ACTN4 Mediates SEPT14 Mutation-Induced Sperm Head Defects

 , , ,
, , ,
Abstract
:1. Introduction
1.1. Male Infertility and Mutations
1.2. Septins and Male Reproduction
1.3. SEPT14
2. Materials and Methods
2.1. Separation and Isolation of Testicular Germ Cell Populations
2.2. Human Sperm Collection and Immunofluorescence Assay
2.3. Cloning and Transfection
2.4. Co-Immunoprecipitation
2.5. Mass Spectrometry Analysis
3. Results
3.1. Dynamic Expression of SEPT14 during Murine Sperm Head Formation
3.2. Identification of SEPT14 Interactors in Male Germ Cells
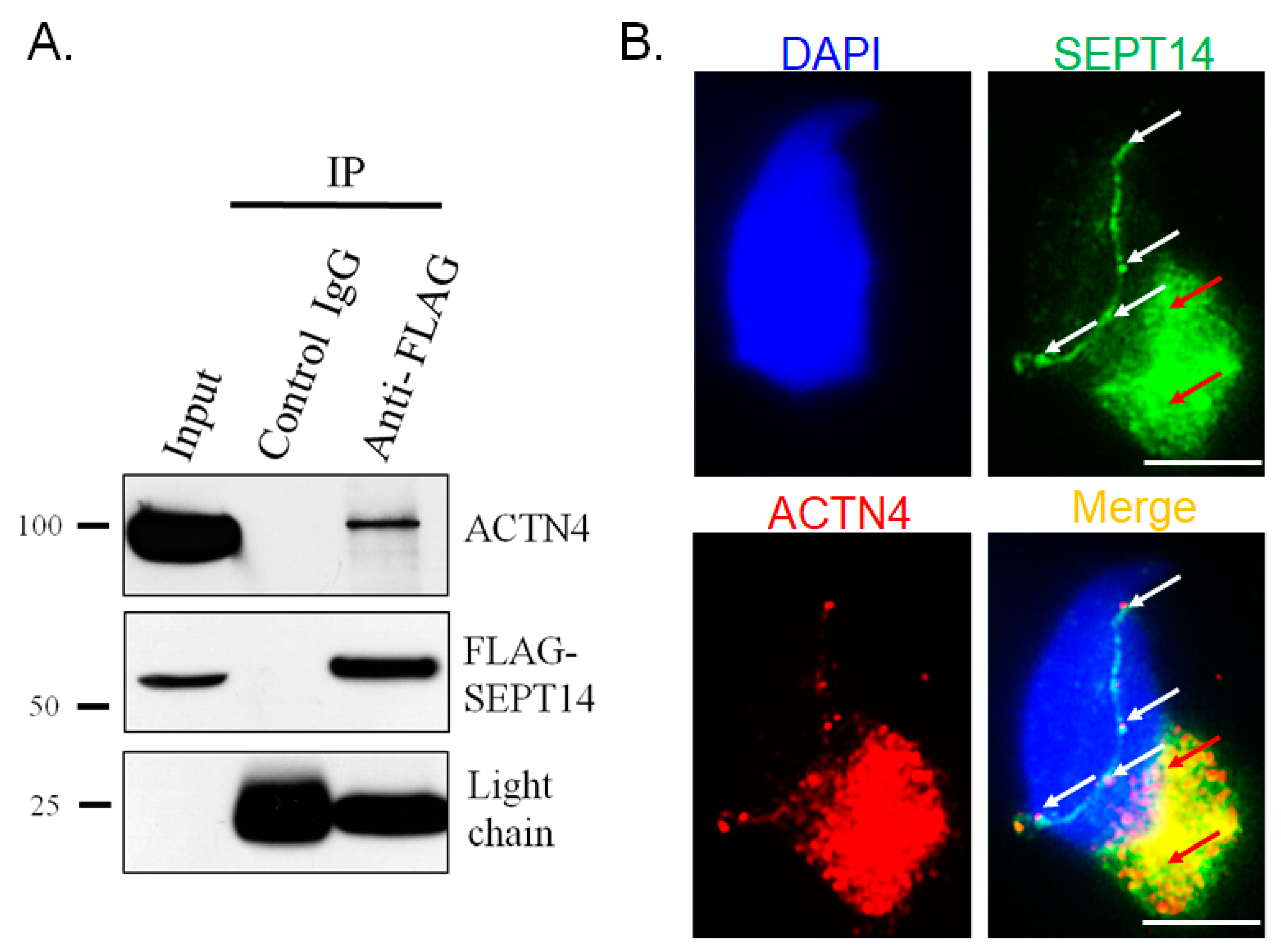
3.3. SEPT14 Interacts and Co-Localizes with ACTN4
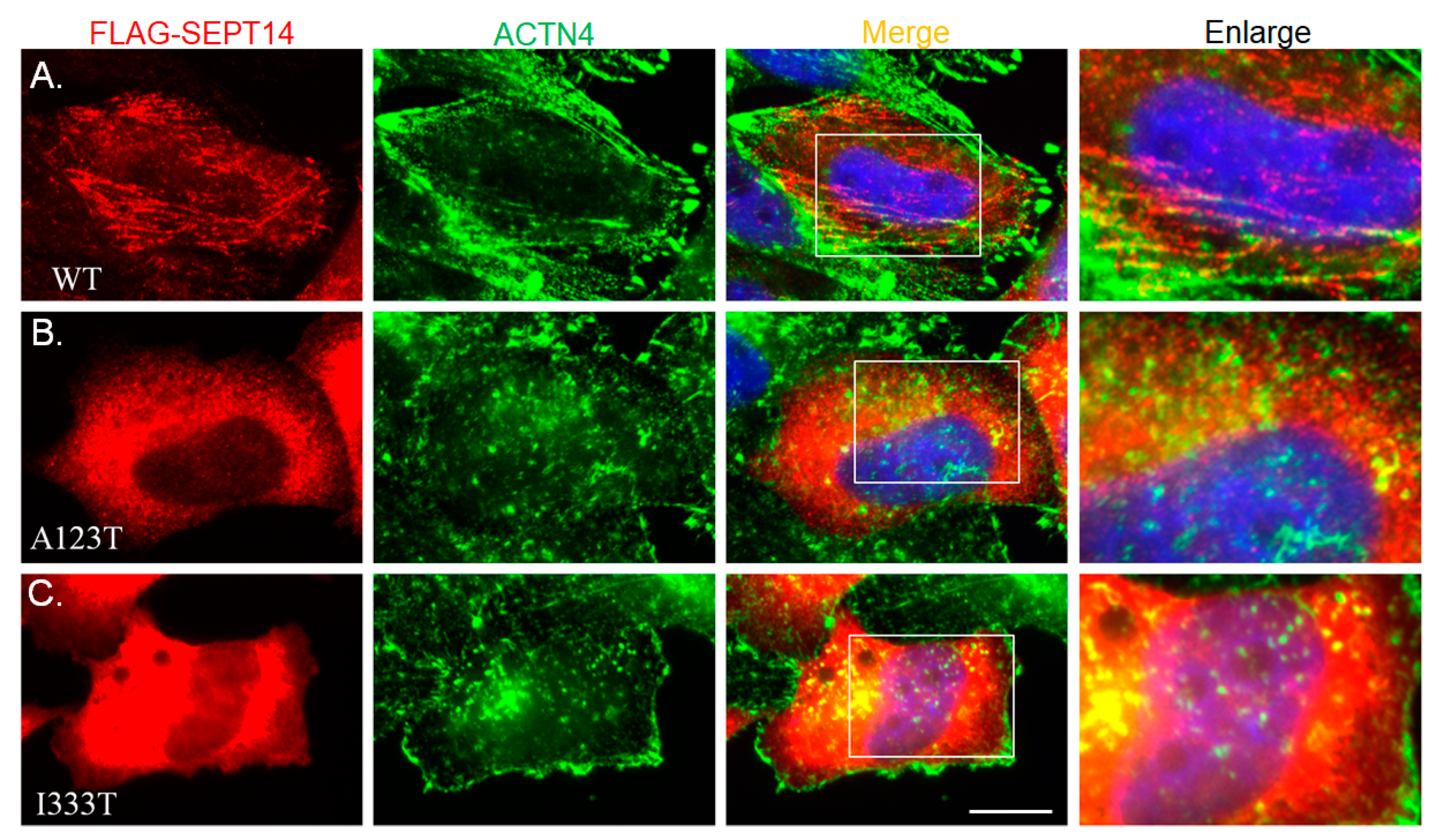
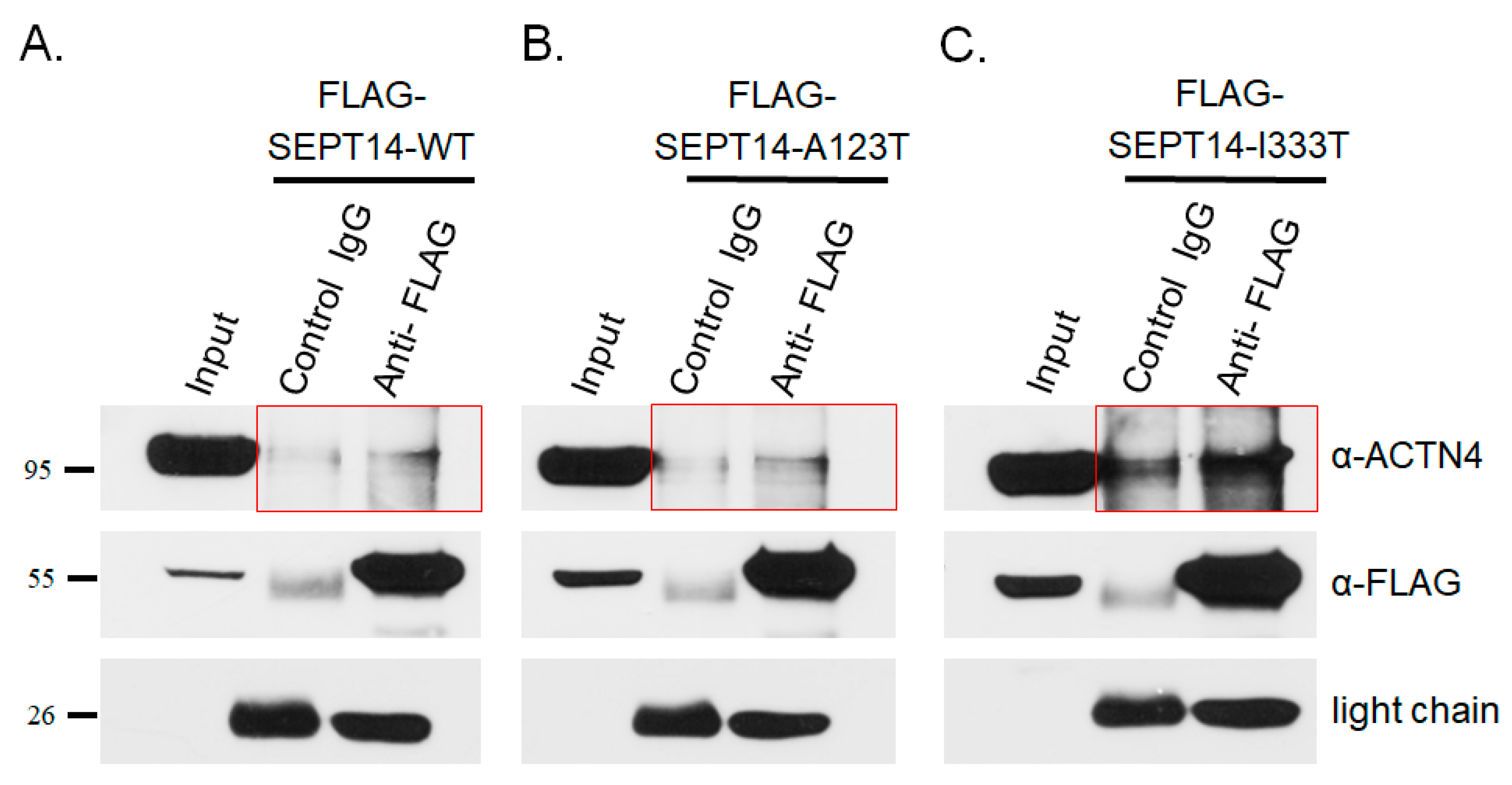
3.4. Mutant SEPT14 Proteins Disturb the ACTN4 Localization Pattern but Do Not Affect the Binding Ability of ACTN4
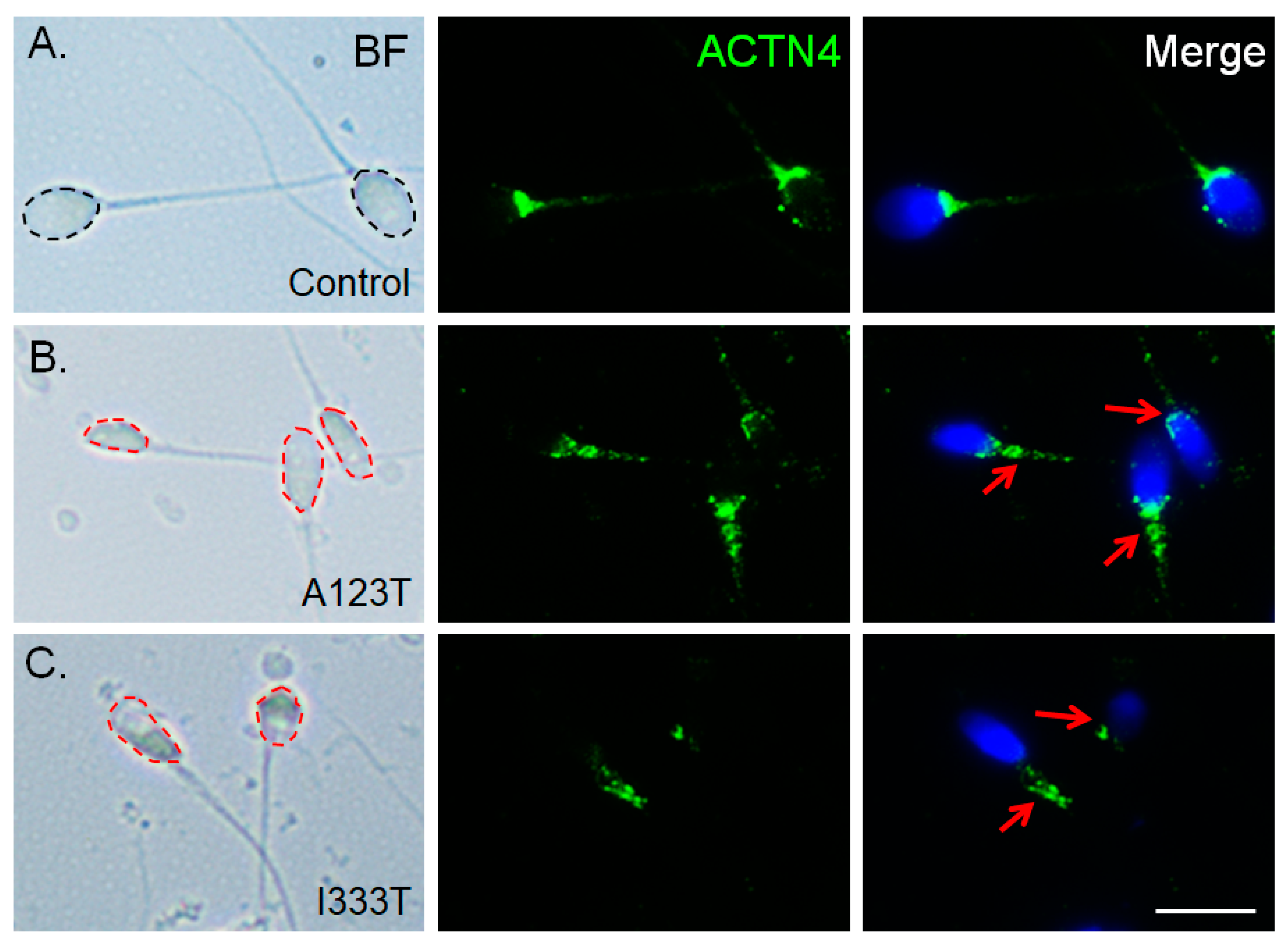
3.5. ACTN4 Patterns Are Disturbed in Human Spermatozoa Bard with the Mutated SEPT14
4. Discussion
4.1. Identification of the SEPT14 Interactor ACTN4 through a Proteomic Assay
4.2. SEPTs Interact with ACTIN in Cells
4.3. Molecular Roles of SEPT14/ACTN4 Complexes during Sperm Head Shaping
4.4. Genetic Changes of SEPT14 in Parkinson’s Disease and Cancer
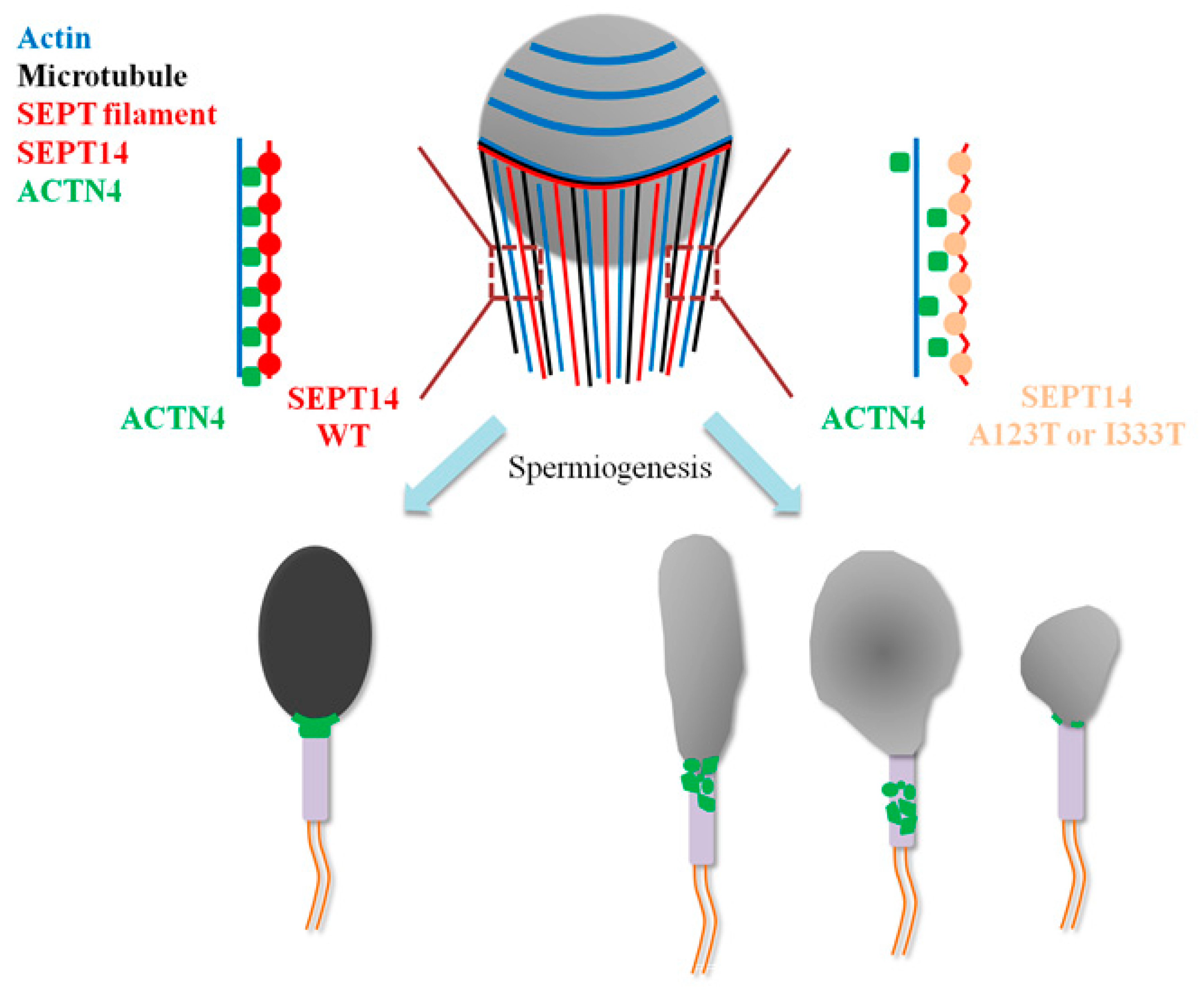
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hirsh, A. Male subfertility. BMJ 2003, 327, 669–672. [Google Scholar] [CrossRef]
- Silber, S.J. Evaluation and treatment of male infertility. Clin. Obstet. Gynecol. 2000, 43, 854–888. [Google Scholar] [CrossRef]
- Madhukar, D.; Rajender, S. Hormonal treatment of male infertility: Promises and pitfalls. J. Androl. 2009, 30, 95–112. [Google Scholar] [CrossRef]
- Aston, K.I. Genetic susceptibility to male infertility: News from genome-wide association studies. Andrology 2014, 2, 315–321. [Google Scholar] [CrossRef]
- Hotaling, J.; Carrell, D.T. Clinical genetic testing for male factor infertility: Current applications and future directions. Andrology 2014, 2, 339–350. [Google Scholar] [CrossRef] [PubMed]
- WHO. WHO Laboratory Manual for the Examination and Processing of Human Semen, 5th ed.; WHO: Geneva, Switzerland, 2010; p. 286. [Google Scholar]
