
Osteopontin Expression in Thyroid Cancer: Deciphering EMT-Related Molecular Mechanisms

 ,
,  ,
,
Abstract
:1. Introduction
2. Thyroid Cancer
3. EMT, MET and Epithelial–Mesenchymal Plasticity
4. EMT in Thyroid Cancer
4.1. EMT-Related Transcription Factors Modulating the Expression of EMT Markers in Thyroid Cancer
4.2. MiRNAs as Regulators of EMT in Thyroid Cancer
5. Osteopontin Expression and Crosstalks with EMT in Thyroid Cancer
6. Final Statements and Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Enternational Agency for Research on Cancer/World Health Organization. Cancer Today, Data Visualization Tools for Ex-Ploring the Global Cancer Burden in 2020. Available online: https://gco.iarc.fr/today/home (accessed on 10 January 2021).
- Raue, F.; Frank-Raue, K. Epidemiology and Clinical Presentation of Medullary Thyroid Carcinoma. Stem Cells Aging 2015, 204, 61–90. [Google Scholar] [CrossRef]
- Nieto, M.A.; Huang, R.Y.-J.; Jackson, R.A.; Thiery, J.P. EMT: 2016. Cell 2016, 166, 21–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shibue, T.; Weinberg, R.A. EMT, CSCs, and Drug Resistance: The Mechanistic Link and Clinical Implications. Nat. Rev. Clin. Oncol. 2017, 14, 611–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kothari, A.N.; Arffa, M.L.; Chang, V.; Blackwell, R.H.; Syn, W.-K.; Zhang, J.; Mi, Z.; Kuo, P.C. Osteopontin—A Master Regulator of Epithelial-Mesenchymal Transition. J. Clin. Med. 2016, 5, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gimba, E.; Tilli, T. Human osteopontin splicing isoforms: Known roles, potential clinical applications and activated signaling pathways. Cancer Lett. 2013, 331, 11–17. [Google Scholar] [CrossRef]
- Briones-Orta, M.A.; Avendaño-Vázquez, S.E.; Aparicio-Bautista, D.I.; Coombes, J.D.; Weber, G.F.; Syn, W.-K. Osteopontin splice variants and polymorphisms in cancer progression and prognosis. Biochim. Biophys. Acta (BBA) Bioenerg. 2017, 1868, 93–108. [Google Scholar] [CrossRef]
- Silva, G.R.; Mattos, D.S.; Bastos, A.C.F.; Viana, B.P.P.B.; Brum, M.C.M.; Ferreira, L.B.; Gimba, E.R.P. Osteopontin-4 and Osteopontin-5 splice variants are expressed in several tumor cell lines. Mol. Biol. Rep. 2020, 47, 8339–8345. [Google Scholar] [CrossRef]
- Ferreira, L.B.; Tavares, C.; Pestana, A.; Pereira, C.L.; Eloy, C.; Pinto, M.; Castro, P.; Batista, R.; Rios, E.; Sobrinho-Simões, M.; et al. Osteopontin-a splice variant is overexpressed in papillary thyroid carcinoma and modulates invasive behavior. Oncotarget 2016, 7, 52003–52016. [Google Scholar] [CrossRef] [Green Version]
- Shakib, H.; Rajabi, S.; Dehghan, M.H.; Mashayekhi, F.J.; Safari-Alighiarloo, N.; Hedayati, M. Epithelial-to-mesenchymal transition in thyroid cancer: A comprehensive review. Endocrine 2019, 66, 435–455. [Google Scholar] [CrossRef] [PubMed]
- Luzón-Toro, B.; Fernández, R.M.; Villalba-Benito, L.; Torroglosa, A.; Antiñolo, G.; Borrego, S. Influencers on Thyroid Cancer Onset: Molecular Genetic Basis. Genes 2019, 10, 913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kitahara, C.M.; Sosa, J.A. The changing incidence of thyroid cancer. Nat. Rev. Endocrinol. 2016, 12, 646–653. [Google Scholar] [CrossRef]
- Elisei, R.; Ugolini, C.; Viola, D.; Lupi, C.; Biagini, A.; Giannini, R.; Romei, C.; Miccoli, P.; Pinchera, A.; Basolo, F. BRAFV600E Mutation and Outcome of Patients with Papillary Thyroid Carcinoma: A 15-Year Median Follow-Up Study. J. Clin. Endocrinol. Metab. 2008, 93, 3943–3949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czarniecka, A.; Oczko-Wojciechowska, M.; Barczyński, M. BRAF V600E mutation in prognostication of papillary thyroid cancer (PTC) recurrence. Gland. Surg. 2016, 5, 495–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damiani, L.; Lupo, S.; Rossi, R.; Bruni, S.; Bartolomei, M.; Panareo, S.; Franceschetti, P.; Carcoforo, P.; Lanza, G.; Pelucchi, S.; et al. Evaluation of the Role of BRAFV600E Somatic Mutation on Papillary Thyroid Cancer Disease Persistence: A Prospective Study. Eur. Thyroid. J. 2018, 7, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Chakravarty, D.; Santos, E.; Ryder, M.; Knauf, J.; Liao, X.-H.; West, B.; Bollag, G.; Kolesnick, R.; Thin, T.H.; Rosen, N.; et al. Small-molecule MAPK inhibitors restore radioiodine incorporation in mouse thyroid cancers with conditional BRAF activation. J. Clin. Investig. 2011, 121, 4700–4711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sobrinho-Simões, M.; Eloy, C.; Magalhães, J.; Lobo, C.; Amaro, T. Follicular Thyroid Carcinoma. Mod. Pathol. 2011, 24 (Suppl. 2), S10–S18. [Google Scholar] [CrossRef]
