
Bronchial Fibroblasts from Asthmatic Patients Display Impaired Responsiveness to Direct Current Electric Fields (dcEFs)
 ,
,  ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Application of Electric Field
2.3. Registration of Cell Migration and Morphology
2.4. Analysis of Cell Migration
2.5. Analysis of Cell Orientation
2.6. Inhibition of Signalling Pathways
2.7. Induction of the Fibroblast-to-Myofibroblast Transition (FMT)
2.8. Immunofluorescent Staining and Fluorescence Microscopy
2.9. Statistical Analysis
3. Results
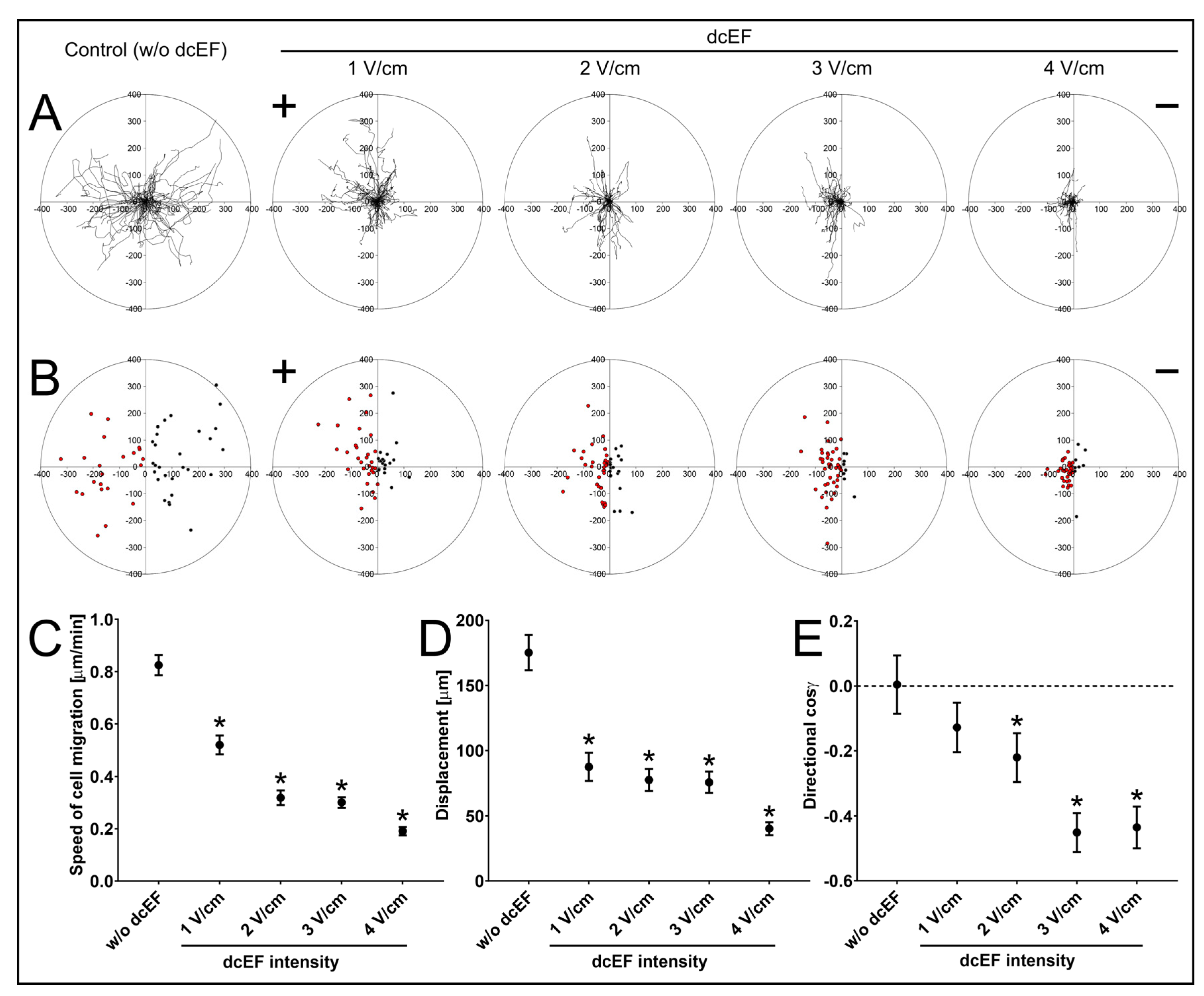
3.1. Electrotactic Responses of NA HBF to dcEFs
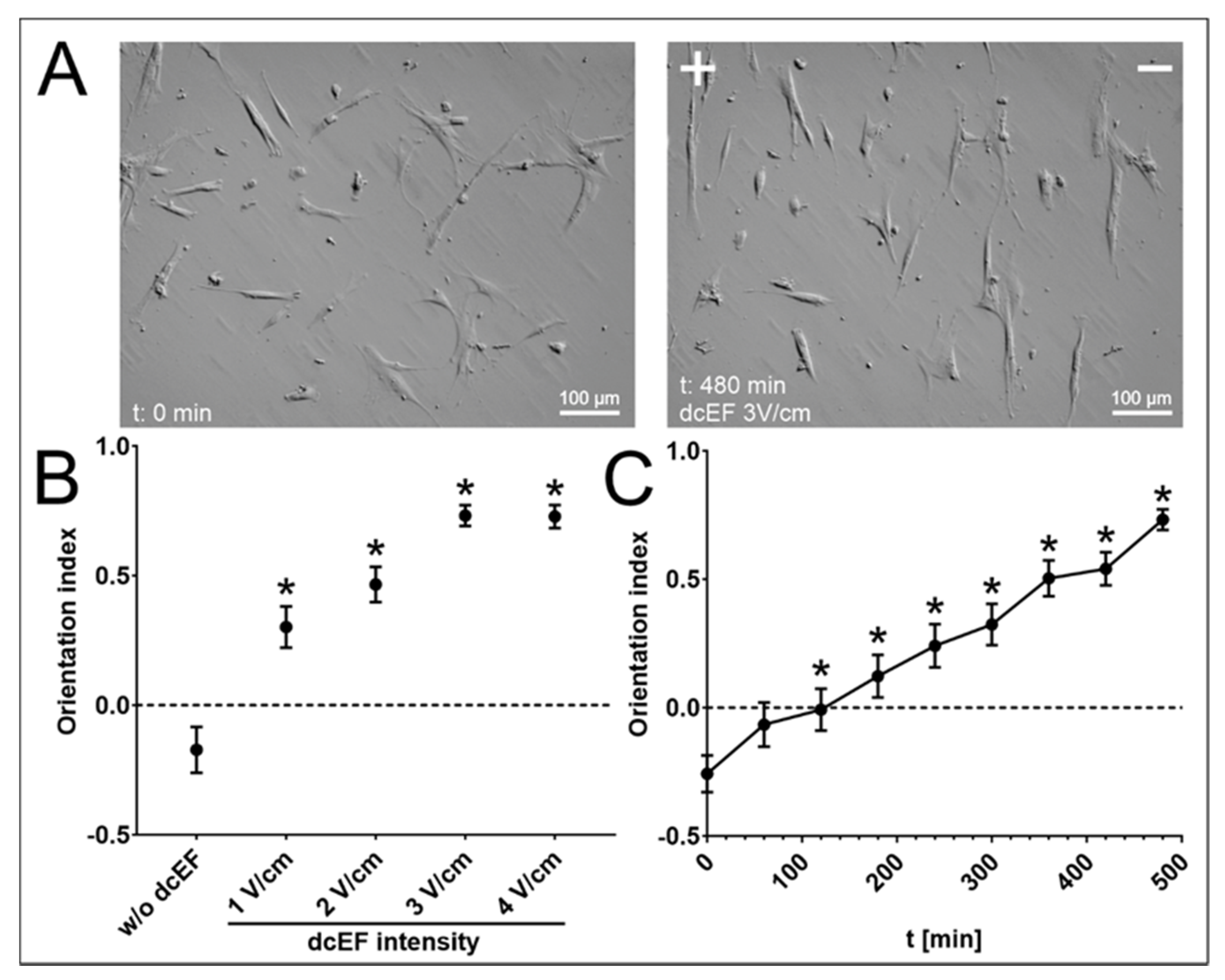
3.2. The Effect of an Electric Field on the Orientation of the Long Axis of NA HBF
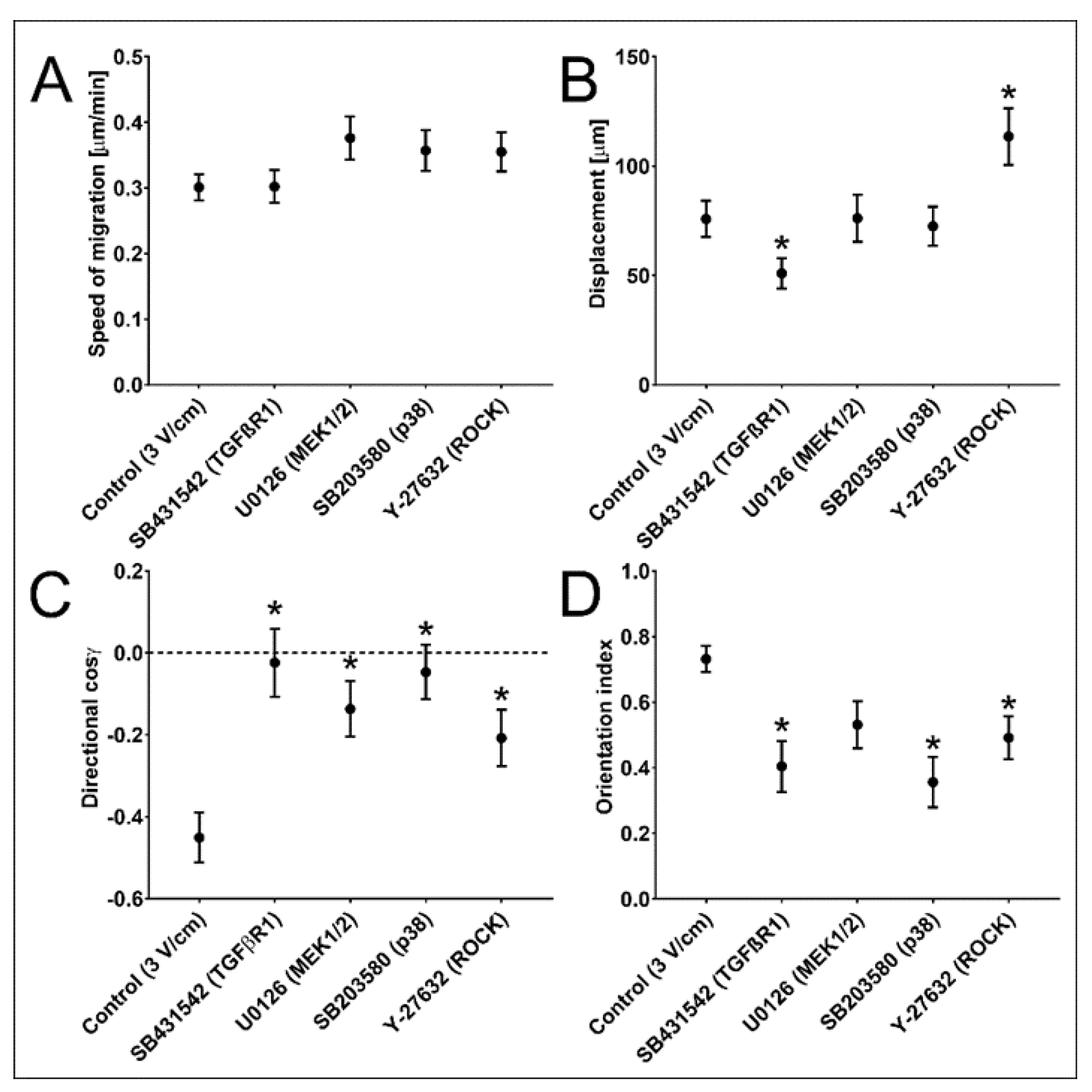
3.3. TGF-β Signalling Participates in NA HBF Responses to dcEFs
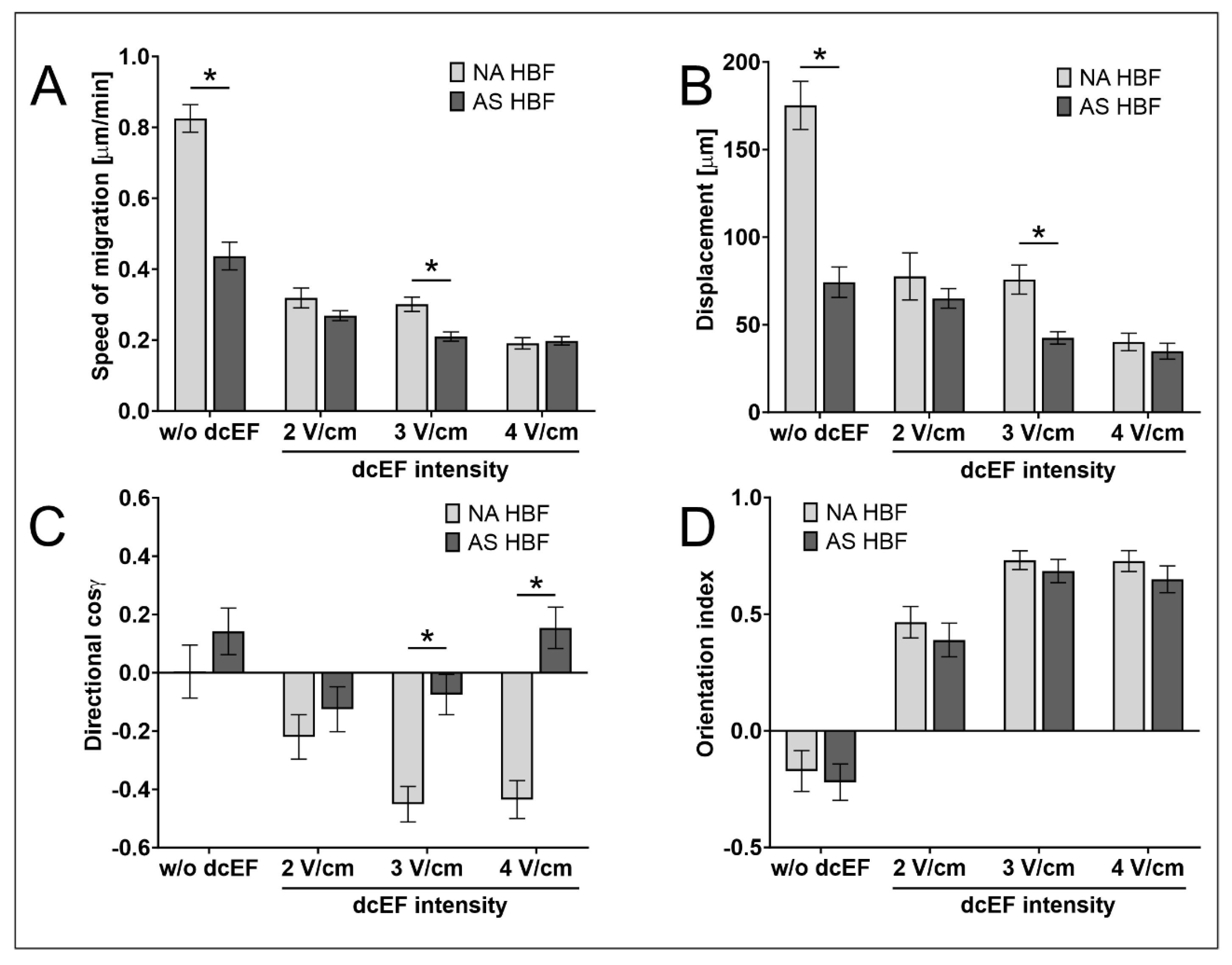
3.4. AS HBF Display Aberrant Reactivity to dcEFs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- du Bois-Reymond, E. Untersuchungen Über Thierische Elektricität; Verlag Von G. Reimer: Berlin, Germany, 1848; Volume 1. [Google Scholar]
- McCaig, C.D.; Rajnicek, A.M.; Song, B.; Zhao, M. Controlling Cell Behavior Electrically: Current Views and Future Potential. Physiol. Rev. 2005, 85, 943–978. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M. Electrical Fields in Wound Healing—An Overriding Signal That Directs Cell Migration. Semin. Cell Dev. Biol. 2009, 20, 674–682. [Google Scholar] [CrossRef]
- Djamgoz, M.B.A.; Mycielska, M.; Madeja, Z.; Fraser, S.P.; Korohoda, W. Directional Movement of Rat Prostate Cancer Cells in Direct-Current Electric Field: Involvement of Voltagegated Na+ Channel Activity. J. Cell Sci. 2001, 114, 2697–2705. [Google Scholar] [CrossRef]
- Mycielska, M.E.; Djamgoz, M.B.A. Cellular Mechanisms of Direct-Current Electric Field Effects: Galvanotaxis and Metastatic Disease. J. Cell Sci. 2004, 117, 1631–1639. [Google Scholar] [CrossRef] [Green Version]
- Sulik, G.L.; Soong, H.K.; Chang, P.C.; Parkinson, W.C.; Elner, S.G.; Elner, V.M. Effects of Steady Electric Fields on Human Retinal Pigment Epithelial Cell Orientation and Migration in Culture. Acta Ophthalmol. 1992, 70, 115–122. [Google Scholar] [CrossRef] [Green Version]
- Trollinger, D.R.; Isseroff, R.R.; Nuccitelli, R. Calcium Channel Blockers Inhibit Galvanotaxis in Human Keratinocytes. J. Cell Physiol. 2002, 193, 1–9. [Google Scholar] [CrossRef]
