohlefeld, F.U.; Villringer, A.; Nikulin, V.V. Power and Temporal Dynamics of Alpha Oscillations at Rest Differentiate Cognitive Performance Involving Sustained and Phasic Cognitive Control. NeuroImage 2019, 188, 135–144. [Google Scholar] [CrossRef]
- Rogala, J.; Kublik, E.; Krauz, R.; Wróbel, A. Resting-State EEG Activity Predicts Frontoparietal Network Reconfiguration and Improved Attentional Performance. Sci. Rep. 2020, 10, 5064. [Google Scholar] [CrossRef]
- Cainelli, E.; Vedovelli, L.; Carretti, B.; Bisiacchi, P. EEG Correlates of Developmental Dyslexia: A Systematic Review. Ann. Dyslexia 2022. [Google Scholar] [CrossRef]
- Klimesch, W.; Doppelmayr, M.; Wimmer, H.; Schwaiger, J.; Röhm, D.; Gruber, W.; Hutzler, F. Theta Band Power Changes in Normal and Dyslexic Children. Clin. Neurophysiol. 2001, 112, 1174–1185. [Google Scholar] [CrossRef]
- Rippon, G. Trait and State EEG Indices of Information Processing in Developmental Dyslexia. Int. J. Psychophysiol. 2000, 36, 251–265. [Google Scholar] [CrossRef]
- Spironelli, C.; Penolazzi, B.; Angrilli, A. Dysfunctional Hemispheric Asymmetry of Theta and Beta EEG Activity during Linguistic Tasks in Developmental Dyslexia. Biol. Psychol. 2008, 77, 123–131. [Google Scholar] [CrossRef]
- Klimesch, W.; Doppelmayr, M.; Wimmer, H.; Gruber, W.; Röhm, D.; Schwaiger, J.; Hutzler, F. Alpha and Beta Band Power Changes in Normal and Dyslexic Children. Clin. Neurophysiol. 2001, 112, 1186–1195. [Google Scholar] [CrossRef]
- Papagiannopoulou, E.A.; Lagopoulos, J. Resting State EEG Hemispheric Power Asymmetry in Children with Dyslexia. Front. Pediatr. 2016, 4, 11. [Google Scholar] [CrossRef]
- Arns, M.; Peters, S.; Breteler, R.; Verhoeven, L. Different Brain Activation Patterns in Dyslexic Children: Evidence from EEG Power and Coherence Patterns for the Double-Deficit Theory of Dyslexia. J. Integr. Neurosci. 2007, 6, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Babiloni, C.; Stella, G.; Buffo, P.; Vecchio, F.; Onorati, P.; Muratori, C.; Miano, S.; Gheller, F.; Antonaci, L.; Albertini, G.; et al. Cortical Sources of Resting State EEG Rhythms Are Abnormal in Dyslexic Children. Clin. Neurophysiol. 2012, 123, 2384–2391. [Google Scholar] [CrossRef]
- Fraga González, G.; Van der Molen, M.J.W.; Žarić, G.; Bonte, M.; Tijms, J.; Blomert, L.; Stam, C.J.; Van der Molen, M.W. Graph Analysis of EEG Resting State Functional Networks in Dyslexic Readers. Clin. Neurophysiol. 2016, 127, 3165–3175. [Google Scholar] [CrossRef] [PubMed]
- Rumsey, J.M.; Coppola, R.; Denckla, M.B.; Hamburger, S.D.; Kruesi, M.J.P. EEG Spectra in Severely Dyslexic Men: Rest and Word and Design Recognition. Electroencephalogr. Clin. Neurophysiol. 1989, 73, 30–40. [Google Scholar] [CrossRef] [PubMed]
- Xue, H.; Wang, Z.; Tan, Y.; Yang, H.; Fu, W.; Xue, L.; Zhao, J. Resting-State EEG Reveals Global Network Deficiency in Dyslexic Children. Neuropsychologia 2020, 138, 107343. [Google Scholar] [CrossRef]
