
Functional Connectivity of the Anterior Cingulate Cortex and the Right Anterior Insula Differentiates between Major Depressive Disorder, Bipolar Disorder and Healthy Controls
 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Methods and Materials
3. MRI Scanning Procedure
4. Image Processing
5. Statistical Analysis
6. Results
6.1. Socio-Demographic and Clinical Characteristics
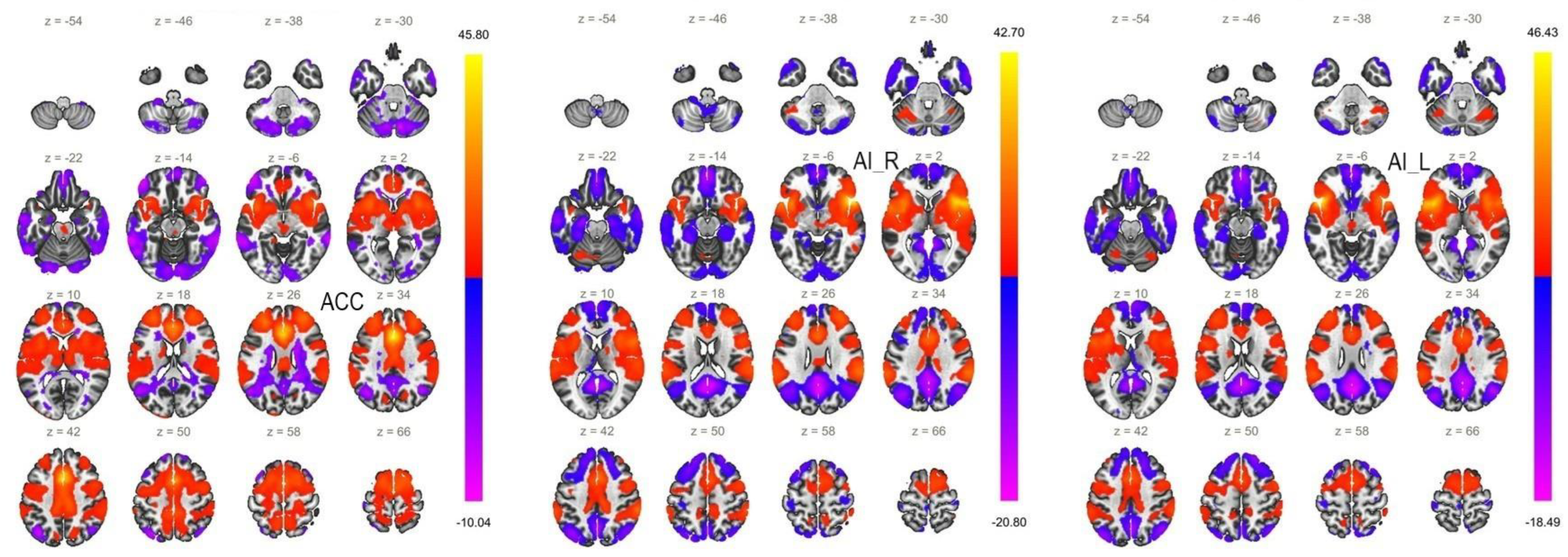
6.2. Functional Connectivity of the ACC Seed
6.3. Functional Connectivity of the AI Seeds
7. Discussion
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ferrari, A. Global, Regional, and National Burden of 12 Mental Disorders in 204 Countries and Territories, 1990–2019: A Systematic Analysis for the Global Burden of Disease Study 2019. Lancet Psychiatry 2022, 9, 137–150. [Google Scholar] [CrossRef]
- Polanczyk, G.V.; Salum, G.A.; Sugaya, L.S.; Caye, A.; Rohde, L.A. Annual Research Review: A Meta-Analysis of the Worldwide Prevalence of Mental Disorders in Children and Adolescents. J. Child Psychol. Psychiatry 2015, 56, 345–365. [Google Scholar] [CrossRef] [PubMed]
- Whitney, D.G.; Peterson, M.D. US National and State-Level Prevalence of Mental Health Disorders and Disparities of Mental Health Care Use in Children. JAMA Pediatr. 2019, 173, 389–391. [Google Scholar] [CrossRef] [PubMed]
- Aragona, M. The Role of Comorbidity in the Crisis of the Current Psychiatric Classification System. Philos. Psychiatry Psychol. 2009, 16, 1–11. [Google Scholar] [CrossRef]
- Schiweck, C.; Arteaga-Henriquez, G.; Aichholzer, M.; Edwin Thanarajah, S.; Vargas-Cáceres, S.; Matura, S.; Grimm, O.; Haavik, J.; Kittel-Schneider, S.; Ramos-Quiroga, J.A.; et al. Comorbidity of ADHD and Adult Bipolar Disorder: A Systematic Review and Meta-Analysis. Neurosci. Biobehav. Rev. 2021, 124, 100–123. [Google Scholar] [CrossRef]
- Dirks, H.; Scherbaum, N.; Kis, B.; Mette, C. ADHD in Adults and Comorbid Substance Use Disorder: Prevalence, Clinical Diagnostics and Integrated Therapy. Fortschr. Neurol. Psychiatr. 2017, 85, 336–344. [Google Scholar] [CrossRef]
- Miller, T.H. Bipolar Disorder. Prim. Care 2016, 43, 269–284. [Google Scholar] [CrossRef] [PubMed]
