
SARS-CoV-2 Gut-Targeted Epitopes: Sequence Similarity and Cross-Reactivity Join Together for Molecular Mimicry



Abstract
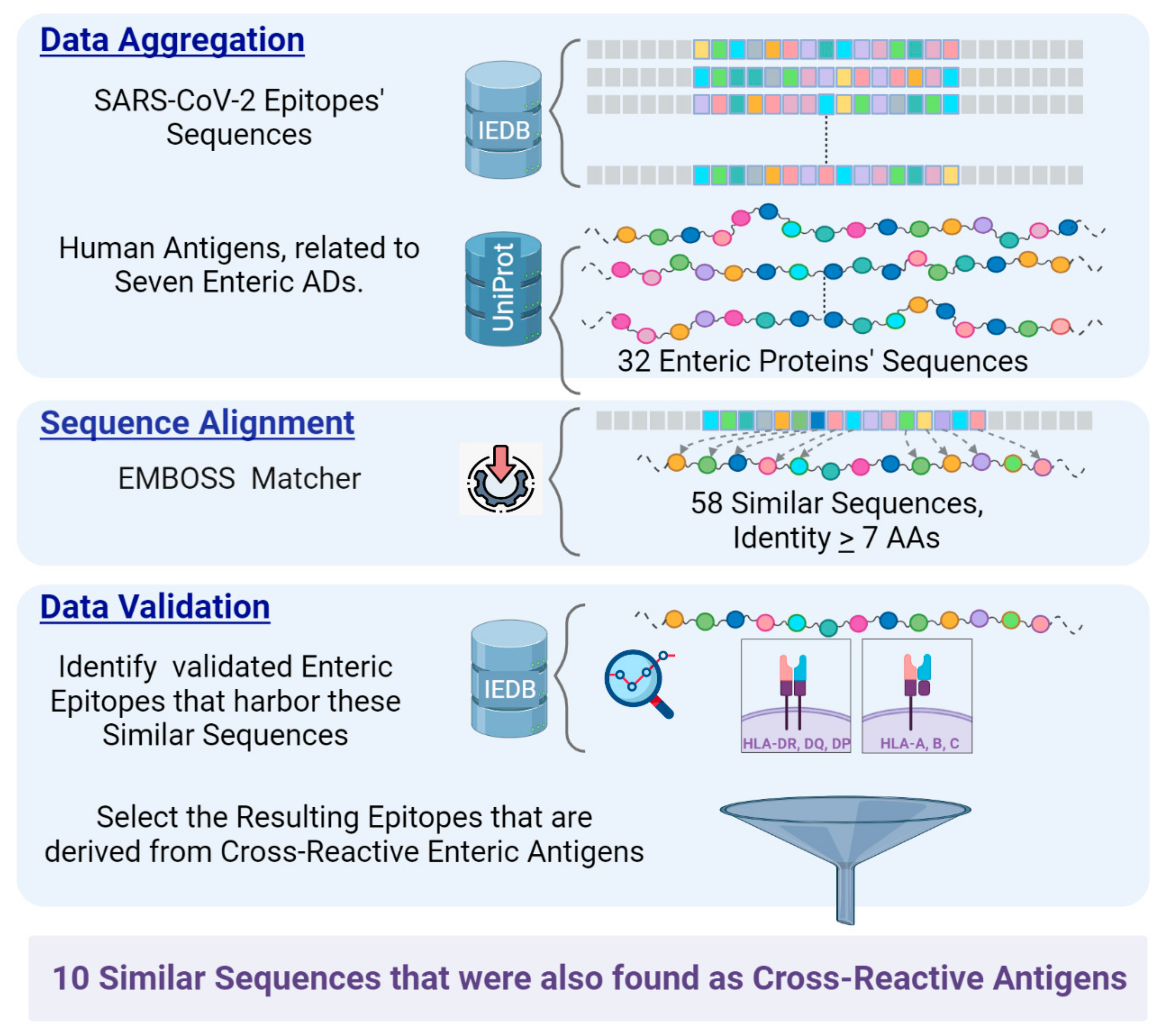
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
- SARS-CoV-2 spike, nucleo, envelope, and membrane protein cross-react with major enteric and hepatic self-antigens.
- Sequence similarity exists between multiple coronavirus and intestinal/hepatic epitopes.
- Cross-reactive autoantibodies and sequence similarity are major potential drivers of molecular mimicry in the auto-immunogenic avalanche.
- The combined shared cross-reactive and sequence identical core in gut-associated epitopes further strengthens the connection to molecular mimicry [39,131]. The two mechanisms detected major essential proteins that represent specific autoantigens for CD, AIH, and PBC or contribute to leaky gut by disrupting tight junction functionality, thus becoming directly involved in the corresponding auto-immuno-genesis [29].
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gusev, E.; Sarapultsev, A.; Solomatina, L.; Chereshnev, V. SARS-CoV-2-Specific Immune Response and the Pathogenesis of COVID-19. Int. J. Mol. Sci. 2022, 23, 1716. [Google Scholar] [CrossRef]
- Liu, Y.; Sawalha, A.H.; Lu, Q. COVID-19 and autoimmune diseases. Curr. Opin. Rheumatol. 2021, 33, 155–162. [Google Scholar] [CrossRef]
- Hosseini, P.; Fallahi, M.S.; Erabi, G.; Pakdin, M.; Zarezadeh, S.M.; Faridzadeh, A.; Entezari, S.; Ansari, A.; Poudineh, M.; Deravi, N. Multisystem Inflammatory Syndrome and Autoimmune Diseases Following COVID-19: Molecular Mechanisms and Therapeutic Opportunities. Front. Mol. Biosci. 2022, 9, 804109. [Google Scholar] [CrossRef]
- Mohkhedkar, M.; Venigalla, S.S.K.; Janakiraman, V. Untangling COVID-19 and autoimmunity: Identification of plausible targets suggests multi organ involvement. Mol. Immunol. 2021, 137, 105–113. [Google Scholar] [CrossRef]
- Yazdanpanah, N.; Rezaei, N. Autoimmune complications of COVID-19. J. Med. Virol. 2022, 94, 54–62. [Google Scholar] [CrossRef] [PubMed]
- Dotan, A.; Muller, S.; Kanduc, D.; David, P.; Halpert, G.; Shoenfeld, Y. The SARS-CoV-2 as an instrumental trigger of autoimmunity. Autoimmun. Rev. 2021, 20, 102792. [Google Scholar] [CrossRef] [PubMed]
- Damoiseaux, J.; Dotan, A.; Fritzler, M.J.; Bogdanos, D.P.; Meroni, P.L.; Roggenbuck, D.; Goldman, M.; Landegren, N.; Bastard, P.; Shoenfeld, Y.; et al. Autoantibodies and SARS-CoV2 infection: The spectrum from association to clinical implication: Report of the 15th Dresden Symposium on Autoantibodies. Autoimmun. Rev. 2022, 21, 103012. [Google Scholar] [CrossRef]
- Dotan, A.; Mahroum, N.; Bogdanos, D.P.; Shoenfeld, Y. COVID-19 as an infectome paradigm of autoimmunity. J. Allergy Clin. Immunol. 2022, 149, 63–64. [Google Scholar] [CrossRef]
- Mobasheri, L.; Nasirpour, M.H.; Masoumi, E.; Azarnaminy, A.F.; Jafari, M.; Esmaeili, S.A. SARS-CoV-2 triggering autoimmune diseases. Cytokine 2022, 154, 155873. [Google Scholar] [CrossRef] [PubMed]
- Darmarajan, T.; Paudel, K.R.; Candasamy, M.; Chellian, J.; Madheswaran, T.; Sakthivel, L.P.; Goh, B.H.; Gupta, P.K.; Jha, N.K.; Devkota, H.P.; et al. Autoantibodies and autoimmune disorders in SARS-CoV-2 infection: Pathogenicity and immune regulation. Environ. Sci. Pollut. Res. Int. 2022, 29, 54072–54087. [Google Scholar] [CrossRef] [PubMed]
- Baiocchi, G.C.; Vojdani, A.; Rosenberg, A.Z.; Vojdani, E.; Halpert, G.; Ostrinski, Y.; Zyskind, I.; Filgueiras, I.S.; Schimke, L.F.; Marques, A.H.C.; et al. Cross-sectional analysis reveals autoantibody signatures associated with COVID-19 severity. J. Med. Virol. 2023, 95, e28538. [Google Scholar] [CrossRef]
- Lavi, Y.; Vojdani, A.; Halpert, G.; Sharif, K.; Ostrinski, Y.; Zyskind, I.; Lattin, M.T.; Zimmerman, J.; Silverberg, J.I.; Rosenberg, A.Z.; et al. Dysregulated Levels of Circulating Autoantibodies against Neuronal and Nervous System Autoantigens in COVID-19 Patients. Diagnostics 2023, 13, 687. [Google Scholar] [CrossRef]
- Dotan, A.; David, P.; Arnheim, D.; Shoenfeld, Y. The autonomic aspects of the post-COVID19 syndrome. Autoimmun. Rev. 2022, 21, 103071. [Google Scholar] [CrossRef] [PubMed]
- Mahroum, N.; Zoubi, M.; Lavine, N.; Ohayon, A.; Amital, H.; Shoenfeld, Y. The mosaic of autoimmunity—A taste for more. The 12th international congress of autoimmunity 2021 (AUTO12) virtual. Autoimmun. Rev. 2021, 20, 102945. [Google Scholar] [CrossRef] [PubMed]
- Miller, F.W. The increasing prevalence of autoimmunity and autoimmune diseases: An urgent call to action for improved understanding, diagnosis, treatment, and prevention. Curr. Opin. Immunol. 2023, 80, 102266. [Google Scholar] [CrossRef] [PubMed]
- Vojdani, A.; Vojdani, E. The Role of Exposomes in the Pathophysiology of Autoimmune Diseases I: Toxic Chemicals and Food. Pathophysiology 2021, 28, 513–543. [Google Scholar] [CrossRef]
- Vojdani, A.; Vojdani, E.; Rosenberg, A.Z.; Shoenfeld, Y. The Role of Exposomes in the Pathophysiology of Autoimmune Diseases II: Pathogens. Pathophysiol. Off. J. Int. Soc. Pathophysiol. 2022, 29, 243–280. [Google Scholar] [CrossRef]
- Abdelhamid, L.; Luo, X.M. Diet and Hygiene in Modulating Autoimmunity During the Pandemic Era. Front. Immunol. 2022, 12, 749774. [Google Scholar] [CrossRef]
- Garabatos, N.; Santamaria, P. Gut Microbial Antigenic Mimicry in Autoimmunity. Front. Immunol. 2022, 13, 873607. [Google Scholar] [CrossRef]