- Ghiani, A.; Maniglia, M.; Battaglini, L.; Melcher, D.; Ronconi, L. Binding Mechanisms in Visual Perception and Their Link With Neural Oscillations: A Review of Evidence From TACS. Front. Psychol. 2021, 12, 643677. [Google Scholar] [CrossRef]
- Klimesch, W. EEG-Alpha Rhythms and Memory Processes. Int. J. Psychophysiol. 1997, 26, 319–340. [Google Scholar] [CrossRef]
- Klimesch, W.; Schmike, H.; Ladurner, G.; Pfurtscheller, G. Alpha Frequency and Memory Performance. J. Psychophysiol. 1990, 4, 381–390. [Google Scholar]
- Suldo, S.M.; Olson, L.A.; Evans, J.R. Quantitative EEG Evidence of Increased Alpha Peak Frequency in Children with Precocious Reading Ability. J. Neurother. 2002, 5, 39–50. [Google Scholar] [CrossRef]
- Angelakis, E.; Lubar, J.F.; Stathopoulou, S. Electroencephalographic Peak Alpha Frequency Correlates of Cognitive Traits. Neurosci. Lett. 2004, 371, 60–63. [Google Scholar] [CrossRef] [PubMed]
- Grandy, T.H.; Werkle-Bergner, M.; Chicherio, C.; Lövdén, M.; Schmiedek, F.; Lindenberger, U. Individual Alpha Peak Frequency Is Related to Latent Factors of General Cognitive Abilities. NeuroImage 2013, 79, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, I.S.; Lynn, P.A.; Schermitzler, B.; Sponheim, S.R. Individual Alpha Peak Frequency Is Slower in Schizophrenia and Related to Deficits in Visual Perception and Cognition. Sci. Rep. 2021, 11, 17852. [Google Scholar] [CrossRef]
- Dickinson, A.; DiStefano, C.; Senturk, D.; Jeste, S.S. Peak Alpha Frequency Is a Neural Marker of Cognitive Function across the Autism Spectrum. Eur. J. Neurosci. 2018, 47, 643–651. [Google Scholar] [CrossRef] [PubMed]
- Edgar, J.C.; Dipiero, M.; McBride, E.; Green, H.L.; Berman, J.; Ku, M.; Liu, S.; Blaskey, L.; Kuschner, E.; Airey, M.; et al. Abnormal Maturation of the Resting-state Peak Alpha Frequency in Children with Autism Spectrum Disorder. Hum. Brain Mapp. 2019, 40, 3288–3298. [Google Scholar] [CrossRef]
- Donoghue, T.; Haller, M.; Peterson, E.J.; Varma, P.; Sebastian, P.; Gao, R.; Noto, T.; Lara, A.H.; Wallis, J.D.; Knight, R.T.; et al. Parameterizing Neural Power Spectra into Periodic and Aperiodic Components. Nat. Neurosci. 2020, 23, 1655–1665. [Google Scholar] [CrossRef]
- He, B.J. Scale-Free Brain Activity: Past, Present, and Future. Trends Cogn. Sci. 2014, 18, 480–487. [Google Scholar] [CrossRef]
- Donoghue, T.; Schaworonkow, N.; Voytek, B. Methodological Considerations for Studying Neural Oscillations. Eur. J. Neurosci. 2022, 55, 3502–3527. [Google Scholar] [CrossRef]
- Ouyang, G.; Hildebrandt, A.; Schmitz, F.; Herrmann, C.S. Decomposing Alpha and 1/f Brain Activities Reveals Their Differential Associations with Cognitive Processing Speed. NeuroImage 2020, 205, 116304. [Google Scholar] [CrossRef]
