
Morphofunctional Changes in Brain and Peripheral Blood in Adult and Aged Wistar Rats with AlCl3-Induced Neurodegeneration
 , , and
, , and
Abstract
:1. Introduction
2. Material and Methods
2.1. Animals
2.2. Neurodegeneration Modelling
2.3. Cognitive Function Evaluation
2.4. Samples Obtained and Histological Preparations
2.5. Morphological Study
2.6. Immunohistochemical Study
2.7. qPCR-RT Study
2.8. Flow Cytometry
2.9. Statistical Analysis
3. Results
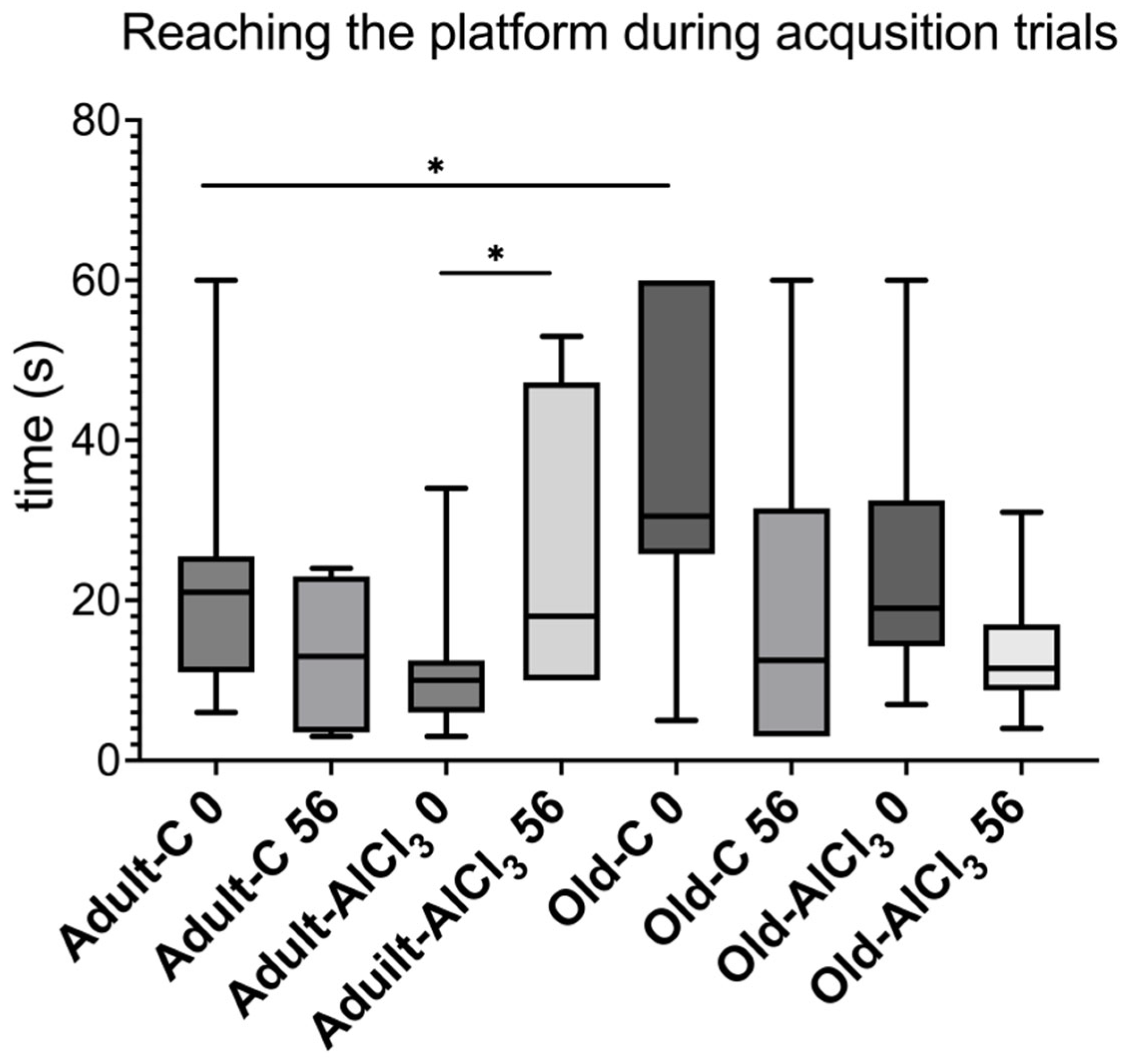
3.1. Short-Term Memory Evaluation
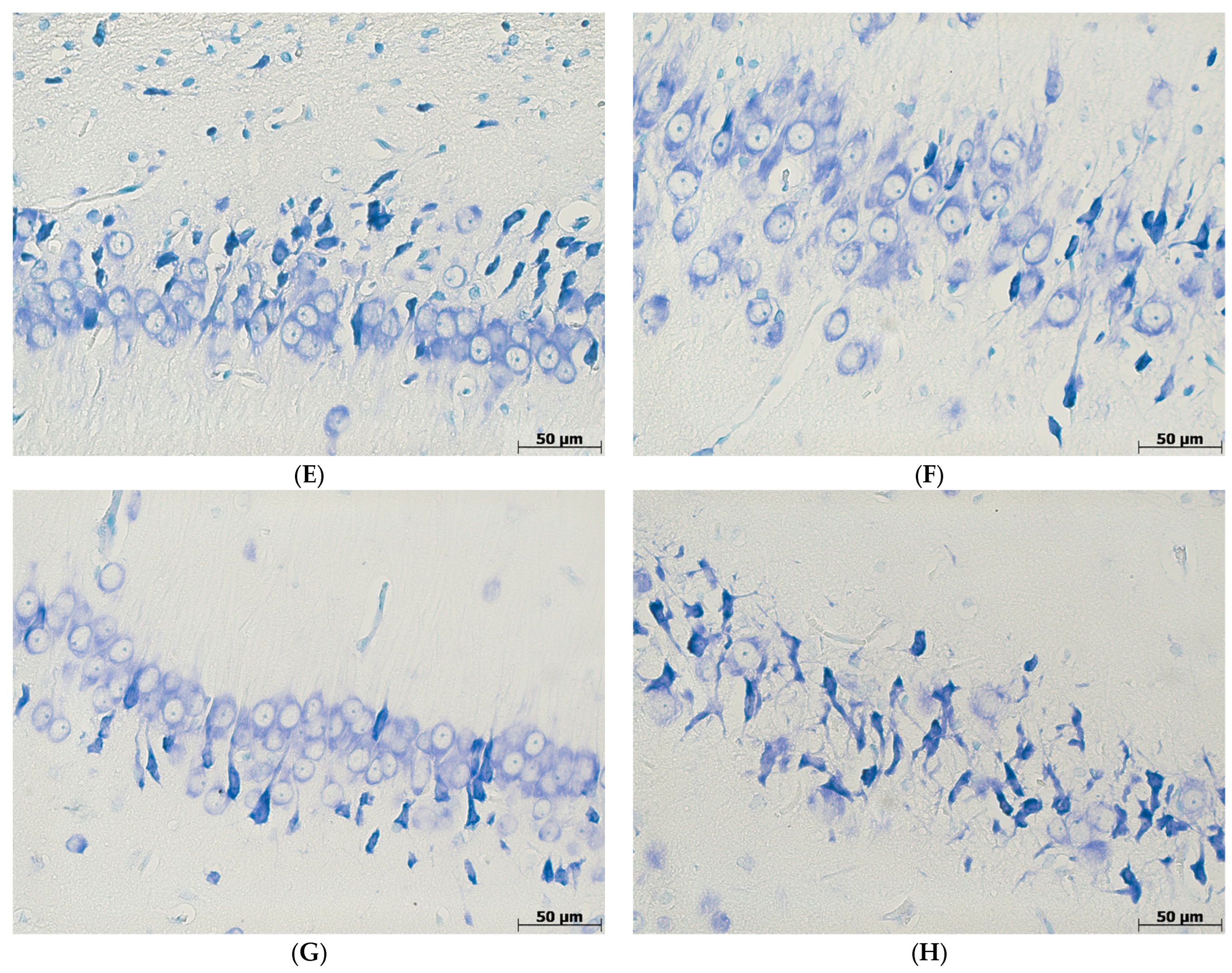
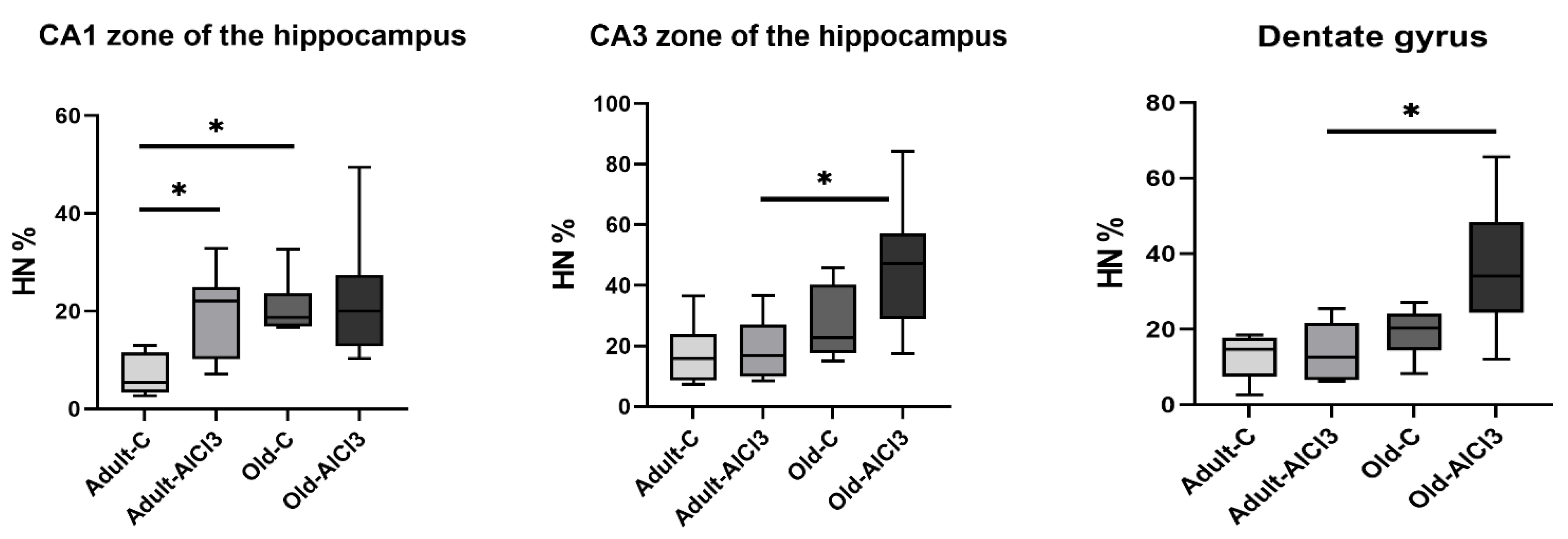
3.2. The Percentage of Hyperchromic Neurons
3.3. Morphological Features of Microglia
3.4. qPCR-RT Examination of the Prefrontal Cortex