- Fang, K.S.; Farboud, B.; Nuccitelli, R.; Isseroff, R.R. Migration of Human Keratinocytes in Electric Fields Requires Growth Factors and Extracellular Calcium. J. Investig. Dermatol. 1998, 111, 751–756. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, K.Y.; Isseroff, R.R.; Nucciteili, R.; Nuccitelli, R. Human Keratinocytes Migrate to the Negative Pole in Direct Current Electric Fields Comparable to Those Measured in Mammalian Wounds. J. Cell Sci. 1996, 109 Pt 1, 199–207. [Google Scholar] [CrossRef]
- Sheridan, D.M.; Isseroff, R.R.; Nuccitelli, R. Imposition of a Physiologic DC Electric Field Alters the Migratory Response of Human Keratinocytes on Extracellular Matrix Molecules. J. Investig. Dermatol. 1996, 106, 642–646. [Google Scholar] [CrossRef] [Green Version]
- Soong, H.K.; Parkinson, W.C.; Bafna, S.; Sulik, G.L.; Huang, S.C. Movements of Cultured Corneal Epithelial Cells and Stromal Fibroblasts in Electric Fields. Investig. Ophthalmol. Vis. Sci. 1990, 31, 2278–2282. [Google Scholar]
- Farboud, B.; Nuccitelli, R.; Schwab, I.R.; Isseroff, R.R. DC Electric Fields Induce Rapid Directional Migration in Cultured Human Corneal Epithelial Cells. Exp. Eye Res. 2000, 70, 667–673. [Google Scholar] [CrossRef]
- Zhao, M.; Agius-Fernandez, A.; Forrester, J.V.; McCaig, C.D. Directed Migration of Corneal Epithelial Sheets in Physiological Electric Fields. Investig. Ophthalmol. Vis. Sci. 1996, 37, 2548–2558. [Google Scholar]
- Li, X.; Kolega, J. Effects of Direct Current Electric Fields on Cell Migration and Actin Filament Distribution in Bovine Vascular Endothelial Cells. J. Vasc. Res. 2002, 39, 391–404. [Google Scholar] [CrossRef]
- Kim, M.S.; Lee, M.H.; Kwon, B.-J.; Seo, H.J.; Koo, M.-A.; You, K.E.; Kim, D.; Park, J.-C. Control of Neonatal Human Dermal Fibroblast Migration on Poly(Lactic-Co-Glycolic Acid)-Coated Surfaces by Electrotaxis. J. Tissue Eng. Regen. Med. 2017, 11, 862–868. [Google Scholar] [CrossRef]
- Stump, R.F.; Robinson, K.R. Xenopus Neural Crest Cell Migration in an Applied Electrical Field. J. Cell Biol. 1983, 97, 1226–1233. [Google Scholar] [CrossRef]
- Nuccitelli, R.; Smart, T. Extracellular Calcium Levels Strongly Influence Neural Crest Cell Galvanotaxis. Biol. Bull. 1989, 176, 130–135. [Google Scholar] [CrossRef]
- Cooper, M.S.; Keller, R.E. Perpendicular Orientation and Directional Migration of Amphibian Neural Crest Cells in Dc Electrical Fields. Proc. Natl. Acad. Sci. USA 1984, 81, 160–164. [Google Scholar] [CrossRef]
- Allen, G.M.; Mogilner, A.; Theriot, J.A. Electrophoresis of Cellular Membrane Components Creates the Directional Cue Guiding Keratocyte Galvanotaxis. Curr. Biol. 2013, 23, 560–568. [Google Scholar] [CrossRef] [Green Version]
- Cooper, M.S.; Schliwa, M. Electrical and Ionic Controls of Tissue Cell Locomotion in DC Electric Fields. J. Neurosci. Res. 1985, 13, 223–244. [Google Scholar] [CrossRef]