- Groppe, D.M.; Bickel, S.; Keller, C.J.; Jain, S.K.; Hwang, S.T.; Harden, C.; Mehta, A.D. Dominant Frequencies of Resting Human Brain Activity as Measured by the Electrocorticogram. NeuroImage 2013, 79, 223–233. [Google Scholar] [CrossRef] [PubMed]
- Waschke, L.; Donoghue, T.; Fiedler, L.; Smith, S.; Garrett, D.D.; Voytek, B.; Obleser, J. Modality-Specific Tracking of Attention and Sensory Statistics in the Human Electrophysiological Spectral Exponent. eLife 2021, 10, e70068. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Peterson, E.J.; Voytek, B. Inferring Synaptic Excitation/Inhibition Balance from Field Potentials. NeuroImage 2017, 158, 70–78. [Google Scholar] [CrossRef]
- Voytek, B.; Knight, R.T. Dynamic Network Communication as a Unifying Neural Basis for Cognition, Development, Aging, and Disease. Biol. Psychiatry 2015, 77, 1089–1097. [Google Scholar] [CrossRef]
- Manning, J.R.; Jacobs, J.; Fried, I.; Kahana, M.J. Broadband Shifts in Local Field Potential Power Spectra Are Correlated with Single-Neuron Spiking in Humans. J. Neurosci. 2009, 29, 13613–13620. [Google Scholar] [CrossRef]
- Schaworonkow, N.; Voytek, B. Longitudinal Changes in Aperiodic and Periodic Activity in Electrophysiological Recordings in the First Seven Months of Life. Dev. Cogn. Neurosci. 2021, 47, 100895. [Google Scholar] [CrossRef]
- Cellier, D.; Riddle, J.; Petersen, I.; Hwang, K. The Development of Theta and Alpha Neural Oscillations from Ages 3 to 24 Years. Dev. Cogn. Neurosci. 2021, 50, 100969. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.T.; Clark, G.M.; Bigelow, F.J.; Lum, J.A.G.; Enticott, P.G. Periodic and Aperiodic Neural Activity Displays Age-Dependent Changes across Early-to-Middle Childhood. Dev. Cogn. Neurosci. 2022, 54, 101076. [Google Scholar] [CrossRef]
- McSweeney, M.; Morales, S.; Valadez, E.A.; Buzzell, G.A.; Fox, N.A. Longitudinal Age- and Sex-Related Change in Background Aperiodic Activity during Early Adolescence. Dev. Cogn. Neurosci. 2021, 52, 101035. [Google Scholar] [CrossRef]
- Merkin, A.; Sghirripa, S.; Graetz, L.; Smith, A.E.; Hordacre, B.; Harris, R.; Pitcher, J.; Semmler, J.; Rogasch, N.C.; Goldsworthy, M. Do Age-Related Differences in Aperiodic Neural Activity Explain Differences in Resting EEG Alpha? Neurobiol. Aging 2023, 121, 78–87. [Google Scholar] [CrossRef]
- Voytek, B.; Kramer, M.A.; Case, J.; Lepage, K.Q.; Tempesta, Z.R.; Knight, R.T.; Gazzaley, A. Age-Related Changes in 1/f Neural Electrophysiological Noise. J. Neurosci. 2015, 35, 13257–13265. [Google Scholar] [CrossRef] [PubMed]
- Colombo, M.A.; Napolitani, M.; Boly, M.; Gosseries, O.; Casarotto, S.; Rosanova, M.; Brichant, J.-F.; Boveroux, P.; Rex, S.; Laureys, S.; et al. The Spectral Exponent of the Resting EEG Indexes the Presence of Consciousness during Unresponsiveness Induced by Propofol, Xenon, and Ketamine. NeuroImage 2019, 189, 631–644. [Google Scholar] [CrossRef] [PubMed]
