
Relationships of Ischemic Stroke Occurrence and Outcome with Gene Variants Encoding Enzymes of Tryptophan Metabolism

 , ,
, , 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Sample Collection and Processing
2.3. DNA Isolation, Genotyping
2.4. RNA Isolation, Gene Expression Analysis
2.5. Statistical Analysis
3. Results
3.1. Genotype and Allele Distribution of the Investigated SNPs


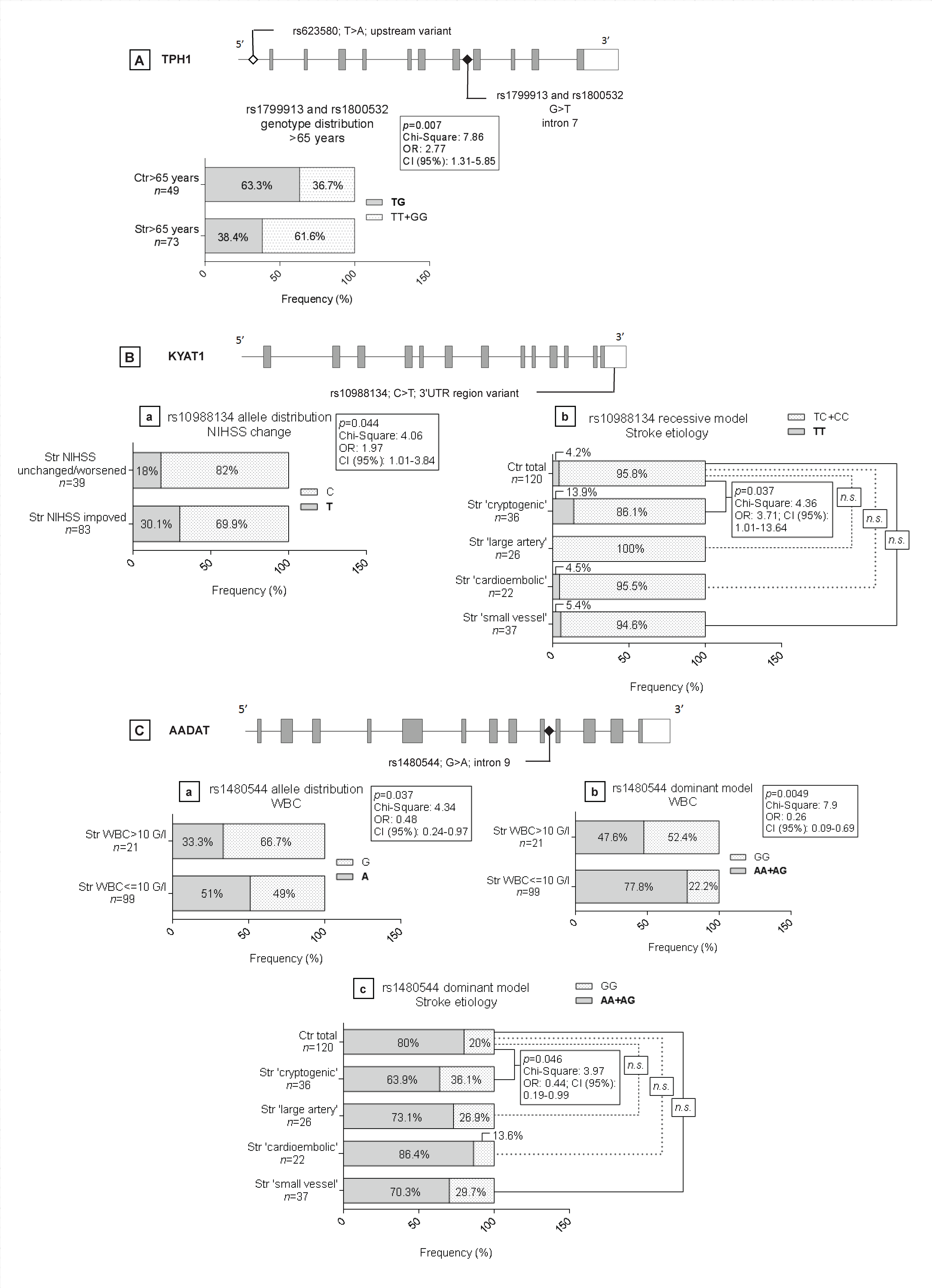
3.1.1. TPH1 Variants rs1799913, rs1800532 and rs623580
3.1.2. TPH2 Variants rs7963803 and rs4570625
3.1.3. KAT1 Variant rs10988134
3.1.4. AADAT Variant rs1480544