- Palermo, G.D.; Neri, Q.V.; Cozzubbo, T.; Rosenwaks, Z. Perspectives on the assessment of human sperm chromatin integrity. Fertil. Steril. 2014, 102, 1508–1517. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, Q.; Wang, Y.; Li, Y. Whether sperm deoxyribonucleic acid fragmentation has an effect on pregnancy and miscarriage after in vitro fertilization/intracytoplasmic sperm injection: A systematic review and meta-analysis. Fertil. Steril. 2014, 102, 998–1005.e8. [Google Scholar] [CrossRef] [PubMed]
- Osman, A.; Alsomait, H.; Seshadri, S.; El-Toukhy, T.; Khalaf, Y. The effect of sperm DNA fragmentation on live birth rate after IVF or ICSI: A systematic review and meta-analysis. Reprod. Biomed. Online 2015, 30, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Wdowiak, A.; Bakalczuk, S.; Bakalczuk, G. The effect of sperm DNA fragmentation on the dynamics of the embryonic development in intracytoplasmatic sperm injection. Reprod. Biol. 2015, 15, 94–100. [Google Scholar] [CrossRef]
- Agarwal, A.; Cho, C.L.; Esteves, S.C. Should we evaluate and treat sperm DNA fragmentation? Curr. Opin. Obstet. Gynecol. 2016, 28, 164–171. [Google Scholar] [CrossRef] [Green Version]
- Zini, A.; Boman, J.M.; Belzile, E.; Ciampi, A. Sperm DNA damage is associated with an increased risk of pregnancy loss after IVF and ICSI: Systematic review and meta-analysis. Hum. Reprod. 2008, 23, 2663–2668. [Google Scholar] [CrossRef] [Green Version]
- Evenson, D.; Wixon, R. Meta-analysis of sperm DNA fragmentation using the sperm chromatin structure assay. Reprod. Biomed. Online 2006, 12, 466–472. [Google Scholar] [CrossRef]
- De Braekeleer, M.; Nguyen, M.H.; Morel, F.; Perrin, A. Genetic aspects of monomorphic teratozoospermia: A review. J. Assist. Reprod. Genet. 2015, 32, 615–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coutton, C.; Escoffier, J.; Martinez, G.; Arnoult, C.; Ray, P.F. Teratozoospermia: Spotlight on the main genetic actors in the human. Hum. Reprod. Update 2015, 21, 455–485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.H.; Wang, Y.Y.; Chen, H.I.; Kuo, Y.C.; Chiou, Y.W.; Lin, H.H.; Wu, C.M.; Hsu, C.C.; Chiang, H.S.; Kuo, P.L. SEPTIN12 Genetic Variants Confer Susceptibility to Teratozoospermia. PLoS ONE 2012, 7, e34011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravel, C.; Chantot-Bastaraud, S.; El Houate, B.; Berthaut, I.; Verstraete, L.; De Larouziere, V.; Lourenco, D.; Dumaine, A.; Antoine, J.M.; Mandelbaum, J.; et al. Mutations in the protamine 1 gene associated with male infertility. Mol. Hum. Reprod. 2007, 13, 461–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mostowy, S.; Cossart, P. Septins: The fourth component of the cytoskeleton. Nat. Rev. Mol. Cell Biol. 2012, 13, 183–194. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, M.; Noda, M. Roles of septins in the mammalian cytokinesis machinery. Cell Struct. Funct. 2001, 26, 667–670. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, M.; Field, C.M.; Coughlin, M.L.; Straight, A.F.; Mitchison, T.J. Self- and actin-templated assembly of Mammalian septins. Dev. Cell 2002, 3, 791–802. [Google Scholar] [CrossRef] [Green Version]
- Bai, X.; Bowen, J.R.; Knox, T.K.; Zhou, K.; Pendziwiat, M.; Kuhlenbaumer, G.; Sindelar, C.V.; Spiliotis, E.T. Novel septin 9 repeat motifs altered in neuralgic amyotrophy bind and bundle microtubules. J. Cell Biol. 2013, 203, 895–905. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.; Dolat, L.; Angelis, D.; Forgacs, E.; Spiliotis, E.T.; Galkin, V.E. Septin 9 Exhibits Polymorphic Binding to F-Actin and Inhibits Myosin and Cofilin Activity. J. Mol. Biol. 2015, 427, 3273–3284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, P.A.; Jung, K.; Hillan, K.J.; Russell, S.E. Expression profiling the human septin gene family. J. Pathol. 2005, 206, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Peterson, E.A.; Petty, E.M. Conquering the complex world of human septins: Implications for health and disease. Clin. Genet. 2010, 77, 511–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sugino, Y.; Ichioka, K.; Soda, T.; Ihara, M.; Kinoshita, M.; Ogawa, O.; Nishiyama, H. Septins as diagnostic markers for a subset of human asthenozoospermia. J. Urol. 2008, 180, 2706–2709. [Google Scholar] [CrossRef]
- Lhuillier, P.; Rode, B.; Escalier, D.; Lores, P.; Dirami, T.; Bienvenu, T.; Gacon, G.; Dulioust, E.; Toure, A. Absence of annulus in human asthenozoospermia: Case report. Hum. Reprod. 2009, 24, 1296–1303. [Google Scholar] [CrossRef] [Green Version]
- Chao, H.C.; Lin, Y.H.; Kuo, Y.C.; Shen, C.J.; Pan, H.A.; Kuo, P.L. The expression pattern of SEPT7 correlates with sperm morphology. J. Assist. Reprod. Genet. 2010, 27, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Ihara, M.; Kinoshita, A.; Yamada, S.; Tanaka, H.; Tanigaki, A.; Kitano, A.; Goto, M.; Okubo, K.; Nishiyama, H.; Ogawa, O.; et al. Cortical organization by the septin cytoskeleton is essential for structural and mechanical integrity of mammalian spermatozoa. Dev. Cell 2005, 8, 343–352. [Google Scholar] [CrossRef] [Green Version]