- Hirschfeld, R.M. Differential Diagnosis of Bipolar Disorder and Major Depressive Disorder. J. Affect. Disord. 2014, 169 (Suppl. S1), S12–S16. [Google Scholar] [CrossRef]
- Benazzi, F. A Continuity between Bipolar II Depression and Major Depressive Disorder? Prog. Neuropsychopharmacol. Biol. Psychiatry 2006, 30, 1043–1050. [Google Scholar] [CrossRef]
- Rosen, C.; Harrow, M.; Tong, L.; Jobe, T.H.; Harrow, H. A Word Is Worth a Thousand Pictures: A 20-Year Comparative Analysis of Aberrant Abstraction in Schizophrenia, Affective Psychosis, and Non-Psychotic Depression. Schizophr. Res. 2021, 238, 1–9. [Google Scholar] [CrossRef]
- Berk, M.; Dodd, S.; Malhi, G.S. “Bipolar Missed States”: The Diagnosis and Clinical Salience of Bipolar Mixed States. Aust. N. Z. J. Psychiatry 2005, 39, 215–221. [Google Scholar] [CrossRef]
- Mistry, S.; Harrison, J.R.; Smith, D.J.; Escott-Price, V.; Zammit, S. The Use of Polygenic Risk Scores to Identify Phenotypes Associated with Genetic Risk of Bipolar Disorder and Depression: A Systematic Review. J. Affect. Disord. 2018, 234, 148–155. [Google Scholar] [CrossRef]
- Aronica, R.; Enrico, P.; Squarcina, L.; Brambilla, P.; Delvecchio, G. Association between Diffusion Tensor Imaging, Inflammation and Immunological Alterations in Unipolar and Bipolar Depression: A Review. Neurosci. Biobehav. Rev. 2022, 143, 104922. [Google Scholar] [CrossRef]
- Siegel-Ramsay, J.E.; Bertocci, M.A.; Wu, B.; Phillips, M.L.; Strakowski, S.M.; Almeida, J.R.C. Distinguishing between Depression in Bipolar Disorder and Unipolar Depression Using Magnetic Resonance Imaging: A Systematic Review. Bipolar Disord. 2022, 24, 474–498. [Google Scholar] [CrossRef] [PubMed]
- Menon, V. Large-Scale Brain Networks and Psychopathology: A Unifying Triple Network Model. Trends Cogn. Sci. 2011, 15, 483–506. [Google Scholar] [CrossRef]
- Sevinc, G.; Gurvit, H.; Nathan Spreng, R. Salience Network Engagement with the Detection of Morally Laden Information. Soc. Cogn. Affect. Neurosci. 2017, 12, 1118. [Google Scholar] [CrossRef]
- Sridharan, D.; Levitin, D.J.; Menon, V. A Critical Role for the Right Fronto-Insular Cortex in Switching between Central-Executive and Default-Mode Networks. Proc. Natl. Acad. Sci. USA 2008, 105, 12569–12574. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Becker, B.; Zhao, W.; Zhao, Z.; Kou, J.; Ma, X.; Geng, Y.; Ren, P.; Kendrick, K.M. Oxytocin Modulates Attention Switching Between Interoceptive Signals and External Social Cues. Neuropsychopharmacology 2018, 43, 294–301. [Google Scholar] [CrossRef]