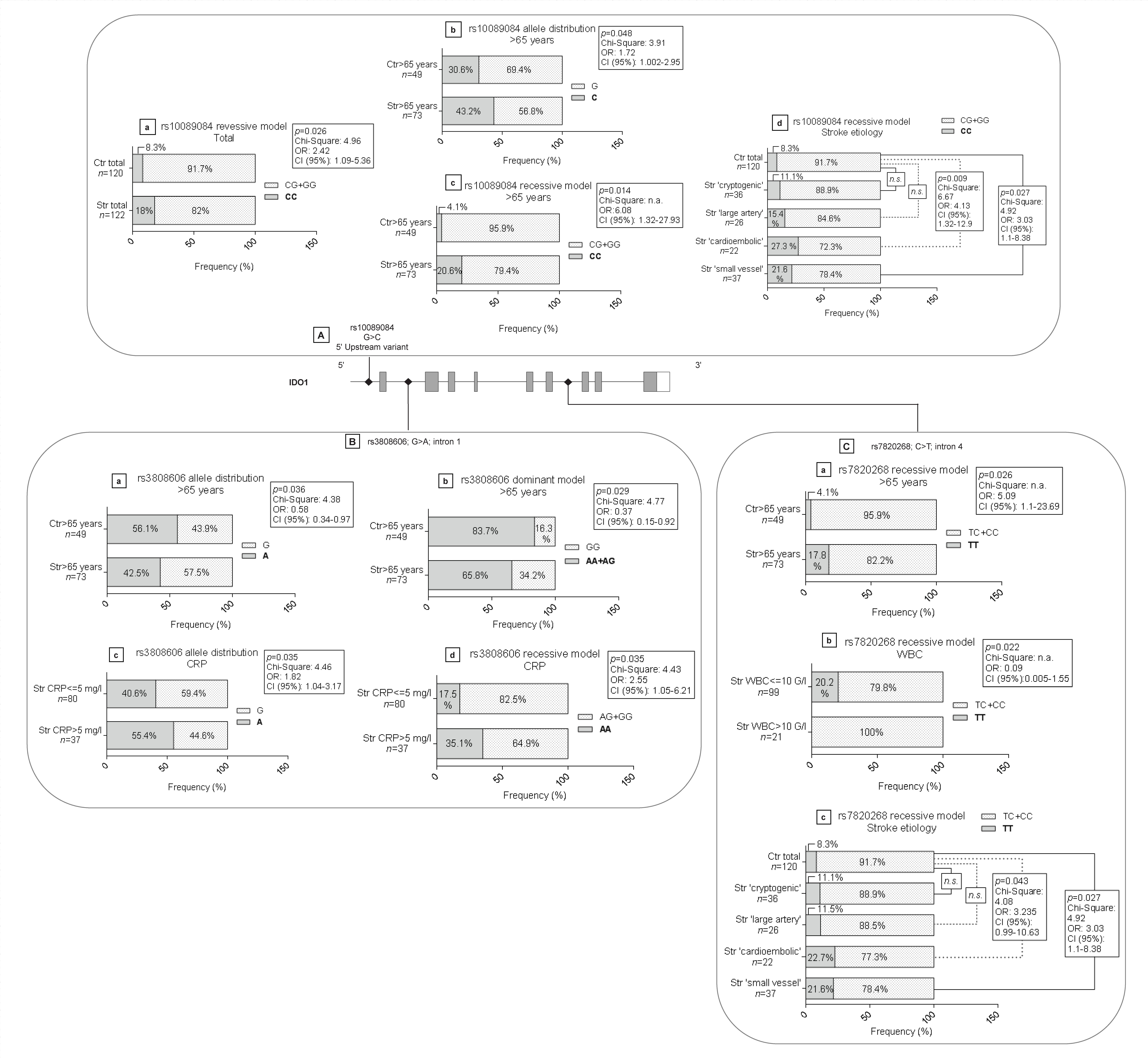
3.1.5. IDO1 Variants rs3808606, rs10089084 and rs7820268
3.2. Haplotype Blocks
3.3. Relationship of Clinical Parameters and Short Term Stroke Outcome
3.4. Gene Expression Analysis
3.4.1. Effects of Gene Variants on mRNA Levels in Blood Samples
3.4.2. Changes in TPH1, IDO1 and KYAT1 mRNA Levels during the Course of Ischemic Stroke
4. Discussion
5. Conclusions
6. Key Summary Points
- Despite the striking improvement of acute stroke management ischemic stroke is still among the leading causes of mortality and long-term disability worldwide.