- Hassan, A.; Blanchard, N. Microbial (co)infections: Powerful immune influencers. PLoS Pathog. 2022, 18, e1010212. [Google Scholar] [CrossRef]
- Johnson, D.; Jiang, W. Infectious diseases, autoantibodies, and autoimmunity. J. Autoimmun. 2023, 137, 102962. [Google Scholar] [CrossRef]
- Jara, L.J.; Vera-Lastra, O.; Mahroum, N.; Pineda, C.; Shoenfeld, Y. Autoimmune post-COVID vaccine syndromes: Does the spectrum of autoimmune/inflammatory syndrome expand? Clin. Rheumatol. 2022, 41, 1603–1609. [Google Scholar] [CrossRef]
- Segal, Y.; Shoenfeld, Y. Vaccine-induced autoimmunity: The role of molecular mimicry and immune crossreaction. Cell. Mol. Immunol. 2018, 15, 586–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguayo-Patrón, S.V.; Calderón de la Barca, A.M. Old Fashioned vs. Ultra-Processed-Based Current Diets: Possible Implication in the Increased Susceptibility to Type 1 Diabetes and Celiac Disease in Childhood. Foods 2017, 6, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, A.; Benzvi, C. Microbial transglutaminase is a very frequently used food additive and is a potential inducer of autoimmune/neurodegenerative diseases. Toxics 2021, 9, 233. [Google Scholar] [CrossRef] [PubMed]
- Lerner, A.; Benzvi, C. “Let food be thy medicine”: Gluten and potential role in neurodegeneration. Cells 2021, 10, 756. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, E.; Katahira, Y.; Mizoguchi, I.; Watanabe, A.; Furusaka, Y.; Sekine, A.; Yamagishi, M.; Sonoda, J.; Miyakawa, S.; Inoue, S.; et al. Chemical- and Drug-Induced Allergic, Inflammatory, and Autoimmune Diseases Via Haptenation. Biology 2023, 12, 123. [Google Scholar] [CrossRef] [PubMed]
- Borba, V.; Malkova, A.; Basantsova, N.; Halpert, G.; Andreoli, L.; Tincani, A.; Amital, H.; Shoenfeld, Y. Classical Examples of the Concept of the ASIA Syndrome. Biomolecules 2020, 10, 436. [Google Scholar] [CrossRef]
- Lerner, A.; Matthias, T. Changes in intestinal tight junction permeability associated with industrial food additives explain the rising incidence of autoimmune disease. Autoimmun. Rev. 2015, 14, 479–489. [Google Scholar] [CrossRef] [Green Version]
- Lerner, A. COVID-19 and the human gut: A new runner on the tract. Int. J. Celiac Dis. 2020, 8, 64–67. [Google Scholar] [CrossRef]
- Lerner, A. Are my patients with celiac disease at higher risk of COVID-19 virus. Int. J. Celiac Dis. 2020, 8, 35–38. [Google Scholar] [CrossRef]
- Lerner, A.; McCarty, M.F. The aging bowel dysfunction and elderly vulnerability towards COVID-19 infection. Life 2021, 11, 97. [Google Scholar] [CrossRef]
- Lerner, A. The COVID-19 vaccination debate: CoV-2 in celiac disease: A Pathogen or just along for the ride? Int. J. Celiac Dis. 2021, 9, 6–9. [Google Scholar] [CrossRef]
- Dvornikova, K.A.; Bystrova, E.Y.; Churilov, L.P.; Lerner, A. Pathogenesis of the inflammatory bowel disease in context of SARS-COV-2 infection. Mol. Biol. Rep. 2021, 48, 5745–5758. [Google Scholar] [CrossRef]
- Samasca, G.; Lerner, A. Celiac disease in the COVID-19 pandemic. J. Transl. Autoimmun. 2021, 4, 100120. [Google Scholar] [CrossRef]
- Lerner, A.; Aminov, R.; Matthias, T. Dysbiosis may trigger autoimmune diseases via inappropriate post-translational modification of host proteins. Front. Microbiol. 2016, 7, 84. [Google Scholar] [CrossRef]
- Lerner, A.; Aminov, R.; Matthias, T. Transglutaminases in dysbiosis as potential environmental drivers of autoimmunity. Front. Microbiol. 2017, 8, 66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, A.; Matthias, T.; Aminov, R. Potential effects of horizontal gene exchange in the human gut. Front. Immunol. 2017, 8, 1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojas, M.; Restrepo-Jiménez, P.; Monsalve, D.M.; Pacheco, Y.; Acosta-Ampudia, Y.; Ramírez-Santana, C.; Leung, P.S.C.; Ansari, A.A.; Gershwin, M.E.; Anaya, J.M. Molecular mimicry and autoimmunity. J. Autoimmun. 2018, 95, 100–123. [Google Scholar] [CrossRef]
- Normatov, M.G.; Karev, V.E.; Kolobov, A.V.; Mayevskaya, V.A.; Ryabkova, V.A.; Utekhin, V.J.; Churilov, L.P. Post-COVID Endocrine Disorders: Putative Role of Molecular Mimicry and Some Pathomorphological Correlates. Diagnostics 2023, 13, 522. [Google Scholar] [CrossRef]
- Carnevale, S.; Giovanetti, M.; Benvenuto, D.; Ciccozzi, M.; Broccolo, F. Is Molecular Mimicry between hPF4 and SARS-CoV-2 Spike Protein a Potential Basis for Autoimmune Responses in Vaccinated and Naturally Infected Patients? Semin. Thromb. Hemost. 2023, 49, 103–104. [Google Scholar] [CrossRef] [PubMed]
- Cornaby, C.; Gibbons, L.; Mayhew, V.; Sloan, C.S.; Welling, A.; Poole, B.D. B cell epitope spreading: Mechanisms and contribution to autoimmune diseases. Immunol. Lett. 2015, 163, 56–68. [Google Scholar] [CrossRef] [PubMed]
- Farris, A.D.; Keech, C.L.; Gordon, T.P.; McCluskey, J. Epitope mimics and determinant spreading: Pathways to autoimmunity. Cell. Mol. Life Sci. 2000, 57, 569–578. [Google Scholar] [CrossRef]
- Pacheco, Y.; Acosta-Ampudia, Y.; Monsalve, D.M.; Chang, C.; Gershwin, M.E.; Anaya, J.M. Bystander activation and autoimmunity. J. Autoimmun. 2019, 103, 102301. [Google Scholar] [CrossRef] [PubMed]
