
Pulsed Radiofrequency Upregulates Serotonin Transporters and Alleviates Neuropathic Pain-Induced Depression in a Spared Nerve Injury Rat Model

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
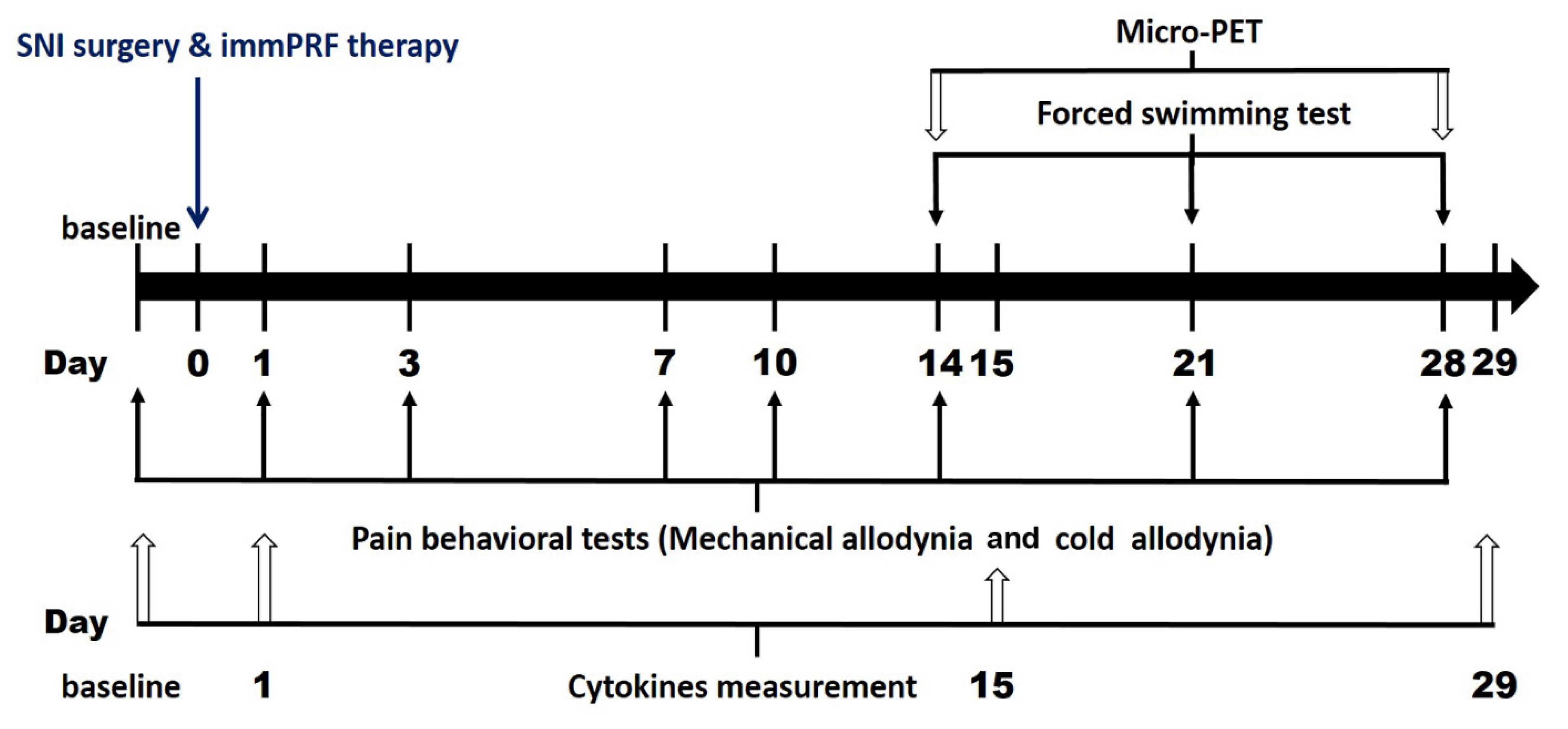