- Our aim was to investigate the possible association between ischemic stroke and alterations of Trp metabolism-related genes in regard to both genomic variants and gene expression changes.
- Out of the ten studied polymorphisms we detected significant differences in the frequencies of seven variants in relation to ischemic stroke occurrence, disease etiology, inflammatory markers, and short-term disease outcome.
- We detected changes in TPH1 and IDO1 mRNA levels during the course of the disease.
- IDO1 variants showing a trend towards elevated mRNA level were more frequent in stroke patients than in controls, while IDO1 variants which lead to a trend of decreasing mRNA level are present among stroke patients less frequently than in non-stroke controls.
- Our results are important novel observations which suggest a causal relationship between elevated IDO1 expression and stroke etiology.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| 18S rRNA | 18S ribosomal RNA |
| 3-HAA | 3-hydroxyanthranilic acid |
| 3-HAO | 3-hydroxyanthranilate 3,4-dioxygenase |
| 3HK | 3-hydroxy-kynurenine |
| AA | anthranilic acid |
| ACMSD | aminocarboxymuconate semialdehyde decarboxylase |
| AADAT | kynurenine/alpha-aminoadipate aminotransferase |
| CI | confidence interval |
| CNS | central nervous system |
| CRP | C-reactive protein |
| CSF | cerebrospinal fluid |
| DM | diabetes mellitus |
| DtN | door-to-needle (time) |
| HT | hypertension |
| HL | hyperlipidaemia |
| IDO1 | indoleamine 2,3-dioxygenase 1 |
| KAT | kynurenine aminotransferase |
| KMO | kynurenine monooxygenase |
| KYAT1 | kynurenine aminotransferase 1 |
| KYN | kynurenine |
| KYNA | kynurenic acid |
| KYNU | kynureninase |
| KP | kynurenine pathway |
| LD | linkage disequilibrium |
| LDL | low density lipoprotein |
| mRS | modified Rankin Scale |
| NIHSS | National Institutes of Health Stroke Scale |
| OR | odds ratio |
| PBS | peripheral blood sample |
| RT-qPCR | reverse transcription-qPCR |
| QA | quinolinic acid |
| qPCR | quantitative polymerase chain reaction |
| SNP | single nucleotide polymorphism |
| TDO2 | tryptophan 2,3-dioxygenase |
| TOAST | Trial of Org 10172 in Acute Stroke Treatment |
| TPH1 | tryptophan hydroxylase 1 |
| TPH2 | tryptophan hydroxylase 2 |
| UN | urea nitrogen |
| WBC | white blood cell |
References
- Boehme, A.K.; Esenwa, C.; Elkind, M.S.V. Stroke Risk Factors, Genetics and Prevention. Circ. Res. 2017, 120, 472–495. [Google Scholar] [CrossRef]
- Colpo, G.D.; Venna, V.R.; McCullough, L.D.; Teixeira, A.L. Systematic Review on the Involvement of the Kynurenine Pathway in Stroke: Pre-clinical and Clinical Evidence. Front. Neurol. 2019, 10, 778. [Google Scholar] [CrossRef] [PubMed]
- Carlsson, G.E.; Möller, A.; Blomstrand, C. Consequences of mild stroke in persons. Cerebrovasc. Dis. 2003, 16, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Sims, K.; Politei, J.; Banikazemi, M.; Lee, P. Stroke in Fabry Disease Frequently Occurs Before Diagnosis and in the Absence of Other Clinical Events. Stroke 2009, 40, 788–794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wityk, R.J.; Zanferrari, C.; Oppenheimer, S. Neurovascular Complications of Marfan Syndrome. Stroke 2002, 33, 680–684. [Google Scholar] [CrossRef]
- Verduzco, L.A.; Nathan, D.G. Sickle cell disease and stroke. Blood 2009, 114, 5117–5125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joutel, A.; Corpechot, C.; Ducros, A.; Vahedi, K.; Chabriat, H.; Mouton, P.; Alamowitch, S.; Domenga, V.; Cécillion, M.; Maréchal, E.; et al. Notch3 mutations in CADASIL, a hereditary adult-onset condition causing stroke and dementia. Nat. Cell Biol. 1996, 383, 707–710. [Google Scholar] [CrossRef]