- Okada, M.; Zhang, V.; Loaiza Naranjo, J.D.; Tillett, B.J.; Wong, F.S.; Steptoe, R.J.; Bergot, A.S.; Hamilton-Williams, E.E. Islet-specific CD8+ T cells gain effector function in the gut lymphoid tissues via bystander activation not molecular mimicry. Immunol. Cell Biol. 2023, 101, 36–48. [Google Scholar] [CrossRef]
- Engelhardt, B.; Vajkoczy, P.; Weller, R.O. The movers and shapers in immune privilege of the CNS. Nat. Immunol. 2017, 18, 123–131. [Google Scholar] [CrossRef]
- Hamdy, A.; Leonardi, A. Superantigens and SARS-CoV-2. Pathogens 2022, 11, 390. [Google Scholar] [CrossRef]
- Vojdani, A.; Kharrazian, D. Potential antigenic cross-reactivity between SARS-CoV-2 and human tissue with a possible link to an increase in autoimmune diseases. Clin. Immunol. 2020, 217, 108480. [Google Scholar] [CrossRef]
- Vojdani, A.; Vojdani, E.; Kharrazian, D. Reaction of Human Monoclonal Antibodies to SARS-CoV-2 Proteins with Tissue Antigens: Implications for Autoimmune Diseases. Front. Immunol. 2021, 11, 617089. [Google Scholar] [CrossRef]
- Vojdani, A.; Monro, J.; Lanzisera, F.; Sadeghi, H. Serological cross-reactivity between viruses and their contribution to autoimmunity. Autoimmun. Rev. 2021, 20, 102840. [Google Scholar] [CrossRef]
- Vojdani, A.; Lerner, A.; Vojdani, E. Cross-Reactivity and Sequence Homology between Al-Pha-Synuclein and Food Products: A Step Further for Parkinson’s Disease Synucleinopathy. Cells 2021, 10, 1111. [Google Scholar] [CrossRef]
- Vojdani, A.; Rosenberg, A.; Vojdani, E.; Shoenfeld, Y. Molecular and immunological evidence for SARS-CoV-2 being the autoimmune virus. In Autoimmunity, COVID-19, Post-COVID19 Syndrome and COVID-19 Vaccination; Shoenfeld, Y., Dotan, A., Eds.; Elsevier: Tel Aviv, Israel, 2023; Volume 1, pp. 339–350. [Google Scholar]
- Kanduc, D.; Shoenfeld, Y. Molecular mimicry between SARS-CoV-2 spike glycoprotein and mammalian proteomes: Implications for the vaccine. Immunol. Res. 2020, 68, 310–313. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Martin, M.J.; Orchard, S.; Magrane, M.; Agivetova, R.; Ahmad, S.; Alpi, E.; Bowler-Barnett, E.H.; Britto, R.; Bursteinas, B.; et al. UniProt: The universal protein knowledgebase in 2021. Nucleic Acids Res. 2021, 49, D480–D489. [Google Scholar] [CrossRef]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef] [PubMed]
- Kanduc, D. Pentapeptides as minimal functional units in cell biology and immunology. Curr. Protein Pept. Sci. 2013, 14, 111–120. [Google Scholar] [CrossRef]
- Kanduc, D. Homology, similarity, and identity in peptide epitope immunodefinition. J. Pept. Sci. 2012, 18, 487–494. [Google Scholar] [CrossRef]
- Kanduc, D. Immunogenicity, Immunopathogenicity, and Immunotolerance in One Graph. Anticancer. Agents Med. Chem. 2015, 15, 1264–1268. [Google Scholar] [CrossRef]
- Lanz, T.V.; Brewer, R.C.; Ho, P.P.; Moon, J.S.; Jude, K.M.; Fernandez, D.; Fernandes, R.A.; Gomez, A.M.; Nadj, G.S.; Bartley, C.M.; et al. Clonally expanded B cells in multiple sclerosis bind EBV EBNA1 and GlialCAM. Nature 2022, 603, 321–327. [Google Scholar] [CrossRef]
- Truong, H.V.; Sgourakis, N.G. Dynamics of MHC-I molecules in the antigen processing and presentation pathway. Curr. Opin. Immunol. 2021, 70, 122–128. [Google Scholar] [CrossRef]
- Yoshimoto, F.K. The Proteins of Severe Acute Respiratory Syndrome Coronavirus-2 (SARS CoV-2 or n-COV19), the Cause of COVID-19. Protein J. 2020, 39, 198–216. [Google Scholar] [CrossRef]
- Christen, U.; Holdener, M.; Hintermann, E. Cytochrome P450 2D6 as a Model Antigen. Dig. Dis. 2010, 28, 80–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutti, S.; Rigamonti, C.; Vidali, M.; Albano, E. CYP2E1 autoantibodies in liver diseases. Redox Biol. 2014, 3, 72–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christen, U.; Hintermann, E. Autoantibodies in Autoimmune Hepatitis: Can Epitopes Tell Us about the Etiology of the Disease? Front. Immunol. 2018, 9, 163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manns, M.P.; Obermayer-Straub, P. Cytochromes P450 and uridine triphosphate-glucuronosyltransferases: Model autoantigens to study drug-induced, virus-induced, and autoimmune liver disease. Hepatology 1997, 26, 1054–1066. [Google Scholar] [CrossRef]
- Jang, K.; Yoon, H.; Lee, J.; Yi, H.; Park, S.; Lee, S.; Lim, Y.; Lee, H.; Cho, J.; Paik, Y.; et al. Liver disease–associated keratin 8 and 18 mutations modulate keratin acetylation and methylation. FASEB J. 2019, 33, 9030–9043. [Google Scholar] [CrossRef] [Green Version]
- Ku, N.-O.; Darling, J.M.; Krams, S.M.; Esquivel, C.O.; Keeffe, E.B.; Sibley, R.K.; Lee, Y.M.; Wright, T.L.; Omary, M.B. Keratin 8 and 18 mutations are risk factors for developing liver disease of multiple etiologies. Proc. Natl. Acad. Sci. USA 2003, 100, 6063–6068. [Google Scholar] [CrossRef]
- McFarlane, I.G. The relationship between autoimmune markers and different clinical syndromes in autoimmune hepatitis. Gut 1998, 42, 599–602. [Google Scholar] [CrossRef]
- Von Mühlen, C.A.; Chan, E.K.L.; Peebles, C.L.; Imai, H.; Kiyosawa, K.; Tan, E.M. Non-muscle myosin as target antigen for human autoantibodies in patients with hepatitis C virus-associated chronic liver diseases. Clin. Exp. Immunol. 2008, 100, 67–74. [Google Scholar] [CrossRef]
- Zamanou, A.; Samiotaki, M.; Panayotou, G.; Margaritis, L.; Lymberi, P. Fine specificity and subclasses of IgG anti-actin autoantibodies differ in health and disease. J. Autoimmun. 2003, 20, 333–344. [Google Scholar] [CrossRef]
