
Expression of Selected miRNAs in Undifferentiated Carcinoma with Osteoclast-like Giant Cells (UCOGC) of the Pancreas: Comparison with Poorly Differentiated Pancreatic Ductal Adenocarcinoma
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Samples
2.2. MicroRNA Isolation and Reverse Transcription
2.3. Real-Time qPCR
2.4. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hruban, R.H.; Adsay, N.V.; Esposito, I.; Fukushima, N.; Furukawa, T.; Klöppel, G.; Maitra, A.; Notohara, K.; Offerhaus, G.J.A.; Ohike, N.; et al. Pancreatic ductal adenocarcinoma. In WHO Classification of Tumours Editorial Board. Digestive System Tumours; International Agency for Research on Cancer: Lyon, France, 2019; pp. 329–330. ISBN 978-92-832-4499-8. [Google Scholar]
- Muraki, T.; Reid, M.D.; Basturk, O.; Jang, K.T.; Bedolla, G.; Bagci, P.; Mittal, P.; Memis, B.; Katabi, N.; Bandyopadhyay, S.; et al. Undifferentiated Carcinoma with Osteoclastic Giant Cells of the Pancreas: Clinicopathologic Analysis of 38 Cases Highlights a More Protracted Clinical Course than Currently Appreciated. Am. J. Surg. Pathol. 2016, 40, 1203–1216. [Google Scholar] [CrossRef]
- Sommers, S.C.; Meissner, W.A. Unusual Carcinomas of the Pancreas. AMA Arch. Pathol. 1954, 58, 101–111. [Google Scholar]
- Rosai, J. Carcinoma of Pancreas Simulating Giant Cell Tumor of Bone. Electron-Microscopic Evidence of Its Acinar Cell Origin. Cancer 1968, 22, 333–344. [Google Scholar] [CrossRef]
- Luchini, C.; Cros, J.; Pea, A.; Pilati, C.; Veronese, N.; Rusev, B.; Capelli, P.; Mafficini, A.; Nottegar, A.; Brosens, L.A.A.; et al. PD-1, PD-L1, and CD163 in Pancreatic Undifferentiated Carcinoma with Osteoclast-like Giant Cells: Expression Patterns and Clinical Implications. Hum. Pathol. 2018, 81, 157–165. [Google Scholar] [CrossRef]
- Bazzichetto, C.; Luchini, C.; Conciatori, F.; Vaccaro, V.; Di Cello, I.; Mattiolo, P.; Falcone, I.; Ferretti, G.; Scarpa, A.; Cognetti, F.; et al. Morphologic and Molecular Landscape of Pancreatic Cancer Variants as the Basis of New Therapeutic Strategies for Precision Oncology. Int. J. Mol. Sci. 2020, 21, 8841. [Google Scholar] [CrossRef]
- Mugaanyi, J.; Lu, C.; Huang, J.; Lu, C. Undifferentiated Pancreatic Carcinomas, Clinical Features and Therapeutic Options: What We Know. Cancers 2022, 14, 6102. [Google Scholar] [CrossRef]
- Hrudka, J.; Kalinová, M.; Ciprová, V.; Moravcová, J.; Dvořák, R.; Matěj, R. Undifferentiated Carcinoma with Osteoclast-like Giant Cells of the Pancreas: Molecular Genetic Analysis of 13 Cases. Int. J. Mol. Sci. 2024, 25, 3285. [Google Scholar] [CrossRef]
- Ali Syeda, Z.; Langden, S.S.S.; Munkhzul, C.; Lee, M.; Song, S.J. Regulatory Mechanism of MicroRNA Expression in Cancer. Int. J. Mol. Sci. 2020, 21, 1723. [Google Scholar] [CrossRef]
- Shang, R.; Lee, S.; Senavirathne, G.; Lai, E.C. microRNAs in Action: Biogenesis, Function and Regulation. Nat. Rev. Genet. 2023, 24, 816–833. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed]
- Sassen, S.; Miska, E.A.; Caldas, C. MicroRNA—Implications for Cancer. Virchows Archiv. 2008, 452, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Iorio, M.V.; Croce, C.M. MicroRNA Dysregulation in Cancer: Diagnostics, Monitoring and Therapeutics. A Comprehensive Review. EMBO Mol. Med. 2012, 4, 143–159. [Google Scholar] [CrossRef]
- Moriyama, T.; Ohuchida, K.; Mizumoto, K.; Yu, J.; Sato, N.; Nabae, T.; Takahata, S.; Toma, H.; Nagai, E.; Tanaka, M. MicroRNA-21 Modulates Biological Functions of Pancreatic Cancer Cells Including Their Proliferation, Invasion, and Chemoresistance. Mol. Cancer Ther. 2009, 8, 1067–1074. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Tao, F.; Wang, W.; Ji, K. Prognostic Value of microRNA-21 in Pancreatic Ductal Adenocarcinoma: A Meta-Analysis. World J. Surg. Oncol. 2016, 14, 82. [Google Scholar] [CrossRef] [PubMed]
- Daoud, A.Z.; Mulholland, E.J.; Cole, G.; McCarthy, H.O. MicroRNAs in Pancreatic Cancer: Biomarkers, Prognostic, and Therapeutic Modulators. BMC Cancer 2019, 19, 1130. [Google Scholar] [CrossRef] [PubMed]