- Cooper, M.S.; Schliwa, M. Motility of Cultured Fish Epidermal Cells in the Presence and Absence of Direct Current Electric Fields. J. Cell Biol. 1986, 102, 1384–1399. [Google Scholar] [CrossRef]
- Sroka, J.; Krecioch, I.; Zimolag, E.; Lasota, S.; Rak, M.; Kedracka-Krok, S.; Borowicz, P.; Gajek, M.; Madeja, Z. Lamellipodia and Membrane Blebs Drive Efficient Electrotactic Migration of Rat Walker Carcinosarcoma Cells WC 256. PLoS ONE 2016, 11, e0149133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrier, J.; Ross, S.M.; Kanehisa, J.; Aubin, J.E. Osteoclasts and Osteoblasts Migrate in Opposite Directions in Response to a Constant Electrical Field. J. Cell Physiol. 1986, 129, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Orida, N.; Feldman, J.D. Directional Protrusive Pseudopodial Activity and Motility in Macrophages Induced by Extracellular Electric Fields. Cell Motil. 1982, 2, 243–255. [Google Scholar] [CrossRef]
- Rapp, B.; de Boisfleury-Chevance, A.; Gruler, H. Galvanotaxis of Human Granulocytes. Dose-Response Curve. Eur. Biophys. J. 1988, 16, 313–319. [Google Scholar] [CrossRef]
- Zhang, J.; Calafiore, M.; Zeng, Q.; Zhang, X.; Huang, Y.; Li, R.A.; Deng, W.; Zhao, M. Electrically Guiding Migration of Human Induced Pluripotent Stem Cells. Stem Cell Rev. 2011, 7, 987–996. [Google Scholar] [CrossRef] [Green Version]
- Bai, H.; McCaig, C.D.; Forrester, J.V.; Zhao, M. DC Electric Fields Induce Distinct Preangiogenic Responses in Microvascular and Macrovascular Cells. Arterioscler. Thromb. Vasc. Biol. 2004, 24, 1234–1239. [Google Scholar] [CrossRef] [Green Version]
- Nuccitelli, R. A Role for Endogenous Electric Fields in Wound Healing. Curr. Top Dev. Biol. 2003, 58, 1–26. [Google Scholar] [CrossRef]
- McCaig, C.D.; Song, B.; Rajnicek, A.M. Electrical Dimensions in Cell Science. J. Cell Sci. 2009, 122, 4267–4276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorg, H.; Tilkorn, D.J.; Hager, S.; Hauser, J.; Mirastschijski, U. Skin Wound Healing: An Update on the Current Knowledge and Concepts. Eur. Surg. Res. 2017, 58, 81–94. [Google Scholar] [CrossRef]
- Yang, J.; Liu, X.; Wang, W.; Chen, Y.; Liu, J.; Zhang, Z.; Wu, C.; Jiang, X.; Liang, Y.; Zhang, J.P. Bioelectric Fields Coordinate Wound Contraction and Re-Epithelialization Process to Accelerate Wound Healing via Promoting Myofibroblast Transformation. Bioelectrochemistry 2022, 148, 108247. [Google Scholar] [CrossRef]
- Sun, Y.-H.Y.-H.; Reid, B.; Fontaine, J.H.; Miller, L.; Hyde, D.M.; Mogilner, A.; Zhao, M. Airway Epithelial Wounds in Rhesus Monkey Generate Ionic Currents That Guide Cell Migration to Promote Healing. J. Appl. Physiol. (1985) 2011, 111, 1031–1041. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadel, J.A.; Davis, B.; Phipps, R.J. Control of Mucus Secretion and Ion Transport in Airways. Annu. Rev. Physiol. 1979, 41, 369–381. [Google Scholar] [CrossRef]