- Cordeiro, I.M.; Nzwalo, H.; Isabel, A.; Ferreira, R.B.; Alonso, I.; Afonso, L.; Basílio, C. Shifting the CARASIL Paradigm. Stroke 2015, 46, 1110–1112. [Google Scholar] [CrossRef] [Green Version]
- Meyer-Schaller, N.; Heck, C.; Tiede, S.; Yilmaz, M.; Christofori, G. Foxf2 plays a dual role during transforming growth factor beta-induced epithelial to mesenchymal transition by promoting apoptosis yet enabling cell junction dissolution and migration. Breast Cancer Res. 2018, 20, 118. [Google Scholar] [CrossRef] [PubMed]
- Scheu, S.; Ruland, C.; Lahl, K.; Zimmer, A.; Werner, H.; Förste, I.; Alferink, J. Tspan2 is involved in anti-infectious immune responses and CNS autoimmunity. J. Immunol. 2020, 204. [Google Scholar]
- Joy, M.T.; Ben Assayag, E.; Shabashov-Stone, D.; Liraz-Zaltsman, S.; Mazzitelli, J.; Arenas, M.; Abduljawad, N.; Kliper, E.; Korczyn, A.D.; Thareja, N.S.; et al. CCR5 Is a Therapeutic Target for Recovery after Stroke and Traumatic Brain Injury. Cell 2019, 176, 1143–1157.e13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vécsei, L.; Szalardy, L.; Fülöp, F.; Toldi, J. Kynurenines in the CNS: Recent advances and new questions. Nat. Rev. Drug Discov. 2012, 12, 64–82. [Google Scholar] [CrossRef] [PubMed]
- Hertelendy, P.; Toldi, J.; Fülöp, F.; Vécsei, L. Ischemic Stroke and Kynurenines: Medicinal Chemistry Aspects. Curr. Med. Chem. 2019, 25, 5945–5957. [Google Scholar] [CrossRef]
- Darlington, L.G.; Mackay, G.M.; Forrest, C.M.; Stoy, N.; George, C.; Stone, T.W. Altered kynurenine metabolism correlates with infarct volume in stroke. Eur. J. Neurosci. 2007, 26, 2211–2221. [Google Scholar] [CrossRef]
- Brouns, R.; Verkerk, R.; Aerts, T.; De Surgeloose, D.; Wauters, A.; Scharpe, S.; De Deyn, P.P. The Role of Tryptophan Catabolism along the Kynurenine Pathway in Acute Ischemic Stroke. Neurochem. Res. 2010, 35, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- Hajsl, M.; Hlavackova, A.; Broulikova, K.; Sramek, M.; Maly, M.; Dyr, J.E.; Suttnar, J. Tryptophan Metabolism, Inflammation, and Oxidative Stress in Patients with Neurovascular Disease. Metabolites 2020, 10, 208. [Google Scholar] [CrossRef]
- Schwarcz, R.; Bruno, J.P.; Muchowski, P.J.; Wu, H.-Q. Kynurenines in the mammalian brain: When physiology meets pathology. Nat. Rev. Neurosci. 2012, 13, 465–477. [Google Scholar] [CrossRef]
- Mo, X.; Pi, L.; Yang, J.; Xiang, Z.; Tang, A. Serum indoleamine 2,3-dioxygenase and kynurenine aminotransferase enzyme activity in patients with ischemic stroke. J. Clin. Neurosci. 2014, 21, 482–486. [Google Scholar] [CrossRef]
- Wigner, P.; Saluk-Bijak, J.; Synowiec, E.; Miller, E.; Sliwinski, T.; Cichon, N.; Bijak, M. Variation of Genes Encoding Tryptophan Catabolites Pathway Enzymes in Stroke. J. Clin. Med. 2019, 8, 2133. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.A.; Dykes, D.D.; Polesky, H.F. A simple salting out procedre for extracting DNA from human nucleated cells. Nucleid Acids Res. 1988, 16, 1215. [Google Scholar] [CrossRef] [Green Version]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.D.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Shaun Purcell. PLINK, version 1.9; Center for Human Genetic Research (CHGR), Massachusetts General Hospital (MGH), Broad Institute of Harvard & MIT: Cambridge, MA, USA, 2014. [Google Scholar]
- Haploview, version 4.2; Broad Institute of Harvard and MIT: Cambridge, MA, USA, 2005.
- Barrett, J.C.; Fry, B.; Maller, J.; Daly, M.J. Haploview: Analysis and visualization of LD and haplotype maps. Bioinformatics 2004, 21, 263–265. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- GraphPad Prism, version 6.01; GraphPad Software: San Diego, CA, USA, 2012.