- Szabo, A.; Gurlich, R.; Liberko, M.; Soumarova, R.; Vernerova, Z.; Mandys, V.; Popov, A. Expression of Selected microRNAs in Pancreatic Ductal Adenocarcinoma: Is There a Relation to Tumor Morphology, Progression and Patient’s Outcome? Neoplasma 2020, 67, 1170–1181. [Google Scholar] [CrossRef] [PubMed]
- Popov, A.; Mandys, V. Senescence-Associated miRNAs and Their Role in Pancreatic Cancer. Pathol. Oncol. Res. 2022, 28, 1610156. [Google Scholar] [CrossRef] [PubMed]
- Prinz, C.; Fehring, L.; Frese, R. MicroRNAs as Indicators of Malignancy in Pancreatic Ductal Adenocarcinoma (PDAC) and Cystic Pancreatic Lesions. Cells 2022, 11, 2374. [Google Scholar] [CrossRef] [PubMed]
- Shenouda, S.K.; Alahari, S.K. MicroRNA Function in Cancer: Oncogene or a Tumor Suppressor? Cancer Metastasis Rev. 2009, 28, 369–378. [Google Scholar] [CrossRef]
- Zhang, B.; Pan, X.; Cobb, G.; Anderson, T. microRNAs as Oncogenes and Tumor Suppressors. Dev. Biol. 2007, 302, 1–12. [Google Scholar] [CrossRef]
- Eun, J.L.; Gusev, Y.; Jiang, J.; Nuovo, G.J.; Lerner, M.R.; Frankel, W.L.; Morgan, D.L.; Postier, R.G.; Brackett, D.J.; Schmittgen, T.D. Expression Profiling Identifies microRNA Signature in Pancreatic Cancer. Int. J. Cancer 2007, 120, 1046–1054. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, M.; Wang, H.; Fisher, W.E.; Lin, P.H.; Yao, Q.; Chen, C. Profiling of 95 MicroRNAs in Pancreatic Cancer Cell Lines and Surgical Specimens by Real-Time PCR Analysis. World J. Surg. 2009, 33, 698–709. [Google Scholar] [CrossRef] [PubMed]
- Negoi, I.; Hostiuc, S.; Sartelli, M.; Negoi, R.I.; Beuran, M. MicroRNA-21 as a Prognostic Biomarker in Patients with Pancreatic Cancer—A Systematic Review and Meta-Analysis. Am. J. Surg. 2017, 214, 515–524. [Google Scholar] [CrossRef]
- Jamieson, N.B.; Morran, D.C.; Morton, J.P.; Ali, A.; Dickson, E.J.; Carter, C.R.; Sansom, O.J.; Evans, T.R.J.; McKay, C.J.; Oien, K.A. MicroRNA Molecular Profiles Associated with Diagnosis, Clinicopathologic Criteria, and Overall Survival in Patients with Resectable Pancreatic Ductal Adenocarcinoma. Clin. Cancer Res. 2012, 18, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Du, Y.; Wang, G.; Gao, J.; Gong, Y.; Li, L.; Zhang, Z.; Zhu, J.; Jing, Q.; Qin, Y.; et al. Detection of Differentially Expressed microRNAs in Serum of Pancreatic Ductal Adenocarcinoma Patients: miR-196a Could Be a Potential Marker for Poor Prognosis. Dig. Dis. Sci. 2011, 56, 602–609. [Google Scholar] [CrossRef]
- Papaconstantinou, I.G.; Manta, A.; Gazouli, M.; Lyberopoulou, A.; Lykoudis, P.M.; Polymeneas, G.; Voros, D. Expression of microRNAs in Patients with Pancreatic Cancer and Its Prognostic Significance. Pancreas 2013, 42, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Bloomston, M.; Frankel, W.L.; Petrocca, F.; Volinia, S.; Alder, H.; Hagan, J.P.; Liu, C.G.; Bhatt, D.; Taccioli, C.; Croce, C.M. MicroRNA Expression Patterns to Differentiate Pancreatic Adenocarcinoma from Normal Pancreas and Chronic Pancreatitis. JAMA 2007, 297, 1901–1908. [Google Scholar] [CrossRef] [PubMed]
- Hong, T.H.; Park, I.Y. MicroRNA Expression Profiling of Diagnostic Needle Aspirates from Surgical Pancreatic Cancer Specimens. Ann. Surg. Treat. Res. 2014, 87, 290. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, J.; Chang, P.; LeBlanc, A.; Li, D.; Abbruzzesse, J.L.; Frazier, M.L.; Killary, A.M.; Sen, S. MicroRNAs in Plasma of Pancreatic Ductal Adenocarcinoma Patients as Novel Blood-Based Biomarkers of Disease. Cancer Prev. Res. 2009, 2, 807–813. [Google Scholar] [CrossRef]
- Greither, T.; Grochola, L.F.; Udelnow, A.; Lautenschläger, C.; Würl, P.; Taubert, H. Elevated Expression of microRNAs 155, 203, 210 and 222 in Pancreatic Tumors Is Associated with Poorer Survival. Int. J. Cancer 2010, 126, 73–80. [Google Scholar] [CrossRef]
- Delpu, Y.; Lulka, H.; Sicard, F.; Saint-Laurent, N.; Lopez, F.; Hanoun, N.; Buscail, L.; Cordelier, P.; Torrisani, J. The Rescue of miR-148a Expression in Pancreatic Cancer: An Inappropriate Therapeutic Tool. PLoS ONE 2013, 8, e55513. [Google Scholar] [CrossRef] [PubMed]