- Tomkiewicz, R.P.; Albers, G.M.; De Sanctis, T.; Ramirez, O.E.; King, M.; Rubin, B.K. Species Differences in the Physical and Transport Properties of Airway Secretions. Can. J. Physiol. Pharmacol. 1995, 73, 165–171. [Google Scholar] [CrossRef]
- Park, J.A.; Kim, J.H.; Bi, D.; Mitchel, J.A.; Qazvini, N.T.; Tantisira, K.; Park, C.Y.; McGill, M.; Kim, S.H.; Gweon, B.; et al. Unjamming and Cell Shape in the Asthmatic Airway Epithelium. Nat. Mater. 2015, 14, 1040–1048. [Google Scholar] [CrossRef]
- Michalik, M.; Wójcik-Pszczoła, K.; Paw, M.; Wnuk, D.; Koczurkiewicz, P.; Sanak, M.; Pękala, E.; Madeja, Z. Fibroblast-to-Myofibroblast Transition in Bronchial Asthma. Cell. Mol. Life Sci. 2018, 75, 3943–3961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hackett, T.L.; Knight, D.A. The Role of Epithelial Injury and Repair in the Origins of Asthma. Curr. Opin. Allergy Clin. Immunol. 2007, 7, 63–68. [Google Scholar] [CrossRef] [PubMed]
- Inoue, H.; Akimoto, K.; Homma, T.; Tanaka, A.; Sagara, H. Airway Epithelial Dysfunction in Asthma: Relevant to Epidermal Growth Factor Receptors and Airway Epithelial Cells. J. Clin. Med. 2020, 9, 1–15. [Google Scholar] [CrossRef]
- Roth, H.M.; Wadsworth, S.J.; Kahn, M.; Knight, D.A. The Airway Epithelium in Asthma: Developmental Issues That Scar the Airways for Life? Pulm. Pharmacol. Ther. 2012, 25, 420–426. [Google Scholar] [CrossRef]
- Davies, D.E.; Wicks, J.; Powell, R.M.; Puddicombe, S.M.; Holgate, S.T. Airway Remodeling in Asthma: New Insights. J. Allergy Clin. Immunol. 2003, 111, 215–226. [Google Scholar] [CrossRef]
- Wnuk, D.; Lasota, S.; Paw, M.; Madeja, Z.; Michalik, M. Asthma-Derived Fibroblast to Myofibroblast Transition Is Enhanced in Comparison to Fibroblasts Derived from Non-Asthmatic Patients in 3D in Vitro Culture Due to Smad2/3 Signalling. Acta Biochim. Pol. 2020, 67, 441–448. [Google Scholar] [CrossRef]
- Wnuk, D.; Paw, M.; Ryczek, K.; Bochenek, G.; Sładek, K.; Madeja, Z.; Michalik, M. Enhanced Asthma-Related Fibroblast to Myofibroblast Transition Is the Result of Profibrotic TGF-β/Smad2/3 Pathway Intensification and Antifibrotic TGF-β/Smad1/5/(8)9 Pathway Impairment. Sci. Rep. 2020, 10, 16492. [Google Scholar] [CrossRef] [PubMed]