- Rolls, E.T. The Cingulate Cortex and Limbic Systems for Emotion, Action, and Memory. Brain Struct. Funct. 2019, 224, 3001. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, R.H.; Andrews-Hanna, J.R.; Wager, T.D.; Pizzagalli, D.A. Large-Scale Network Dysfunction in Major Depressive Disorder: A Meta-Analysis of Resting-State Functional Connectivity. JAMA Psychiatry 2015, 72, 603–611. [Google Scholar] [CrossRef]
- Huang, H.; Chen, C.; Rong, B.; Wan, Q.; Chen, J.; Liu, Z.; Zhou, Y.; Wang, G.; Wang, H. Resting-State Functional Connectivity of Salience Network in Schizophrenia and Depression. Sci. Rep. 2022, 12, 11204. [Google Scholar] [CrossRef]
- McTeague, L.M.; Rosenberg, B.M.; Lopez, J.W.; Carreon, D.M.; Huemer, J.; Jiang, Y.; Chick, C.F.; Eickhoff, S.B.; Etkin, A. Identification of Common Neural Circuit Disruptions in Emotional Processing Across Psychiatric Disorders. Am. J. Psychiatry 2020, 177, 411–421. [Google Scholar] [CrossRef]
- Goodkind, M.; Eickhoff, S.B.; Oathes, D.J.; Jiang, Y.; Chang, A.; Jones-Hagata, L.B.; Ortega, B.N.; Zaiko, Y.V.; Roach, E.L.; Korgaonkar, M.S.; et al. Identification of a Common Neurobiological Substrate for Mental Illness. JAMA Psychiatry 2015, 72, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Redlich, R.; Almeida, J.R.; Grotegerd, D.; Opel, N.; Kugel, H.; Heindel, W.; Arolt, V.; Phillips, M.L.; Dannlowski, U. Brain Morphometric Biomarkers Distinguishing Unipolar and Bipolar Depression. A Voxel-Based Morphometry-Pattern Classification Approach. JAMA Psychiatry 2014, 71, 1222–1230. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.; Li, M.L.; Li, Y.F.; Li, X.J.; Meng, Y.; Liang, S.; Li, Z.; Guo, W.; Wang, Q.; Deng, W.; et al. Anterior Cingulate Cortex, Insula and Amygdala Seed-Based Whole Brain Resting-State Functional Connectivity Differentiates Bipolar from Unipolar Depression. J. Affect. Disord. 2020, 274, 38–47. [Google Scholar] [CrossRef]
- Zeng, C.; Xue, Z.; Ross, B.; Zhang, M.; Liu, Z.; Wu, G.; Ouyang, X.; Li, D.; Pu, W. Salience-Thalamic Circuit Uncouples in Major Depressive Disorder, but Not in Bipolar Depression. J. Affect. Disord. 2020, 269, 43–50. [Google Scholar] [CrossRef]