- Feng, J.; Yu, J.; Pan, X.; Li, Z.; Chen, Z.; Zhang, W.; Wang, B.; Yang, L.; Xu, H.; Zhang, G.; et al. HERG1 Functions as an Oncogene in Pancreatic Cancer and Is Downregulated by miR-96. Oncotarget 2014, 5, 5832–5844. [Google Scholar] [CrossRef]
- Hrudka, J.; Lawrie, K.; Waldauf, P.; Ciprová, V.; Moravcová, J.; Matěj, R. Negative Prognostic Impact of PD-L1 Expression in Tumor Cells of Undifferentiated (Anaplastic) Carcinoma with Osteoclast-like Giant Cells of the Pancreas: Study of 13 Cases Comparing Ductal Pancreatic Carcinoma and Review of the Literature. Virchows Arch. 2020, 477, 687–696. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A New Mathematical Model for Relative Quantification in Real-Time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Du Rieu, M.C.; Torrisani, J.; Selves, J.; Al Saati, T.; Souque, A.; Dufresne, M.; Tsongalis, G.J.; Suriawinata, A.A.; Carrère, N.; Buscail, L.; et al. MicroRNA-21 Is Induced Early in Pancreatic Ductal Adenocarcinoma Precursor Lesions. Clin. Chem. 2010, 56, 603–612. [Google Scholar] [CrossRef]
- Qi, L.; Bart, J.; Tan, L.P.; Platteel, I.; van der Sluis, T.; Huitema, S.; Harms, G.; Fu, L.; Hollema, H.; van den Berg, A. Expression of miR-21 and Its Targets (PTEN, PDCD4, TM1) in Flat Epithelial Atypia of the Breast in Relation to Ductal Carcinoma in Situ and Invasive Carcinoma. BMC Cancer 2009, 9, 163. [Google Scholar] [CrossRef]
- Habbe, N.; Koorstra, J.B.M.; Mendell, J.T.; Offerhaus, G.J.; Ji, K.R.; Feldmann, G.; Mullendore, M.E.; Goggins, M.G.; Hong, S.M.; Maitra, A. MicroRNA miR-155 Is a Biomarker of Early Pancreatic Neoplasia. Cancer Biol. Ther. 2009, 8, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Gironella, M.; Seux, M.; Xie, M.-J.; Cano, C.; Tomasini, R.; Gommeaux, J.; Garcia, S.; Nowak, J.; Yeung, M.L.; Jeang, K.-T.; et al. Tumor Protein 53-Induced Nuclear Protein 1 Expression Is Repressed by miR-155, and Its Restoration Inhibits Pancreatic Tumor Development. Proc. Natl. Acad. Sci. USA 2007, 104, 16170–16175. [Google Scholar] [CrossRef]
- Yu, S.; Lu, Z.; Liu, C.; Meng, Y.; Ma, Y.; Zhao, W.; Liu, J.; Yu, J.; Chen, J. miRNA-96 Suppresses KRAS and Functions as a Tumor Suppressor Gene in Pancreatic Cancer. Cancer Res. 2010, 70, 6015–6025. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.-G.; Yu, S.-N.; Lu, Z.-H.; Ma, Y.-H.; Gu, Y.-M.; Chen, J. The miR-217 microRNA Functions as a Potential Tumor Suppressor in Pancreatic Ductal Adenocarcinoma by Targeting KRAS. Carcinogenesis 2010, 31, 1726–1733. [Google Scholar] [CrossRef]
- Szafranska, A.E.; Davison, T.S.; John, J.; Cannon, T.; Sipos, B.; Maghnouj, A.; Labourier, E.; Hahn, S.A. MicroRNA Expression Alterations Are Linked to Tumorigenesis and Non-Neoplastic Processes in Pancreatic Ductal Adenocarcinoma. Oncogene 2007, 26, 4442–4452. [Google Scholar] [CrossRef]
- Szafranska, A.E.; Doleshal, M.; Edmunds, H.S.; Gordon, S.; Luttges, J.; Munding, J.B.; Barth, R.J.; Gutmann, E.J.; Suriawinata, A.A.; Pipas, J.M.; et al. Analysis of microRNAs in Pancreatic Fine-Needle Aspirates Can Classify Benign and Malignant Tissues. Clin. Chem. 2008, 54, 1716–1724. [Google Scholar] [CrossRef]
- Link, A.; Becker, V.; Goel, A.; Wex, T.; Malfertheiner, P. Feasibility of Fecal microRNAs as Novel Biomarkers for Pancreatic Cancer. PLoS ONE 2012, 7, e42933. [Google Scholar] [CrossRef]
- Molberg, K.H.; Heffess, C.; Delgado, R.; Albores-Saavedra, J. Undifferentiated Carcinoma with Osteoclast-like Giant Cells of the Pancreas and Periampullary Region. Cancer 1998, 82, 1279–1287. [Google Scholar] [CrossRef]
- Paal, E.; Thompson, L.D.; Frommelt, R.A.; Przygodzki, R.M.; Heffess, C.S. A Clinicopathologic and Immunohistochemical Study of 35 Anaplastic Carcinomas of the Pancreas with a Review of the Literature. Ann. Diagn. Pathol. 2001, 5, 129–140. [Google Scholar] [CrossRef]
- Rosai and Ackerman’s Surgical Pathology—2 Volume Set—11th Edition. Available online: https://www.elsevier.com/books/rosai-and-ackermans-surgical-pathology-2-volume-set/goldblum/978-0-323-26339-9 (accessed on 23 May 2023).
- Dworak, O.; Wittekind, C.; Koerfgen, H.P.; Gall, F.P. Osteoclastic Giant Cell Tumor of the Pancreas. An Immunohistological Study and Review of the Literature. Pathol. Res. Pract. 1993, 189, 228–234. [Google Scholar] [CrossRef]