- Nikiforova, M.N.; Lynch, R.A.; Biddinger, P.W.; Alexander, E.K.; Dorn, G.W.; Tallini, G.; Kroll, T.G.; Nikiforov, Y.E. RAS Point Mutations and PAX8-PPAR Gamma Rearrangement in Thyroid Tumors: Evidence for Distinct Molecular Pathways in Thyroid Follicular Carcinoma. J. Clin. Endocrinol. Metab. 2003, 88, 2318–2326. [Google Scholar] [CrossRef] [Green Version]
- Viola, D.; Valerio, L.; Molinaro, E.; Agate, L.; Bottici, V.; Biagini, A.; Lorusso, L.; Cappagli, V.; Pieruzzi, L.; Giani, C.; et al. Treatment of advanced thyroid cancer with targeted therapies: Ten years of experience. Endocr.-Relat. Cancer 2016, 23, R185–R205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khairy, G. Anaplastic Transformation of Differentiated Thyroid Carcinoma. Int. J. Health Sci. 2009, 3, 93–96. [Google Scholar]
- Yu, M.G.; Rivera, J.; Jimeno, C. Poorly Differentiated Thyroid Carcinoma: 10-Year Experience in a Southeast Asian Population. Endocrinol. Metab. 2017, 32, 288–295. [Google Scholar] [CrossRef]
- Landa, I.; Ganly, I.; Chan, T.A.; Mitsutake, N.; Matsuse, M.; Ibrahimpasic, T.; Ghossein, R.A.; Fagin, J.A. Frequent Somatic TERT Promoter Mutations in Thyroid Cancer: Higher Prevalence in Advanced Forms of the Disease. J. Clin. Endocrinol. Metab. 2013, 98, E1562–E1566. [Google Scholar] [CrossRef] [Green Version]
- Modigliani, E.; Cohen, R.; Campos, J.-M.; Conte-Devolx, B.; Maes, B.; Boneu, A.; Schlumberger, M.; Bigorgne, J.-C.; Dumontier, P.; Leclerc, L.; et al. Prognostic factors for survival and for biochemical cure in medullary thyroid carcinoma: Results in 899 patients. Clin. Endocrinol. 1998, 48, 265–273. [Google Scholar] [CrossRef]
- Keiser, H.R. Sipple’s Syndrome: Medullary Thyroid Carcinoma, Pheochromocytoma, and Parathyroid Disease: Studies in a Large Family. Ann. Intern. Med. 1973, 78, 561–579. [Google Scholar] [CrossRef]
- Cabanillas, M.E.; McFadden, D.G.; Durante, C. Thyroid Cancer. Lancet 2016, 388, 2783–2795. [Google Scholar] [CrossRef]
- Dongre, A.; Weinberg, R.A. New insights into the mechanisms of epithelial–mesenchymal transition and implications for cancer. Nat. Rev. Mol. Cell Biol. 2019, 20, 69–84. [Google Scholar] [CrossRef]
- Lu, W.; Kang, Y. Epithelial-Mesenchymal Plasticity in Cancer Progression and Metastasis. Dev. Cell 2019, 49, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.D.; Gao, D.; Redfern, A.; Thompson, E.W. Controversies around epithelial–mesenchymal plasticity in cancer metastasis. Nat. Rev. Cancer 2019, 19, 716–732. [Google Scholar] [CrossRef] [Green Version]
- Yadav, A.K.; Desai, N.S. Cancer Stem Cells: Acquisition, Characteristics, Therapeutic Implications, Targeting Strategies and Future Prospects. Stem Cell Rev. Rep. 2019, 15, 331–355. [Google Scholar] [CrossRef] [PubMed]
- Liao, T.-T.; Yang, M.-H. Revisiting epithelial-mesenchymal transition in cancer metastasis: The connection between epithelial plasticity and stemness. Mol. Oncol. 2017, 11, 792–804. [Google Scholar] [CrossRef]
- Palona, I.; Namba, H.; Mitsutake, N.; Starenki, D.; Podtcheko, A.; Sedliarou, I.; Ohtsuru, A.; Saenko, V.; Nagayama, Y.; Umezawa, K.; et al. BRAF V600E Promotes Invasiveness of Thyroid Cancer Cells through Nuclear Factor ΚB Activation. Endocrinology 2006, 147, 5699–5707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasko, V.; Espinosa, A.V.; Scouten, W.; He, H.; Auer, H.; Liyanarachchi, S.; Larin, O.; Savchenko, V.; Francis, G.L.; de la Chapelle, A.; et al. Gene expression and functional evidence of epithelial-to-mesenchymal transition in papillary thyroid carcinoma invasion. Proc. Natl. Acad. Sci. USA 2007, 104, 2803–2808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braun, J.; Hoangvu, C.; Dralle, H.; Huttelmaier, S. Downregulation of microRNAs directs the EMT and invasive potential of anaplastic thyroid carcinomas. Oncogene 2010, 29, 4237–4244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.; Kakudo, K.; Bai, Y.; Li, Y.; Ozaki, T.; Miyauchi, A.; Taniguchi, E.; Mori, I. Loss of cellular polarity/cohesiveness in the invasive front of papillary thyroid carcinoma, a novel predictor for lymph node metastasis; possible morphological indicator of epithelial mesenchymal transition. J. Clin. Pathol. 2011, 64, 325–329. [Google Scholar] [CrossRef] [PubMed]