- Kissel, H.; Georgescu, M.M.; Larisch, S.; Manova, K.; Hunnicutt, G.R.; Steller, H. The Sept4 septin locus is required for sperm terminal differentiation in mice. Dev. Cell 2005, 8, 353–364. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.H.; Lin, Y.M.; Wang, Y.Y.; Yu, I.S.; Lin, Y.W.; Wang, Y.H.; Wu, C.M.; Pan, H.A.; Chao, S.C.; Yen, P.H.; et al. The expression level of septin12 is critical for spermiogenesis. Am. J. Pathol. 2009, 174, 1857–1868. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.H.; Chou, C.K.; Hung, Y.C.; Yu, I.S.; Pan, H.A.; Lin, S.W.; Kuo, P.L. SEPT12 deficiency causes sperm nucleus damage and developmental arrest of preimplantation embryos. Fertil. Steril. 2011, 95, 363–365. [Google Scholar] [CrossRef]
- Kuo, Y.C.; Lin, Y.H.; Chen, H.I.; Wang, Y.Y.; Chiou, Y.W.; Lin, H.H.; Pan, H.A.; Wu, C.M.; Su, S.M.; Hsu, C.C.; et al. SEPT12 mutations cause male infertility with defective sperm annulus. Hum. Mutat. 2012, 33, 710–719. [Google Scholar] [CrossRef] [PubMed]
- Peterson, E.A.; Kalikin, L.M.; Steels, J.D.; Estey, M.P.; Trimble, W.S.; Petty, E.M. Characterization of a SEPT9 interacting protein, SEPT14, a novel testis-specific septin. Mamm. Genome 2007, 18, 796–807. [Google Scholar] [CrossRef] [PubMed]
- Shinoda, T.; Ito, H.; Sudo, K.; Iwamoto, I.; Morishita, R.; Nagata, K. Septin 14 is involved in cortical neuronal migration via interaction with Septin 4. Mol. Biol. Cell 2010, 21, 1324–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozenkrantz, L.; Gan-Or, Z.; Gana-Weisz, M.; Mirelman, A.; Giladi, N.; Bar-Shira, A.; Orr-Urtreger, A. SEPT14 Is Associated with a Reduced Risk for Parkinson’s Disease and Expressed in Human Brain. J. Mol. Neurosci. 2016, 59, 343–350. [Google Scholar] [CrossRef]
- Hosseinifar, H.; Gilani, S.; Sabbaghian, M. Localization of septin 14 protein in sperm. Iran. J. Reprod. Med. 2015, 2015, 70. [Google Scholar]
- Shafipour, M.; Sabbaghian, M.; Shahhoseini, M.; Sadighi Gilani, M.A. Comparative expression analysis of Septin 14 in testes of infertile men with normal spermatogenesis and spermatogenic failure. Iran. J. Reprod. Med. 2014, 12, 205–208. [Google Scholar]
- Wang, Y.Y.; Lai, T.H.; Chen, M.F.; Lee, H.L.; Kuo, P.L.; Lin, Y.H. SEPT14 Mutations and Teratozoospermia: Genetic Effects on Sperm Head Morphology and DNA Integrity. J. Clin. Med. 2019, 8, 1297. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.H.; Lin, Y.M.; Kuo, Y.C.; Wang, Y.Y.; Kuo, P.L. Identification and characterization of a novel Rab GTPase-activating protein in spermatids. Int. J. Androl. 2011, 34 Pt 2, e358–e367. [Google Scholar] [CrossRef]