- Reid, M.D.; Muraki, T.; HooKim, K.; Memis, B.; Graham, R.P.; Allende, D.; Shi, J.; Schaeffer, D.F.; Singh, R.; Basturk, O.; et al. Cytologic Features and Clinical Implications of Undifferentiated Carcinoma with Osteoclastic Giant Cells of the Pancreas: An Analysis of 15 Cases. Cancer Cytopathol. 2017, 125, 563–575. [Google Scholar] [CrossRef]
- Lehrke, H.D.; Graham, R.P.; McWilliams, R.R.; Lam-Himlin, D.M.; Smyrk, T.C.; Jenkins, S.; Dong, H.; Zhang, L. Undifferentiated Pancreatic Carcinomas Display Enrichment for Frequency and Extent of PD-L1 Expression by Tumor Cells. Am. J. Clin. Pathol. 2017, 148, 441–449. [Google Scholar] [CrossRef]
- Besaw, R.J.; Terra, A.R.; Malvar, G.L.; Chapman, T.R.; Hertan, L.M.; Schlechter, B.L. Durable Response to PD-1 Blockade in a Patient with Metastatic Pancreatic Undifferentiated Carcinoma with Osteoclast-Like Giant Cells. J. Natl. Compr. Canc. Netw. 2021, 19, 247–252. [Google Scholar] [CrossRef]
- Obayashi, M.; Shibasaki, Y.; Koakutsu, T.; Hayashi, Y.; Shoji, T.; Hirayama, K.; Yamazaki, M.; Takayanagi, Y.; Shibata, H.; Nakamura, M.; et al. Pancreatic Undifferentiated Carcinoma with Osteoclast-like Giant Cells Curatively Resected after Pembrolizumab Therapy for Lung Metastases: A Case Report. BMC Gastroenterol. 2020, 20, 220. [Google Scholar] [CrossRef]
- Mattiolo, P.; Fiadone, G.; Paolino, G.; Chatterjee, D.; Bernasconi, R.; Piccoli, P.; Parolini, C.; El Aidi, M.; Sperandio, N.; Malleo, G.; et al. Epithelial-Mesenchymal Transition in Undifferentiated Carcinoma of the Pancreas with and without Osteoclast-like Giant Cells. Virchows Arch. 2021, 478, 319–326. [Google Scholar] [CrossRef]
- Sakakida, T.; Ishikawa, T.; Doi, T.; Morita, R.; Kataoka, S.; Miyake, H.; Yamaguchi, K.; Moriguchi, M.; Sogame, Y.; Yasuda, H.; et al. Genomic Landscape and Clinical Features of Rare Subtypes of Pancreatic Cancer: Analysis with the National Database of Japan. J. Gastroenterol. 2023, 58, 575–585. [Google Scholar] [CrossRef]
- Bergmann, F.; Moldenhauer, G.; Herpel, E.; Gaida, M.M.; Strobel, O.; Werner, J.; Esposito, I.; Müerköster, S.S.; Schirmacher, P.; Kern, M.A. Expression of L1CAM, COX-2, EGFR, c-KIT and Her2/Neu in Anaplastic Pancreatic Cancer: Putative Therapeutic Targets? Histopathology 2010, 56, 440–448. [Google Scholar] [CrossRef]
- Wang, X.; Miao, J.; Wang, S.; Shen, R.; Zhang, S.; Tian, Y.; Li, M.; Zhu, D.; Yao, A.; Bao, W.; et al. Single-Cell RNA-Seq Reveals the Genesis and Heterogeneity of Tumor Microenvironment in Pancreatic Undifferentiated Carcinoma with Osteoclast-like Giant-Cells. Mol. Cancer 2022, 21, 133. [Google Scholar] [CrossRef]
- Nakata, K.; Ohuchida, K.; Mizumoto, K.; Kayashima, T.; Ikenaga, N.; Sakai, H.; Lin, C.; Fujita, H.; Otsuka, T.; Aishima, S.; et al. MicroRNA-10b Is Overexpressed in Pancreatic Cancer, Promotes Its Invasiveness, and Correlates with a Poor Prognosis. Surgery 2011, 150, 916–922. [Google Scholar] [CrossRef]
- Park, J.-K.; Lee, E.J.; Esau, C.; Schmittgen, T.D. Antisense Inhibition of microRNA-21 or -221 Arrests Cell Cycle, Induces Apoptosis, and Sensitizes the Effects of Gemcitabine in Pancreatic Adenocarcinoma. Pancreas 2009, 38, e190. [Google Scholar] [CrossRef]
- Wang, P.; Zhu, C.; Ma, M.; Chen, G.; Song, M.; Zeng, Z.; Lu, W.; Yang, J.; Wen, S.; Chiao, P.J.; et al. Micro-RNA-155 Is Induced by K-Ras Oncogenic Signal and Promotes ROS Stress in Pancreatic Cancer. Oncotarget 2015, 6, 21148–21158. [Google Scholar] [CrossRef]
- Fasanaro, P.; D’Alessandra, Y.; Di Stefano, V.; Melchionna, R.; Romani, S.; Pompilio, G.; Capogrossi, M.C.; Martelli, F. MicroRNA-210 Modulates Endothelial Cell Response to Hypoxia and Inhibits the Receptor Tyrosine Kinase Ligand Ephrin-A3. J. Biol. Chem. 2008, 283, 15878–15883. [Google Scholar] [CrossRef]
- Zhang, Z.; Sun, H.; Dai, H.; Walsh, R.M.; Imakura, M.; Schelter, J.; Burchard, J.; Dai, X.; Chang, A.N.; Diaz, R.L.; et al. MicroRNA miR-210 Modulates Cellular Response to Hypoxia through the MYC Antagonist MNT. Cell Cycle 2009, 8, 2756–2768. [Google Scholar] [CrossRef]