- Ellard, K.K.; Zimmerman, J.P.; Kaur, N.; Van Dijk, K.R.A.; Roffman, J.L.; Nierenberg, A.A.; Dougherty, D.D.; Deckersbach, T.; Camprodon, J.A. Functional Connectivity Between Anterior Insula and Key Nodes of Frontoparietal Executive Control and Salience Networks Distinguish Bipolar Depression from Unipolar Depression and Healthy Control Subjects. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, D.V.; Lecrubier, Y.; Sheehan, K.H.; Amorim, P.; Janavs, J.; Weiller, E.; Hergueta, T.; Baker, R.; Dunbar, G.C. The Mini-International Neuropsychiatric Interview (M.I.N.I.): The Development and Validation of a Structured Diagnostic Psychiatric Interview for DSM-IV and ICD-10. J. Clin. Psychiatry 1998, 59 (Suppl. S20), 22–33, quiz 34. [Google Scholar]
- Montgomery, S.A.; Asberg, M. A New Depression Scale Designed to Be Sensitive to Change. Br. J. Psychiatry 1979, 134, 382–389. [Google Scholar] [CrossRef]
- Whitfield-Gabrieli, S.; Nieto-Castanon, A. Conn: A Functional Connectivity Toolbox for Correlated and Anticorrelated Brain Networks. Brain Connect. 2012, 2, 125–141. [Google Scholar] [CrossRef] [PubMed]
- Sladky, R.; Friston, K.J.; Tröstl, J.; Cunnington, R.; Moser, E.; Windischberger, C. Slice-Timing Effects and Their Correction in Functional MRI. Neuroimage 2011, 58, 588–594. [Google Scholar] [CrossRef]
- Bates, J.F.; Goldman-Rakic, P.S. Prefrontal Connections of Medial Motor Areas in the Rhesus Monkey. J. Comp. Neurol. 1993, 336, 211–228. [Google Scholar] [CrossRef] [PubMed]
- King, J.B.; Anderson, J.S.; Yurgelun-Todd, D.A.; Subramaniam, P.; Ehrler, M.R.; Lopez-Larson, M.P. Decreased Anterior Cingulate Activation in a Motor Task in Youths with Bipolar Disorder. J. Child Psychol. Psychiatry 2018, 59, 900–907. [Google Scholar] [CrossRef]
- Drachman, R.; Colic, L.; Sankar, A.; Spencer, L.; Goldman, D.A.; Villa, L.M.; Kim, J.A.; Oquendo, M.A.; Pittman, B.; Blumberg, H.P. Rethinking “Aggression” and Impulsivity in Bipolar Disorder: Risk, Clinical and Brain Circuitry Features. J. Affect. Disord. 2022, 303, 331–339. [Google Scholar] [CrossRef] [PubMed]