- Yeh, Y.C.; Yang, V.C.; Huang, S.C.; Lo, N.W. Stage-dependent expression of extra-embryonic tissue-spermatogenesis-homeobox gene 1 (ESX1) protein, a candidate marker for X chromosome-bearing sperm. Reprod. Fertil. Dev. 2005, 17, 447–455. [Google Scholar] [CrossRef] [Green Version]
- Yeh, C.H.; Kuo, P.L.; Wang, Y.Y.; Wu, Y.Y.; Chen, M.F.; Lin, D.Y.; Lai, T.H.; Chiang, H.S.; Lin, Y.H. SEPT12/SPAG4/LAMINB1 complexes are required for maintaining the integrity of the nuclear envelope in postmeiotic male germ cells. PLoS ONE 2015, 10, e0120722. [Google Scholar] [CrossRef]
- Kuo, P.L.; Chiang, H.S.; Wang, Y.Y.; Kuo, Y.C.; Chen, M.F.; Yu, I.S.; Teng, Y.N.; Lin, S.W.; Lin, Y.H. SEPT12-microtubule complexes are required for sperm head and tail formation. Int. J. Mol. Sci. 2013, 14, 22102–22116. [Google Scholar] [CrossRef] [PubMed]
- Lai, T.H.; Wu, Y.Y.; Wang, Y.Y.; Chen, M.F.; Wang, P.; Chen, T.M.; Wu, Y.N.; Chiang, H.S.; Kuo, P.L.; Lin, Y.H. SEPT12-NDC1 Complexes Are Required for Mammalian Spermiogenesis. Int. J. Mol. Sci. 2016, 17, 1911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Y.H.; Ke, C.C.; Wang, Y.Y.; Chen, M.F.; Chen, T.M.; Ku, W.C.; Chiang, H.S.; Yeh, C.H. RAB10 Interacts with the Male Germ Cell-Specific GTPase-Activating Protein during Mammalian Spermiogenesis. Int. J. Mol. Sci. 2017, 18, 97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, C.C.; Lin, Y.H.; Wang, Y.Y.; Wu, Y.Y.; Chen, M.F.; Ku, W.C.; Chiang, H.S.; Lai, T.H. TBC1D21 Potentially Interacts with and Regulates Rap1 during Murine Spermatogenesis. Int. J. Mol. Sci. 2018, 19, 3292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boersema, P.J.; Raijmakers, R.; Lemeer, S.; Mohammed, S.; Heck, A.J. Multiplex peptide stable isotope dimethyl labeling for quantitative proteomics. Nat. Protoc. 2009, 4, 484–494. [Google Scholar] [CrossRef] [PubMed]
- Rappsilber, J.; Mann, M.; Ishihama, Y. Protocol for micro-purification, enrichment, pre-fractionation and storage of peptides for proteomics using StageTips. Nat. Protoc. 2007, 2, 1896–1906. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.C.; Lin, C.M.; Huang, C.J.; Chen, S.K.; Wu, S.T.; Chiang, H.S.; Ku, W.C. Dual Roles of 17-beta Estradiol in Estrogen Receptor-dependent Growth Inhibition in Renal Cell Carcinoma. Cancer Genom. Proteomics 2016, 13, 219–230. [Google Scholar]