- Paw, M.; Borek, I.; Wnuk, D.; Ryszawy, D.; Piwowarczyk, K.; Kmiotek, K.; Wojcik-Pszczoła, K.A.; Pierzchalska, M.; Madeja, Z.; Sanak, M.; et al. Connexin43 Controls the Myofibroblastic Differentiation of Bronchial Fibroblasts from Patients with Asthma. Am. J. Respir. Cell. Mol. Biol. 2017, 57, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Michalik, M.; Pierzchalska, M.; Włodarczyk, A.; Wójcik, K.A.; Czy, J.; Sanak, M.; Madeja, Z. Transition of Asthmatic Bronchial Fibroblasts to Myofibroblasts Is Inhibited by Cell-Cell Contacts. Respir. Med. 2011, 105, 1467–1475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Michalik, M.; Pierzchalska, M.; Legutko, A.; Ura, M.; Ostaszewska, A.; Soja, J.; Sanak, M. Asthmatic Bronchial Fibroblasts Demonstrate Enhanced Potential to Differentiate into Myofibroblasts in Culture. Med. Sci. Monit. 2009, 15, BR194–BR201. [Google Scholar]
- Paw, M.; Wnuk, D.; Jakieła, B.; Bochenek, G.; Sładek, K.; Madeja, Z.; Michalik, M. Responsiveness of Human Bronchial Fibroblasts and Epithelial Cells from Asthmatic and Non-Asthmatic Donors to the Transforming Growth Factor-Β1 in Epithelial-Mesenchymal Trophic Unit Model. BMC Mol. Cell. Biol. 2021, 22, 19. [Google Scholar] [CrossRef]
- Sarna, M.; Wojcik, K.A.; Hermanowicz, P.; Wnuk, D.; Burda, K.; Sanak, M.; Czyö, J.; Michalik, M. Undifferentiated Bronchial Fibroblasts Derived from Asthmatic Patients Display Higher Elastic Modulus than Their Non-Asthmatic Counterparts. PLoS ONE 2015, 10, e0116840. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wójcik, K.A.; Skoda, M.; Koczurkiewicz, P.; Sanak, M.; Czyz, J.; Michalik, M. Apigenin Inhibits TGF-Β1 Induced Fibroblast-to-Myofibroblast Transition in Human Lung Fibroblast Populations. Pharmacol. Rep. 2013, 65, 164–172. [Google Scholar] [CrossRef]
- Sroka, J.; Zimolag, E.; Lasota, S.; Korohoda, W.; Madeja, Z. Electrotaxis: Cell Directional Movement in Electric Fields. In Cell Migration; Gautreau, A., Ed.; Methods in Molecular Biology; Humana Press: New York, NY, USA, 2018; Volume 1749, pp. 325–340. ISBN 978-1-4939-7701-7. [Google Scholar]
- Krecioch, I.; Madeja, Z.; Lasota, S.; Zimolag, E.; Sroka, J. The Role of Microtubules in Electrotaxis of Rat Walker Carcinosarcoma WC256 Cells. Acta Biochim. Pol. 2015, 62, 401–406. [Google Scholar] [CrossRef]
- Zimolag, E.; Borowczyk-Michalowska, J.; Kedracka-Krok, S.; Skupien-Rabian, B.; Karnas, E.; Lasota, S.; Sroka, J.; Drukala, J.; Madeja, Z. Electric Field as a Potential Directional Cue in Homing of Bone Marrow-Derived Mesenchymal Stem Cells to Cutaneous Wounds. Biochim. Et Biophys. Acta (BBA) -Mol. Cell Res. 2017, 1864, 267–279. [Google Scholar] [CrossRef] [Green Version]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzsch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An Open-Source Platform for Biological-Image Analysis. Nat. Methods 2012, 9, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Paw, M.; Wnuk, D.; Nit, K.; Bobis-Wozowicz, S.; Szychowski, R.; Ślusarczyk, A.; Madeja, Z.; Michalik, M. SB203580-A Potent P38 MAPK Inhibitor Reduces the Profibrotic Bronchial Fibroblasts Transition Associated with Asthma. Int. J. Mol. Sci. 2021, 22, 12790. [Google Scholar] [CrossRef]