- Montemayor-Garcia, C.; Hardin, H.; Guo, Z.; Larrain, C.; Buehler, D.; Asioli, S.; Chen, H.; Lloyd, R.V. The role of epithelial mesenchymal transition markers in thyroid carcinoma progression. Endocr. Pathol. 2013, 24, 206–212. [Google Scholar] [CrossRef] [Green Version]
- Yasui, K.; Shimamura, M.; Mitsutake, N.; Nagayama, Y. SNAIL Induces Epithelial-to-Mesenchymal Transition and Cancer Stem Cell–Like Properties in Aldehyde Dehydroghenase–Negative Thyroid Cancer Cells. Thyroid 2013, 23, 989–996. [Google Scholar] [CrossRef] [Green Version]
- Hardin, H.; Guo, Z.; Shan, W.; Montemayor-Garcia, C.; Asioli, S.; Yu, X.-M.; Harrison, A.D.; Chen, H.; Lloyd, R.V. The Roles of the Epithelial-Mesenchymal Transition Marker PRRX1 and miR-146b-5p in Papillary Thyroid Carcinoma Progression. Am. J. Pathol. 2014, 184, 2342–2354. [Google Scholar] [CrossRef] [Green Version]
- Ma, R.; Minsky, N.; Morshed, S.A.; Davies, T.F. Stemness in Human Thyroid Cancers and Derived Cell Lines: The Role of Asymmetrically Dividing Cancer Stem Cells Resistant to Chemotherapy. J. Clin. Endocrinol. Metab. 2014, 99, E400–E409. [Google Scholar] [CrossRef]
- Lv, N.; Shan, Z.; Gao, Y.; Guan, H.; Fan, C.; Wang, H.; Teng, W. Twist1 Regulates the Epithelial–Mesenchymal Transition via the NF-ΚB Pathway in Papillary Thyroid Carcinoma. Endocrine 2016, 51, 469–477. [Google Scholar] [CrossRef]
- Byeon, H.K.; Na, H.J.; Yang, Y.J.; Ko, S.; Yoon, S.O.; Ku, M.; Yang, J.; Kim, J.W.; Ban, M.J.; Kim, J.-H.; et al. Acquired resistance to BRAF inhibition induces epithelial-to-mesenchymal transition in BRAF (V600E) mutant thyroid cancer by c-Met-mediated AKT activation. Oncotarget 2016, 8, 596–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werner, T.A.; Forster, C.M.; Dizdar, L.; Verde, P.E.; Raba, K.; Schott, M.; Knoefel, W.T.; Krieg, A. CXCR4/CXCR7/CXCL12 Axis Promotes an Invasive Phenotype in Medullary Thyroid Carcinoma. Br. J. Cancer 2017, 117, 1837–1845. [Google Scholar] [CrossRef]
- Lee, S.; Bae, J.S.; Jung, C.K.; Chung, W.Y. Extensive lymphatic spread of papillary thyroid microcarcinoma is associated with an increase in expression of genes involved in epithelial-mesenchymal transition and cancer stem cell-like properties. Cancer Med. 2019, 8, 6528–6537. [Google Scholar] [CrossRef]
- Wu, J.; Zhang, Y.; Cheng, R.; Gong, W.; Ding, T.; Zhai, Q.; Wang, Y.; Meng, B.; Sun, B. Expression of epithelial-mesenchymal transition regulators TWIST, SLUG and SNAIL in follicular thyroid tumours may relate to widely invasive, poorly differentiated and distant metastasis. Histopathology 2018, 74, 780–791. [Google Scholar] [CrossRef]
- Zhao, Y.-L.; Yuan, B.-Q.; Shen, G.-S. Mechanism of RET Gene Mediated EGFR Signaling Pathway on Epithelial-Mesenchymal Transition, Proliferation and Apoptosis of Papillary Thyroid Carcinoma Cells. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 8036–8047. [Google Scholar] [CrossRef]
- Niu, D.-F.; Kondo, T.; Nakazawa, T.; Oishi, N.; Kawasaki, T.; Mochizuki, K.; Yamane, T.; Katoh, R. Transcription factor Runx2 is a regulator of epithelial–mesenchymal transition and invasion in thyroid carcinomas. Lab. Investig. 2012, 92, 1181–1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Zhou, G.; Nakamura, M.; Ozaki, T.; Mori, I.; Taniguchi, E.; Miyauchi, A.; Ito, Y.; Kakudo, K. Survival impact of psammoma body, stromal calcification, and bone formation in papillary thyroid carcinoma. Mod. Pathol. 2009, 22, 887–894. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, M.; Onoda, N.; Noda, S.; Kashiwagi, S.; Aomatsu, N.; Kurata, K.; Kawajiri, H.; Takashima, T.; Ishikawa, T.; Hirakawa, K. E-cadherin expression and cell proliferation in the primary tumor and metastatic lymph nodes of papillary thyroid microcarcinoma. Mol. Clin. Oncol. 2014, 2, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Buehler, D.; Hardin, H.; Shan, W.; Montemayor-Garcia, C.; Rush, P.S.; Asioli, S.; Chen, H.; Lloyd, R.V. Expression of epithelial-mesenchymal transition regulators SNAI2 and TWIST1 in thyroid carcinomas. Mod. Pathol. 2012, 26, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhang, Y.; Liu, G.; Wu, S.; Jiang, F.; Xie, J. Zinc finger E-box-binding homeobox 1: Its clinical significance and functional role in human thyroid cancer. OncoTargets Ther. 2016, 9, 1303–1310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cerutti, J.; Ebina, K.N.; Matsuo, S.E.; Martins, L.; Maciel, R.M.B.; Kimura, E.T. Expression of Smad4 and Smad7 in human thyroid follicular carcinoma cell lines. J. Endocrinol. Investig. 2003, 26, 516–521. [Google Scholar] [CrossRef]
- He, H.; Jazdzewski, K.; Li, W.; Liyanarachchi, S.; Nagy, R.; Volinia, S.; Calin, G.; Liu, C.-G.; Franssila, K.; Suster, S.; et al. The role of microRNA genes in papillary thyroid carcinoma. Proc. Natl. Acad. Sci. USA 2005, 102, 19075–19080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, W.; Lan, X.; Wang, Z.; Dong, W.; He, L.; Zhang, T.; Zhang, H. MicroRNA-144 inhibits proliferation by targeting WW domain-containing transcription regulator protein 1 in papillary thyroid cancer. Oncol. Lett. 2017, 15, 1007–1013. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Moya, J.; Wert-Lamas, L.; Santisteban, P. MicroRNA-146b promotes PI3K/AKT pathway hyperactivation and thyroid cancer progression by targeting PTEN. Oncogene 2018, 37, 3369–3383. [Google Scholar] [CrossRef]
- Wei, Z.-L.; Gao, A.-B.; Wang, Q.; Lou, X.-E.; Zhao, J.; Lu, Q.-J. MicroRNA-221 promotes papillary thyroid carcinoma cells migration and invasion via targeting RECK and regulating epithelial–mesenchymal transition. OncoTargets Ther. 2019, 12, 2323–2333. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Su, C.; Chen, Y.; Li, G. MiR-144-3p promotes the tumor growth and metastasis of papillary thyroid carcinoma by targeting paired box gene 8. Cancer Cell Int. 2018, 18, 54. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, B.-G.; Zhou, C.-X. MicroRNA-31 inhibits papillary thyroid carcinoma cell biological progression by directly targeting SOX11 and regulating epithelial-to-mesenchymal transition, ERK and Akt signaling pathways. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5863–5873. [Google Scholar] [CrossRef]
- Bi, C.; Zhang, Y.; Li, B.; Guo, M.; Fu, Y. MicroRNA-520a-3p suppresses epithelial–mesenchymal transition, invasion, and migration of papillary thyroid carcinoma cells via the JAK1 -mediated JAK/STAT signaling pathway. J. Cell. Physiol. 2019, 234, 4054–4067. [Google Scholar] [CrossRef]
- Hébrant, A.; Dom, G.; Dewaele, M.; Andry, G.; Trésallet, C.; Leteurtre, E.; Dumont, J.E.; Maenhaut, C. mRNA Expression in Papillary and Anaplastic Thyroid Carcinoma: Molecular Anatomy of a Killing Switch. PLoS ONE 2012, 7, e37807. [Google Scholar] [CrossRef]
- Anborgh, P.H.; Mutrie, J.C.; Tuck, A.B.; Chambers, A.F. Role of the metastasis-promoting protein osteopontin in the tumour microenvironment. J. Cell. Mol. Med. 2010, 14, 2037–2044. [Google Scholar] [CrossRef] [Green Version]
- Zohar, R.; Lee, W.; Arora, P.; Cheifetz, S.; McCulloch, C.; Sodek, J. Single Cell Analysis of Intracellular Osteopontin in Osteogenic Cultures of Fetal Rat Calvarial Cells. J. Cell Physiol. 1997, 170, 88–100. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, Q.; Alam, A.; Cui, J.; Suen, K.C.; Soo, A.P.; Eguchi, S.; Gu, J.; Ma, D. The role of osteopontin in the progression of solid organ tumour. Cell Death Dis. 2018, 9, 356. [Google Scholar] [CrossRef] [PubMed]
- Icer, M.A.; Gezmen-Karadag, M. The multiple functions and mechanisms of osteopontin. Clin. Biochem. 2018, 59, 17–24. [Google Scholar] [CrossRef] [PubMed]