- The 1000 Genomes Project Consortium; Auton, A.; Brooks, L.D.; Durbin, R.M.; Garrison, E.P.; Kang, H.M.; Korbel, J.O.; Marchini, J.L.; McCarthy, S.; McVean, G.A.; et al. A global reference for human genetic variation. Nature 2015, 526, 68–74. [Google Scholar] [CrossRef] [Green Version]
- Moore, L.E.; Boffetta, P.; Karami, S.; Brennan, P.; Stewart, P.S.; Hung, R.; Zaridze, D.; Matveev, V.; Janout, V.; Kollárová, H.; et al. Occupational Trichloroethylene Exposure and Renal Carcinoma Risk: Evidence of Genetic Susceptibility by Reductive Metabolism Gene Variants. Cancer Res. 2010, 70, 6527–6536. [Google Scholar] [CrossRef] [Green Version]
- Královičová, J.; Vořechovský, I. Global control of aberrant splice-site activation by auxiliary splicing sequences: Evidence for a gradient in exon and intron definition. Nucleic Acids Res. 2007, 35, 6399–6413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Souza, F.R.S.; Fontes, F.L.; Da Silva, T.A.; Coutinho, L.G.; Leib, S.L.; Agnez-Lima, L.F. Association of kynurenine aminotransferase II gene C401T polymorphism with immune response in patients with meningitis. BMC Med. Genet. 2011, 12, 51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Napolioni, V.; Pariano, M.; Borghi, M.; Oikonomou, V.; Galosi, C.; De Luca, A.; Stincardini, C.; Vacca, C.; Renga, G.; Lucidi, V.; et al. Genetic Polymorphisms Affecting IDO1 or IDO2 Activity Differently Associate with Aspergillosis in Humans. Front. Immunol. 2019, 10, 890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Guillemin, G.J. Kynurenine Pathway Metabolites in Humans: Disease and Healthy States. Int. J. Tryptophan Res. 2009, 2, IJTR.S2097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, S.-J.; Wang, Y.-C.; Chen, J.-Y.; Hong, C.-J. Allelic variants of the tryptophan hydroxylase (A218C) and serotonin 1B receptor (A-161T) and personality traits. Neuropsychobiology 2003, 48, 68–71. [Google Scholar] [CrossRef]
- Suzuki, A.; Fukasawa, T.; Shiraishi, H.; Ishii, G.; Oshino, S.; Aoshima, T.; Otani, K. No association between the TPH A218C polymorphism and personality traits in Japanese healthy subjects. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2007, 31, 395–398. [Google Scholar] [CrossRef] [PubMed]
- Anghelescu, I.; Klawe, C.; Fehr, C.; Singer, P.; Schleicher, A.; Himmerich, H.; Hiemke, C.; Dahmen, N.; Szegedi, A. The TPH intron 7 A218C polymorphism and TCI dimension scores in alcohol-dependent patients: Hints to nonspecific psychopathology. Addict. Behav. 2005, 30, 1135–1143. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, D.A.; Barral, S.; Proudnikov, D.; Kellogg, S.; Ho, A.; Ott, J.; Kreek, M.J. TPH2 and TPH1: Association of Variants and Interactions with Heroin Addiction. Behav. Genet. 2008, 38, 133–150. [Google Scholar] [CrossRef]
- Li, D.; He, L. Meta-analysis shows association between the tryptophan hydroxylase (TPH) gene and schizophrenia. Qual. Life Res. 2006, 120, 22–30. [Google Scholar] [CrossRef]
- González-Castro, T.B.; Juárez-Rojop, I.; López-Narváez, M.L.; Tovilla-Zárate, C.A. Association of TPH-1 and TPH-2 gene polymorphisms with suicidal behavior: A systematic review and meta-analysis. BMC Psychiatry 2014, 14, 196. [Google Scholar] [CrossRef] [Green Version]