- Chang, H.-F.; Cheng, H.-T.; Chen, H.-Y.; Yeung, W.K.; Cheng, J.-Y. Doxycycline Inhibits Electric Field-Induced Migration of Non-Small Cell Lung Cancer (NSCLC) Cells. Sci. Rep. 2019, 9, 8094. [Google Scholar] [CrossRef] [Green Version]
- Frederich, B.J.; Timofeyev, V.; Thai, P.N.; Haddad, M.J.; Poe, A.J.; Lau, V.C.; Moshref, M.; Knowlton, A.A.; Sirish, P.; Chiamvimonvat, N. Electrotaxis of Cardiac Progenitor Cells, Cardiac Fibroblasts, and Induced Pluripotent Stem Cell-Derived Cardiac Progenitor Cells Requires Serum and Is Directed via PI3′K Pathways. Heart Rhythm 2017, 14, 22. [Google Scholar] [CrossRef]
- Guo, A.; Song, B.; Reid, B.; Gu, Y.; Forrester, J.V.; Jahoda, C.A.B.; Zhao, M. Effects of Physiological Electric Fields on Migration of Human Dermal Fibroblasts. J. Investig. Dermatol. 2010, 130, 2320–2327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Guo, X.; Ren, X.; Tian, H.; Liang, Y.; Luo, Z.; Wang, W.; Wang, Y.; Zhang, D.; Huang, Y.; et al. A Novel FPCL Model Producing Directional Contraction through Induction of Fibroblast Alignment by Biphasic Pulse Direct Current Electric Field. Exp. Cell Res. 2018, 371, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Onuma, E.K.; Hui, S.W. Electric Field-Directed Cell Shape Changes, Displacement, and Cytoskeletal Reorganization Are Calcium Dependent. J. Cell Biol. 1988, 106, 2067–2075. [Google Scholar] [CrossRef] [PubMed]
- Finkelstein, E.; Chang, W.; Chao, P.-H.G.; Gruber, D.; Minden, A.; Hung, C.T.; Bulinski, J.C. Roles of Microtubules, Cell Polarity and Adhesion in Electric-Field-Mediated Motility of 3T3 Fibroblasts. J. Cell Sci. 2004, 117, 1533–1545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.-J.; Samorajski, J.; Kreimer, R.; Searson, P.C. The Influence of Electric Field and Confinement on Cell Motility. PLoS ONE 2013, 8, e59447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, M.J.; Loew, L.M. Electric Field-Directed Fibroblast Locomotion Involves Cell Surface Molecular Reorganization and Is Calcium Independent. J. Cell Biol. 1994, 127, 117–128. [Google Scholar] [CrossRef]
- Guo, L.; Xu, C.; Li, D.; Zheng, X.; Tang, J.; Bu, J.; Sun, H.; Yang, Z.; Sun, W.; Yu, X. Calcium Ion Flow Permeates Cells through SOCs to Promote Cathode-Directed Galvanotaxis. PLoS ONE 2015, 10, e0139865. [Google Scholar] [CrossRef] [Green Version]