- Roa-Espitia, A.L.; Hernandez-Rendon, E.R.; Baltierrez-Hoyos, R.; Munoz-Gotera, R.J.; Cote-Velez, A.; Jimenez, I.; Gonzalez-Marquez, H.; Hernandez-Gonzalez, E.O. Focal adhesion kinase is required for actin polymerization and remodeling of the cytoskeleton during sperm capacitation. Biol. Open 2016, 5, 1189–1199. [Google Scholar] [CrossRef] [Green Version]
- Sjoblom, B.; Salmazo, A.; Djinovic-Carugo, K. Alpha-actinin structure and regulation. Cell. Mol. Life Sci. 2008, 65, 2688–2701. [Google Scholar] [CrossRef]
- van Son, M.; Tremoen, N.H.; Gaustad, A.H.; Myromslien, F.D.; Vage, D.I.; Stenseth, E.B.; Zeremichael, T.T.; Grindflek, E. RNA sequencing reveals candidate genes and polymorphisms related to sperm DNA integrity in testis tissue from boars. BMC Vet. Res. 2017, 13, 362. [Google Scholar] [CrossRef] [Green Version]
- Dunleavy, J.E.M.; O’Bryan, M.; Stanton, P.G.; O’Donnell, L. The Cytoskeleton in Spermatogenesis. Reproduction 2018, 157, R53–R72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pleuger, C.; Lehti, M.S.; Dunleavy, J.E.; Fietz, D.; O’Bryan, M.K. Haploid male germ cells-the Grand Central Station of protein transport. Hum. Reprod. Update 2020, 26, 474–500. [Google Scholar] [CrossRef] [PubMed]
- Tilney, L.G. Actin filaments in the acrosomal reaction of Limulus sperm. Motion generated by alterations in the packing of the filaments. J. Cell Biol. 1975, 64, 289–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goode, N.P.; Shires, M.; Khan, T.N.; Mooney, A.F. Expression of alpha-actinin-4 in acquired human nephrotic syndrome: A quantitative immunoelectron microscopy study. Nephrol. Dial. Transplant. 2004, 19, 844–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyrieleis, H.A.; Lowik, M.M.; Pronk, I.; Cruysberg, H.R.; Kremer, J.A.; Oyen, W.J.; van den Heuvel, B.L.; Wetzels, J.F.; Levtchenko, E.N. Long-term outcome of biopsy-proven, frequently relapsing minimal-change nephrotic syndrome in children. Clin. J. Am. Soc. Nephrol. CJASN 2009, 4, 1593–1600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safarikova, M.; Reiterova, J.; Safrankova, H.; Stekrova, J.; Zidkova, A.; Obeidova, L.; Kohoutova, M.; Tesar, V. Mutational analysis of ACTN4, encoding alpha-actinin 4, in patients with focal segmental glomerulosclerosis using HRM method. Folia Biol. 2013, 59, 110–115. [Google Scholar]