- Duan, X.; Gan, J.; Xu, F.; Li, L.; Han, L.; Peng, C.; Bao, Q.; Xiao, L.; Peng, D. RNA Sequencing for Gene Expression Profiles in a Rat Model of Middle Cerebral Artery Occlusion. BioMed Res. Int. 2018, 2018, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Walther, D.J.; Peter, J.-U.; Winter, S.; Höltje, M.; Paulmann, N.; Grohmann, M.; Vowinckel, J.; Alamo-Bethencourt, V.; Wilhelm, C.S.; Ahnert-Hilger, G.; et al. Serotonylation of Small GTPases Is a Signal Transduction Pathway that Triggers Platelet α-Granule Release. Cell 2003, 115, 851–862. [Google Scholar] [CrossRef] [Green Version]
- Peter, J.-U.; Alenina, N.; Bader, M.; Walther, D.J. Development of antithrombotic miniribozymes that target peripheral tryptophan hydroxylase. Mol. Cell. Biochem. 2006, 295, 205–215. [Google Scholar] [CrossRef]
- Orabona, C.; Mondanelli, G.; Pallotta, M.T.; Carvalho, A.; Albini, E.; Fallarino, F.; Vacca, C.; Volpi, C.; Belladonna, M.L.; Berioli, M.G.; et al. Deficiency of immunoregulatory indoleamine 2,3-dioxygenase 1in juvenile diabetes. JCI Insight 2018, 3, 96244. [Google Scholar] [CrossRef]
- Castell, J.V.; Gómez-Lechón, M.J.; David, M.; Andus, T.; Geiger, T.; Trullenque, R.; Fabra, R.; Heinrich, P.C. Interleukin-6 is the major regulator of acute phase protein synthesis in adult human hepatocytes. FEBS Lett. 1989, 242, 237–239. [Google Scholar] [CrossRef] [Green Version]
- Castell, J.V.; Andus, T.; Kunz, D.; Heinrich, P.C. Interleukin-6. Ann. N. Y. Acad. Sci. 1989, 557, 87–101. [Google Scholar] [CrossRef]
- Sehgal, P.B. Interleukin-6: A regulator of plasma protein gene expression in hepatic and non-hepatic tissues. Mol. Boil. Med. 1990, 7, 117–130. [Google Scholar]
- Mondanelli, G.; Carvalho, A.; Puccetti, P.; Grohmann, U.; Volpi, C. Reply to Han et al.: On track for an IDO1-based personalized therapy in autoimmunity. Proc. Natl. Acad. Sci. USA 2020, 117, 24037–24038. [Google Scholar] [CrossRef]
- Tardito, S.; Negrini, S.; Conteduca, G.; Ferrera, F.; Parodi, A.; Battaglia, F.; Kalli, F.; Fenoglio, D.; Cutolo, M.; Filaci, G. Indoleamine 2,3 dioxygenase gene polymorphisms correlate with CD8+ Treg impairment in systemic sclerosis. Hum. Immunol. 2013, 74, 166–169. [Google Scholar] [CrossRef] [PubMed]
- Han, Z.; He, D.; Zhang, Y. Genetic variant rs7820258 regulates the expression of indoleamine 2,3-dioxygenase 1 in brain regions. Proc. Natl. Acad. Sci. USA 2020, 117, 24035–24036. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, S.; Lawrence, C.B. Old Dog New Tricks; Revisiting How Stroke Modulates the Systemic Immune Landscape. Front. Neurol. 2019, 10, 718. [Google Scholar] [CrossRef] [PubMed]
- Chamorro, A.; Meisel, A.; Planas, A.M.; Urra, X.; van de Beek, D.; Veltkamp, R. The immunology of acute stroke. Nat. Rev. Neurol. 2012, 8, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Emsley, H.C.A.; Smith, C.; Tyrrell, P.; Hopkins, S. Inflammation in Acute Ischemic Stroke and its Relevance to Stroke Critical Care. Neurocrit. Care 2007, 9, 125–138. [Google Scholar] [CrossRef] [PubMed]
- Gruys, E.; Toussaint, M.J.M.; Niewold, T.A.; Koopmans, S.J. Acute phase reaction and acute phase proteins. J. Zhejiang Univ. Sci. B 2005, 6, 1045–1056. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oxenkrug, G.F. Genetic and Hormonal Regulation of Tryptophan-Kynurenine Metabolism. Ann. N. Y. Acad. Sci. 2007, 1122, 35–49. [Google Scholar] [CrossRef] [PubMed]