- Huang, X.; Le, Q.-T.; Giaccia, A.J. MiR-210–Micromanager of the Hypoxia Pathway. Trends Mol. Med. 2010, 16, 230–237. [Google Scholar] [CrossRef]
- Buscaglia, L.E.B.; Li, Y. Apoptosis and the Target Genes of microRNA-21. Chin. J. Cancer 2011, 30, 371–380. [Google Scholar] [CrossRef]
- Huang, C.; Li, H.; Wu, W.; Jiang, T.; Qiu, Z. Regulation of miR-155 Affects Pancreatic Cancer Cell Invasiveness and Migration by Modulating the STAT3 Signaling Pathway through SOCS1. Oncol. Rep. 2013, 30, 1223–1230. [Google Scholar] [CrossRef]
- Nakamura, S.; Sadakari, Y.; Ohtsuka, T.; Okayama, T.; Nakashima, Y.; Gotoh, Y.; Saeki, K.; Mori, Y.; Nakata, K.; Miyasaka, Y.; et al. Pancreatic Juice Exosomal MicroRNAs as Biomarkers for Detection of Pancreatic Ductal Adenocarcinoma. Ann. Surg. Oncol. 2019, 26, 2104–2111. [Google Scholar] [CrossRef]
- Huang, F.; Tang, J.; Zhuang, X.; Zhuang, Y.; Cheng, W.; Chen, W.; Yao, H.; Zhang, S. MiR-196a Promotes Pancreatic Cancer Progression by Targeting Nuclear Factor Kappa-B-Inhibitor Alpha. PLoS ONE 2014, 9, e87897. [Google Scholar] [CrossRef]
- Giovannetti, E.; Funel, N.; Peters, G.J.; Del Chiaro, M.; Erozenci, L.A.; Vasile, E.; Leon, L.G.; Pollina, L.E.; Groen, A.; Falcone, A.; et al. MicroRNA-21 in Pancreatic Cancer: Correlation with Clinical Outcome and Pharmacologic Aspects Underlying Its Role in the Modulation of Gemcitabine Activity. Cancer Res. 2010, 70, 4528–4538. [Google Scholar] [CrossRef]
- Huang, X.; Zuo, J. Emerging Roles of miR-210 and Other Non-Coding RNAs in the Hypoxic Response. Acta Biochim. Et Biophys. Sin. 2014, 46, 220–232. [Google Scholar] [CrossRef]
- Zhao, Q.; Chen, S.; Zhu, Z.; Yu, L.; Ren, Y.; Jiang, M.; Weng, J.; Li, B. miR-21 Promotes EGF-Induced Pancreatic Cancer Cell Proliferation by Targeting Spry2. Cell Death Dis. 2018, 9, 1157. [Google Scholar] [CrossRef]
- Mok, E.T.Y.; Chitty, J.L.; Cox, T.R. miRNAs in Pancreatic Cancer Progression and Metastasis. Clin. Exp. Metastasis 2024. [Google Scholar] [CrossRef]
- Wang, P.; Zhuang, L.; Zhang, J.; Fan, J.; Luo, J.; Chen, H.; Wang, K.; Liu, L.; Chen, Z.; Meng, Z. The Serum miR-21 Level Serves as a Predictor for the Chemosensitivity of Advanced Pancreatic Cancer, and miR-21 Expression Confers Chemoresistance by Targeting FasL. Mol. Oncol. 2013, 7, 334. [Google Scholar] [CrossRef]
- Karásek, P.; Gablo, N.; Hlavsa, J.; Kiss, I.; Vychytilová-Faltejsková, P.; Hermanová, M.; Kala, Z.; Slabý, O.; Procházka, V. Pre-Operative Plasma miR-21-5p Is a Sensitive Biomarker and Independent Prognostic Factor in Patients with Pancreatic Ductal Adenocarcinoma Undergoing Surgical Resection. Cancer Genom. Proteom. 2018, 15, 321–327. [Google Scholar] [CrossRef]
- Khan, K.; Cunningham, D.; Peckitt, C.; Barton, S.; Tait, D.; Hawkins, M.; Watkins, D.; Starling, N.; Rao, S.; Begum, R.; et al. miR-21 Expression and Clinical Outcome in Locally Advanced Pancreatic Cancer: Exploratory Analysis of the Pancreatic Cancer Erbitux, Radiotherapy and UFT (PERU) Trial. Oncotarget 2016, 7, 12672–12681. [Google Scholar] [CrossRef] [PubMed]
- Nishiwada, S.; Sho, M.; Banwait, J.K.; Yamamura, K.; Akahori, T.; Nakamura, K.; Baba, H.; Goel, A. A MicroRNA Signature Identifies Pancreatic Ductal Adenocarcinoma Patients at Risk for Lymph Node Metastases. Gastroenterology 2020, 159, 562–574. [Google Scholar] [CrossRef] [PubMed]
- Mikamori, M.; Yamada, D.; Eguchi, H.; Hasegawa, S.; Kishimoto, T.; Tomimaru, Y.; Asaoka, T.; Noda, T.; Wada, H.; Kawamoto, K.; et al. MicroRNA-155 Controls Exosome Synthesis and Promotes Gemcitabine Resistance in Pancreatic Ductal Adenocarcinoma. Sci. Rep. 2017, 7, 42339. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Zubair, H.; Srivastava, S.K.; Singh, S.; Singh, A.P. Insights into the Role of microRNAs in Pancreatic Cancer Pathogenesis: Potential for Diagnosis, Prognosis, and Therapy. Adv. Exp. Med. Biol. 2015, 889, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Pang, W.; Su, J.; Wang, Y.; Feng, H.; Dai, X.; Yuan, Y.; Chen, X.; Yao, W. Pancreatic Cancer-Secreted miR-155 Implicates in the Conversion from Normal Fibroblasts to Cancer-Associated Fibroblasts. Cancer Sci. 2015, 106, 1362–1369. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Hong, Q.; Lu, F.; Zhang, Z.; Li, J.; Nie, Z.; He, B. The Diagnostic and Prognostic Value of miR-155 in Cancers: An Updated Meta-Analysis. Mol. Diagn. Ther. 2023, 27, 283–301. [Google Scholar] [CrossRef]