- A Takafuji, V.; Forgues, M.; Unsworth, E.; Goldsmith, P.; Wang, X.W. An osteopontin fragment is essential for tumor cell invasion in hepatocellular carcinoma. Oncogene 2007, 26, 6361–6371. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Chang, S.; Lu, Y.; Wang, J.; Si, Y.; Zhang, L.; Cheng, S.; Jiang, W.G. Enhanced osteopontin splicing regulated by RUNX2 is HDAC-dependent and induces invasive phenotypes in NSCLC cells. Cancer Cell Int. 2019, 19, 306. [Google Scholar] [CrossRef]
- Jia, R.; Liang, Y.; Chen, R.; Liu, G.; Wang, H.; Tang, M.; Zhou, X.; Wang, H.; Yang, Y.; Wei, H.; et al. Osteopontin facilitates tumor metastasis by regulating epithelial–mesenchymal plasticity. Cell Death Dis. 2016, 7, e2564. [Google Scholar] [CrossRef]
- Brown, L.F.; Papadopoulos-Sergiou, A.; Berse, B.; Manseau, E.J.; Tognazzi, K.; Perruzzi, C.A.; Dvorak, H.F.; Senger, D.R. Osteopontin expression and distribution in human carcinomas. Am. J. Pathol. 1994, 145, 610–623. [Google Scholar]
- Fisher, L.W.; Jain, A.; Tayback, M.; Fedarko, N.S. Small Integrin Binding LigandN-Linked Glycoprotein Gene Family Expression in Different Cancers. Clin. Cancer Res. 2004, 10, 8501–8511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fluge, Φ.; Bruland, O.; Akslen, L.A.; Lillehaug, J.R.; Varhaug, J.E. Gene Expression in Poorly Differentiated Papillary Thyroid Carcinomas. Thyroid 2006, 16, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Castellone, M.; Celetti, A.; Guarino, V.; Cirafici, A.M.; Basolo, F.; Giannini, R.; Medico, E.; Kruhoffer, M.; Orntoft, T.F.; Curcio, F.; et al. Autocrine stimulation by osteopontin plays a pivotal role in the expression of the mitogenic and invasive phenotype of RET/PTC-transformed thyroid cells. Oncogene 2004, 23, 2188–2196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borrello, M.G.; Alberti, L.; Fischer, A.; Degl’Innocenti, D.; Ferrario, C.; Gariboldi, M.; Marchesi, F.; Allavena, P.; Greco, A.; Collini, P.; et al. Induction of a proinflammatory program in normal human thyrocytes by the RET/PTC1 oncogene. Proc. Natl. Acad. Sci. USA 2005, 102, 14825–14830. [Google Scholar] [CrossRef] [Green Version]
- Guarino, V.; Faviana, P.; Salvatore, G.; Castellone, M.; Cirafici, A.M.; De Falco, V.; Celetti, A.; Giannini, R.; Basolo, F.; Melillo, R.M.; et al. Osteopontin Is Overexpressed in Human Papillary Thyroid Carcinomas and Enhances Thyroid Carcinoma Cell Invasiveness. J. Clin. Endocrinol. Metab. 2005, 90, 5270–5278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oler, G.; Camacho, C.P.; Hojaij, F.C., Jr.; Michaluart, P.M.; Riggins, G.J.; Cerutti, J. Gene Expression Profiling of Papillary Thyroid Carcinoma Identifies Transcripts Correlated with BRAF Mutational Status and Lymph Node Metastasis. Clin. Cancer Res. 2008, 14, 4735–4742. [Google Scholar] [CrossRef] [Green Version]
- Briese, J.; Cheng, S.; Ezzat, S.; Liu, W.; Winer, D.; Wagener, C.; Bamberger, A.-M.; Asa, S. Osteopontin (OPN) expression in thyroid carcinoma. Anticancer Res. 2010, 30, 1681–1688. [Google Scholar]
- Wang, X.; Chao, L.; Zhen, J.; Chen, L.; Ma, G.; Li, X. Phosphorylated c-Jun NH2-terminal kinase is overexpressed in human papillary thyroid carcinomas and associates with lymph node metastasis. Cancer Lett. 2010, 293, 175–180. [Google Scholar] [CrossRef]
- Sun, Y.; Fang, S.; Dong, H.; Zhao, C.; Yang, Z.; Li, P.; Wang, J. Correlation between osteopontin messenger RNA expression and microcalcification shown on sonography in papillary thyroid carcinoma. J. Ultrasound Med. 2011, 30, 765–771. [Google Scholar] [CrossRef]
- Kang, K.H. Osteopontin Expression in Papillary Thyroid Carcinoma and Its Relationship with the BRAF Mutation and Tumor Characteristics. J. Korean Surg. Soc. 2013, 84, 9–17. [Google Scholar] [CrossRef]
- Mu, G.; Wang, H.; Cai, Z.; Ji, H. OPN -443C>T Genetic Polymorphism and Tumor OPN Expression are Associated with the Risk and Clinical Features of Papillary Thyroid Cancer in a Chinese Cohort. Cell. Physiol. Biochem. 2013, 32, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Sepe, R.; Formisano, U.; Federico, A.; Forzati, F.; Bastos, A.U.; Angelo, D.D.; Cacciola, N.A.; Fusco, A.; Pallante, P. CBX7 and HMGA1b proteins act in opposite way on the regulation of the SPP1 gene expression. Oncotarget 2015, 6, 2680–2692. [Google Scholar] [CrossRef] [Green Version]