3.4.1. Pro-inflammatory Markers
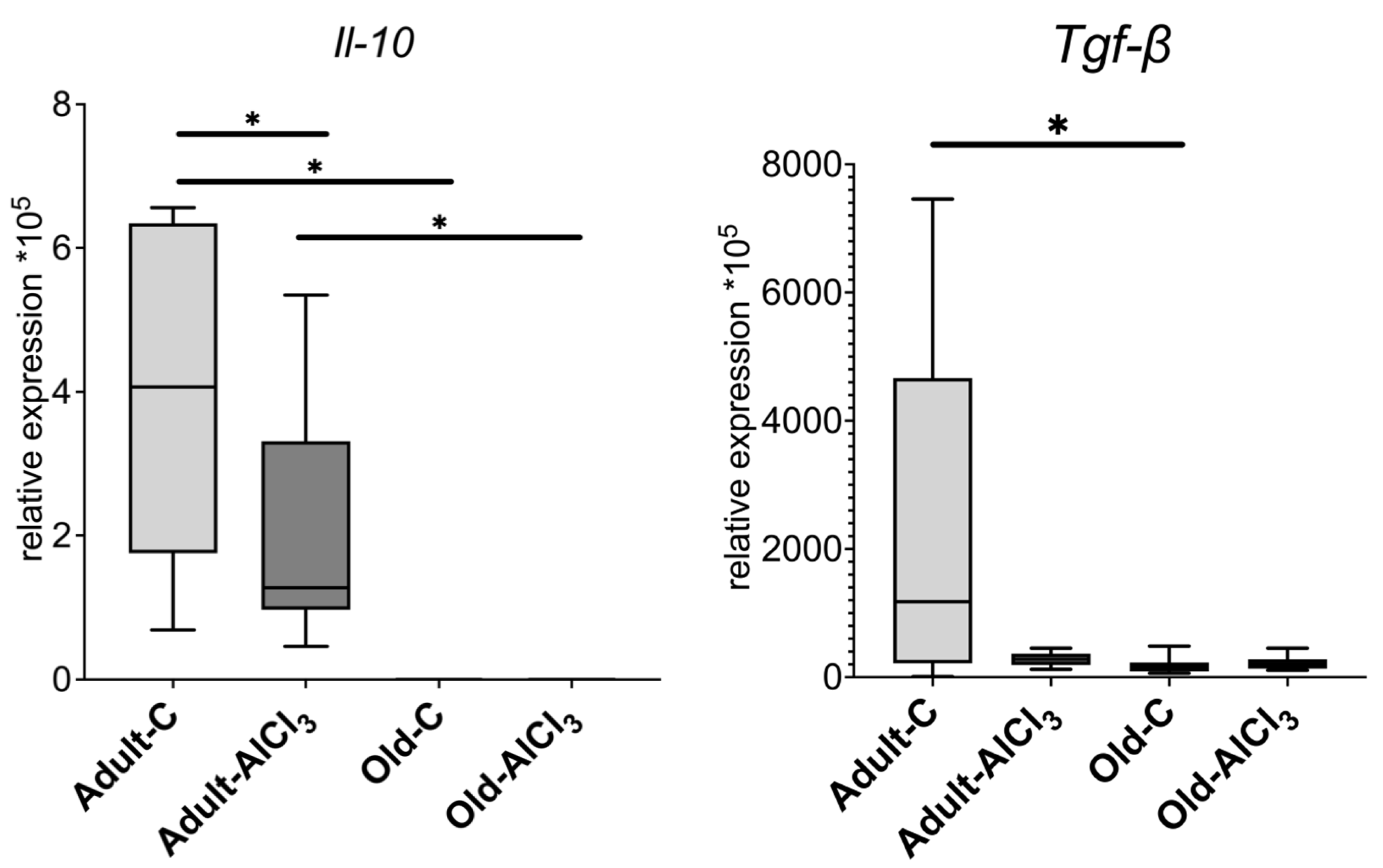
3.4.2. Anti-Inflammatory Markers
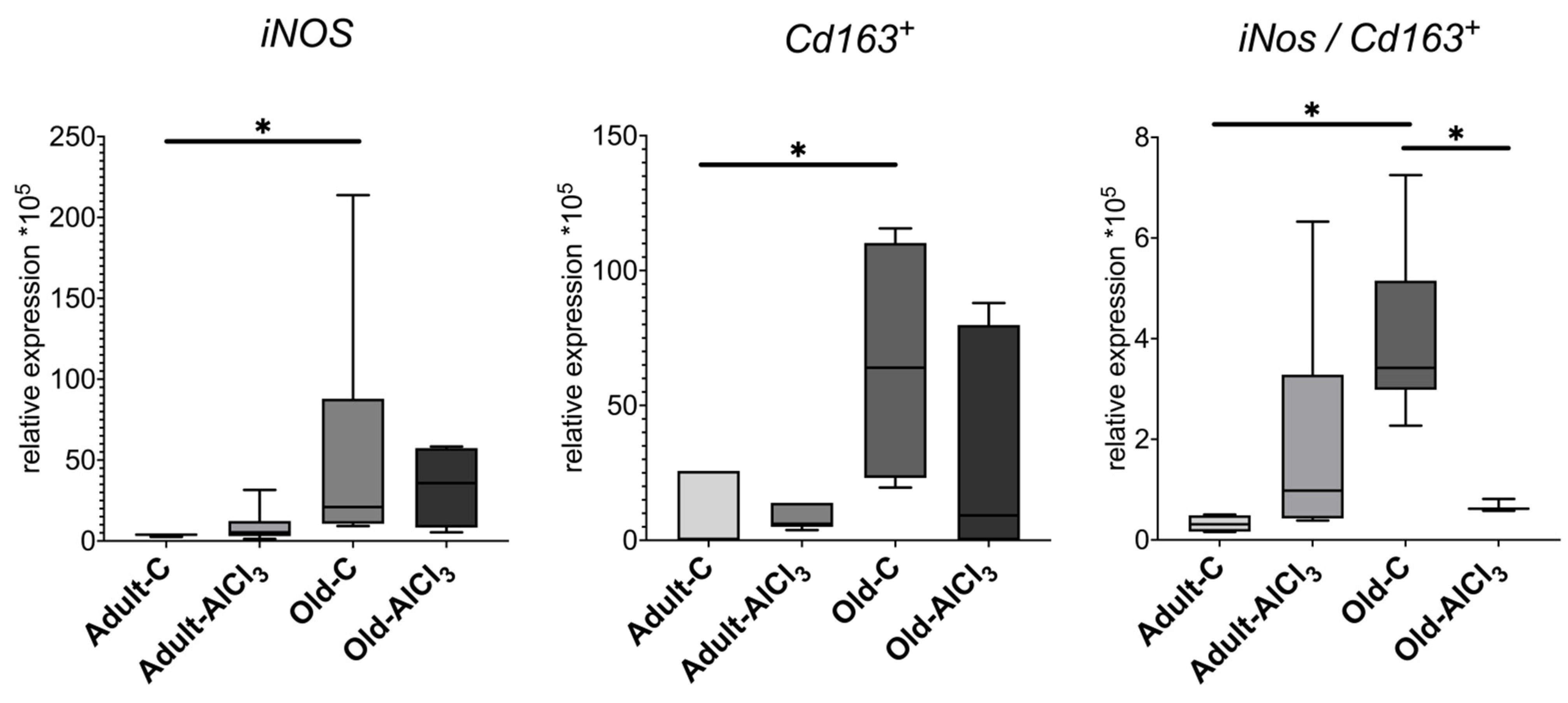
3.4.3. Microglia Activation Markers
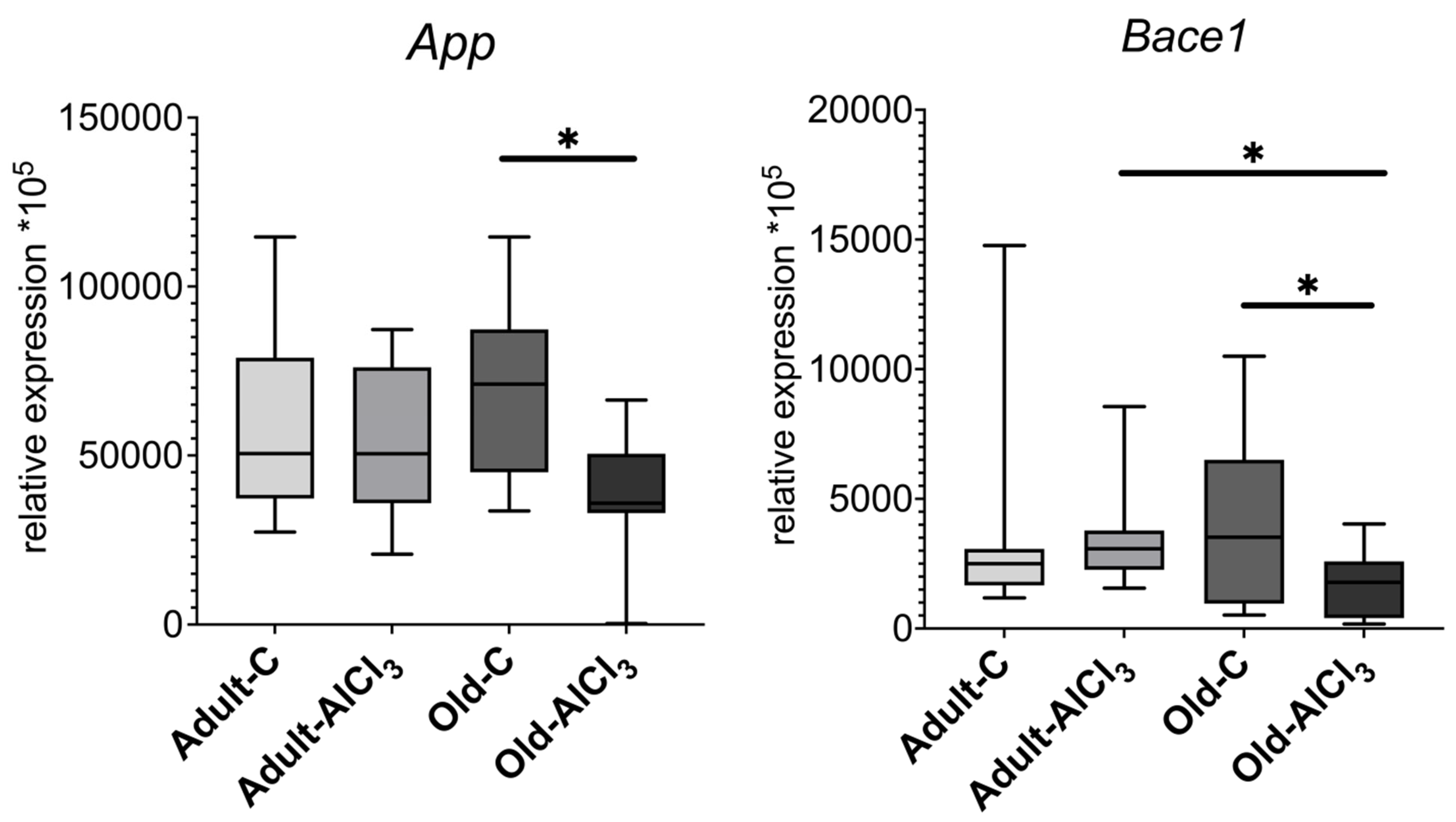
3.4.4. Amyloid-Related Proteins
3.4.5. Cellular Senescence Markers
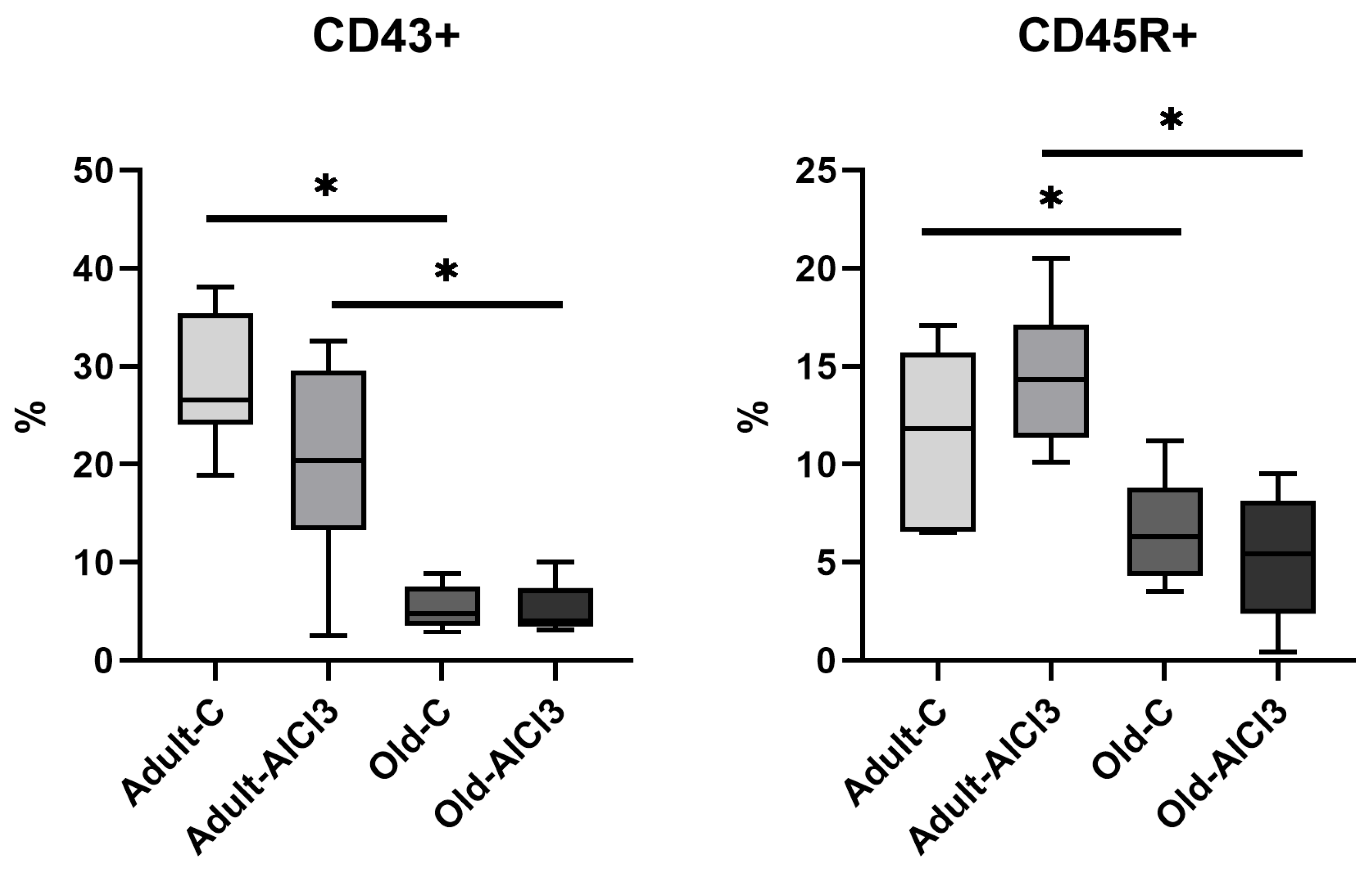
3.5. The Relative Numbers of Lymphocytes and Monocyte in Peripheral Blood
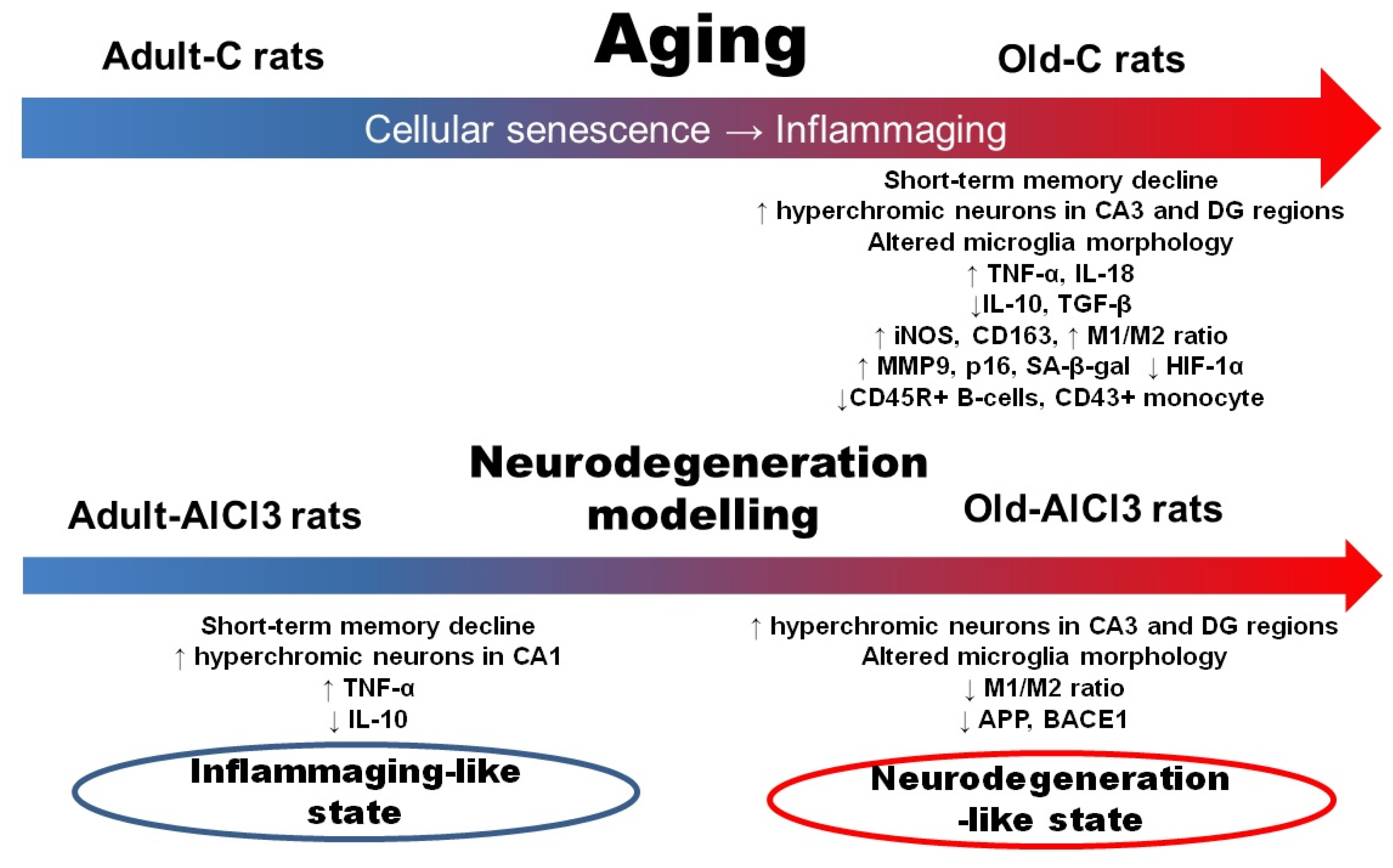
4. Discussion
4.1. Morphofunctional Changes in Brain and Peripheral Blood of Wistar Rats Due to Aging
4.2. Morphofunctional Changes in Brain and Peripheral Blood of Adult Wistar Rats with AlCl3-Induced Neurodegeneration
4.3. Morphofunctional Changes in Brain and Peripheral Blood of Old Wistar Rats with AlCl3-Induced Neurodegeneration