- Hanford, L.C.; Nazarov, A.; Hall, G.B.; Sassi, R.B. Cortical Thickness in Bipolar Disorder: A Systematic Review. Bipolar Disord. 2016, 18, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Angst, J.; Gamma, A.; Benazzi, F.; Ajdacic, V.; Rössler, W. Does Psychomotor Agitation in Major Depressive Episodes Indicate Bipolarity? Evidence from the Zurich Study. Eur. Arch. Psychiatry Clin. Neurosci. 2009, 259, 55–63. [Google Scholar] [CrossRef]
- Iwanami, T.; Maeshima, H.; Baba, H.; Satomura, E.; Namekawa, Y.; Shimano, T.; Suzuki, T.; Arai, H. Psychomotor Agitation in Major Depressive Disorder Is a Predictive Factor of Mood-Switching. J. Affect. Disord. 2015, 170, 185–189. [Google Scholar] [CrossRef]
- Terziivanova, P.D.; Haralanov, S.H. Latent Bipolarity In Unipolar Depression: Experimental Findings, Conceptual Analysis And Implications For Treatment Strategies. Folia Med. 2014, 56, 282–288. [Google Scholar] [CrossRef]
- Zamfir, M.; Sharif, B.; Locke, S.; Ehrlich, A.T.; Ochandarena, N.E.; Scherrer, G.; Ribeiro-da-Silva, A.; Kieffer, B.L.; Séguéla, P. Distinct and Sex-Specific Expression of Mu Opioid Receptors in Anterior Cingulate and Somatosensory S1 Cortical Areas. Pain 2023, 164, 703–716. [Google Scholar] [CrossRef]
- Kang, L.; Zhang, A.; Sun, N.; Liu, P.; Yang, C.; Li, G.; Liu, Z.; Wang, Y.; Zhang, K. Functional Connectivity between the Thalamus and the Primary Somatosensory Cortex in Major Depressive Disorder: A Resting-State FMRI Study. BMC Psychiatry 2018, 18, 339. [Google Scholar] [CrossRef]
- Zhu, X.; Tang, H.D.; Dong, W.Y.; Kang, F.; Liu, A.; Mao, Y.; Xie, W.; Zhang, X.; Cao, P.; Zhou, W.; et al. Distinct Thalamocortical Circuits Underlie Allodynia Induced by Tissue Injury and by Depression-like States. Nat. Neurosci. 2021, 24, 542–553. [Google Scholar] [CrossRef]
- Liang, S.; Deng, W.; Li, X.; Greenshaw, A.J.; Wang, Q.; Li, M.; Ma, X.; Bai, T.J.; Bo, Q.J.; Cao, J.; et al. Biotypes of Major Depressive Disorder: Neuroimaging Evidence from Resting-State Default Mode Network Patterns. Neuroimage Clin. 2020, 28, 102514. [Google Scholar] [CrossRef]
- Feng, J.; Cheng, W.; Rolls, E.T.; Ruan, H. Functional Connectivities in the Brain That Mediate the Association Between Depressive Problems and Sleep Quality. JAMA Psychiatry 2018, 75, 1052–1061. [Google Scholar] [CrossRef]