- Thuwal, K.; Banerjee, A.; Roy, D. Aperiodic and Periodic Components of Ongoing Oscillatory Brain Dynamics Link Distinct Functional Aspects of Cognition across Adult Lifespan. Eneuro 2021, 8, ENEURO.0224-21.2021. [Google Scholar] [CrossRef]
- Demuru, M.; Fraschini, M. EEG Fingerprinting: Subject-Specific Signature Based on the Aperiodic Component of Power Spectrum. Comput. Biol. Med. 2020, 120, 103748. [Google Scholar] [CrossRef]
- Robertson, M.M.; Furlong, S.; Voytek, B.; Donoghue, T.; Boettiger, C.A.; Sheridan, M.A. EEG Power Spectral Slope Differs by ADHD Status and Stimulant Medication Exposure in Early Childhood. J. Neurophysiol. 2019, 122, 2427–2437. [Google Scholar] [CrossRef] [PubMed]
- Manyukhina, V.O.; Prokofyev, A.O.; Galuta, I.A.; Goiaeva, D.E.; Obukhova, T.S.; Schneiderman, J.F.; Altukhov, D.I.; Stroganova, T.A.; Orekhova, E.V. Globally Elevated Excitation–Inhibition Ratio in Children with Autism Spectrum Disorder and below-Average Intelligence. Mol. Autism 2022, 13, 20. [Google Scholar] [CrossRef] [PubMed]
- Molina, J.L.; Voytek, B.; Thomas, M.L.; Joshi, Y.B.; Bhakta, S.G.; Talledo, J.A.; Swerdlow, N.R.; Light, G.A. Memantine Effects on Electroencephalographic Measures of Putative Excitatory/Inhibitory Balance in Schizophrenia. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2020, 5, 562–568. [Google Scholar] [CrossRef]
- Hancock, R.; Pugh, K.R.; Hoeft, F. Neural Noise Hypothesis of Developmental Dyslexia. Trends Cogn. Sci. 2017, 21, 434–448. [Google Scholar] [CrossRef]
- Barry, R.J.; De Blasio, F.M. EEG Differences between Eyes-Closed and Eyes-Open Resting Remain in Healthy Ageing. Biol. Psychol. 2017, 129, 293–304. [Google Scholar] [CrossRef]
- Barry, R.J.; Clarke, A.R.; Johnstone, S.J.; Brown, C.R. EEG Differences in Children between Eyes-Closed and Eyes-Open Resting Conditions. Clin. Neurophysiol. 2009, 120, 1806–1811. [Google Scholar] [CrossRef]
- Barry, R.J.; Clarke, A.R.; Johnstone, S.J.; Magee, C.A.; Rushby, J.A. EEG Differences between Eyes-Closed and Eyes-Open Resting Conditions. Clin. Neurophysiol. 2007, 118, 2765–2773. [Google Scholar] [CrossRef]
- Arthur, W.; Day, D.V. Development of a Short Form for the Raven Advanced Progressive Matrices Test. Educ. Psychol. Meas. 1994, 54, 394–403. [Google Scholar] [CrossRef]
- Ronconi, L.; Melcher, D.; Franchin, L. Investigating the Role of Temporal Processing in Developmental Dyslexia: Evidence for a Specific Deficit in Rapid Visual Segmentation. Psychon. Bull. Rev. 2020, 27, 724–734. [Google Scholar] [CrossRef] [PubMed]
- Judica, A.; De Luca, M. Prove Di Velocità Di Lettura Brani per La Scuola Media Superiore [Tests of Text-Reading Speed for High School]. Available online: https://www.hsantalucia.it/sites/default/files/fsl_labdislessia_lettura_brani_test.pdf (accessed on 1 November 2020).