- Wu, G.; Guo, J.-J.; Ma, Z.-Y.; Wang, J.; Zhou, Z.-W.; Wang, Y. Correlation between calcification and bone sialoprotein and osteopontin in papillary thyroid carcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 2010–2017. [Google Scholar] [PubMed]
- Ferreira, L.B.; Eloy, C.; Pestana, A.; Lyra, J.; Moura, M.; Prazeres, H.; Tavares, C.; Sobrinho-Simões, M.; Gimba, E.R.P.; Soares, P. Osteopontin expression is correlated with differentiation and good prognosis in medullary thyroid carcinoma. Eur. J. Endocrinol. 2016, 174, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Chernaya, G.; Mikhno, N.; Хабалoва, T.; Svyatchenko, S.; Mostovich, L.; Shevchenko, S.; Gulyaeva, L. The expression profile of integrin receptors and osteopontin in thyroid malignancies varies depending on the tumor progression rate and presence of BRAF V600E mutation. Surg. Oncol. 2018, 27, 702–708. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, L.B.; Lima, R.T.; Bastos, A.C.S.D.F.; Silva, A.M.; Tavares, C.; Pestana, A.; Rios, E.; Eloy, C.; Sobrinho-Simões, M.; Gimba, E.R.P.; et al. OPNa Overexpression Is Associated with Matrix Calcification in Thyroid Cancer Cell Lines. Int. J. Mol. Sci. 2018, 19, 2990. [Google Scholar] [CrossRef] [Green Version]
- Park, S.; Zhu, J.; Altan-Bonnet, G.; Cheng, S.-Y. Monocyte recruitment and activated inflammation are associated with thyroid carcinogenesis in a mouse model. Am. J. Cancer Res. 2019, 9, 1439–1453. [Google Scholar]
- Lloyd, R.V.; Buehler, D.; Khanafshar, E. Papillary Thyroid Carcinoma Variants. Head Neck Pathol. 2011, 5, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-H.; Park, C.-S.; Kim, Y.-I.; Nam-Goong, I.-S.; Kim, Y.-S.; Lee, J.-C.; Choi, J.-I.; Park, J.-W.; Kim, E.-S. Osteopontin levels in patients with papillary thyroid cancer according to the presence of Hashimoto’s thyroiditis. Asian Pac. J. Cancer Prev. 2015, 16, 2447–2451. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, L.B.; Gimba, E.; Vinagre, J.; Sobrinho-Simões, M.; Soares, P. Molecular Aspects of Thyroid Calcification. Int. J. Mol. Sci. 2020, 21, 7718. [Google Scholar] [CrossRef] [PubMed]
- De Falco, V.; Tamburrino, A.; Ventre, S.; Castellone, M.; Malek, M.; Manié, S.N.; Santoro, M. CD44 Proteolysis Increases CREB Phosphorylation and Sustains Proliferation of Thyroid Cancer Cells. Cancer Res. 2012, 72, 1449–1458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.; Fan, Y.; Zhang, X. Molecular Mechanisms in Differentiated Thyroid Cancer. Front. Biosci. 2016, 21, 119–129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikiforov, Y.E.; Nikiforova, M.N. Molecular genetics and diagnosis of thyroid cancer. Nat. Rev. Endocrinol. 2011, 7, 569–580. [Google Scholar] [CrossRef] [PubMed]
- Knauf, J.A.; Kuroda, H.; Basu, S.; Fagin, J.A. RET/PTC-induced dedifferentiation of thyroid cells is mediated through Y1062 signaling through SHC-RAS-MAP kinase. Oncogene 2003, 22, 4406–4412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baquero, P.; Sánchez-Hernández, I.; Jiménez-Mora, E.; Orgaz, J.; Jiménez, B.; Chiloeches, A. V600EBRAF promotes invasiveness of thyroid cancer cells by decreasing E-cadherin expression through a Snail-dependent mechanism. Cancer Lett. 2013, 335, 232–241. [Google Scholar] [CrossRef] [Green Version]
- Baquero, P.; Jiménez-Mora, E.; Santos, A.; Lasa, M.; Chiloeches, A. TGFβ induces epithelial-mesenchymal transition of thyroid cancer cells by both the BRAF/MEK/ERK and Src/FAK pathways. Mol. Carcinog. 2016, 55, 1639–1654. [Google Scholar] [CrossRef] [PubMed]
- Wai, P.Y.; Kuo, P.C. Osteopontin: Regulation in tumor metastasis. Cancer Metastasis Rev. 2007, 27, 103–118. [Google Scholar] [CrossRef] [PubMed]
- Kaomongkolgit, R.; Manokawinchoke, J.; Sanchavanakit, N.; Pavasant, P.; Sumrejkanchanakij, P. Fibronectin supports TNF-α-induced osteopontin expression through β1 integrin and ERK in HN-22 cells. Arch. Oral Biol. 2010, 55, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Liu, C.; Zhang, Q.-H.; Chen, L.; Shen, Y.-Y.; Chen, Y.-J.; Zeng, X.; Zu, X.-Y.; Cao, R.-X. TGF-β1 induces HMGA1 expression: The role of HMGA1 in thyroid cancer proliferation and invasion. Int. J. Oncol. 2017, 50, 1567–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]