- Yu, H.; Li, M.L.; Meng, Y.; Li, X.J.; Wei, W.; Li, Y.F.; Li, L.; Guo, W.; Wang, Q.; Deng, W.; et al. Inferior Frontal Gyrus Seed-Based Resting-State Functional Connectivity and Sustained Attention across Manic/Hypomanic, Euthymic and Depressive Phases of Bipolar Disorder. J. Affect. Disord. 2021, 282, 930–938. [Google Scholar] [CrossRef] [PubMed]
- Martens, M.; Filippini, N.; Masaki, C.; Godlewska, B.R. Functional Connectivity between Task-Positive Networks and the Left Precuneus as a Biomarker of Response to Lamotrigine in Bipolar Depression: A Pilot Study. Pharmaceuticals 2021, 14, 534. [Google Scholar] [CrossRef] [PubMed]
- Kumral, E.; Çetin, F.E.; Özdemir, H.N. Cognitive and Behavioral Disorders in Patients with Superior Parietal Lobule Infarcts. Can. J. Neurol. Sci. 2022, 1–9. [Google Scholar] [CrossRef]
- Vialatte, A.; Yeshurun, Y.; Khan, A.Z.; Rosenholtz, R.; Pisella, L. Superior Parietal Lobule: A Role in Relative Localization of Multiple Different Elements. Cereb. Cortex 2021, 31, 658–671. [Google Scholar] [CrossRef]
- Rolls, E.T.; Cheng, W.; Gong, W.; Qiu, J.; Zhou, C.; Zhang, J.; Lv, W.; Ruan, H.; Wei, D.; Cheng, K.; et al. Functional Connectivity of the Anterior Cingulate Cortex in Depression and in Health. Cereb. Cortex 2019, 29, 3617–3630. [Google Scholar] [CrossRef]
- Su, L.; Cai, Y.; Xu, Y.; Dutt, A.; Shi, S.; Bramon, E. Cerebral Metabolism in Major Depressive Disorder: A Voxel-Based Meta-Analysis of Positron Emission Tomography Studies. BMC Psychiatry 2014, 14, 321. [Google Scholar] [CrossRef]
- Chen, G.; Zhao, L.; Jia, Y.; Zhong, S.; Chen, F.; Luo, X.; Qiu, S.; Lai, S.; Qi, Z.; Huang, L.; et al. Abnormal Cerebellum-DMN Regions Connectivity in Unmedicated Bipolar II Disorder. J. Affect. Disord. 2019, 243, 441–447. [Google Scholar] [CrossRef]
- Meerwijk, E.L.; Ford, J.M.; Weiss, S.J. Brain Regions Associated with Psychological Pain: Implications for a Neural Network and Its Relationship to Physical Pain. Brain Imaging Behav. 2013, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Güntürkün, O.; Ströckens, F.; Ocklenburg, S. Brain Lateralization: A Comparative Perspective. Physiol. Rev. 2020, 100, 1019–1063. [Google Scholar] [CrossRef]