- Sartori, G.; Job, R.; Tressoldi, P. Batteria per La Valutazione Della Dislessia e Della Disortografia Evolutiva [Battery for the Assessment of Dyslexia and Developmental Dysorthography]; Giunti, O.S.: Florence, Italy, 1995. [Google Scholar]
- Delorme, A.; Makeig, S. EEGLAB: An Open Source Toolbox for Analysis of Single-Trial EEG Dynamics Including Independent Component Analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef]
- Oostenveld, R.; Fries, P.; Maris, E.; Schoffelen, J.-M. FieldTrip: Open Source Software for Advanced Analysis of MEG, EEG, and Invasive Electrophysiological Data. Comput. Intell. Neurosci. 2011, 2011, 156869. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Ripley, B.; Venables, B.; Bates, D.M.; Hornik, K.; Gebhardt, A.; Firth, D.; Ripley, M.B. Package ‘Mass’. Cran R 2013, 538, 113–120. [Google Scholar]
- Lenth, R.; Singmann, H.; Love, J.; Buerkner, P.; Herve, M. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 2018, 1, 3. [Google Scholar]
- Sperling, A.J.; Lu, Z.-L.; Manis, F.R.; Seidenberg, M.S. Motion-Perception Deficits and Reading Impairment: It’s the Noise, Not the Motion. Psychol. Sci. 2006, 17, 1047–1053. [Google Scholar] [CrossRef]
- Miller, K.J.; Honey, C.J.; Hermes, D.; Rao, R.P.; Dennijs, M.; Ojemann, J.G. Broadband Changes in the Cortical Surface Potential Track Activation of Functionally Diverse Neuronal Populations. NeuroImage 2014, 85, 711–720. [Google Scholar] [CrossRef]
- Bertone, A.; Mottron, L.; Jelenic, P.; Faubert, J. Enhanced and Diminished Visuo-Spatial Information Processing in Autism Depends on Stimulus Complexity. Brain 2005, 128, 2430–2441. [Google Scholar] [CrossRef] [PubMed]
- Senkowski, D.; Schneider, T.R.; Foxe, J.J.; Engel, A.K. Crossmodal Binding through Neural Coherence: Implications for Multisensory Processing. Trends Neurosci. 2008, 31, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Ding, N.; Simon, J.Z. Cortical Entrainment to Continuous Speech: Functional Roles and Interpretations. Front. Hum. Neurosci. 2014, 8, 311. [Google Scholar] [CrossRef] [PubMed]
- Gross, J.; Hoogenboom, N.; Thut, G.; Schyns, P.; Panzeri, S.; Belin, P.; Garrod, S. Speech Rhythms and Multiplexed Oscillatory Sensory Coding in the Human Brain. PLoS Biol. 2013, 11, e1001752. [Google Scholar] [CrossRef]
- Melby-Lervåg, M.; Lyster, S.-A.H.; Hulme, C. Phonological Skills and Their Role in Learning to Read: A Meta-Analytic Review. Psychol. Bull. 2012, 138, 322–352. [Google Scholar] [CrossRef]
- Ziegler, J.C.; Pech-Georgel, C.; George, F.; Lorenzi, C. Speech-Perception-in-Noise Deficits in Dyslexia. Dev. Sci. 2009, 12, 732–745. [Google Scholar] [CrossRef] [PubMed]
- Gori, S.; Facoetti, A. How the Visual Aspects Can Be Crucial in Reading Acquisition: The Intriguing Case of Crowding and Developmental Dyslexia. J. Vis. 2015, 15, 8. [Google Scholar] [CrossRef]
- Capilla, A.; Arana, L.; García-Huéscar, M.; Melcón, M.; Gross, J.; Campo, P. The Natural Frequencies of the Resting Human Brain: An MEG-Based Atlas. NeuroImage 2022, 258, 119373. [Google Scholar] [CrossRef]
- Samaha, J.; Gosseries, O.; Postle, B.R. Distinct Oscillatory Frequencies Underlie Excitability of Human Occipital and Parietal Cortex. J. Neurosci. 2017, 37, 2824–2833. [Google Scholar] [CrossRef]