- Erickson, C.A.; Nuccitelli, R. Embryonic Fibroblast Motility and Orientation Can Be Influenced by Physiological Electric Fields. J. Cell Biol. 1984, 98, 296–307. [Google Scholar] [CrossRef]
- Ross, S.M.; Ferrier, J.M.; Aubin, J.E. Studies on the Alignment of Fibroblasts in Uniform Applied Electrical Fields. Bioelectromagnetics 1989, 10, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Sillman, A.L.; Quang, D.M.; Farboud, B.; Fang, K.S.; Nuccitelli, R.; Isseroff, R.R. Human Dermal Fibroblasts Do Not Exhibit Directional Migration on Collagen I in Direct-Current Electric Fields of Physiological Strength. Exp. Dermatol. 2003, 12, 396–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, M.; Dick, A.; Forrester, J.V.; McCaig, C.D. Electric Field-Directed Cell Motility Involves up-Regulated Expression and Asymmetric Redistribution of the Epidermal Growth Factor Receptors and Is Enhanced by Fibronectin and Laminin. Mol. Biol. Cell 1999, 10, 1259–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, F.; Wang, H.; Li, L.; Huang, C.; Lin, J.; Zhu, G.; Chen, Z.; Wu, N.; Feng, H. Superoxide Plays Critical Roles in Electrotaxis of Fibrosarcoma Cells via Activation of ERK and Reorganization of the Cytoskeleton. Free Radic. Biol. Med. 2012, 52, 1888–1896. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Chen, T.; Hu, S.; Lin, J.; Hu, R.; Feng, H. Superoxide Mediates Direct Current Electric Field-Induced Directional Migration of Glioma Cells through the Activation of AKT and ERK. PLoS ONE 2013, 8, e61195. [Google Scholar] [CrossRef] [Green Version]
- Sun, S.; Wise, J.; Cho, M. Human Fibroblast Migration in Three-Dimensional Collagen Gel in Response to Noninvasive Electrical Stimulus. I. Characterization of Induced Three-Dimensional Cell Movement. Tissue Eng. 2004, 10, 1548–1557. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NA HBF; n = 50 | Speed of Cell Migration [μm/min] | Cell Displacement [μm] | Directionality (cos γ) [No Unit] |
|---|---|---|---|
| dcEF Intensity | Mean ± SEM | Mean ± SEM | Mean ± SEM |
| w/o (Control) | 0.825 ± 0.039 | 175.248 ± 13.693 | 0.004 ± 0.091 |
| 1 V/cm | 0.520 ± 0.036 * | 87.540 ± 10.835 * | −0.128 ± 0.077 |
| 2 V/cm | 0.319 ± 0.028 * | 77.610 ± 13.450 * | −0.220 ± 0.076 * |
| 3 V/cm | 0.301 ± 0.020 * | 75.796 ± 8.270 * | −0.451 ± 0.061 * |
| 4 V/cm | 0.191 ± 0.016 * | 40.143 ± 4.967 * | −0.435 ± 0.065 * |
| NA HBF; 3 V/cm dcEF; n = 50 | Speed of Cell Migration [μm/min] | Cell Displacement [μm] | Directionality (cos γ) [No Unit] | Orientation Index [No Unit] |
|---|---|---|---|---|
| Inhibitor (Target Protein) | Mean ± SEM | Mean ± SEM | Mean ± SEM | Mean ± SEM |
| w/o (Control) | 0.301 ± 0.020 | 75.796 ± 8.270 | −0.451 ± 0.061 | 0.731 ± 0.040 |