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Adult-AlCl3 | adult rats that consumed AlCl3 |
| Adult-C | adult rats control group |
| AlCl3 | aluminum chloride |
| APP | amyloid precursor protein |
| BACE1 | β-Site APP-cleaving enzyme 1 |
| BBB | blood–brain barrier |
| DG | dentate gyrus |
| CNS | central nervous system |
| GAPDH | glyceraldehyde 3-phosphate dehydrogenase |
| HIF-1α | hypoxia-inducible factor 1α |
| HRP | horseradish peroxidase |
| Iba1 | ionized calcium binding adaptor molecule 1 |
| ICH-P | Immunohistochemistry-paraffin protocol |
| IL-10 | interleukin 10 |
| IL-18 | interleukin 18 |
| iNOS | inducible nitric oxide synthase |
| MMP9 | matrix metallopeptidase 9 |
| MWM | Morris water maze |
| NF-κB | nuclear factor kappa-light-chain-enhancer of activated B cells |
| ODG | oxygen-glucose deprivation |
| Old-AlCl3 | old rats that consumed AlCl3 |
| Old-C | old rats control group |
| p16 | cyclin-dependent kinase inhibitor 2A |
| qPCR-RT | quantitative polymerase chain reaction, real-time regime |
| ROS | reactive oxygen species |
| Sa-β-gal | senescence-associated β-galactosidase |
| SASP | senescence-associated secretory phenotype |
| TGF-β | transforming growth factor β |
| TNF-α | tumor necrosis factor α |
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Forward Sequence | Reverse Sequence |
|---|---|---|
| GAPDH | GCGAGATCCCGCTAACATCA | CCCTTCCACGATGCCAAAGT |
| IL-18 | GACAAAAGAAACCCGCCTG | ACATCCTTCCATCCTTCACAG |