- Kim, M.W.; Koh, H.; Kim, J.Y.; Lee, S.; Lee, H.; Kim, Y.; Hwang, H.K.; Kim, S.I. Tumor-Specific miRNA Signatures in Combination with CA19-9 for Liquid Biopsy-Based Detection of PDAC. Int. J. Mol. Sci. 2021, 22, 13621. [Google Scholar] [CrossRef]
- Wang, J.; Raimondo, M.; Guha, S.; Chen, J.; Diao, L.; Dong, X.; Wallace, M.B.; Killary, A.M.; Frazier, M.L.; Woodward, T.A.; et al. Circulating microRNAs in Pancreatic Juice as Candidate Biomarkers of Pancreatic Cancer. J. Cancer 2014, 5, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Lian, M.; Mortoglou, M.; Uysal-Onganer, P. Impact of Hypoxia-Induced miR-210 on Pancreatic Cancer. Curr. Issues Mol. Biol. 2023, 45, 9778–9792. [Google Scholar] [CrossRef]
- Wu, G.; Ding, X.; Quan, G.; Xiong, J.; Li, Q.; Li, Z.; Wang, Y. Hypoxia-Induced miR-210 Promotes Endothelial Cell Permeability and Angiogenesis via Exosomes in Pancreatic Ductal Adenocarcinoma. Biochem. Res. Int. 2022, 2022, 7752277. [Google Scholar] [CrossRef]
- Yu, Q.; Xu, C.; Yuan, W.; Wang, C.; Zhao, P.; Chen, L.; Ma, J. Evaluation of Plasma MicroRNAs as Diagnostic and Prognostic Biomarkers in Pancreatic Adenocarcinoma: miR-196a and miR-210 Could Be Negative and Positive Prognostic Markers, Respectively. BioMed Res. Int. 2017, 2017, 6495867. [Google Scholar] [CrossRef] [PubMed]
- Wnuk, J.; Strzelczyk, J.K.; Gisterek, I. Clinical Value of Circulating miRNA in Diagnosis, Prognosis, Screening and Monitoring Therapy of Pancreatic Ductal Adenocarcinoma–A Review of the Literature. Int. J. Mol. Sci. 2023, 24, 5113. [Google Scholar] [CrossRef] [PubMed]
- Luthra, R.; Singh, R.R.; Luthra, M.G.; Li, Y.X.; Hannah, C.; Romans, A.M.; Barkoh, B.A.; Chen, S.S.; Ensor, J.; Maru, D.M.; et al. MicroRNA-196a Targets Annexin A1: A microRNA-Mediated Mechanism of Annexin A1 Downregulation in Cancers. Oncogene 2008, 27, 6667–6678. [Google Scholar] [CrossRef]
- Lee, Y.S.; Kim, H.; Kim, H.W.; Lee, J.-C.; Paik, K.-H.; Kang, J.; Kim, J.; Yoon, Y.-S.; Han, H.-S.; Sohn, I.; et al. High Expression of MicroRNA-196a Indicates Poor Prognosis in Resected Pancreatic Neuroendocrine Tumor. Medicine 2015, 94, e2224. [Google Scholar] [CrossRef] [PubMed]
- Balzeau, J.; Menezes, M.R.; Cao, S.; Hagan, J.P. The LIN28/Let-7 Pathway in Cancer. Front. Genet. 2017, 8, 31. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chirshev, E.; Hojo, N.; Suzuki, T.; Bertucci, A.; Pierce, M.; Perry, C.; Wang, R.; Zink, J.; Glackin, C.A.; et al. The Epithelial–Mesenchymal Transcription Factor SNAI1 Represses Transcription of the Tumor Suppressor miRNA Let-7 in Cancer. Cancers 2021, 13, 1469. [Google Scholar] [CrossRef] [PubMed]
- Bhutia, Y.D.; Hung, S.W.; Krentz, M.; Patel, D.; Lovin, D.; Manoharan, R.; Thomson, J.M.; Govindarajan, R. Differential Processing of Let-7a Precursors Influences RRM2 Expression and Chemosensitivity in Pancreatic Cancer: Role of LIN-28 and SET Oncoprotein. PLoS ONE 2013, 8, e53436. [Google Scholar] [CrossRef] [PubMed]
- Idichi, T.; Seki, N.; Kurahara, H.; Fukuhisa, H.; Toda, H.; Shimonosono, M.; Okato, A.; Arai, T.; Kita, Y.; Mataki, Y.; et al. Molecular Pathogenesis of Pancreatic Ductal Adenocarcinoma: Impact of Passenger Strand of Pre-miR-148a on Gene Regulation. Cancer Sci. 2018, 109, 2013–2026. [Google Scholar] [CrossRef]
- Hanoun, N.; Delpu, Y.; Suriawinata, A.A.; Bournet, B.; Bureau, C.; Selves, J.; Tsongalis, G.J.; Dufresne, M.; Buscail, L.; Cordelier, P.; et al. The Silencing of MicroRNA 148a Production by DNA Hypermethylation Is an Early Event in Pancreatic Carcinogenesis. Clin. Chem. 2010, 56, 1107–1118. [Google Scholar] [CrossRef]
- Deng, S.; Zhu, S.; Wang, B.; Li, X.; Liu, Y.; Qin, Q.; Gong, Q.; Niu, Y.; Xiang, C.; Chen, J.; et al. Chronic Pancreatitis and Pancreatic Cancer Demonstrate Active Epithelial-Mesenchymal Transition Profile, Regulated by miR-217-SIRT1 Pathway. Cancer Lett. 2014, 355, 184–191. [Google Scholar] [CrossRef]