- Stengel, C.; Vernet, M.; Amengual, J.L.; Valero-Cabré, A. Causal Modulation of Right Hemisphere Fronto-Parietal Phase Synchrony with Transcranial Magnetic Stimulation during a Conscious Visual Detection Task. Sci. Rep. 2021, 11, 3807. [Google Scholar] [CrossRef]
- Capotosto, P.; Babiloni, C.; Romani, G.L.; Corbetta, M. Frontoparietal Cortex Controls Spatial Attention through Modulation of Anticipatory Alpha Rhythms. J. Neurosci. 2009, 29, 5863–5872. [Google Scholar] [CrossRef] [PubMed]
- Veniero, D.; Gross, J.; Morand, S.; Duecker, F.; Sack, A.T.; Thut, G. Top-down Control of Visual Cortex by the Frontal Eye Fields through Oscillatory Realignment. Nat. Commun. 2021, 12, 1757. [Google Scholar] [CrossRef] [PubMed]
- Yordanova, J.; Kolev, V.; Verleger, R.; Heide, W.; Grumbt, M.; Schürmann, M. Synchronization of Fronto-Parietal Beta and Theta Networks as a Signature of Visual Awareness in Neglect. NeuroImage 2017, 146, 341–354. [Google Scholar] [CrossRef] [PubMed]
- Battaglini, L.; Ghiani, A.; Casco, C.; Ronconi, L. Parietal TACS at Beta Frequency Improves Vision in a Crowding Regime. NeuroImage 2020, 208, 116451. [Google Scholar] [CrossRef] [PubMed]
- Ronconi, L.; Bertoni, S.; Bellacosa Marotti, R. The Neural Origins of Visual Crowding as Revealed by Event-Related Potentials and Oscillatory Dynamics. Cortex 2016, 79, 87–98. [Google Scholar] [CrossRef]
- Ronconi, L.; Bellacosa Marotti, R. Awareness in the Crowd: Beta Power and Alpha Phase of Prestimulus Oscillations Predict Object Discrimination in Visual Crowding. Conscious. Cogn. 2017, 54, 36–46. [Google Scholar] [CrossRef]
- Bertoni, S.; Franceschini, S.; Ronconi, L.; Gori, S.; Facoetti, A. Is Excessive Visual Crowding Causally Linked to Developmental Dyslexia? Neuropsychologia 2019, 130, 107–117. [Google Scholar] [CrossRef]
- Martelli, M.; Di Filippo, G.; Spinelli, D.; Zoccolotti, P. Crowding, Reading, and Developmental Dyslexia. J. Vis. 2009, 9, 14. [Google Scholar] [CrossRef]
- Atilgan, N.; Yu, S.M.; He, S. Visual Crowding Effect in the Parvocellular and Magnocellular Visual Pathways. J. Vis. 2020, 20, 6. [Google Scholar] [CrossRef]
- Livingstone, M.S.; Rosen, G.D.; Drislane, F.W.; Galaburda, A.M. Physiological and Anatomical Evidence for a Magnocellular Defect in Developmental Dyslexia. Proc. Natl. Acad. Sci. USA 1991, 88, 7943–7947. [Google Scholar] [CrossRef]
- McLean, G.M.; Stuart, G.W.; Coltheart, V.; Castles, A. Visual Temporal Processing in Dyslexia and the Magnocellular Deficit Theory: The Need for Speed? J. Exp. Psychol. Hum. Percept. Perform. 2011, 37, 1957. [Google Scholar] [CrossRef] [PubMed]
- Stein, J. To See but Not to Read; the Magnocellular Theory of Dyslexia. Trends Neurosci. 1997, 20, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Stein, J. The Current Status of the Magnocellular Theory of Developmental Dyslexia. Neuropsychologia 2019, 130, 66–77. [Google Scholar] [CrossRef] [PubMed]
- Vidyasagar, T.R.; Pammer, K. Impaired Visual Search in Dyslexia Relates to the Role of the Magnocellular Pathway in Attention. Neuroreport 1999, 10, 1283–1287. [Google Scholar] [CrossRef] [PubMed]