| SB431542 (TGFβR1) | 0.302 ± 0.025 | 50.990 ± 6.972 * | −0.024 ± 0.083 * | 0.404 ± 0.078 * |
| U0126 (MEK1/2) | 0.376 ± 0.033 | 76.163 ± 10.766 | −0.137 ± 0.068 * | 0.531 ± 0.072 |
| SB203580 (p38) | 0.357 ± 0.031 | 72.527 ± 8.941 | −0.047 ± 0.066 * | 0.356 ± 0.077 * |
| Y-27632 (ROCK) | 0.355 ± 0.030 | 113.532 ± 12.974 * | −0.208 ± 0.069 * | 0.492 ± 0.065 * |
| AS HBF; n = 50 | Speed of Cell Migration [μm/min] | Cell Displacement [μm] | Directionality (cos γ) [No Unit] | Orientation Index [No Unit] |
|---|---|---|---|---|
| dcEF Intensity | Mean ± SEM | Mean ± SEM | Mean ± SEM | Mean ± SEM |
| w/o (Control) | 0.437 ± 0.039 * | 74.273 ± 8.689 * | 0.142 ± 0.080 | −0.220 ± 0.078 |
| 2 V/cm | 0.269 ± 0.014 | 64.999 ± 5.579 | −0.125 ± 0.077 | 0.389 ± 0.072 |
| 3 V/cm | 0.210 ± 0.013 * | 42.496 ± 3.504 * | −0.075 ± 0.069 * | 0.685 ± 0.050 |
| 4 V/cm | 0.198 ± 0.012 | 34.867 ± 4.502 | 0.154 ± 0.071 * | 0.649 ± 0.057 |
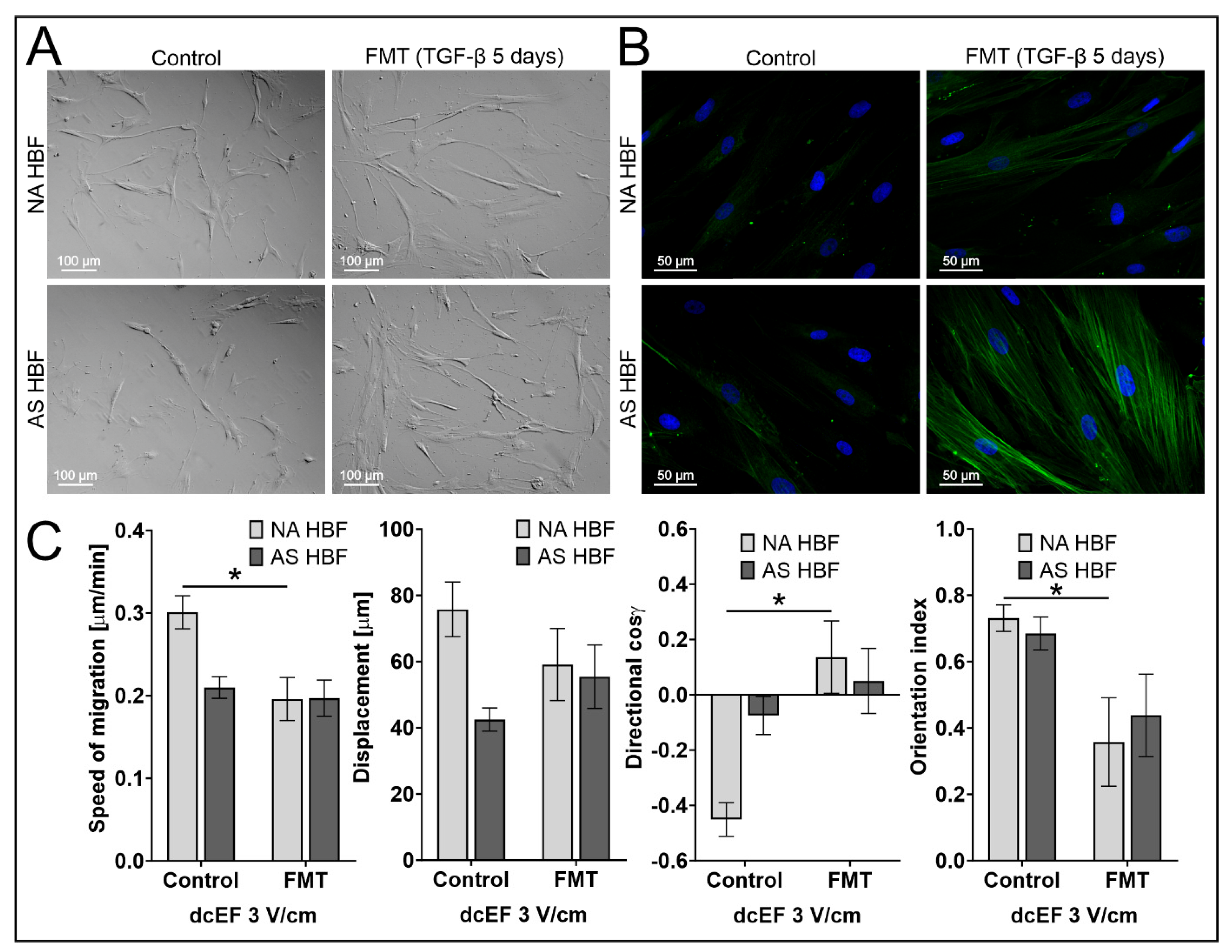
| 3 V/cm dcEF; FMT (TGF-β 5 ng/mL for 5 days); n = 20 | Speed of Cell Migration [μm/min] | Cell Displacement [μm] | Directionality (cos γ) [No Unit] | Orientation Index [No Unit] |
|---|---|---|---|---|
| Cell Population | Mean ± SEM | Mean ± SEM | Mean ± SEM | Mean ± SEM |
| NA HBF | 0.196 ± 0.026 * | 59.106 ± 10.895 | 0.136 ± 0.131 * | 0.357 ± 0.133 * |
| AS HBF | 0.197 ± 0.022 | 55.431 ± 9.580 | 0.050 ± 0.118 | 0.438 ± 0.124 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pavlenko, A.; Lasota, S.; Wnuk, D.; Paw, M.; Czyż, J.; Michalik, M.; Madeja, Z. Bronchial Fibroblasts from Asthmatic Patients Display Impaired Responsiveness to Direct Current Electric Fields (dcEFs). Biomedicines 2023, 11, 2138. https://doi.org/10.3390/biomedicines11082138
Pavlenko A, Lasota S, Wnuk D, Paw M, Czyż J, Michalik M, Madeja Z. Bronchial Fibroblasts from Asthmatic Patients Display Impaired Responsiveness to Direct Current Electric Fields (dcEFs). Biomedicines. 2023; 11(8):2138. https://doi.org/10.3390/biomedicines11082138
Chicago/Turabian StylePavlenko, Anastasiia, Sławomir Lasota, Dawid Wnuk, Milena Paw, Jarosław Czyż, Marta Michalik, and Zbigniew Madeja. 2023. "Bronchial Fibroblasts from Asthmatic Patients Display Impaired Responsiveness to Direct Current Electric Fields (dcEFs)" Biomedicines 11, no. 8: 2138. https://doi.org/10.3390/biomedicines11082138
APA StylePavlenko, A., Lasota, S., Wnuk, D., Paw, M., Czyż, J., Michalik, M., & Madeja, Z. (2023). Bronchial Fibroblasts from Asthmatic Patients Display Impaired Responsiveness to Direct Current Electric Fields (dcEFs). Biomedicines, 11(8), 2138. https://doi.org/10.3390/biomedicines11082138