- Mori, T.; Ono, K.; Hakozaki, M.; Kochi, H. Epitope mapping on E1α subunit of pyruvate dehydrogenase complex with autoantibodies of patients with primary biliary cirrhosis. Liver Int. 2003, 23, 355–362. [Google Scholar] [CrossRef]
- Fregeau, D.R.; Roche, T.E.; Davis, P.A.; Coppel, R.; Gershwin, M.E. Primary biliary cirrhosis. Inhibition of pyruvate dehydrogenase complex activity by autoantibodies specific for E1 alpha, a non-lipoic acid containing mitochondrial enzyme. J. Immunol. 1990, 144, 1671–1676. [Google Scholar] [CrossRef] [PubMed]
- Jones, D.E.J. Pathogenesis of primary biliary cirrhosis. Postgrad. Med. J. 2008, 84, 23–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, P.S.C.; Choi, J.; Yang, G.; Woo, E.; Kenny, T.P.; Gershwin, M.E. A contemporary perspective on the molecular characteristics of mitochondrial autoantigens and diagnosis in primary biliary cholangitis. Expert Rev. Mol. Diagn. 2016, 16, 697–705. [Google Scholar] [CrossRef] [Green Version]
- Eggena, M.; Cohavy, O.; Parseghian, M.H.; Hamkalo, B.A.; Clemens, D.; Targan, S.R.; Gordon, L.K.; Braun, J. Identification of histone H1 as a cognate antigen of the ulcerative colitis-associated marker antibody pANCA. J. Autoimmun. 2000, 14, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Bogdanos, D.P.; Hussain, M.J.; Underhill, J.; Bansal, S.; Longhi, M.S.; Cheeseman, P.; Mieli-Vergani, G.; Vergani, D. Polyclonal T-cell responses to cytochrome P450IID6 are associated with disease activity in autoimmune hepatitis type 2. Gastroenterology 2006, 130, 868–882. [Google Scholar] [CrossRef] [PubMed]
- Tarke, A.; Sidney, J.; Kidd, C.K.; Dan, J.M.; Ramirez, S.I.; Yu, E.D.; Mateus, J.; da Silva Antunes, R.; Moore, E.; Rubiro, P.; et al. Comprehensive analysis of T cell immunodominance and immunoprevalence of SARS-CoV-2 epitopes in COVID-19 cases. Cell Rep. Med. 2021, 2, 100204. [Google Scholar] [CrossRef]
- Schwarz, T.; Heiss, K.; Mahendran, Y.; Casilag, F.; Kurth, F.; Sander, L.E.; Wendtner, C.M.; Hoechstetter, M.A.; Müller, M.A.; Sekul, R.; et al. SARS-CoV-2 proteome-wide analysis revealed significant epitope signatures in COVID-19 Patients. Front. Immunol. 2021, 12, 629185. [Google Scholar] [CrossRef]
- Sarkizova, S.; Klaeger, S.; Le, P.M.; Li, L.W.; Oliveira, G.; Keshishian, H.; Hartigan, C.R.; Zhang, W.; Braun, D.A.; Ligon, K.L.; et al. A large peptidome dataset improves HLA class I epitope prediction across most of the human population. Nat. Biotechnol. 2020, 38, 199–209. [Google Scholar] [CrossRef]
- Yair-Sabag, S.; Tedeschi, V.; Vitulano, C.; Barnea, E.; Glaser, F.; Melamed Kadosh, D.; Taurog, J.D.; Fiorillo, M.T.; Sorrentino, R.; Admon, A. The peptide repertoire of HLA-B27 may include ligands with lysine at P2 anchor position. Proteomics 2018, 18, e1700249. [Google Scholar] [CrossRef]
- Liu, R.; Jiang, W.; Mellins, E.D. Yeast display of MHC-II enables rapid identification of peptide ligands from protein antigens (RIPPA). Cell. Mol. Immunol. 2021, 18, 1847–1860. [Google Scholar] [CrossRef]
- Mishra, N.; Huang, X.; Joshi, S.; Guo, C.; Ng, J.; Thakkar, R.; Wu, Y.; Dong, X.; Li, Q.; Pinapati, R.S.; et al. Immunoreactive peptide maps of SARS-CoV-2. Commun. Biol. 2021, 4, 1–4. [Google Scholar] [CrossRef]
- Weingarten-Gabbay, S.; Klaeger, S.; Sarkizova, S.; Pearlman, L.R.; Chen, D.Y.; Gallagher, K.M.E.; Bauer, M.R.; Taylor, H.B.; Dunn, W.A.; Tarr, C.; et al. Profiling SARS-CoV-2 HLA-I peptidome reveals T cell epitopes from out-of-frame ORFs. Cell 2021, 184, 3962–3980.e17. [Google Scholar] [CrossRef] [PubMed]
- Tarke, A.; Coelho, C.H.; Zhang, Z.; Dan, J.M.; Yu, E.D.; Methot, N.; Bloom, N.I.; Goodwin, B.; Phillips, E.; Mallal, S.; et al. SARS-CoV-2 vaccination induces immunological T cell memory able to cross-recognize variants from Alpha to Omicron. Cell 2022, 185, 847–859.e11. [Google Scholar] [CrossRef] [PubMed]
- Voss, C.; Esmail, S.; Liu, X.; Knauer, M.J.; Ackloo, S.; Kaneko, T.; Lowes, L.; Stogios, P.; Seitova, A.; Hutchinson, A.; et al. Epitope-specific antibody responses differentiate COVID-19 outcomes and variants of concern. JCI Insight 2021, 6, e148855. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Chen, Y.; Prévost, J.; Ullah, I.; Lu, M.; Gong, S.Y.; Tauzin, A.; Gasser, R.; Vézina, D.; Anand, S.P.; et al. Structural basis and mode of action for two broadly neutralizing antibodies against SARS-CoV-2 emerging variants of concern. Cell Rep. 2022, 38, 2. [Google Scholar] [CrossRef]
- Chen, L.; Shi, H.; Koftori, D.; Sekine, T.; Nicastri, A.; Ternette, N.; Bowness, P. Identification of an unconventional subpeptidome bound to the Behçet’s disease-associated HLA-B*51:01 that is regulated by endoplasmic reticulum aminopeptidase 1 (ERAP1). Mol. Cell. Proteom. 2020, 19, 871–883. [Google Scholar] [CrossRef] [Green Version]
- Heide, J.; Schulte, S.; Kohsar, M.; Brehm, T.T.; Herrmann, M.; Karsten, H.; Marget, M.; Peine, S.; Johansson, A.M.; Sette, A.; et al. Correction: Broadly directed SARS-CoV-2-specific CD4+ T cell response includes frequently detected peptide specificities within the membrane and nucleoprotein in patients with acute and resolved COVID-19. PLoS Pathog. 2022, 18, e1010220. [Google Scholar] [CrossRef]