2.2. Experiments Schedule
2.3. Establishment of the Neuropathic Pain Model
2.4. Pulsed Radiofrequency Therapy
2.5. Pain and Depression-Like Behavioral Testing
2.6. PET Imaging
2.7. Assay of Serum Cytokines
2.8. Statistical Analysis
3. Results
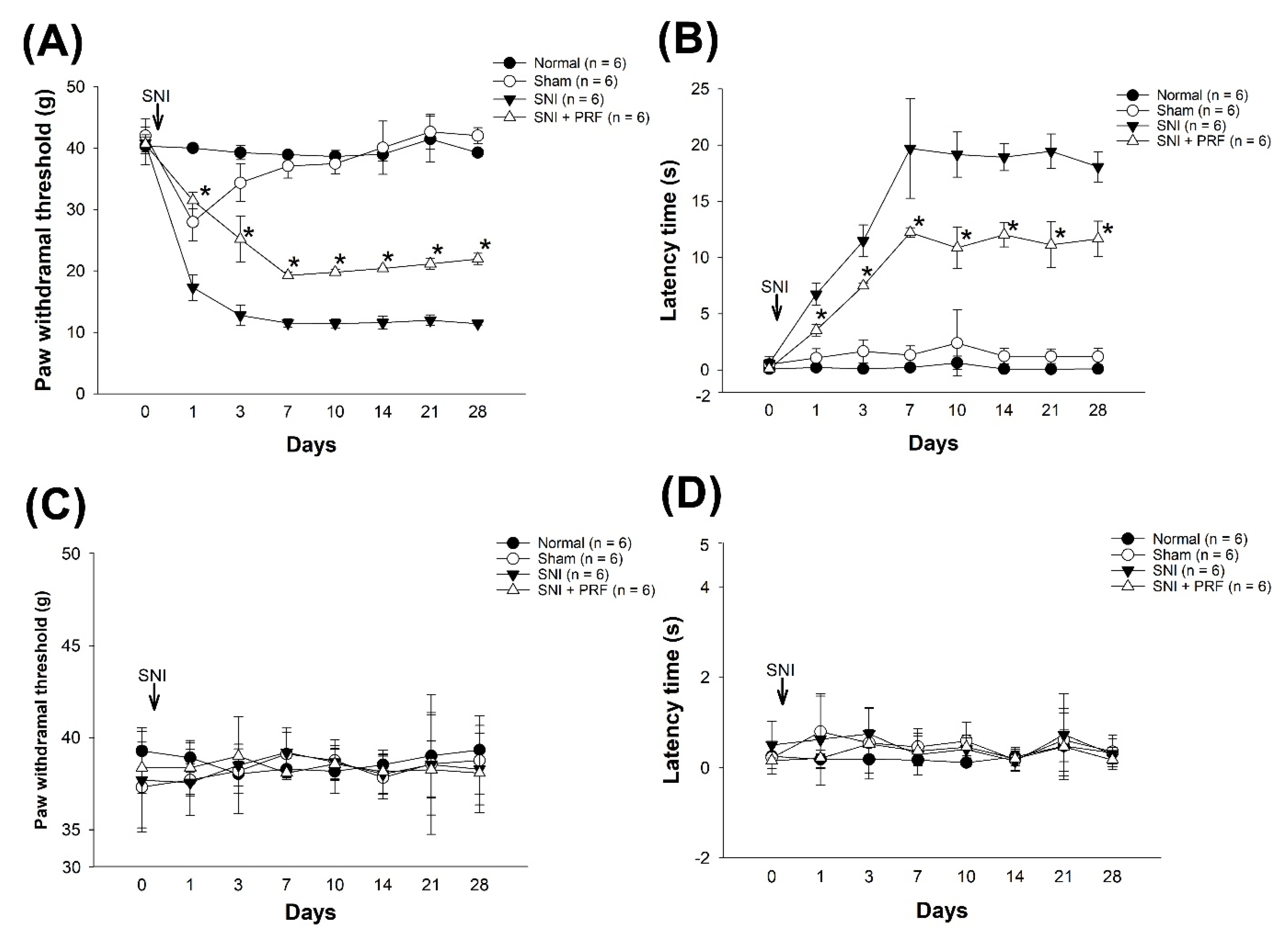
3.1. PRF Treatment Alleviated Mechanical and Cold Allodynia in an SNI-Induced Neuropathic Pain Model
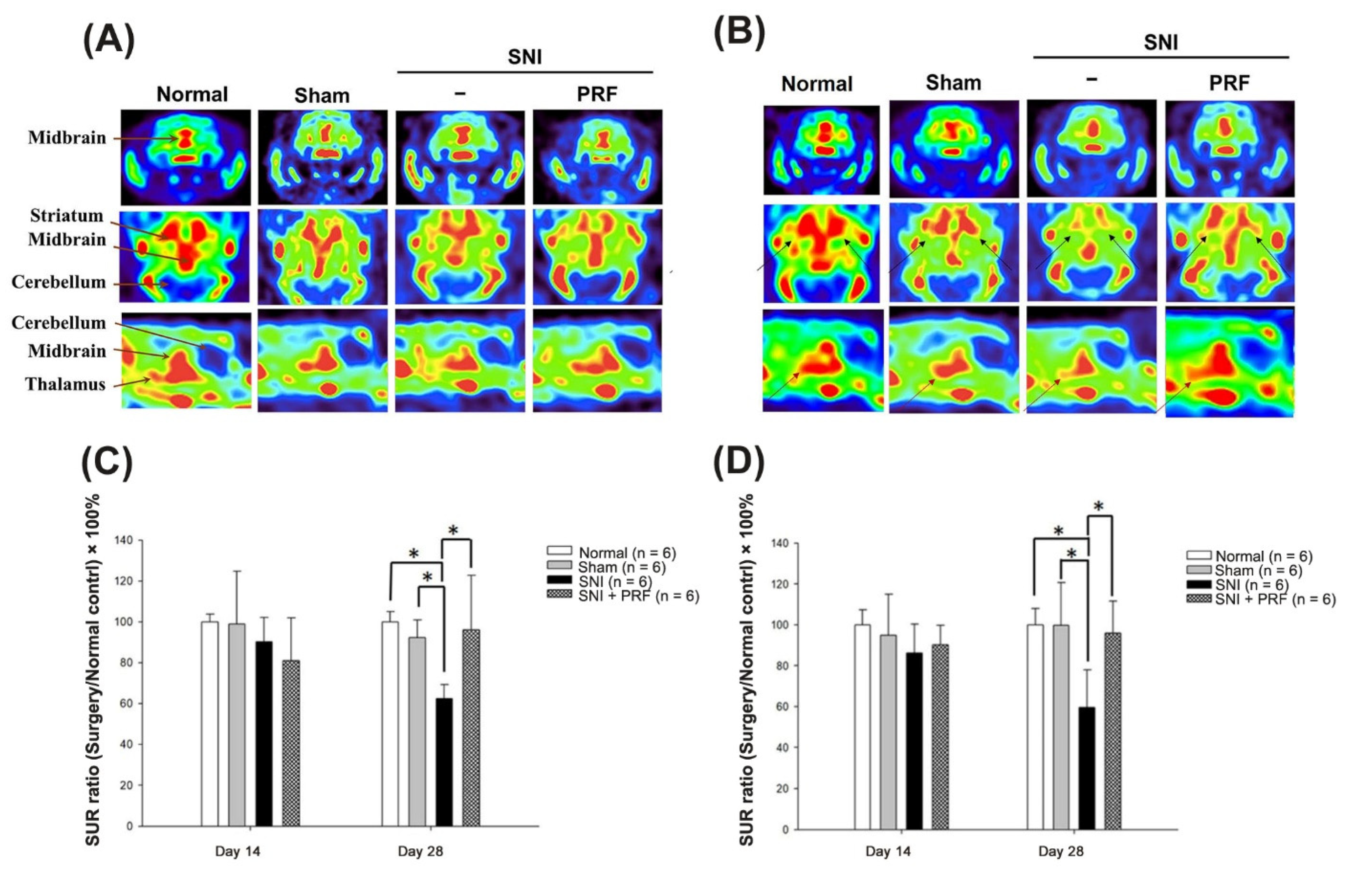
3.2. PET Images of 4-[18F]-ADAM in Normal Rats