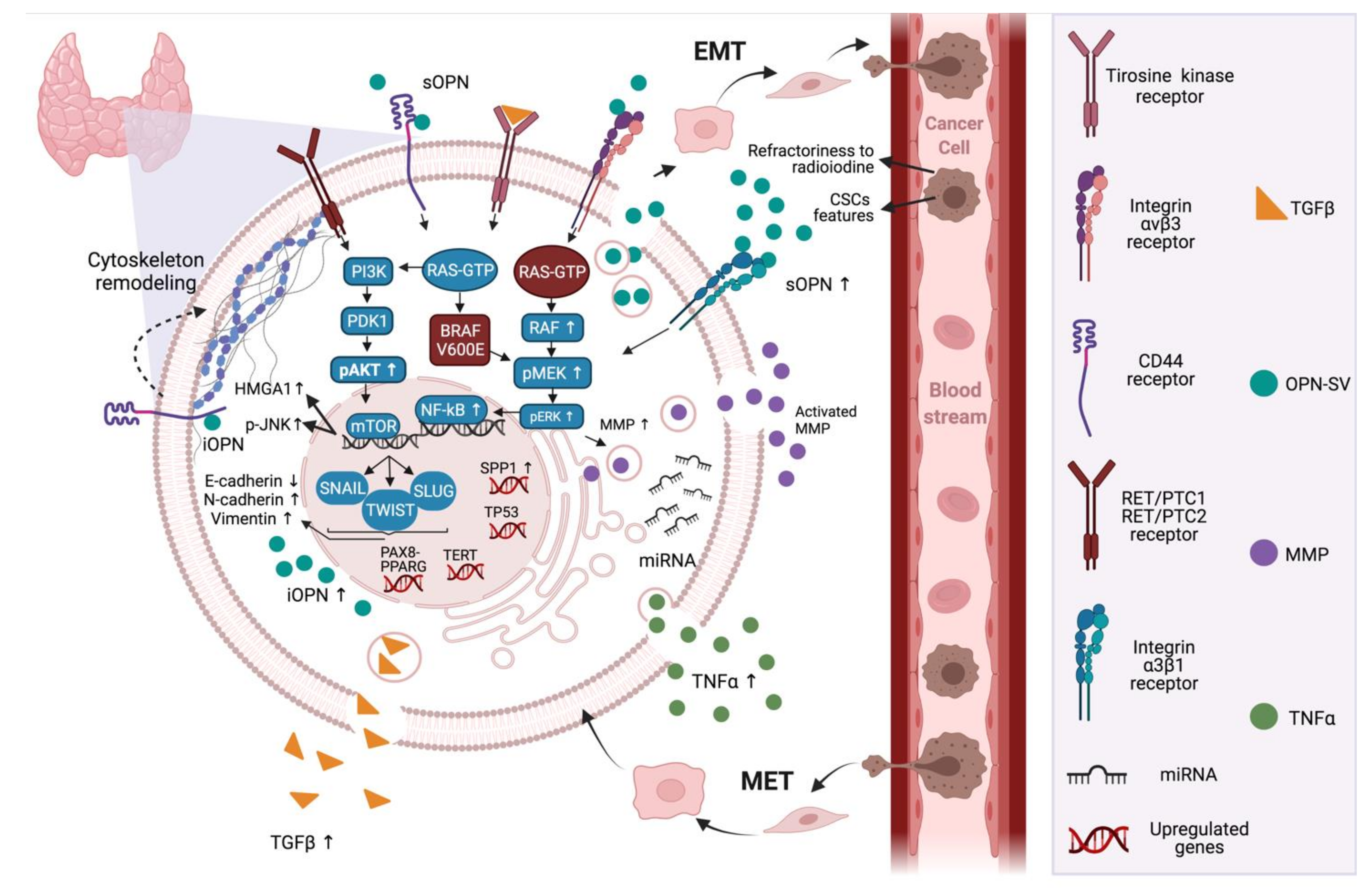{kind=link}
| Thyroid Tumor Model | EMT-Associated Biological Role | Bibliographic Reference |
|---|---|---|
| FTC cell line (WRO) PTC cell line (NPA) ATC cell line (KTC-3) | Induced BRAF V600E mutation expression was related to high pMEK, pERK, NF-κB and MMPs expression and increased cell invasion | [31] |
| PTC samples PTC cell line (NPA) ATC cell line (ARO) | In papillary thyroid carcinoma, RUNX2 and fibronectin1 had high expression in the tumor invasive area. Also, vimentin presented high levels in PTC, which was related to cell invasion and metastatic tumors | [32] |
| ATC, PTC and FTC samples ATC-derived cells | In ATC samples, miR-200 and miR-30 were decreased while ZEB1/2 and SMAD2 were upregulated in ATC samples in comparison to papillary and follicular tumors. Also, miR-200 and miR-30 overexpression in ATC induced an epithelial-like phenotype, harboring low vimentin levels and decreased cell invasion | [33] |
| PTC samples | Cells in papillary tumors invasive front with loss of cohesiveness and cell polarity had decreased membrane E-cadherin expression and were related to lymph node metastases | [34] |
| cPTC, FV-PTC, FTC and ATC samples PTC cell line (TPC-1) | Anaplastic tumors were related to decreased survival rates, high ZEB1 and SMAD7 and low E-cadherin expression in comparison to papillary and follicular carcinomas | [35] |
| ATC cell lines (ACT-1, FRO, KTC-2 and KTC-3) PTC cell line (TPC-1) | SNAIL overexpression in cell lines derived from anaplastic carcinomas generated E-cadherin downregulation, vimentin high expression and enhanced cell migration | [36] |
| PTC cell lines (BCPAP and TPC-1) ATC cell lines (THJ-16T and THJ-21T) | TGFβ treatment induced PRRX1, TWIST, SLUG, SNAIL and vimentin high expression and E-cadherin and cytokeratin 19 downregulation in papillary carcinoma derived cell lines, besides stemness properties and efficient tumor formation in mice | [37] |
| PTC samples PTC cell line (TPC-1) FTC cell lines (ML1, FTC236 and FTC 238) ATC cell lines (T238 and SW1736) | Stemness-related genes OCT4, ABCG2, CD44, NANOG, and SSEA-1 exhibited high expression in thyroid tumor cells when compared to normal thyroid, which were upregulated by Fluorouracil chemotherapy treatment | [38] |
| PTC tissue samples PTC cell lines (IHH-4, TPC1 and BCPAP) | In papillary thyroid carcinoma, TWIST1 exhibited high expression in comparison to normal adjacent thyroid cells and was correlated to lymph node metastasis, cell invasion and vimentin expression | [39] |
| PTC cell line (BCPAP) ATC cell line (8505c) Athymic nude BALB/c mice | ATC cells showed enhanced cell migration and high p-c-MET, p-AKT, vimentin, β-catenin and CD44 expression, besides high tumorigenicity, upon BRAF V600E inhibiton, demonstrating EMT-related chemoresistance | [40] |
| MTC samples MTC cell line (TT) | The chemokine receptor CXCR4 expression was high in medullary thyroid carcinoma in comparison to normal thyroid cells, which was also related to bigger and advanced stage tumors and lymph node metastasis. Cell treatment with chemokine SDF1α, ligand to CXCR4, enhanced MTC cell invasion | [41] |
| Papillary thyroid microcarcinoma (PTMC) samples | Stemness genes ALDH1A3, TM4SF1, PROM1 and EMT-associated CAV1 gene had high expression in PMTC with neck-node metastases in comparison with non-metastatic papillary microcarcinomas | [42] |
| FTC, PDTC and ATC samples | SNAIL, SLUG and TWIST had high expression in widely invasive FTC, ATC and PDTC. Also, high TWIST levels were correlated with vascular invasion and poor disease-free survival | [43] |
| Thyroid Tumor Model | OPN Expression | EMT-Associated Biological Role | Bibliographic Reference |
|---|---|---|---|
| PCCL3, TPC-1 and FB2 cell lines | High OPN expression was related to RET/PTC1, RET/PTC3, CD44, HA-RAS and MEK1 | OPN treatment generated increased cell proliferation, invasion and dissemination | [69] |