- Hotop, S.K.; Reimering, S.; Shekhar, A.; Asgari, E.; Beutling, U.; Dahlke, C.; Fathi, A.; Khan, F.; Lütgehetmann, M.; Ballmann, R.; et al. Peptide microarrays coupled to machine learning reveal individual epitopes from human antibody responses with neutralizing capabilities against SARS-CoV-2. Emerg. Microbes Infect. 2022, 11, 1037–1048. [Google Scholar] [CrossRef]
- Neidert, M.C.; Kowalewski, D.J.; Silginer, M.; Kapolou, K.; Backert, L.; Freudenmann, L.K.; Peper, J.K.; Marcu, A.; Wang, S.S.Y.; Walz, J.S.; et al. The natural HLA ligandome of glioblastoma stem-like cells: Antigen discovery for T cell-based immunotherapy. Acta Neuropathol. 2018, 135, 923–938. [Google Scholar] [CrossRef] [Green Version]
- Jin, X.; Ding, Y.; Sun, S.; Wang, X.; Zhou, Z.; Liu, X.; Li, M.; Chen, X.; Shen, A.; Wu, Y.; et al. Screening HLA-A-restricted T cell epitopes of SARS-CoV-2 and the induction of CD8+ T cell responses in HLA-A transgenic mice. Cell. Mol. Immunol. 2021, 18, 2588–2608. [Google Scholar] [CrossRef]
- Nagler, A.; Kalaora, S.; Barbolin, C.; Gangaev, A.; Ketelaars, S.L.C.; Alon, M.; Pai, J.; Benedek, G.; Yahalom-Ronen, Y.; Erez, N.; et al. Identification of presented SARS-CoV-2 HLA class I and HLA class II peptides using HLA peptidomics. Cell Rep. 2021, 35, 109305. [Google Scholar] [CrossRef]
- Marcu, A.; Bichmann, L.; Kuchenbecker, L.; Kowalewski, D.J.; Freudenmann, L.K.; Backert, L.; Mühlenbruch, L.; Szolek, A.; Lübke, M.; Wagner, P.; et al. HLA Ligand Atlas: A benign reference of HLA-presented peptides to improve T-cell-based cancer immunotherapy. J. Immunother. Cancer 2021, 9, e002071. [Google Scholar] [CrossRef] [PubMed]
- Mateus, J.; Grifoni, A.; Tarke, A.; Sidney, J.; Ramirez, S.I.; Dan, J.M.; Burger, Z.C.; Rawlings, S.A.; Smith, D.M.; Phillips, E.; et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science 2020, 370, 89–94. [Google Scholar] [CrossRef]
- Knierman, M.D.; Lannan, M.B.; Spindler, L.J.; McMillian, C.L.; Konrad, R.J.; Siegel, R.W. The human leukocyte antigen class II immunopeptidome of the SARS-CoV-2 spike glycoprotein. Cell Rep. 2020, 33, 108454. [Google Scholar] [CrossRef]
- Mifsud, N.A.; Illing, P.T.; Lai, J.W.; Fettke, H.; Hensen, L.; Huang, Z.; Rossjohn, J.; Vivian, J.P.; Kwan, P.; Purcell, A.W. Carbamazepine induces focused T Cell responses in resolved Stevens-Johnson syndrome and toxic epidermal necrolysis cases but does not perturb the immunopeptidome for T Cell recognition. Front. Immunol. 2021, 12, 653710. [Google Scholar] [CrossRef]
- Faridi, P.; Woods, K.; Ostrouska, S.; Deceneux, C.; Aranha, R.; Duscharla, D.; Wong, S.Q.; Chen, W.; Ramarathinam, S.H.; Lim Kam Sian, T.C.C.; et al. Spliced peptides and cytokine-driven changes in the immunopeptidome of melanoma. Cancer Immunol. Res. 2020, 8, 1322–1334. [Google Scholar] [CrossRef]
- Chen, L.; Pang, P.; Qi, H.; Yan, K.; Ren, Y.; Ma, M.; Cao, R.; Li, H.; Hu, C.; Li, Y.; et al. Evaluation of spike protein epitopes by assessing the dynamics of humoral immune responses in moderate COVID-19. Front. Immunol. 2022, 13, 770982. [Google Scholar] [CrossRef] [PubMed]
- Ramarathinam, S.H.; Ho, B.K.; Dudek, N.L.; Purcell, A.W. HLA class II immunopeptidomics reveals that co-inherited HLA-allotypes within an extended haplotype can improve proteome coverage for immunosurveillance. Proteomics 2021, 21, e2000160. [Google Scholar] [CrossRef] [PubMed]
- Braun, S.; Berg, C.; Buck, S.; Gregor, M.; Klein, R. Catalytic domain of PDC-E2 contains epitopes recognized by antimitochondrial antibodies in primary biliary cirrhosis. World J. Gastroenterol. 2010, 16, 973–981. [Google Scholar] [CrossRef]
- Low, J.S.; Vaqueirinho, D.; Mele, F.; Foglierini, M.; Jerak, J.; Perotti, M.; Jarrossay, D.; Jovic, S.; Perez, L.; Cacciatore, R.; et al. Clonal analysis of immunodominance and cross-reactivity of the CD4 T cell response to SARS-CoV-2. Science 2021, 372, 1336–1341. [Google Scholar] [CrossRef]
- Polyiam, K.; Phoolcharoen, W.; Butkhot, N.; Srisaowakarn, C.; Thitithanyanont, A.; Auewarakul, P.; Hoonsuwan, T.; Ruengjitchatchawalya, M.; Mekvichitsaeng, P.; Roshorm, Y.M. Immunodominant linear B cell epitopes in the spike and membrane proteins of SARS-CoV-2 identified by immunoinformatics prediction and immunoassay. Sci. Rep. 2021, 11, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Holenya, P.; Lange, P.J.; Reimer, U.; Woltersdorf, W.; Panterodt, T.; Glas, M.; Wasner, M.; Eckey, M.; Drosch, M.; Hollidt, J.M.; et al. Peptide microarray-based analysis of antibody responses to SARS-CoV-2 identifies unique epitopes with potential for diagnostic test development. Eur. J. Immunol. 2021, 51, 1839–1849. [Google Scholar] [CrossRef] [PubMed]
- Kaabinejadian, S.; Barra, C.; Alvarez, B.; Yari, H.; Hildebrand, W.H.; Nielsen, M. Accurate MHC motif deconvolution of immunopeptidomics data reveals a significant contribution of DRB3, 4 and 5 to the total DR immunopeptidome. Front. Immunol. 2022, 13, 835454. [Google Scholar] [CrossRef] [PubMed]
- Cerrada-Romero, C.; Berastegui-Cabrera, J.; Camacho-Martínez, P.; Goikoetxea-Aguirre, J.; Pérez-Palacios, P.; Santibáñez, S.; José Blanco-Vidal, M.; Valiente, A.; Alba, J.; Rodríguez-Álvarez, R.; et al. Excretion and viability of SARS-CoV-2 in feces and its association with the clinical outcome of COVID-19. Sci. Rep. 2022, 12, 7397. [Google Scholar] [CrossRef]
- Lavania, M.; Joshi, M.S.; Ranshing, S.S.; Potdar, V.A.; Shinde, M.; Chavan, N.; Jadhav, S.M.; Sarkale, P.; Mohandas, S.; Sawant, P.M.; et al. Prolonged shedding of SARS-CoV-2 in feces of COVID-19 positive patients: Trends in genomic variation in first and second wave. Front. Med. 2022, 9, 835168. [Google Scholar] [CrossRef]