- Schmidt, K.; Nichols, B.J. Functional interdependence between septin and actin cytoskeleton. BMC Cell Biol. 2004, 5, 43. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, M.; Kumar, S.; Mizoguchi, A.; Ide, C.; Kinoshita, A.; Haraguchi, T.; Hiraoka, Y.; Noda, M. Nedd5, a mammalian septin, is a novel cytoskeletal component interacting with actin-based structures. Genes Dev. 1997, 11, 1535–1547. [Google Scholar] [CrossRef] [Green Version]
- Joo, E.; Surka, M.C.; Trimble, W.S. Mammalian SEPT2 is required for scaffolding nonmuscle myosin II and its kinases. Dev. Cell 2007, 13, 677–690. [Google Scholar] [CrossRef] [Green Version]
- Kremer, B.E.; Adang, L.A.; Macara, I.G. Septins regulate actin organization and cell-cycle arrest through nuclear accumulation of NCK mediated by SOCS7. Cell 2007, 130, 837–850. [Google Scholar] [CrossRef] [Green Version]
- de Boer, P.; de Vries, M.; Ramos, L. A mutation study of sperm head shape and motility in the mouse: Lessons for the clinic. Andrology 2015, 3, 174–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelis, D.; Spiliotis, E.T. Septin Mutations in Human Cancers. Front. Cell dev. Biol. 2016, 4, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frattini, V.; Trifonov, V.; Chan, J.M.; Castano, A.; Lia, M.; Abate, F.; Keir, S.T.; Ji, A.X.; Zoppoli, P.; Niola, F.; et al. The integrated landscape of driver genomic alterations in glioblastoma. Nat. Genet. 2013, 45, 1141–1149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, H.B.; Chen, X.; Yang, X.; Ye, Y.; Bekaii-Saab, T.; Zheng, Y.; Zhang, Y. A Rare EGFR-SEPT14 Fusion in a Patient with Colorectal Adenocarcinoma Responding to Erlotinib. Oncologist 2020, 25, 203–207. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Symbol | Gene | Functions |
|---|---|---|
| SEPT-related proteins | ||
| SEPT2 | Septin 2 | Cell division, cilium assembly |
| SEPT7 | Septin 7 | Cytokinesis, cilium morphogenesis |
| SEPT9 | Septin 9 | Cell division |
| SEPT11 | Septin 11 | Cytokinesis, vesicle trafficking |
| Microtubule-related proteins | ||
| TUBB2A | Tubulin beta 2A | Mitosis, intracellular transport |
| TUBB2B | Tubulin beta 2B | Mitosis, intracellular transport |
| TUBB4B | Tubulin, beta 4B | Mitosis, intracellular transport |
| STMN1 | Stathmin | Microtubule depolymerization, axonogenesis |
| KIF5B | Kinesin family member 5B | Cytoskeleton-dependent intracellular transport |
| DYNC1LI1 | Dynein, cytoplasmic 1, light intermediate chain 1 | Cell division, microtubule-based movement |