| TNF-a | CCACCACGCTCTTCTGTCTA | GCTACGGGCTTGTCACTCG |
| IL-10 | GCCCAGAAATCAAGGAGCAT | TGAGTGTCACGTAGGCTTCTA |
| TGF-β | CCGCAACAACGCAATCTATG | AGCCCTGTATTCCGTCTCCTT |
| iNOS | CGCTGGTTTGAAACTTCTCAG | GGCAAGCCATGTCTGTGAC |
| CD163 | TCTTGTGGACTCTGAAGCGA | TCTTAAATGCCAACCCGAGG |
| APP | TGGATGATCTCCAACCGTG | CGTCGACAGGCTCAACTTC |
| BACE1 | GGGCAGTAGTAATTTTGCAGT | TTCGGAGGTCTCGGTATGT |
| p16 | GTACCCCGATACAGGTGATG | GGTGCAGTACTACCAGAGTG |
| Sa-β-Gal | CTTCCGGATACCCCGATTCT | AGGGCACGTACGTCTGGAT |
| MMP9 | ATGGTTTCTGCCCCAGTGAG | CACCAGCGATAACCATCCGA |
| HIF-1a | TCACAGTCGGACAACCTCAC | TGCTGCAGTAACGTTCCAATTC |
References
- WHO. Dementia. 2021. Available online: https://www.who.int/publications/i/item/9789241550543 (accessed on 19 June 2023).