- Holm-Jacobsen, J.N.; Bundgaard-Nielsen, C.; Rold, L.S.; Jensen, A.M.; Shakar, S.; Ludwig, M.; Kirk, K.F.; Donneborg, M.L.; Vonasek, J.H.; Pedersen, B.; et al. The prevalence and clinical implications of rectal SARS-CoV-2 shedding in Danish COVID-19 patients and the general population. Front. Med. 2022, 8, 804804. [Google Scholar] [CrossRef]
- Maan, H.S.; Chaurasia, D.; Kapoor, G.; Dave, L.; Siddiqui, A.; Pal, S.; Singh, H.O.; Biswas, D.; Chowdhary, R. Intestinal viral infections of nSARS-CoV2 in the Indian community: Risk of virus spread in India. J. Med. Virol. 2022, 94, 1315–1329. [Google Scholar] [CrossRef]
- Jafarzadeh, A.; Chauhan, P.; Saha, B.; Jafarzadeh, S.; Nemati, M. Contribution of monocytes and macrophages to the local tissue inflammation and cytokine storm in COVID-19: Lessons from SARS and MERS, and potential therapeutic interventions. Life Sci. 2020, 257, 118102. [Google Scholar] [CrossRef]
- Lyons-Weiler, J. Pathogenic priming likely contributes to serious and critical illness and mortality in COVID-19 via autoimmunity. J. Transl. Autoimmun. 2020, 3, 100051. [Google Scholar] [CrossRef]
- Nunez-Castilla, J.; Stebliankin, V.; Baral, P.; Balbin, C.A.; Sobhan, M.; Cickovski, T.; Mondal, A.M.; Narasimhan, G.; Chapagain, P.; Mathee, K.; et al. Potential autoimmunity resulting from molecular mimicry between SARS-CoV-2 spike and human proteins. Viruses 2022, 14, 1415. [Google Scholar] [CrossRef]
- Oldstone, M.B.A. Molecular mimicry and immune-mediated diseases. FASEB J. 1998, 12, 1255–1265. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Ballester, M.P.; Soffientini, U.; Jalan, R.; Mehta, G. SARS-CoV-2 infection and liver involvement. Hepatol. Int. 2022, 16, 755–774. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.T.; Yu, Y.; Zhang, C.; Wang, H.; Zhao, L.; Wang, H.; Su, Y.; Yang, M. Mechanism of SARS-CoV-2 invasion into the liver and hepatic injury in patients with COVID-19. Mediterr. J. Hematol. Infect. Dis. 2022, 14, e2022003. [Google Scholar] [CrossRef]
- Ekpanyapong, S.; Bunchorntavakul, C.; Reddy, K.R. COVID-19 and the Liver: Lessons learnt from the EAST and the WEST, a year later. J. Viral Hepat. 2022, 29, 4–20. [Google Scholar] [CrossRef] [PubMed]
- Floreani, A.; De Martin, S. COVID-19 and autoimmune liver diseases. J. Clin. Med. 2022, 11, 2681. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Domínguez, L.; Rodríguez, Y.; Polo, F.; Restrepo Gutierrez, J.C.; Zapata, E.; Rojas, M.; Anaya, J.M. COVID-19 vaccine and autoimmunity. A new case of autoimmune hepatitis and review of the literature. J. Transl. Autoimmun. 2022, 5, 100140. [Google Scholar] [CrossRef] [PubMed]
- Pruthvishree, B.S.; Vinodh Kumar, O.R.; Sinha, D.K.; Malik, Y.P.S.; Dubal, Z.B.; Desingu, P.A.; Shivakumar, M.; Krishnaswamy, N.; Singh, B.R. Spatial molecular epidemiology of carbapenem-resistant and New Delhi metallo beta-lactamase (blaNDM)-producing Escherichia coli in the piglets of organized farms in India. J. Appl. Microbiol. 2017, 122, 1537–1546. [Google Scholar] [CrossRef]
- Ampuero, J.; Lucena, A.; Hernández-Guerra, M.; Moreno-Moraleda, I.; Arenas, J.; Conde, I.; Muñoz, L.; Canillas, L.; Fernandez, E.; Quiñones, R.; et al. Primary biliary cholangitis and SARS-CoV-2 infection: Incidence, susceptibility and outcomes. Gut 2021, 71, 2138–2140. [Google Scholar] [CrossRef]
- Cirillo, C.; Sarnelli, G.; Esposito, G.; Turco, F.; Steardo, L.; Cuomo, R. S100B protein in the gut: The evidence for enteroglial-sustained intestinal inflammation. World J. Gastroenterol. 2011, 17, 1261–1266. [Google Scholar] [CrossRef]
- Lerner, A.; Neidhöfer, S.; Matthias, T. The gut microbiome feelings of the brain: A perspective for non-microbiologists. Microorganisms 2017, 5, 66. [Google Scholar] [CrossRef]
- Obrenovich, M. Leaky gut, leaky brain? Microorganisms 2018, 6, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerner, A.; Matthias, T. GUT-the Trojan Horse in remote organs’ Autoimmunity. J. Clin. Cell. Immunol. 2016, 7, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Izcovich, A.; Ragusa, M.A.; Tortosa, F.; Marzio, M.A.L.; Agnoletti, C.; Bengolea, A.; Ceirano, A.; Espinosa, F.; Saavedra, E.; Sanguine, V.; et al. Prognostic factors for severity and mortality in patients infected with COVID-19: A systematic review. PLoS ONE 2020, 15, e0241955. [Google Scholar] [CrossRef]
- Chen, J.; Vitetta, L. The gut-liver axis in chronic liver disease associated with severe COVID-19. Eur. J. Gastroenterol. Hepatol. 2021, 33, e1103. [Google Scholar] [CrossRef] [PubMed]
- Prasad, R.; Patton, M.J.; Floyd, J.L.; Fortmann, S.; DuPont, M.; Harbour, A.; Wright, J.; Lamendella, R.; Stevens, B.R.; Oudit, G.Y.; et al. Plasma Microbiome in COVID-19 Subjects: An Indicator of Gut Barrier Defects and Dysbiosis. Int. J. Mol. Sci. 2022, 23, 9141. [Google Scholar] [CrossRef] [PubMed]
- Zachou, K.; Rigopoulou, E.; Dalekos, G.N. Autoantibodies and autoantigens in autoimmune hepatitis: Important tools in clinical practice and to study pathogenesis of the disease. J. Autoimmune Dis. 2004, 1, 2. [Google Scholar] [CrossRef] [Green Version]
- Paolella, G.; Sposito, S.; Romanelli, A.M.; Caputo, I. Type 2 Transglutaminase in Coeliac Disease: A Key Player in Pathogenesis, Diagnosis and Therapy. Int. J. Mol. Sci. 2022, 23, 7513. [Google Scholar] [CrossRef]