| Non-tumor thyroid cells overexpressing RET/PTC1 PTC and FTC samples | High OPN expression in RET/PTC1-thyrocytes and in PTC samples | High OPN expression generated increased proliferation and high p-AKT, p-ERK1/2, MMP-2 and MMP-9 | [70] |
| cPTC, FV-PTC and TCV-PTC samples PTC cell lines (TPC1, FB2, BHP2-7, BHP7-13, BHP10-3, BHP5-16, BHP14-9, BHP17-10, NPA and BCPAP) | OPN had higher expression in cPTC and TCV-PTC | OPN was related to lymph node metastasis, cell invasion and high levels of CD44, p44/42 MAPK and p-AKT | [71] |
| PTC and FTC samples FTC cell line (WRO) PTC cell line (NPA) | Higher OPN expression in PTC | OPN expression was correlated with the occurrence of lymph node metastasis | [72] |
| PTC, FTC, ATC and MTC samples | Tumor tissues had high OPN expression | MTC and ATC lymph node metastasis had higher OPN expression compared to their primary tumors | [73] |
| cPTC, FV-PTC and other PTC variants samples | OPN had higher expression in PTC | OPN expression was positively correlated with p-JNK | [74] |
| PTC samples | PTC samples had high OPN expression | OPN expression was related to microcalcification and lymph node metastasis | [75] |
| cPTC, FV-PTC and TCV-PTC samples | OPN had high expression in PTC | High OPN expression was related to lymph node metastasis, advanced N stage and BRAF V599E mutation | [76] |
| PTC samples | OPN -443 C>T polymorphism exhibited high expression in PTC | OPN -443 C>T was related to higher PTC and cervical lymph node metastasis risk and angiolymphatic invasion | [77] |
| PTC cell lines (B-CPAP and TPC1) | OPN expression was upregulated and downregulated by HGMA1b and CBX7, respectively | OPN expression induced increased cell migration | [78] |
| PTC samples | PTC samples exhibited high OPN expression | High OPN expression was related to calcification areas | [79] |
| MTC samples MTC cell lines (TT and MZ-CRC-1) | C-cells and C-cell hyperplasia exhibited higher OPN expression than MTC samplesOPNa was the most expressed OPN-SV | High expression of OPN in small, non-invasive tumors with PTEN nuclear expression and wild-type RAS | [80] |
| cPTC, FV-PTC and FTC samples PTC cell lines (TPC1, K1 and BCPAP) ATC cell lines (KAT4, Hth74, 8505c and c643) | cPTC exhibited higher OPN expression OPNa was the most expressed OPN-SV | OPNa expression was related to vascular invasion, extrathyroid extension, cell migration and invasion and MMPs activity | [9] |
| PTC samples | PTC lymph node metastasis samples showed OPNb high expression | OPNb and integrin α5had high expression in metastatic samples | [81] |
| cPTC samples PTC cell line (TPC1) ATC cell line (c643) | OPN exhibited high expression in cPTC samples Among OPN-SV, OPNa and OPNb were the most expressed | OPN expression in psammoma bodies was related to lymph node metastasis OPNa overexpression induced matrix calcification and collagen synthesis | [82] |
| ThrbPV/PVPten+/− mice with aggressive FTC | OPN showed high levels in ThrbPV/PVPten+/− mice | In ThrbPV/PVPten+/− tumors, OPN, integrin β1, pAKT, p65, TGF-β1 and TNF-α had high expression | [83] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Viana, B.P.P.B.; Gomes, A.V.P.; Gimba, E.R.P.; Ferreira, L.B. Osteopontin Expression in Thyroid Cancer: Deciphering EMT-Related Molecular Mechanisms. Biomedicines 2021, 9, 1372. https://doi.org/10.3390/biomedicines9101372
Viana BPPB, Gomes AVP, Gimba ERP, Ferreira LB. Osteopontin Expression in Thyroid Cancer: Deciphering EMT-Related Molecular Mechanisms. Biomedicines. 2021; 9(10):1372. https://doi.org/10.3390/biomedicines9101372
Chicago/Turabian StyleViana, Bruna Prunes Pena Baroni, Amanda Vitória Pampolha Gomes, Etel Rodrigues Pereira Gimba, and Luciana Bueno Ferreira. 2021. "Osteopontin Expression in Thyroid Cancer: Deciphering EMT-Related Molecular Mechanisms" Biomedicines 9, no. 10: 1372. https://doi.org/10.3390/biomedicines9101372
APA StyleViana, B. P. P. B., Gomes, A. V. P., Gimba, E. R. P., & Ferreira, L. B. (2021). Osteopontin Expression in Thyroid Cancer: Deciphering EMT-Related Molecular Mechanisms. Biomedicines, 9(10), 1372. https://doi.org/10.3390/biomedicines9101372