- Streit, W.J.; Braak, H.; Del Tredici, K.; Leyh, J.; Lier, J.; Khoshbouei, H.; Eisenlöffel, C.; Müller, W.; Bechmann, I. Microglial Activation Occurs Late During Preclinical Alzheimer’s Disease. Glia 2018, 66, 2550–2562. [Google Scholar] [CrossRef]
- van Dyck, C.H.; Swanson, C.J.; Aisen, P.; Bateman, R.J.; Chen, C.; Gee, M.; Kanekiyo, M.; Li, D.; Reyderman, L.; Cohen, S.; et al. Lecanemab in Early Alzheimer’s Disease. N. Engl. J. Med. 2023, 388, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Campisi, J. Chronic Inflammation (Inflammaging) and Its Potential Contribution to Age-Associated Diseases. J. Gerontol. A Ser. Biol. Sci. Med. Sci. 2014, 69 (Suppl. S1), S4–S9. [Google Scholar] [CrossRef] [PubMed]
- Dafsari, F.S.; Jessen, F. Depression—An underrecognized target for prevention of dementia in Alzheimer’s disease. Transl. Psychiatry 2020, 10, 160. [Google Scholar] [CrossRef] [PubMed]
- Alzheimer’s Disease Facts and Figures. Alzheimers Dement. 2022, 18, 700–789.
- Willhite, C.C.; Karyakina, N.A.; Yokel, R.A.; Yenugadhati, N.; Wisniewski, T.M.; Arnold, I.M.; Momoli, F.; Krewski, D. Systematic review of potential health risks posed by pharmaceutical, occupational and consumer exposures to metallic and nanoscale aluminum, aluminum oxides, aluminum hydroxide and its soluble salts. Crit. Rev. Toxicol. 2014, 44, 1–80. [Google Scholar] [CrossRef]
- Hesamian, M.S.; Eskandari, N. Potential Role of Trace Elements (Al, Cu, Zn, and Se) in Multiple Sclerosis Physiopathology. Neuroimmunomodulation 2020, 27, 163–177. [Google Scholar] [CrossRef]
- Firdaus, Z.; Kumar, D.; Singh, S.K.; Singh, T.D. Centella asiatica Alleviates AlCl3-induced Cognitive Impairment, Oxidative Stress, and Neurodegeneration by Modulating Cholinergic Activity and Oxidative Burden in Rat Brain. Biol. Trace Elem. Res. 2022, 200, 5115–5126. [Google Scholar] [CrossRef]
- Abramova, O.; Zorkina, Y.; Syunyakov, T.; Zubkov, E.; Ushakova, V.; Silantyev, A.; Soloveva, K.; Gurina, O.; Majouga, A.; Morozova, A.; et al. Brain Metabolic Profile after Intranasal vs. Intraperitoneal Clomipramine Treatment in Rats with Ultrasound Model of Depression. Int. J. Mol. Sci. 2021, 22, 9598. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, C.R.; Emson, P.C. Ache-Stained Horizontal Sections of the Rat Brain in Stereotaxic Coordinates. J. Neurosci. Methods 1980, 3, 129–149. [Google Scholar] [CrossRef]
- Shahidehpour, R.K.; Higdon, R.E.; Crawford, N.G.; Neltner, J.H.; Ighodaro, E.T.; Patel, E.; Price, D.; Nelson, P.T.; Bachstetter, A.D. Dystrophic microglia are associated with neurodegenerative disease and not healthy aging in the human brain. Neurobiol. Aging 2021, 99, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Rider, D.A.; Ruan, R. Identification of Valid Housekeeping Genes and Antioxidant Enzyme Gene Expression Change in the Aging Rat Liver. J. Gerontol. Ser. A 2006, 61, 20–27. [Google Scholar] [CrossRef] [PubMed]
- Franceschi, C.; Bonafè, M.; Valensin, S.; Olivieri, F.; De Luca, M.; Ottaviani, E.; De Benedictis, G. Inflammaging. An Evolutionary Perspective on Immunosenescence. Ann. N. Y. Acad. Sci. 2000, 908, 244–254. [Google Scholar] [CrossRef] [PubMed]
- Villarreal-Silva, E.E.; González-Navarro, A.R.; Salazar-Ybarra, R.A.; Quiroga-García, O.; Cruz-Elizondo, M.A.D.J.; García-García, A.; Rodríguez-Rocha, H.; Morales-Gómez, J.A.; Quiroga-Garza, A.; Elizondo-Omaña, R.E.; et al. Aged rats learn Morris Water maze using non-spatial search strategies evidenced by a parameter-based algorithm. Transl. Neurosci. 2022, 13, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Gil-Mohapel, J.; Brocardo, P.S.; Choquette, W.; Gothard, R.; Simpson, J.M.; Christie, B.R. Hippocampal Neurogenesis Levels Predict WATERMAZE Search Strategies in the Aging Brain. PLoS ONE 2013, 8, e75125. [Google Scholar] [CrossRef]
- Burger, C.; López, M.C.; Feller, J.A.; Baker, H.V.; Muzyczka, N.; Mandel, R.J. Changes in transcription within the CA1 field of the hippocampus are associated with age-related spatial learning impairments. Neurobiol. Learn. Mem. 2007, 87, 21–41. [Google Scholar] [CrossRef]
- Ekova, M.; Smirnov, A.V.; Tyurenkov, I.N. Interrelation Structural and Functional Alterations in the Dentate Gyrus of Rats with Aging. Alzheimer’s Dement. 2021, 17, e052607. [Google Scholar]
- Zimatkin, S.M.; Bon’, E.I. Dark Neurons of the Brain. Neurosci. Behav. Physiol. 2018, 48, 908–912. [Google Scholar] [CrossRef]
- Ooigawa, H.; Nawashiro, H.; Fukui, S.; Otani, N.; Osumi, A.; Toyooka, T.; Shima, K. The fate of Nissl-stained dark neurons following traumatic brain injury in rats: Difference between neocortex and hippocampus regarding survival rate. Acta Neuropathol. 2006, 112, 471–481. [Google Scholar] [CrossRef]
- Korzhevskii, D.E. Neurodegeneration and Assessment of the Response of Nerve Cells to Damage. In Molecular Neuromorphology; SpetsLit.: Saint-Petersburg, Russia, 2015; p. 110. [Google Scholar]
- Streit, W.J.; Khoshbouei, H.; Bechmann, I. The Role of Microglia in Sporadic Alzheimer’s Disease. J. Alzheimer’s Dis. 2021, 79, 961–968. [Google Scholar] [CrossRef]