- Rauhavirta, T.; Hietikko, M.; Salmi, T.; Lindfors, K. Transglutaminase 2 and Transglutaminase 2 Autoantibodies in Celiac Disease: A Review. Clin. Rev. Allergy Immunol. 2019, 57, 23–38. [Google Scholar] [CrossRef]
- Iannaccone, M.; Titta, F.; Serretiello, E.; Monfregola, M.; Gentile, V. Possible physiopathological effects of the transglutaminase activity on the molecular mechanisms responsible for human neurodegenerative diseases. Recent Pat. CNS Drug Discov. 2014, 9, 76–84. [Google Scholar] [CrossRef]
- Smatti, M.K.; Cyprian, F.S.; Nasrallah, G.K.; Al Thani, A.A.; Almishal, R.O.; Yassine, H.M. Viruses and Autoimmunity: A Review on the Potential Interaction and Molecular Mechanisms. Viruses 2019, 11, 762. [Google Scholar] [CrossRef] [Green Version]
- Moreno Fernández-Ayala, D.J.; Navas, P.; López-Lluch, G. Age-related mitochondrial dysfunction as a key factor in COVID-19 disease. Exp. Gerontol. 2020, 142, 111147. [Google Scholar] [CrossRef] [PubMed]
- Tereshin, A.E.; Kiryanova, V.V.; Reshetnik, D.A. Correction of Mitochondrial Dysfunction in the Complex Rehabilitation of COVID-19 Patients. Neurosci. Behav. Physiol. 2022, 52, 511–514. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.J.; Cullen, S. Etiopathogenesis of primary biliary cirrhosis: An overview of recent developments. Hepatol. Int. 2013, 7, 28–47. [Google Scholar] [CrossRef] [PubMed]
- Bril, F.; Al Diffalha, S.; Dean, M.; Fettig, D.M. Autoimmune hepatitis developing after coronavirus disease 2019 (COVID-19) vaccine: Causality or casualty? J. Hepatol. 2021, 75, 222–224. [Google Scholar] [CrossRef]
- Kordopati, G.G.; Tzoupis, H.; Troganis, A.N.; Tsivgoulis, G.M.; Golic Grdadolnik, S.; Simal, C.; Tselios, T.V. Biologically relevant conformational features of linear and cyclic proteolipid protein (PLP) peptide analogues obtained by high-resolution nuclear magnetic resonance and molecular dynamics. J. Comput.-Aided Mol. Des. 2017, 31, 841–854. [Google Scholar] [CrossRef]
- Kanaan, Z.; Qadan, M.; Eichenberger, M.R.; Galandiuk, S. The actin-cytoskeleton pathway and its potential role in inflammatory bowel disease-associated human colorectal cancer. Genet. Test. Mol. Biomark. 2010, 14, 347–353. [Google Scholar] [CrossRef]
- Puga Yung, G.L.; Fidler, M.; Albani, E.; Spermon, N.; Teklenburg, G.; Newbury, R.; Schechter, N.; van den Broek, T.; Prakken, B.; Billetta, R.; et al. Heat shock protein-derived T-cell epitopes contribute to autoimmune inflammation in pediatric Crohn’s disease. PLoS ONE 2009, 4, e7714. [Google Scholar] [CrossRef]
- Vuille-Lessard, É.; Montani, M.; Bosch, J.; Semmo, N. Autoimmune hepatitis triggered by SARS-CoV-2 vaccination. J. Autoimmun. 2021, 123, 102710. [Google Scholar] [CrossRef]
- Awaya, T.; Moroi, M.; Enomoto, Y.; Kunimasa, T.; Nakamura, M. What Should We Do after the COVID-19 Vaccination? Vaccine-Associated Diseases and Precautionary Measures against Adverse Reactions. Vaccines 2022, 10, 866. [Google Scholar] [CrossRef]
- Reche, P.A. Potential cross-reactive immunity to SARS-CoV-2 from common human pathogens and vaccines. Front. Immunol. 2020, 11, 586984. [Google Scholar] [CrossRef]
- Vojdani, A.; Vojdani, E.; Melgar, A.L.; Redd, J. Reaction of SARS-CoV-2 antibodies with other pathogens, vaccines, and food antigens. Front. Immunol. 2022, 13, 5254. [Google Scholar] [CrossRef] [PubMed]
- Perricone, C.; Agmon-Levin, N.; Shoenfeld, Y. Novel pebbles in the mosaic of autoimmunity. BMC Med. 2013, 11, 101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahroum, N.; Elsalti, A.; Alwani, A.; Seida, I.; Alrais, M.; Seida, R.; Nil, S.; Abali, T.; Kiyak, Z.; Zoubi, M. Autoimmunity Reviews The mosaic of autoimmunity—Finally discussing in person. The 13th international congress on autoimmunity 2022 (AUTO13) Athens. Autoimmun. Rev. 2022, 21, 103166. [Google Scholar] [CrossRef] [PubMed]
- Vita, R.; Mahajan, S.; Overton, J.A.; Dhanda, S.K.; Martini, S.; Cantrell, J.R.; Wheeler, D.K.; Sette, A.; Peters, B. The immune epitope database (IEDB): 2018 update. Nucleic Acids Res. 2019, 47, D339–D343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleri, W.; Paul, S.; Dhanda, S.K.; Mahajan, S.; Xu, X.; Peters, B.; Sette, A. The immune epitope database and analysis resource in epitope discovery and synthetic vaccine design. Front. Immunol. 2017, 8, 278. [Google Scholar] [CrossRef] [Green Version]
- Rice, P.; Longden, L.; Bleasby, A. EMBOSS: The European Molecular Biology Open Software Suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Human Protein (Autoimmune-Related Disease) 1 | SARS-CoV-2 Protein | Human vs. SARS-CoV-2 | Ln | Identity % | Similarity % | Score 2 |
|---|---|---|---|---|---|---|
| Cytochrome P450 2D6 (AIH, CD) | Membrane glycoprotein | KKSLEQW | 7 | 85.7 | 85.7 | 33 |
| KKLLEQW | ||||||
| Keratin, type I cytoskeletal 18 (AIH) | Spike glycoprotein | VSETNDTK | 8 | 75 | 75 | 26 |
| VSGTNGTK | ||||||
| Spike glycoprotein | EELDKYW | 7 | 85.7 | 100 | 33 | |
| EELDKYF | ||||||
| nucleocapsid phosphoprotein | NARIVLQI | 8 | 75 | 87.5 | 28 | |
| NAAIVLQL | ||||||
| Myosin-11 (AIH) | Envelope small membrane protein | VKNDNSSR | 8 | 87.5 | 87.5 | 30 |
| VKNLNSSR | ||||||
| Actin, alpha skeletal muscle (AIH) | Spike glycoprotein | GDGVTHNV | 8 | 75 | 75 | 28 |
| GIGVTQNV | ||||||