- Chang, X.; Yu, C.; Li, J.; Yu, S.; Chen, J. Hsa-miR-96 and Hsa-miR-217 Expression Down-Regulates with Increasing Dysplasia in Pancreatic Intraepithelial Neoplasias and Intraductal Papillary Mucinous Neoplasms. Int. J. Med. Sci. 2017, 14, 412–418. [Google Scholar] [CrossRef] [PubMed]
- Lemberger, M.; Loewenstein, S.; Lubezky, N.; Nizri, E.; Pasmanik-Chor, M.; Barazovsky, E.; Klausner, J.M.; Lahat, G. MicroRNA Profiling of Pancreatic Ductal Adenocarcinoma (PDAC) Reveals Signature Expression Related to Lymph Node Metastasis. Oncotarget 2019, 10, 2644–2656. [Google Scholar] [CrossRef] [PubMed]
- Kent, O.A.; Mullendore, M.; Wentzel, E.A.; López-Romero, P.; Tan, A.C.; Alvarez, H.; West, K.; Ochs, M.F.; Hidalgo, M.; Arking, D.E.; et al. A Resource for Analysis of microRNA Expression and Function in Pancreatic Ductal Adenocarcinoma Cells. Cancer Biol. Ther. 2009, 8, 2013–2024. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, Y.; Li, Z.; Hou, T. miR-155 Promotes Proliferation and Epithelial-Mesenchymal Transition of MCF-7 Cells. Exp. Ther. Med. 2021, 21, 218. [Google Scholar] [CrossRef] [PubMed]
- Kong, W.; Yang, H.; He, L.; Zhao, J.; Coppola, D.; Dalton, W.S.; Cheng, J.Q. MicroRNA-155 Is Regulated by the Transforming Growth Factor Beta/Smad Pathway and Contributes to Epithelial Cell Plasticity by Targeting RhoA. Mol. Cell. Biol. 2008, 28, 6773–6784. [Google Scholar] [CrossRef] [PubMed]
- Kong, X.; Liu, F.; Gao, J. MiR-155 Promotes Epithelial-Mesenchymal Transition in Hepatocellular Carcinoma Cells through the Activation of PI3K/SGK3/β-Catenin Signaling Pathways. Oncotarget 2016, 7, 66051–66060. [Google Scholar] [CrossRef]
- Kulkarni, P.; Dasgupta, P.; Hashimoto, Y.; Shiina, M.; Shahryari, V.; Tabatabai, Z.L.; Yamamura, S.; Tanaka, Y.; Saini, S.; Dahiya, R.; et al. A lncRNA TCL6-miR-155 Interaction Regulates the Src-Akt-EMT Network to Mediate Kidney Cancer Progression and Metastasis. Cancer Res. 2021, 81, 1500–1512. [Google Scholar] [CrossRef]


{kind=link}
| miRNA Name: | Stem–Loop Primer Sequence: |
|---|---|
| miR-39 C. elegans | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACT- GGATACGACTATTAC |
| mir-21 | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCAC- TGGATACGACTCAACA |
| miR-96 | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATA- CGACAGCAAAAATGTG |
| miR-148a | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTG- GATACGACAGTCGGAG |
| miR-155 | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATA- CGACACCCCTATCACG |
| miR-196a | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATA- CGACCCCAACAACATG |
| miR-210 | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATAC- GACTCAGCCGCTGTC |
| miR-217 | GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATA- CGACTCCAATCAGTTC |
| Primer Name: | Primer Sequence: |
|---|---|
| Universal primer | ATCCAGTGCAGGGTCCGAGG |
| mir-39 C. elegans | GCGGCGGAGCTGATTTCGTCTTG |
| mir-21 | GCGGCGGTAGCTTATCAGACTG |
| miR-96 | GCGGCGGTTTGGCACTAGCAC |
| miR-148a | GCGGCGGAAAGTTCTGAGACACTCC |
| miR-155 | GCGGCGGTTAATGCTAATCGTG |
| miR-196a | GCGGCGGTAGGTAGTTTCATGTTG |
| miR-210 | GCGGCGGCTGTGCGTGTGACAG |
| ID | Clinical Data | miRNA | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Diagnosis | Sex | Age | Stage | Survival (Months) | Status | miR-21 | miR-96 | miR-148a | miR-155 | miR-196a | miR-210 | miR-217 | |
| 1 | UCOGC | M | 73 | pTXN1 | 4 | Dead | 13.25 | 0.42 | 0.48 | 6.34 | 1.48 | 45.88 | 0.06 |
| 2 | UCOGC | F | 76 | pT3N0 | 0.1 | Dead | 12.42 | 0.26 | 0.61 | 3.73 | 0.96 | 14.84 | 0.03 |
| 3 | UCOGC | M | 61 | pT3N0 | 9 | Dead | 10.92 | 0.56 | 0.39 | 3.35 | 0.91 | 4.49 | 0.026 |
| 4 | UCOGC | M | 53 | pT3N1 | 58 | Alive | 16.68 | 0.27 | 1.06 | 2.18 | 0.43 | 7.92 | 0.02 |
| 5 | UCOGC | M | 63 | pT1cN0 | 57 | Alive | 44.79 | 0.38 | 0.76 | 5.61 | 0.89 | 39.98 | 0.08 |
| 6 | UCOGC | M | 67 | pT3N1 | 1 | Dead | 7 | 1.99 | 0.79 | 1.42 | 0.96 | 4.06 | 0.17 |
| 7 | UCOGC + PDAC | M | 63 | pT2N1 | 6 | Dead | 15.79 | 0.92 | 0.17 | 16.63 | 0.44 | 90.7 | 0.07 |
| 8 | UCOGC + IPMN | F | 50 | pT1bN0 | 86 | Alive | 34.39 | 2.21 | 0.46 | 3.19 | 0.45 | 15.88 | 0.06 |