- Maes, M.; Leonard, B.; Myint, A.; Kubera, M.; Verkerk, R. The new ‘5-HT’ hypothesis of depression: Cell-mediated immune activation induces indoleamine 2,3-dioxygenase, which leads to lower plasma tryptophan and an increased synthesis of detrimental tryptophan catabolites (TRYCATs), both of which contribute to the onset of depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2011, 35, 702–721. [Google Scholar] [CrossRef]
- Kiank, C.; Zeden, J.-P.; Drude, S.; Domanska, G.; Fusch, G.; Otten, W.; Schuett, C. Psychological Stress-Induced, IDO1-Dependent Tryptophan Catabolism: Implications on Immunosuppression in Mice and Humans. PLoS ONE 2010, 5, e11825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mellor, A.L.; Lemos, H.D.P.; Huang, L. Indoleamine 2,3-Dioxygenase and Tolerance: Where Are We Now? Front. Immunol. 2017, 8, 1360. [Google Scholar] [CrossRef]
- Chang, C.; Fonseca, K.R.; Li, C.; Horner, W.; Zawadzke, L.E.; Salafia, M.A.; Welch, K.A.; Strick, C.A.; Campbell, B.M.; Gernhardt, S.S.; et al. Quantitative Translational Analysis of Brain Kynurenic Acid Modulation via Irreversible Kynurenine Aminotransferase II Inhibition. Mol. Pharmacol. 2018, 94, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Wigner, P.; Czarny, P.; Synowiec, E.; Bijak, M.; Talarowska, M.; Galecki, P.; Szemraj, J.; Sliwinski, T. Variation of genes encoding KAT1, AADAT and IDO1 as a potential risk of depression development. Eur. Psychiatry 2018, 52, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Rossi, F.; Miggiano, R.; Ferraris, D.M.; Rizzi, M. The Synthesis of Kynurenic Acid in Mammals: An Updated Kynurenine Aminotransferase Structural KATalogue. Front. Mol. Biosci. 2019, 6, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douet, V.; Tanizaki, N.; Franke, A.; Li, X.; Chang, L. Polymorphism of Kynurenine Pathway-Related Genes, Kynurenic Acid, and Psychopathological Symptoms in HIV. J. Neuroimmune Pharmacol. 2016, 11, 549–561. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | n | Age (Mean ± SD; Years) | Male/Female |
|---|---|---|---|
| Control (Ctr) | 120 | 64.94 ± 9.46 | 57/63 |
| Ctr > 65 years | 49 | 74.31 ± 6.75 | 16/33 |
| Stroke (Str) | 122 | 67.22 ± 10.28 | 69/53 |
| Str > 65 years | 73 | 74.19 ± 4.7 | 37/36 |
| Str TOAST: Large-artery atherosclerosis | 26 | 68.46 ± 9.45 | 20/6 |
| Str TOAST: Small-vessel occlusion | 37 | 65.95 ± 8.87 | 18/19 |
| Str TOAST: Cardioembolic | 22 | 72.68 ± 8.9 | 8/14 |
| Str TOAST: Cryptogenic | 36 | 64.47 ± 11.9 | 22/14 |
| Str (CRP ≤ 5 mg/L) | 80 | 67.54 ± 10.47 | 44/36 |
| Str (CRP > 5 mg/L) | 37 | 66.97 ± 10.45 | 22/15 |
| Str (WBC ≤ 10 G/L) | 99 | 68.21 ± 9.64 | 52/47 |
| Str (WBC > 10 G/L) | 21 | 61.95 ± 12.39 | 16/5 |
| Str (NIHSS: improved) | 83 | 67.8 ± 10.71 | 46/37 |
| Str (NIHSS: unchanged/worsened) | 39 | 65.82 ± 9.42 | 23/16 |
| (A) | ||||
| Clinical Parameter | n | % | Missing Data | |
| Alcohol consumption | No | 81 | 68.07 | 3 |
| Yes | 38 | 31.93 | ||
| Antiplatelet therapy | No | 82 | 67.21 | 0 |
| Yes | 40 | 32.79 | ||
| DM | No | 86 | 70.49 | 0 |
| Yes | 36 | 29.51 | ||
| ECG | SR | 92 | 79.31 | 6 |
| AF | 24 | 20.69 | ||
| TOAST classification | Cryptogenic | 36 | 29.75 | 1 |
| Cardioembolic | 22 | 18.18 | ||
| Large-artery atherosclerosis | 26 | 21.49 | ||
| Small-vessel occlusion | 37 | 30.58 | ||
| HT | No | 16 | 13.11 | 0 |
| Yes | 106 | 86.89 | ||
| CRP | ≤5 mg/L | 80 | 68.38 | 5 |
| >5 mg/L | 37 | 31.62 | ||
| WBC | ≤10 G/L | 99 | 82.5 | 2 |
| >10 G/L | 21 | 17.5 | ||
| HL | No | 21 | 18.1 | 6 |
| Yes | 95 | 81.9 | ||