| Pyruvate dehydrogenase E1 component subunit alpha, somatic form, mitochondrial (PBC) | Spike glycoprotein | ATRFAAAY | 8 | 75 | 87.5 | 32 |
| ATRFASVY | ||||||
| Dihydrolipoyllysine-residue acetyltransferase component of pyruvate dehydrogenase complex, mitochondrial (PBC) | Spike glycoprotein | PATPAGPK | 8 | 75 | 75 | 32 |
| PATVCGPK | ||||||
| Spike glycoprotein | DVPIGAIIC | 9 | 77.8 | 88.9 | 39 | |
| DIPIGAGIC | ||||||
| Histone H1.0 (UC) | Nucleocapsid phosphoprotein | PKKAKKPK | 8 | 75 | 75 | 29 |
| PKKDKKKK |
| Similar Peptides | Enteric Protein and Potential Function/Pathogenesis | AD 1 | Ref. |
|---|---|---|---|
| KKSLEQW | Cytochrome P450 2D6 (CYP2D6, UniProt:P10635) Has been recognized as the major autoantigen in type 2 AIH. In patients with AIH-2, the target for anti-LKM-1 antibodies has been identified as the 2D6 isoform of the large cytochrome P450 enzyme family. | AIH CD | [62,63,64,65] |
| VSETNDTK EELDKYW NARIVLQI | Keratin, type I cytoskeletal 18 (Keratin-18, UniProt:P05783) Keratin 18 (K18) is an intermediate filament protein whose phosphorylation/transamidation associates with its aggregation in Mallory-Denk bodies found in patients with various liver diseases: cirrhosis, PBC, and AIH. | AIH PBC | [66,67] |
| VKNDNSSR | Myosin-11 (SMMHC, UniProt:P35749) Autoantibodies to non-muscle myosin heavy chain were reported in patients with chronic liver diseases. Patients presenting with ANA and/or smooth muscle antibodies (SMA) account for about 80% of cases of AIH. | AIH | [68,69] |
| GDGVTHNV | Actin, alpha skeletal muscle (Alpha-actin-1, UniProt:P68133) Anti-actin antibodies were the first anti-cytoskeleton autoantibodies described in liver diseases. They are considered a marker of AIH-1 and are also frequently detected in PBC. | AIH PBC | [70] |
| ATRFAAAY | Pyruvate dehydrogenase E1 component subunit alpha, somatic form, mitochondrial (PDH-E1α, UniProt:P08559) Autoantibodies against multiple antigens of the PDH complex are found in sera of more than 90% of patients with PBC, in particular against the PDH-E1α subunit. Anti-mitochondrial autoantibodies (AMA) appear to be directed to a functional site of PDC-E1α inasmuch as they are able to inhibit enzyme function. | PBC | [71,72] |
| PATPAGPKDVPIGAIIC | Dihydrolipoyllysine-residue acetyltransferase component of pyruvate dehydrogenase complex, mitochondrial (PDC-E2, UniProt:P10515) PBC patients have been characterized to have autoreactive T-cell and B-cell responses directed at self-PDC-E2. The diagnosis of PBC is readily reached by the detection of specific AMA directed against PDH-E2. | PBC | [73,74] |
| PKKAKKPK | Histone H1.0 (UniProt:P07305) Histone H1 bears a recurring COOH-terminal epitope recognized by monoclonal ulcerative colitis-associated pANCA marker antibodies. | IBD | [75] |
| Human Epitope | SARS-CoV-2 Epitope | Human vs. SARS-CoV-2 | IEDB Human Assays’ References | IEDB SARS-CoV-2 Assays’ References |
|---|---|---|---|---|
| STLRNLGLGKKSLEQWVTEE | EELKKLLEQWNLVIG | KKSLEQW | Tcell [76] | Tcell [77]; Bcell(IgA) [78] |
| KKLLEQW | ||||
| VVSETNDTK | WFHAIHVSGTNGTKRFD | VSETNDTK | HLA-C*06:02 [79]; HLA-B*27:09 [80] | HLA-I/II [81]; Tcell [77]; Bcell(IgM) [82] |
| VSGTNGTK | ||||
| EELDKYWSQ | DSFKEELDKYFKNHT | EELDKYW | HLA-I [83] | Tcell [84]; Bcell(IgG) [85]; HLA-DRA*01:01/DRB1*04:01 [81]; Bcell(IgG1) [86] |
| EELDKYF | ||||
| NARIVLQI | PANNAAIVLQLPQGT | NARIVLQI | HLA-B*51:01 [87] | Tcell [88]; Bcell(IgM) [89] |
| NAAIVLQL | ||||
| NAKTVKNDNSSRFG | RVKNLNSSR | VKNDNSSR | HLA-II [90] | HLA-A*30:01; Tcell [91]; HLA-A*01:01 [92]; Bcell(IgM) [89] |
| VKNLNSSR | ||||
| SGDGVTHNVPI | QMAYRFNGIGVTQNV | GDGVTHNV | HLA-II [93] | Tcell [94]; Bcell(IgM) [89]; HLA-II [95] |
| GIGVTQNV | ||||
| EATRFAAAY | ATRFASVYA | ATRFAAAY | HLA-B*15:02 [96]; HLA-B*44:02 [97] | HLA-A*30:01; Tcell [91]; HLA-II [95]; Bcell(IgG) [98] |
| ATRFASVY | ||||
| VPPTPQPLAPTPSAPCPATPAGPK | VLSFELLHAPATVCGPK | PATPAGPK | HLA-DQ [99]; Bcell(IgG) [100] | HLA-DRA*01:01/DRB1*04:01 [81]; Tcell [101]; Bcell(IgG) [102] |
| PATVCGPK | ||||
| GTRDVPIGAIICITVGKPEDIEAFK | SYECDIPIGAGICASYQ | DVPIGAIIC | Bcell(IgG) [100] | HLA-I/II [81]; Tcell [94]; Bcell(IgG) [103] |
| DIPIGAGIC | ||||
| AATPKKAKKPKT | TEPKKDKKKKADETQ | PKKAKKPK | HLA-II [93]; HLA-DR [104]; HLA-DRB1*11:03 [93] | Tcell [88]; Bcell(IgG) [85]; Bcell(IgM) [82] |
| PKKDKKKK |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lerner, A.; Benzvi, C.; Vojdani, A. SARS-CoV-2 Gut-Targeted Epitopes: Sequence Similarity and Cross-Reactivity Join Together for Molecular Mimicry. Biomedicines 2023, 11, 1937. https://doi.org/10.3390/biomedicines11071937
Lerner A, Benzvi C, Vojdani A. SARS-CoV-2 Gut-Targeted Epitopes: Sequence Similarity and Cross-Reactivity Join Together for Molecular Mimicry. Biomedicines. 2023; 11(7):1937. https://doi.org/10.3390/biomedicines11071937
Chicago/Turabian StyleLerner, Aaron, Carina Benzvi, and Aristo Vojdani. 2023. "SARS-CoV-2 Gut-Targeted Epitopes: Sequence Similarity and Cross-Reactivity Join Together for Molecular Mimicry" Biomedicines 11, no. 7: 1937. https://doi.org/10.3390/biomedicines11071937
APA StyleLerner, A., Benzvi, C., & Vojdani, A. (2023). SARS-CoV-2 Gut-Targeted Epitopes: Sequence Similarity and Cross-Reactivity Join Together for Molecular Mimicry. Biomedicines, 11(7), 1937. https://doi.org/10.3390/biomedicines11071937