- Critchley, H.D.; Wiens, S.; Rotshtein, P.; Öhman, A.; Dolan, R.J. Neural Systems Supporting Interoceptive Awareness. Nat. Neurosci. 2004, 7, 189–195. [Google Scholar] [CrossRef]
- Scalabrini, A.; Wolman, A.; Northoff, G. The Self and Its Right Insula-Differential Topography and Dynamic of Right vs. Left Insula. Brain Sci. 2021, 11, 1312. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Duncan, N.W.; de Greck, M.; Northoff, G. Is There a Core Neural Network in Empathy? An FMRI Based Quantitative Meta-Analysis. Neurosci. Biobehav. Rev. 2011, 35, 903–911. [Google Scholar] [CrossRef] [PubMed]
- Stoyanov, D. Molecular Pathway Phenotypes and Endophenotypes in Psychiatry- Part II. Curr. Top. Med. Chem. 2021, 21, 1439–1440. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HC n = 43 | MDD n = 35 | BD n = 25 | Significance Level between MDD and BD | Significance Level between HC and MDD & BD | |
|---|---|---|---|---|---|
| Age (mean ± SD) | 40.25 (±10.73) | 40.97 (±10.86) | 41.72 (±9.62) | p = 1.000 c | p = 1.000 a/1.000 a |
| Gender (M/F) | 43 (14/29) | 35 (14/21) | 25 (9/16) | p = 0.793 b | |
| Education (years) | 15.93 (±3.26) | 13.8 (±2.93) | 13.88 (±2.71) | p = 0.915 c | p = 0.012 a/0.035 a |
| MADRS | 2.28 (±2.25) | 30.2 (±4.14) | 29.76 (±6.17) | p = 0.742 c | p < 0.001 a/0.001 a |
| Age at onset (mean ± SD) | - | 30.24 (11.01) | 29.72 (9.19) | p = 0.850 c | - |
| Illness duration (mean ± SD) | - | 130.97 (113.93) | 148.6 (101.64) | p = 0.544 c | - |
| Episode duration (mean ± SD) | - | 21 (36.65) | 15.8 (14.99) | p = 0.508 c | - |
| Number of previous episodes (mean ± SD) | - | 3.97 (3.56) | 6.36 (4.72) | p = 0.035 c | - |
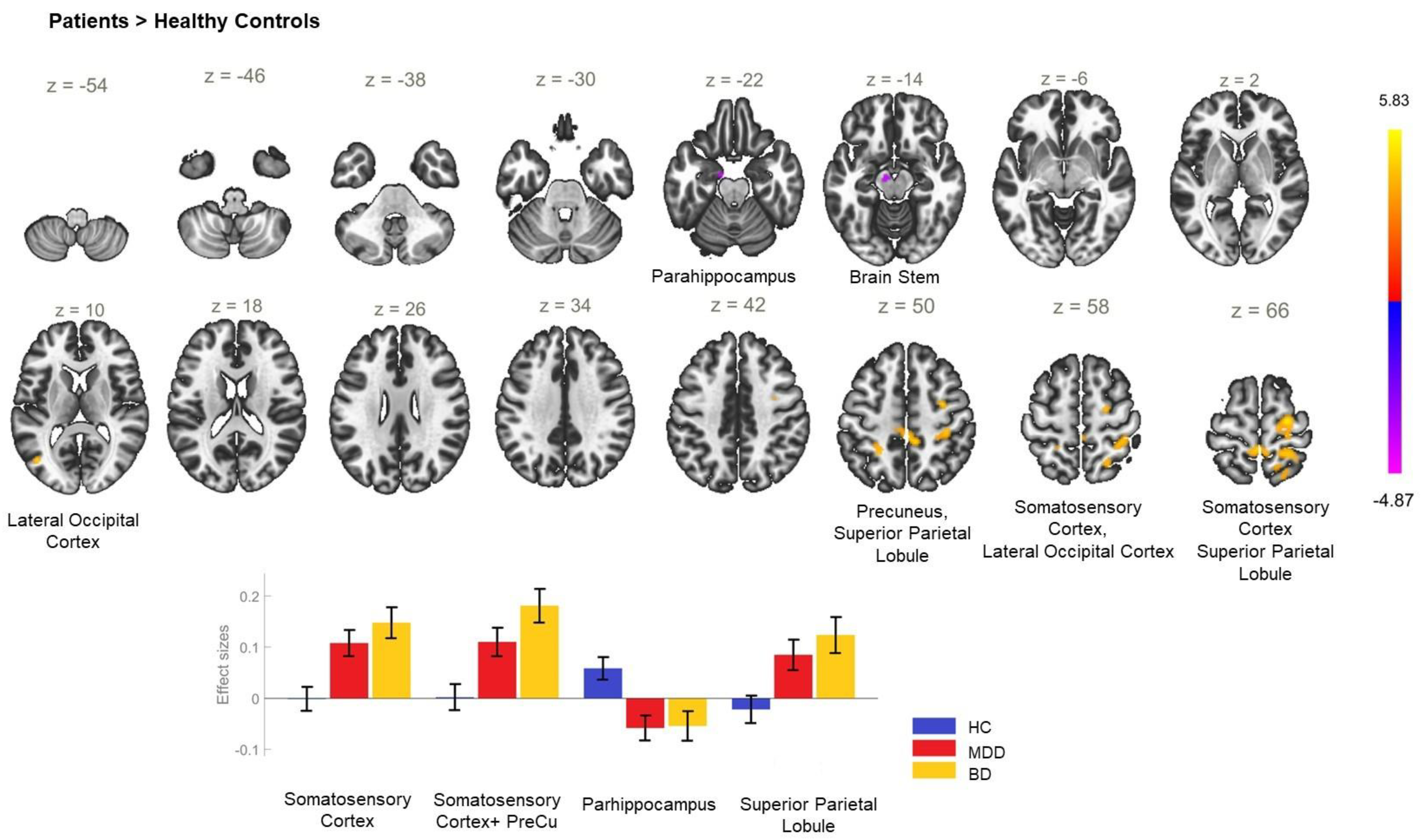
| Between-Group Contrast | MNI Coordinates x, y, z | Cluster Size | Cluster Threshold (p < 0.05, FWE) | Regions within the Cluster |
|---|---|---|---|---|
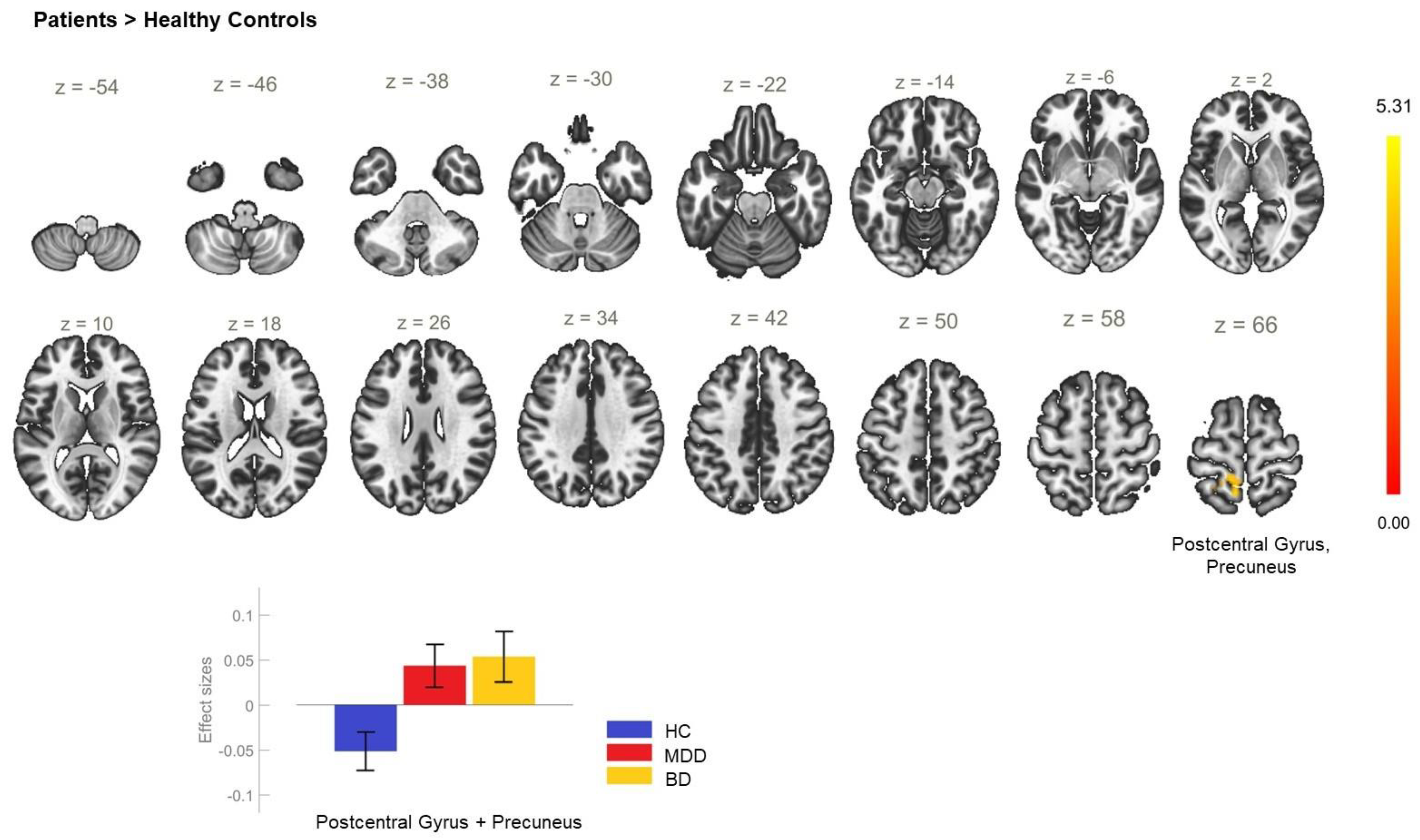
| P > HC | +32 −4 +46 | 697 | <0.001 | PostCG_R, PreCG_R, SPL_R |
| +2 −34 +52 | 526 | <0.001 | PostCG_R, SPL_R, PreCu, LOC_R—superior division, PostCG_L | |
| −24 −48 +52 | 106 | 0.017 | SPL_L | |
| HC > P | −10 −10 −24 | 109 | 0.015 | PHPG_L—anterior division, Brain stem |
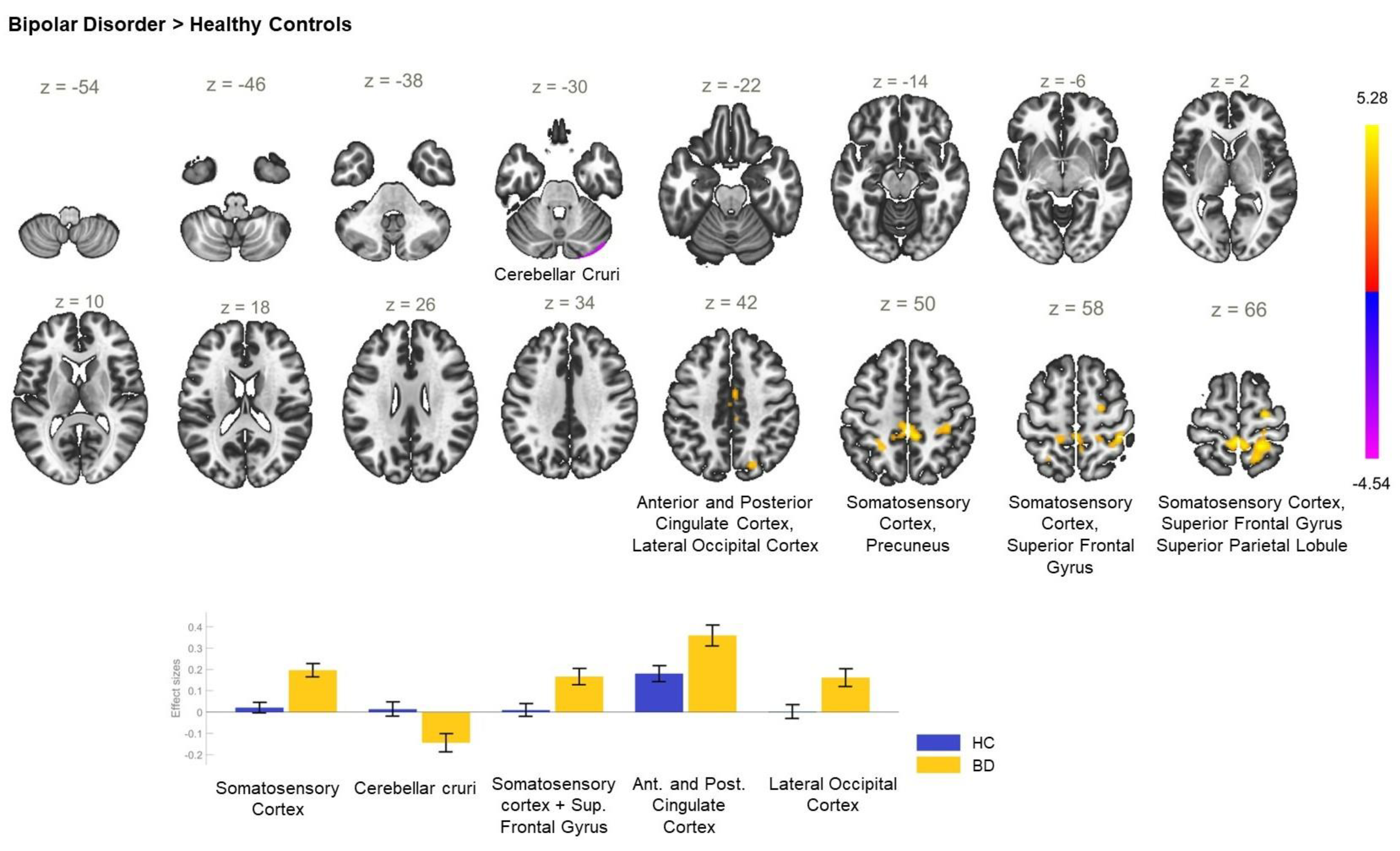
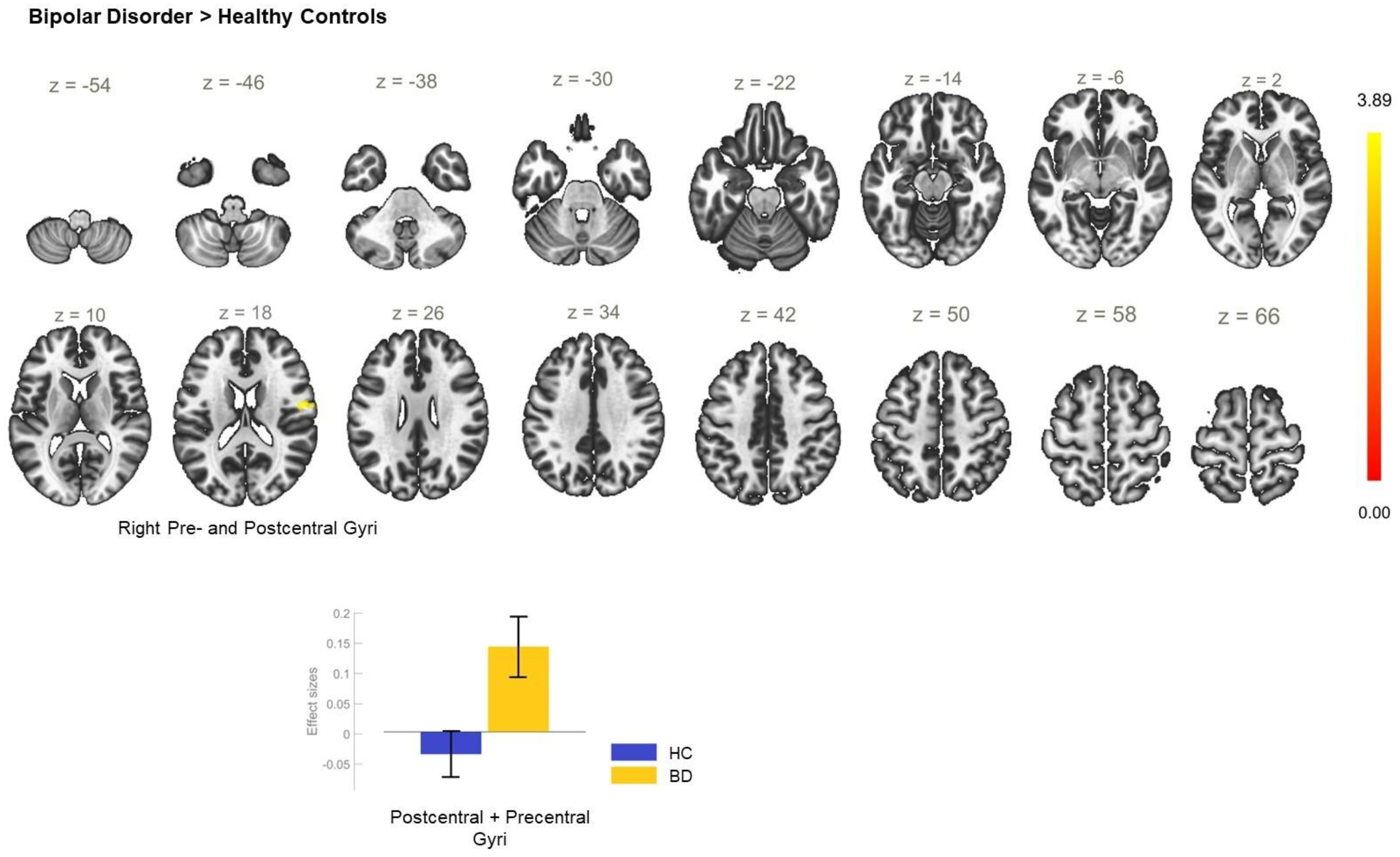
| BD > HC | −02 −36 +66 | 1393 | <0.001 | PostCG_R, SPL_R, PostCG_L, PreCu |