| 9 | UCOGC | M | 59 | pT2N0 | 168 | Dead | 11.62 | 6.22 | 2.12 | 1.69 | 1.14 | 4.39 | 0.05 |
| 10 | UCOGC + PDAC | M | 75 | pT3N1 | 6 | Dead | 19.39 | 0.14 | 0.37 | 4.92 | 0.75 | 3.99 | 0.01 |
| ID | Clinical Data | miRNA | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Diagnosis | Sex | Age | Stage | Survival (Months) | Status | miR-21 | miR-96 | miR-148a | miR-155 | miR-196a | miR-210 | miR-217 | |
| 1 | PDAC | M | 69 | pT3N1 | 23 | Dead | 7.83 | 0.456 | 0.68 | 3.46 | 1.372 | 11.17 | 0.025 |
| 2 | PDAC | M | 68 | pT3N1 | 18 | Dead | 13.16 | 0.824 | 1.4 | 16.62 | 0.797 | 28.07 | 0.105 |
| 3 | PDAC | M | 83 | pT3N1 | 29 | Dead | 3.76 | 0.396 | 0.847 | 2.27 | 0.55 | 3.9 | 0.083 |
| 4 | PDAC | F | 71 | pT3N1 | 7 | Dead | 11.11 | 0.663 | 1.27 | 10.58 | 1.069 | 7.96 | 0.031 |
| 5 | PDAC | M | 63 | pT3N1 | 45 | Dead | 53.51 | 1.45 | 0.137 | 80.67 | 1.51 | 26.88 | 0.001 |
| 6 | PDAC | M | 59 | pT3N1 | 15 | Dead | 8.08 | 0.55 | 0.178 | 0.86 | 1.04 | 3.61 | 0.006 |
| 7 | PDAC | M | 60 | pT3N1 | 9 | Dead | 4.61 | 1.91 | 2.16 | 7.74 | 1.205 | 9.44 | 0.075 |
| 8 | PDAC | F | 66 | pT3N1 | 23 | Dead | 12.28 | 1 | 0.493 | 9.18 | 1.28 | 13.81 | 0.125 |
| 9 | PDAC | M | 64 | pT3N1 | 12 | Dead | 5.9 | 0.41 | 0.116 | 54.26 | 0.996 | 41.98 | 0.002 |
| 10 | PDAC | F | 53 | pT4N1 | 11 | Dead | 14.96 | 0.76 | 0.232 | 232.36 | 1.09 | 5.9 | 0.031 |
| 11 | PDAC | M | 67 | pT3N0 | 27 | Dead | 28.43 | 1.497 | 0.519 | 121.05 | 1.42 | 21.35 | 0.236 |
| 12 | PDAC | F | 77 | pT3N0 | 9 | Dead | 35.4 | 0.792 | 0.053 | 10.56 | 1.4 | 65.34 | 0.103 |
| Cancer Type | miRNAs Fold Change | ||||||
|---|---|---|---|---|---|---|---|
| miR-21 | miR-96 | miR-148a | miR-155 | miR-196a | miR-210 | miR-217 | |
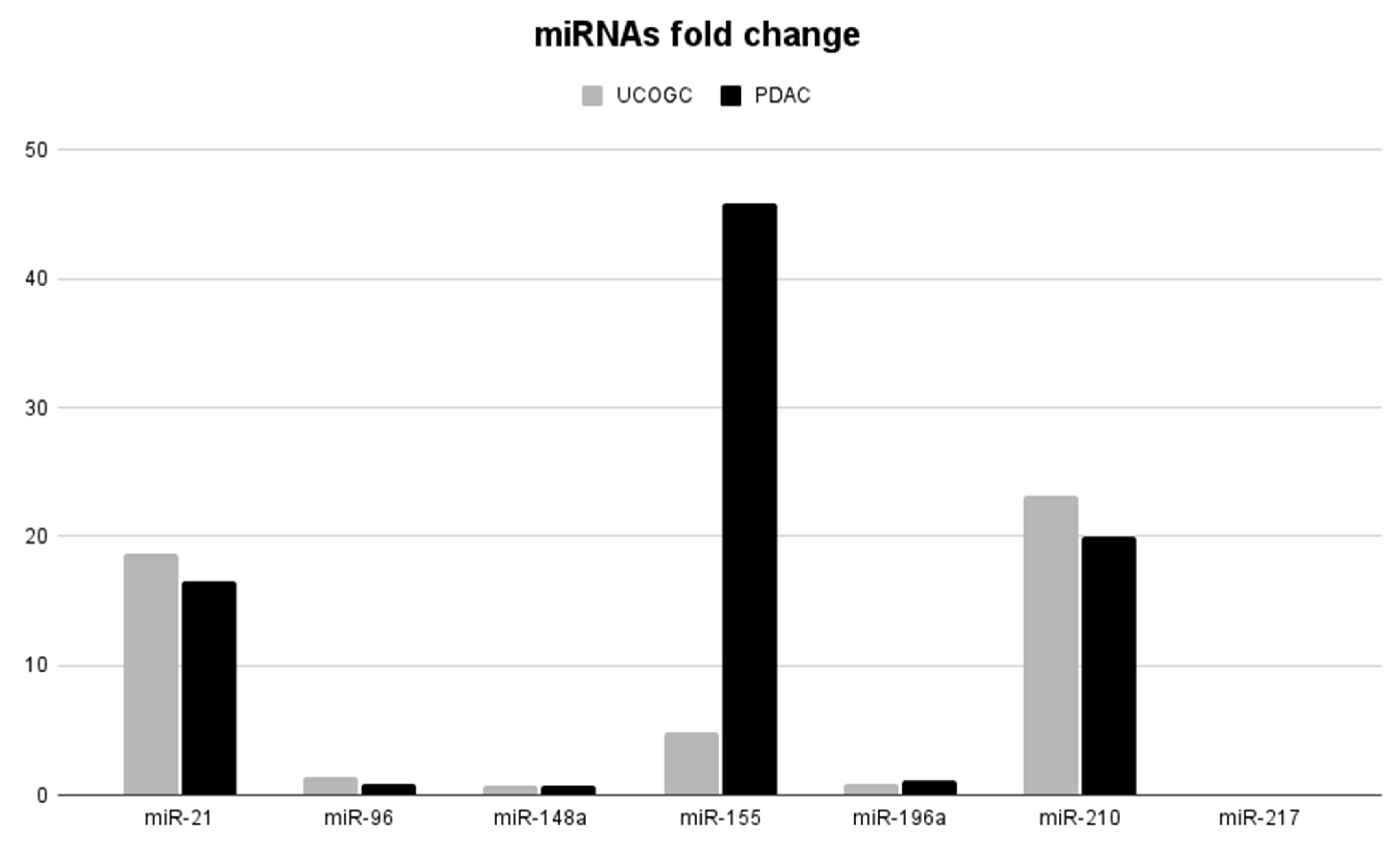
| UCOGC | 18.62± 8.539 | 1.34 ± 1.281 | 0.72± 0.369 | 4.91± 2.775 | 0.84± 0.259 | 23.21±21.384 | 0.05± 0.031 |
| PDAC | 16.58± 11.263 | 0.89 ± 0.381 | 0.67± 0.498 | 45.8± 50.856 | 1.14 ± 0.22 | 19.95± 13.977 | 0.06± 0.052 |
| Oncomirs (Upregulated) | miRNA Targets | miRNA Enhances | References |
|---|---|---|---|
| miR-21 | CDK2AP1, Pdcd, BCL2, PTEN, and almost 30 genes | proliferation, invasion, chemoresistance, tumor survival | [14,15,63,64] |
| miR-155 | TP53INP1, FOXO3a, and SOCS1 | proliferation, transformation, migration, invasion | [38,59,65] |
| miR-196a | NFKBIA | proliferation, migration, invasion | [66] |
| miR-210 | Ephrin-A3, MNT | proliferation, angiogenesis, tumor growth, survival | [26,67] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Popov, A.; Hrudka, J.; Szabó, A.; Oliverius, M.; Šubrt, Z.; Vránová, J.; Ciprová, V.; Moravcová, J.; Mandys, V. Expression of Selected miRNAs in Undifferentiated Carcinoma with Osteoclast-like Giant Cells (UCOGC) of the Pancreas: Comparison with Poorly Differentiated Pancreatic Ductal Adenocarcinoma. Biomedicines 2024, 12, 962. https://doi.org/10.3390/biomedicines12050962
Popov A, Hrudka J, Szabó A, Oliverius M, Šubrt Z, Vránová J, Ciprová V, Moravcová J, Mandys V. Expression of Selected miRNAs in Undifferentiated Carcinoma with Osteoclast-like Giant Cells (UCOGC) of the Pancreas: Comparison with Poorly Differentiated Pancreatic Ductal Adenocarcinoma. Biomedicines. 2024; 12(5):962. https://doi.org/10.3390/biomedicines12050962
Chicago/Turabian StylePopov, Alexey, Jan Hrudka, Arpád Szabó, Martin Oliverius, Zdeněk Šubrt, Jana Vránová, Vanda Ciprová, Jana Moravcová, and Václav Mandys. 2024. "Expression of Selected miRNAs in Undifferentiated Carcinoma with Osteoclast-like Giant Cells (UCOGC) of the Pancreas: Comparison with Poorly Differentiated Pancreatic Ductal Adenocarcinoma" Biomedicines 12, no. 5: 962. https://doi.org/10.3390/biomedicines12050962
APA StylePopov, A., Hrudka, J., Szabó, A., Oliverius, M., Šubrt, Z., Vránová, J., Ciprová, V., Moravcová, J., & Mandys, V. (2024). Expression of Selected miRNAs in Undifferentiated Carcinoma with Osteoclast-like Giant Cells (UCOGC) of the Pancreas: Comparison with Poorly Differentiated Pancreatic Ductal Adenocarcinoma. Biomedicines, 12(5), 962. https://doi.org/10.3390/biomedicines12050962