| Actin-related proteins | ||
| ACTA1 | Actin, alpha 1 | Cell motility, structure, and integrity |
| ACTN2 | Actinin, alpha 2 | Actin filament binding, muscle filament sliding, cell adhesion |
| ACTN3 | Actinin, alpha 3 | Crosslinking actin containing thin filaments, focal adhesion assembly |
| ACTN4 | Actinin, alpha 4 | Binding actin to the membrane, vesicle transport along actin filament |
| CTTN | Cortactin | Regulation of adherens-type junctions, organization of the actin structure |
| CFL2 | Cofilin 2 | Actin filament depolymerization, regulation of dendritic spine morphogenesis |
| ITGB1 | Integrin beta-1 | Cell adhesion, embryogenesis |
| MYO1C | Myosin IC | Membrane binding, lipid raft trafficking |
| MYL6B | Myosin light chain 6B | Muscle myosin |
| MYH14 | Myosin, heavy chain 14 | Cytokinesis, cell motility, cell polarity, axon guidance |
| TPM3 | Tropomyosin 3 | Provide stability to actin filaments and regulate access of other actin-binding proteins |
| LAMA2 | Laminin subunit alpha 2 | Major component of the basement membrane |
| ZYX | Zyxin | Modulate the cytoskeletal organization of actin bundles |
| Sperm-related proteins | ||
| ODF2 | Outer dense fiber of sperm tails 2 | Spermatid development, maintaining the elastic structure and recoil of the sperm tail |
| FSIP2 | Fibrous sheath interacting protein 2 | AKAP4-interacting protein |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Y.-H.; Huang, C.-Y.; Ke, C.-C.; Wang, Y.-Y.; Lai, T.-H.; Liu, H.-C.; Ku, W.-C.; Chan, C.-C.; Lin, Y.-H. ACTN4 Mediates SEPT14 Mutation-Induced Sperm Head Defects. Biomedicines 2020, 8, 518. https://doi.org/10.3390/biomedicines8110518
Lin Y-H, Huang C-Y, Ke C-C, Wang Y-Y, Lai T-H, Liu H-C, Ku W-C, Chan C-C, Lin Y-H. ACTN4 Mediates SEPT14 Mutation-Induced Sperm Head Defects. Biomedicines. 2020; 8(11):518. https://doi.org/10.3390/biomedicines8110518
Chicago/Turabian StyleLin, Yu-Hua, Chia-Yen Huang, Chih-Chun Ke, Ya-Yun Wang, Tsung-Hsuan Lai, Hsuan-Che Liu, Wei-Chi Ku, Chying-Chyuan Chan, and Ying-Hung Lin. 2020. "ACTN4 Mediates SEPT14 Mutation-Induced Sperm Head Defects" Biomedicines 8, no. 11: 518. https://doi.org/10.3390/biomedicines8110518
APA StyleLin, Y. -H., Huang, C. -Y., Ke, C. -C., Wang, Y. -Y., Lai, T. -H., Liu, H. -C., Ku, W. -C., Chan, C. -C., & Lin, Y. -H. (2020). ACTN4 Mediates SEPT14 Mutation-Induced Sperm Head Defects. Biomedicines, 8(11), 518. https://doi.org/10.3390/biomedicines8110518