- Vidyasagar, T.R.; Pammer, K. Dyslexia: A Deficit in Visuo-Spatial Attention, Not in Phonological Processing. Trends Cogn. Sci. 2010, 14, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Franceschini, S.; Gori, S.; Ruffino, M.; Pedrolli, K.; Facoetti, A. A Causal Link Between Visual Spatial Attention and Reading Acquisition. Curr Biol. 2012, 22, 814–819. [Google Scholar] [CrossRef]
- Wang, Y.; Hou, P.; Li, W.; Zhang, M.; Zheng, H.; Chen, X. The Influence of Different Current-Intensity Transcranial Alternating Current Stimulation on the Eyes-Open and Eyes-Closed Resting-State Electroencephalography. Front. Hum. Neurosci. 2022, 16, 934382. [Google Scholar] [CrossRef]
- Peterson, R.L.; Pennington, B.F. Developmental Dyslexia. Annu. Rev. Clin. Psychol. 2015, 11, 283–307. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Measure | Controls (n = 28) Mean (SD) | DD (n = 23) Mean (SD) | p-Value |
|---|---|---|---|
| Raven’s APM | 7.96 (2.26) | 7.13 (2.80) | 0.256 |
| Words Reading Speed | 0.79 (1.15) | −1.05 (1.05) | <0.001 |
| Pseudowords Reading Speed | 0.82 (1.47) | −1.30 (0.91) | <0.001 |
| Text Reading Time | 0.02 (0.77) | −3.57 (4.12) | <0.001 |
| Words Reading Errors | 0.12 (0.65) | −0.76 (1.67) | 0.023 |
| Pseudowords Reading Errors | 0.65 (0.54) | −0.28 (1.40) | 0.007 |
| Text Reading Errors | 0.75 (0.60) | −0.75 (1.48) | <0.001 |
| Words Reading Speed | Words Reading Errors | ||||
| F (df) | p | F (df) | p | ||
| Group | 37.63 (1, 48) | <0.001 | Group | 8.23 (1, 47) | 0.006 |
| Offset | 4.40 (1, 48) | 0.041 | Beta (FOOOF-corr.) | 4.75 (1, 47) | 0.034 |
| Group x Beta (FOOOF-corr.) | 8.66 (1, 47) | 0.005 | |||
| Pseudowords Reading Speed | Pseudowords Reading Errors | ||||
| F (df) | p | F (df) | p | ||
| Group | 36.37 (1, 49) | <0.001 | Group | 10.24 (1, 49) | 0.002 |
| Text Reading Time (Syll/Sec) | Text Reading Errors | ||||
| F (df) | p | F (df) | p | ||
| Group | 27.16 (1, 47) | <0.001 | Group | 24.18 (1, 49) | <0.001 |
| Offset | 8.85 (1, 47) | 0.004 | |||
| Group x Offset | 9.03 (1, 47) | 0.004 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Turri, C.; Di Dona, G.; Santoni, A.; Zamfira, D.A.; Franchin, L.; Melcher, D.; Ronconi, L. Periodic and Aperiodic EEG Features as Potential Markers of Developmental Dyslexia. Biomedicines 2023, 11, 1607. https://doi.org/10.3390/biomedicines11061607
Turri C, Di Dona G, Santoni A, Zamfira DA, Franchin L, Melcher D, Ronconi L. Periodic and Aperiodic EEG Features as Potential Markers of Developmental Dyslexia. Biomedicines. 2023; 11(6):1607. https://doi.org/10.3390/biomedicines11061607
Chicago/Turabian StyleTurri, Chiara, Giuseppe Di Dona, Alessia Santoni, Denisa Adina Zamfira, Laura Franchin, David Melcher, and Luca Ronconi. 2023. "Periodic and Aperiodic EEG Features as Potential Markers of Developmental Dyslexia" Biomedicines 11, no. 6: 1607. https://doi.org/10.3390/biomedicines11061607
APA StyleTurri, C., Di Dona, G., Santoni, A., Zamfira, D. A., Franchin, L., Melcher, D., & Ronconi, L. (2023). Periodic and Aperiodic EEG Features as Potential Markers of Developmental Dyslexia. Biomedicines, 11(6), 1607. https://doi.org/10.3390/biomedicines11061607