- Harry, G.J. Microglia during development and aging. Pharmacol. Ther. 2013, 139, 313–326. [Google Scholar] [CrossRef] [PubMed]
- Campuzano, O.; Castillo-Ruiz, M.M.; Acarin, L.; Castellano, B.; Gonzalez, B. Increased Levels of Pro-inflammatory Cyto-kines in the Aged Rat Brain Attenuate Injury-Induced Cytokine Response after Excitotoxic Damage. J. Neurosci. Res. 2009, 87, 2484–2497. [Google Scholar] [CrossRef] [PubMed]
- Ojala, J.O.; Sutinen, E.M. The Role of Interleukin-18, Oxidative Stress and Metabolic Syndrome in Alzheimer’s Disease. J. Clin. Med. 2017, 6, 55. [Google Scholar] [CrossRef] [PubMed]
- Lecca, D.; Jung, Y.J.; Scerba, M.T.; Hwang, I.; Kim, Y.K.; Kim, S.; Modrow, S.; Tweedie, D.; Hsueh, S.; Liu, D.; et al. Role of chronic neuroinflammation in neuroplasticity and cognitive function: A hypothesis. Alzheimer’s Dement. 2022, 18, 2327–2340. [Google Scholar] [CrossRef]
- Kosyreva, A.M.; Sentyabreva, A.V.; Tsvetkov, I.S.; Makarova, O.V. Alzheimer’s Disease and Inflammaging. Brain Sci. 2022, 12, 1237. [Google Scholar] [CrossRef] [PubMed]
- Porro, C.; Cianciulli, A.; Panaro, M.A. The Regulatory Role of IL-10 in Neurodegenerative Diseases. Biomolecules 2020, 10, 1017. [Google Scholar] [CrossRef]
- Pereira, L.; Font-Nieves, M.; Haute, C.V.D.; Baekelandt, V.; Planas, A.M.; Pozas, E. IL-10 regulates adult neurogenesis by modulating ERK and STAT3 activity. Front. Cell. Neurosci. 2015, 9, 57. [Google Scholar] [CrossRef]
- Norden, D.M.; Fenn, A.M.; Dugan, A.; Godbout, J.P. TGFβ produced by IL-10 redirected astrocytes attenuates microglial activation. Glia 2014, 62, 881–895. [Google Scholar] [CrossRef]
- Vogel, T.; Ahrens, S.; Büttner, N.; Krieglstein, K. Transforming Growth Factor Beta Promotes Neuronal Cell Fate of Mouse Cortical and Hippocampal Progenitors in Vitro and in Vivo: Identification of Nedd9 as an Essential Signaling Component. Cereb. Cortex 2010, 20, 661–671. [Google Scholar] [CrossRef]
- Fukushima, T.; Liu, R.Y.; Byrne, J.H. Transforming Growth Factor-Beta2 Modulates Synaptic Efficacy and Plasticity and In-duces Phosphorylation of Creb in Hippocampal Neurons. Hippocampus 2007, 17, 5–9. [Google Scholar] [CrossRef]
- Wu, Z.; Hayashi, Y.; Zhang, J.; Nakanishi, H. Involvement of Prostaglandin E2 Released from Leptomeningeal Cells in In-creased Expression of Transforming Growth Factor-Beta in Glial Cells and Cortical Neurons During Systemic Inflammation. J. Neurosci. Res. 2007, 85, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Lenzlinger, P.M.; Morganti-Kossmann, M.C.; Laurer, H.L.; McIntosh, T.K. The Duality of the Inflammatory Response to Traumatic Brain Injury. Mol. Neurobiol. 2001, 24, 169–181. [Google Scholar] [CrossRef] [PubMed]
- Tominaga, K.; Suzuki, H.I. TGF-β Signaling in Cellular Senescence and Aging-Related Pathology. Int. J. Mol. Sci. 2019, 20, 5002. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Molina, P.; Almolda, B.; Giménez-Llort, L.; González, B.; Castellano, B. Chronic Il-10 Overproduction Disrupts Microglia-Neuron Dialogue Similar to Aging, Resulting in Impaired Hippocampal Neurogenesis and Spatial Memory. Brain Behav. Immun. 2022, 101, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Wang, H.; Yin, Y. Microglia Polarization from M1 to M2 in Neurodegenerative Diseases. Front. Aging Neurosci. 2022, 14, 815347. [Google Scholar] [CrossRef]
- Das, B.; Singh, N.; Yao, A.Y.; Zhou, J.; He, W.; Hu, X.; Yan, R. BACE1 controls synaptic function through modulating release of synaptic vesicles. Mol. Psychiatry 2021, 26, 6394–6410. [Google Scholar] [CrossRef]
- Jack, C.R., Jr.; Bennett, D.A.; Blennow, K.; Carrillo, M.C.; Dunn, B.; Haeberlein, S.B.; Holtzman, D.M.; Jagust, W.; Jessen, F.; Karlawish, J.; et al. Nia-Aa Research Framework: Toward a Biological Definition of Alzheimer’s Disease. Alzheimer’s Dement. 2018, 14, 535–562. [Google Scholar] [CrossRef]
- Hernandez-Segura, A.; de Jong, T.V.; Melov, S.; Guryev, V.; Campisi, J.; Demaria, M. Unmasking Transcriptional Hetero-geneity in Senescent Cells. Curr. Biol. 2017, 27, 2652–2660. [Google Scholar] [CrossRef]
- Huang, W.; Hickson, L.J.; Eirin, A.; Kirkland, J.L.; Lerman, L.O. Cellular Senescence: The Good, the Bad and the Unknown. Nat. Rev. Nephrol. 2022, 18, 611–627. [Google Scholar] [CrossRef]
- Rayess, H.; Wang, M.B.; Srivatsan, E.S. Cellular senescence and tumor suppressor gene p16. Int. J. Cancer 2012, 130, 1715–1725. [Google Scholar] [CrossRef]
- Lee, B.Y.; Han, J.A.; Im, J.S.; Morrone, A.; Johung, K.; Goodwin, E.C.; Kleijer, W.J.; DiMaio, D.; Hwang, E.S. Senescence-Associated Beta-Galactosidase Is Lysosomal Beta-Galactosidase. Aging Cell 2006, 5, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Yabluchanskiy, A.; Ma, Y.; Iyer, R.P.; Hall, M.E.; Lindsey, M.L. Matrix Metalloproteinase-9: Many Shades of Function in Cardiovascular Disease. Physiology 2013, 28, 391–403. [Google Scholar] [CrossRef]
- Fang, L.; Du, X.-J.; Gao, X.-M.; Dart, A.M. Activation of peripheral blood mononuclear cells and extracellular matrix and inflammatory gene profile in acute myocardial infarction. Clin. Sci. 2010, 119, 175–183. [Google Scholar] [CrossRef]