| +24 −12 +66 | 120 | 0.009 | PreCG_R, SFG_R | |
| −2 −14 +44 | 105 | 0.018 | ACC, PCC, SMC_R | |
| +18 −72 +40 | 86 | 0.047 | LOC_R—superior division, PreCu, CC_R | |
| HC > BD | +24 −92 −28 | 138 | 0.004 | CerC1_R, CerC2_R |
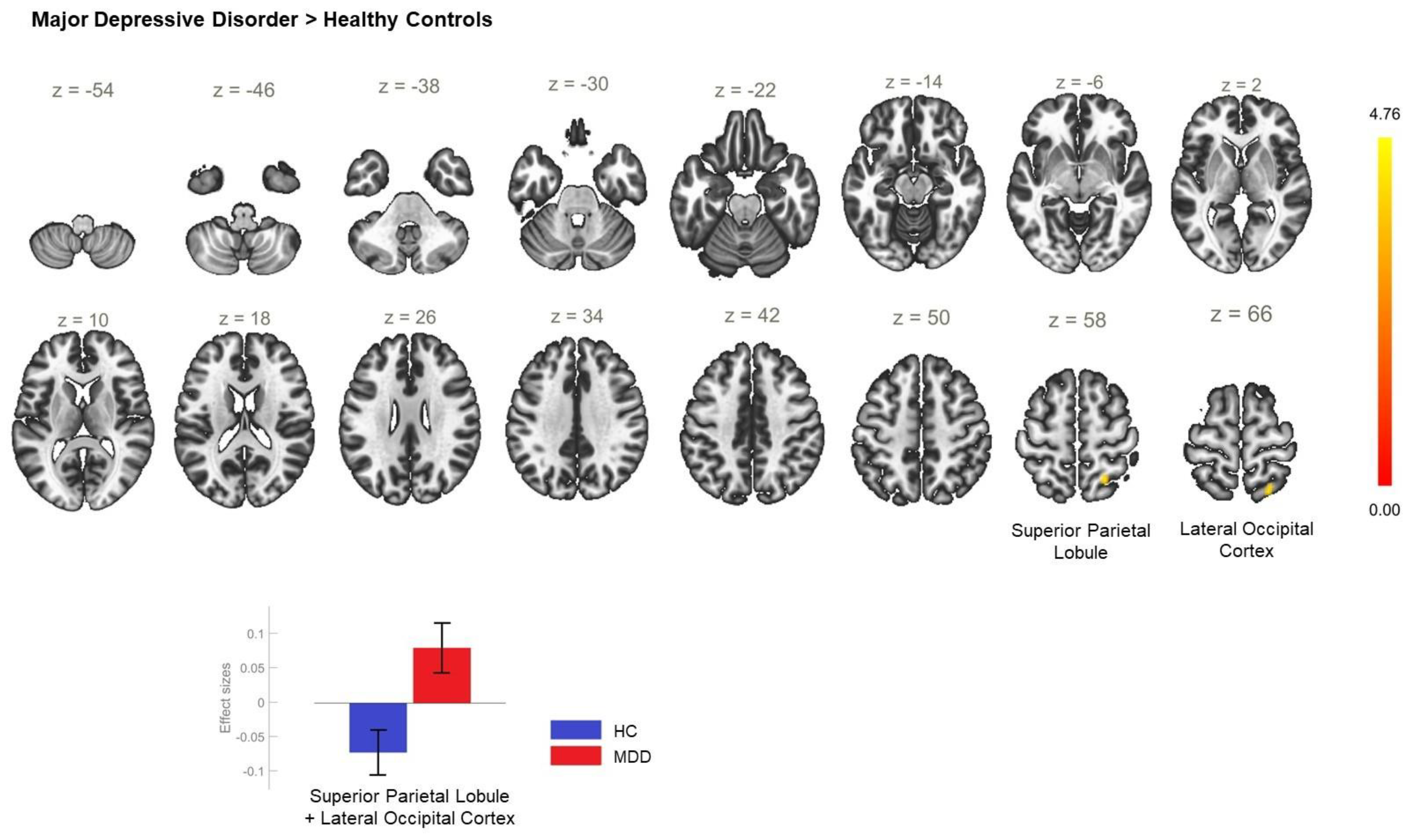
| MDD > HC | +22 −56 +56 | 112 | 0.013 | SPL_R, LOC_R, superior division |
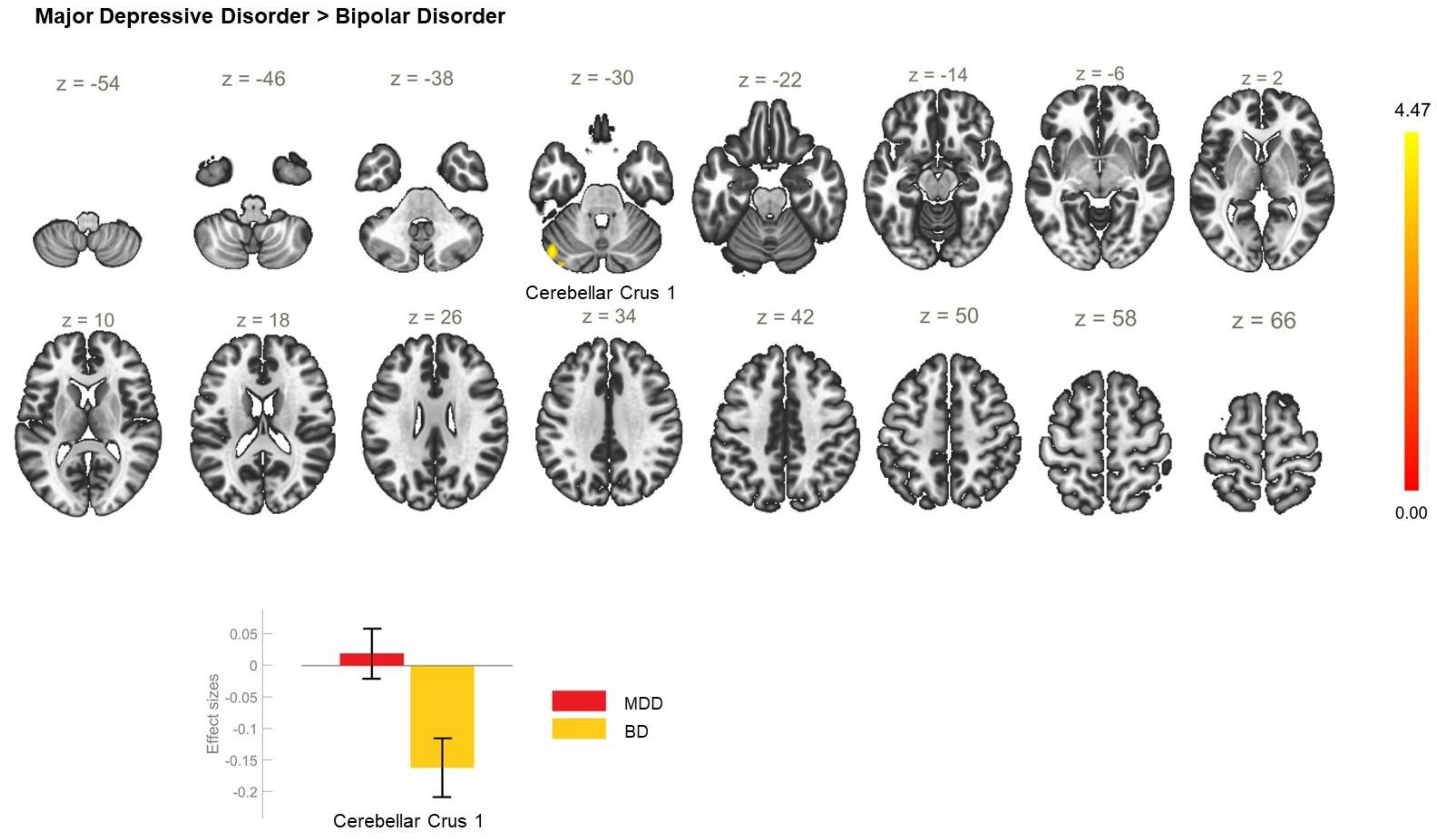
| MDD > BD | −46 −70 −32 | 191 | <0.001 | CerC1_L |
| Between-Group Contrast | MNI Coordinates x, y, z | Cluster Size | Cluster Threshold (p < 0.05, FWE) | Regions within the Clusters |
|---|---|---|---|---|
| P > HC | −8 −52 +64 | 201 | <0.001 | PostCG_L, PreCu |
| BD > HC | +54 −06 +18 | 108 | 0.017 | PostCG_R, PreCG_R |
| MDD > BD | −4 +12 +58 | 80 | 0.068 | SFG_L, SFG_R, SMC_R |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Todeva-Radneva, A.; Kandilarova, S.; Paunova, R.; Stoyanov, D.; Zdravkova, T.; Sladky, R. Functional Connectivity of the Anterior Cingulate Cortex and the Right Anterior Insula Differentiates between Major Depressive Disorder, Bipolar Disorder and Healthy Controls. Biomedicines 2023, 11, 1608. https://doi.org/10.3390/biomedicines11061608
Todeva-Radneva A, Kandilarova S, Paunova R, Stoyanov D, Zdravkova T, Sladky R. Functional Connectivity of the Anterior Cingulate Cortex and the Right Anterior Insula Differentiates between Major Depressive Disorder, Bipolar Disorder and Healthy Controls. Biomedicines. 2023; 11(6):1608. https://doi.org/10.3390/biomedicines11061608
Chicago/Turabian StyleTodeva-Radneva, Anna, Sevdalina Kandilarova, Rositsa Paunova, Drozdstoy Stoyanov, Tina Zdravkova, and Ronald Sladky. 2023. "Functional Connectivity of the Anterior Cingulate Cortex and the Right Anterior Insula Differentiates between Major Depressive Disorder, Bipolar Disorder and Healthy Controls" Biomedicines 11, no. 6: 1608. https://doi.org/10.3390/biomedicines11061608
APA StyleTodeva-Radneva, A., Kandilarova, S., Paunova, R., Stoyanov, D., Zdravkova, T., & Sladky, R. (2023). Functional Connectivity of the Anterior Cingulate Cortex and the Right Anterior Insula Differentiates between Major Depressive Disorder, Bipolar Disorder and Healthy Controls. Biomedicines, 11(6), 1608. https://doi.org/10.3390/biomedicines11061608