| Previous stroke | No | 97 | 79.51 | 0 |
| Yes | 25 | 20.49 | ||
| Smoking | No | 78 | 66.1 | 4 |
| Yes | 40 | 33.9 | ||
| Thrombectomy | No | 103 | 84.43 | 0 |
| Yes | 19 | 15.57 | ||
| Baseline imaging | CT angio | 64 | 52.46 | 0 |
| Noncontrast CT | 32 | 26.23 | ||
| MRI | 26 | 21.31 | ||
| Type of lesion | Nonlacunar | 60 | 72.29 | 39 |
| (B) | ||||
| Clinical parameter | average ± SD | missing data | ||
| DtN (min) | 60.13 ± 41.14 | 2 | ||
| pre- stroke mRS (score) | 0.95 ± 1 | 5 | ||
| UN (mmol/L) | 6.43 ± 2 | 2 | ||
| Creat (μmol/L) | 89.3 ± 23.86 | 2 | ||
| WBC (G/L) | 8.38 ± 2.34 | 2 | ||
| LDL (mmol/L) | 2.85 ± 0.94 | 15 | ||
| Cholesterol (mmol/L) | 4.78 ± 1.19 | 4 | ||
| HgbA1c (%) | 6.47 ± 1.4 | 26 | ||
| TG (mmol/L) | 1.56 ± 1.21 | 4 | ||
| NIHSS baseline | 8.05 ± 4.69 | 0 | ||
| NIHSS discharge | 5.79 ± 5.96 | 0 | ||
| Group | n | versus | Group | n |
| Str total | 122 | Ctr total | 120 | |
| Str ≤ 65 years | 49 | Ctr ≤ 65 years | 71 | |
| Str > 65 years | 73 | Ctr > 65 years | 49 | |
| Str TOAST: Large-artery atherosclerosis | 26 | Ctr total | 120 | |
| Str TOAST: Small-vessel occlusion | 37 | Ctr total | 120 | |
| Str TOAST: Cardioembolic | 22 | Ctr total | 120 | |
| Str TOAST: Cryprogenic | 36 | Ctr total | 120 | |
| Str (CRP ≤ 5 mg/L) | 80 | Str (CRP > 5 mg/L) | 37 | |
| Str (WBC ≤ 10 G/L) | 99 | Str (WBC > 10 G/L) | 21 | |
| Str (NIHSS: improved) | 83 | Str (NIHSS: unchanged/worsened) | 39 |
| Primer Sequences | ||
|---|---|---|
| TPH1 | FW | 5′-TGCAGACCATCCTGGCTTCA-3′ |
| REV | 5′-GGAATACGGTTCCCCAGGTC-3′ | |
| IDO1 | FW | 5′-TCACAGACCACAAGTCACAG-3′ |
| REV | 5′-GCAAGACCTTACGGACATCT-3′ | |
| KYAT1 | FW | 5′-GCCATCCCTGTCTCCATCTT-3′ |
| REV | 5′-AGCGTGGCTTCATCCTTCAC-3′ | |
| 18S rRNA | FW | 5′-GCTTAATTTGACTCAACACGGGA-3′ |
| REV | 5′-AGCTATCAATCTGTCAATCCTGTC-3′ | |
| Cycling conditions | ||
| TPH1 | ||
| 95 °C | 3 min | |
| 95 °C | 15 s | 40× |
| 69 °C | 30 s | |
| IDO1 | ||
| 95 °C | 3 min | |
| 95 °C | 15 s | 40× |
| 68.4 °C | 30 s | |
| KYAT1 | ||
| 95 °C | 3 min | |
| 95 °C | 15 s | 40× |
| 64.8 °C | 15 s | |
| 18S rRNA | ||
| 50 °C | 2 min | |
| 95 °C | 2 min | |
| 95 °C | 30 s | 35× |
| 57 °C | 45 s | |
| 72 °C | 30 s | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boros, F.A.; Maszlag-Török, R.; Szűcs, M.; Annus, Á.; Klivényi, P.; Vécsei, L. Relationships of Ischemic Stroke Occurrence and Outcome with Gene Variants Encoding Enzymes of Tryptophan Metabolism. Biomedicines 2021, 9, 1441. https://doi.org/10.3390/biomedicines9101441
Boros FA, Maszlag-Török R, Szűcs M, Annus Á, Klivényi P, Vécsei L. Relationships of Ischemic Stroke Occurrence and Outcome with Gene Variants Encoding Enzymes of Tryptophan Metabolism. Biomedicines. 2021; 9(10):1441. https://doi.org/10.3390/biomedicines9101441
Chicago/Turabian StyleBoros, Fanni Annamária, Rita Maszlag-Török, Mónika Szűcs, Ádám Annus, Péter Klivényi, and László Vécsei. 2021. "Relationships of Ischemic Stroke Occurrence and Outcome with Gene Variants Encoding Enzymes of Tryptophan Metabolism" Biomedicines 9, no. 10: 1441. https://doi.org/10.3390/biomedicines9101441
APA StyleBoros, F. A., Maszlag-Török, R., Szűcs, M., Annus, Á., Klivényi, P., & Vécsei, L. (2021). Relationships of Ischemic Stroke Occurrence and Outcome with Gene Variants Encoding Enzymes of Tryptophan Metabolism. Biomedicines, 9(10), 1441. https://doi.org/10.3390/biomedicines9101441