- Ndubuizu, O.I.; Chavez, J.C.; LaManna, J.C. Increased Prolyl 4-Hydroxylase Expression and Differential Regulation of Hy-poxia-Inducible Factors in the Aged Rat Brain. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 297, R158–R165. [Google Scholar] [CrossRef] [PubMed]
- Dzhalilova, D.; Kosyreva, A.; Vishnyakova, P.; Zolotova, N.; Tsvetkov, I.; Mkhitarov, V.; Mikhailova, L.; Kakturskiy, L.; Makarova, O. Age-related differences in hypoxia-associated genes and cytokine profile in male Wistar rats. Heliyon 2021, 7, e08085. [Google Scholar] [CrossRef] [PubMed]
- Dowery, R.; Benhamou, D.; Benchetrit, E.; Harel, O.; Nevelsky, A.; Zisman-Rozen, S.; Braun-Moscovici, Y.; Balbir-Gurman, A.; Avivi, I.; Shechter, A.; et al. Peripheral B Cells Repress B-Cell Regeneration in Aging through a Tnf-A/Igfbp-1/Igf-1 Immune-Endocrine Axis. Blood 2021, 138, 1817–1829. [Google Scholar] [CrossRef]
- Snodgrass, R.G.; Jiang, X.; Stephensen, C.B. Monocyte subsets display age-dependent alterations at fasting and undergo non-age-dependent changes following consumption of a meal. Immun. Ageing 2022, 19, 41. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Xin, T.R.; Liang, J.J.; Wang, W.M.; Zhang, Y.Y. Memory Performance, Brain Excitatory Amino Acid and Acetyl-cholinesterase Activity of Chronically Aluminum Exposed Mice in Response to Soy Isoflavones Treatment. Phytother. Res. 2010, 24, 1451–1456. [Google Scholar] [CrossRef]
- Gupta, G.; Chellappan, D.K.; Agarwal, M.; Ashwathanarayana, M.; Nammi, S.; Pabreja, K.; Dua, K. Pharmacological Evaluation of the Recuperative Effect of Morusin against Aluminium Trichloride (Alcl3)-Induced Memory Impairment in Rats. Cent. Nerv. Syst. Agents Med. Chem. 2017, 17, 196–200. [Google Scholar] [CrossRef]
- Lana, D.; Ugolini, F.; Giovannini, M.G. An Overview on the Differential Interplay among Neurons–Astrocytes–Microglia in CA1 and CA3 Hippocampus in Hypoxia/Ischemia. Front. Cell. Neurosci. 2020, 14, 585833. [Google Scholar] [CrossRef]
- Mizuseki, K.; Royer, S.; Diba, K.; Buzsáki, G. Activity dynamics and behavioral correlates of CA3 and CA1 hippocampal pyramidal neurons. Hippocampus 2012, 22, 1659–1680. [Google Scholar] [CrossRef] [PubMed]
- Padurariu, M.; Ciobica, A.; Mavroudis, I.; Fotiou, D.; Baloyannis, S. Hippocampal Neuronal Loss in the CA1 and CA3 Areas of Alzheimer’s Disease Patients. Psychiatr Danub. 2012, 24, 152–158. [Google Scholar] [PubMed]
- Nafea, M.; Elharoun, M.; Abd-Alhaseeb, M.M.; Helmy, M.W. Leflunomide Abrogates Neuroinflammatory Changes in a Rat Model of Alzheimer’s Disease: The Role of Tnf-A/Nf-Κb/Il-1β Axis Inhibition. Naunyn Schmiedeberg’s Arch. Pharmacol. 2023, 396, 485–498. [Google Scholar] [CrossRef]
- Lahiri, D.K.; Chen, D.; Vivien, D.; Ge, Y.W.; Greig, N.H.; Rogers, J.T. Role of Cytokines in the Gene Expression of Amyloid Beta-Protein Precursor: Identification of a 5′-Utr-Binding Nuclear Factor and Its Implications in Alzheimer’s Disease. J. Alzheimer’s Dis. 2003, 5, 81–90. [Google Scholar] [CrossRef] [PubMed]
- She, Y.; Wang, N.; Chen, C.; Zhu, Y.; Xia, S.; Hu, C.; Li, Y. Effects of Aluminum on Immune Functions of Cultured Splenic T and B Lymphocytes in Rats. Biol. Trace Elem. Res. 2011, 147, 246–250. [Google Scholar] [CrossRef] [PubMed]












| Peripheral Blood T-Cells, % | Adult-C | Adult-AlCl3 | Old-C | Old-AlCl3 | p |
|---|---|---|---|---|---|
| CD3+ | 25.8 (19.4–36) | 29.25 (27.8–36.4) | 23.35 (18.5–37.1) | 20.9 (11.3–24.4) | >0.05 |
| CD3+CD4+ | 11.3 (9.3–13.5) | 13 (12.4–17.9) | 12.75 (8.1–14.7) | 11.3 (8.6–12.7) | >0.05 |
| CD3+CD8+ | 12.3 (5.7–18.7) | 12.1 (8–22.1) | 12.65 (5.6–27.1) | 6.2 (5.5–11.3) | >0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sentyabreva, A.V.; Miroshnichenko, E.A.; Melnikova, E.A.; Tsvetkov, I.S.; Kosyreva, A.M. Morphofunctional Changes in Brain and Peripheral Blood in Adult and Aged Wistar Rats with AlCl3-Induced Neurodegeneration. Biomedicines 2023, 11, 2336. https://doi.org/10.3390/biomedicines11092336
Sentyabreva AV, Miroshnichenko EA, Melnikova EA, Tsvetkov IS, Kosyreva AM. Morphofunctional Changes in Brain and Peripheral Blood in Adult and Aged Wistar Rats with AlCl3-Induced Neurodegeneration. Biomedicines. 2023; 11(9):2336. https://doi.org/10.3390/biomedicines11092336
Chicago/Turabian StyleSentyabreva, Alexandra Vladislavovna, Ekaterina Alexandrovna Miroshnichenko, Ekaterina Andreevna Melnikova, Ivan Sergeevich Tsvetkov, and Anna Mikhailovna Kosyreva. 2023. "Morphofunctional Changes in Brain and Peripheral Blood in Adult and Aged Wistar Rats with AlCl3-Induced Neurodegeneration" Biomedicines 11, no. 9: 2336. https://doi.org/10.3390/biomedicines11092336
APA StyleSentyabreva, A. V., Miroshnichenko, E. A., Melnikova, E. A., Tsvetkov, I. S., & Kosyreva, A. M. (2023). Morphofunctional Changes in Brain and Peripheral Blood in Adult and Aged Wistar Rats with AlCl3-Induced Neurodegeneration. Biomedicines, 11(9), 2336. https://doi.org/10.3390/biomedicines11092336