3.3. PET Images and the SUR of 4-[18F]-ADAM in Rats after Surgery
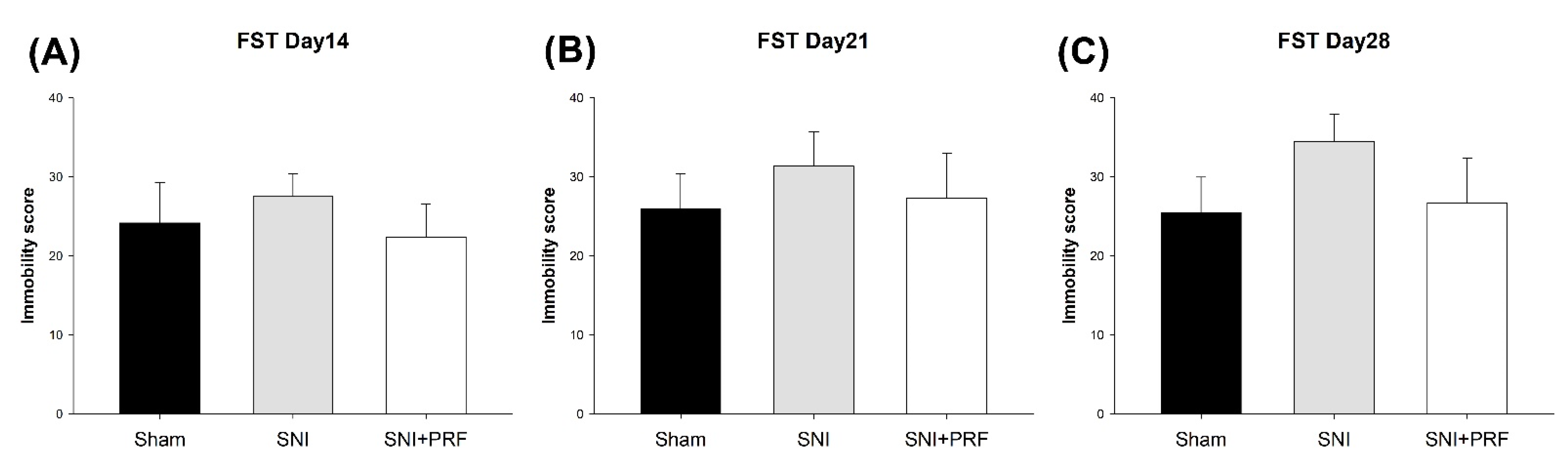
3.4. Depression-Like Behavior
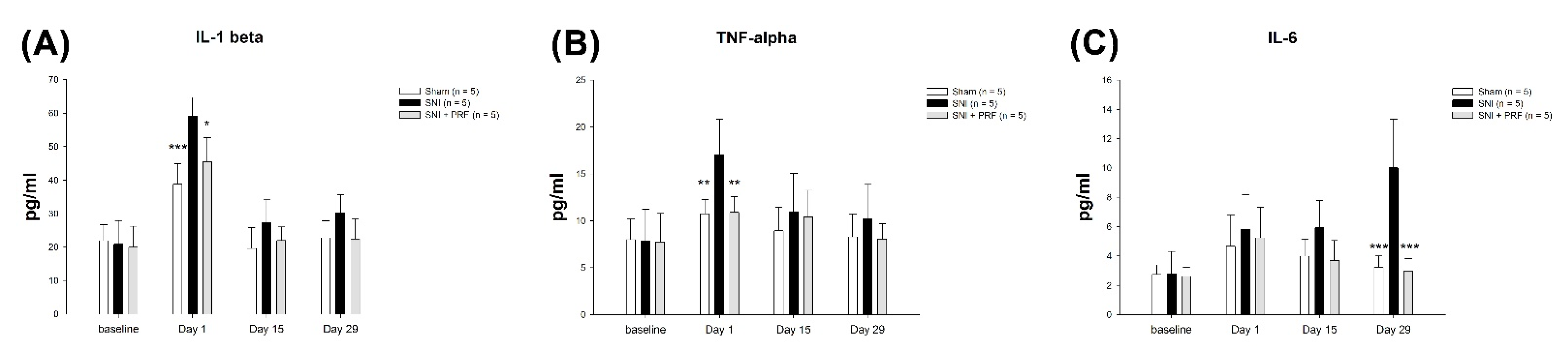
3.5. Proinflammatory Cytokines in Rats
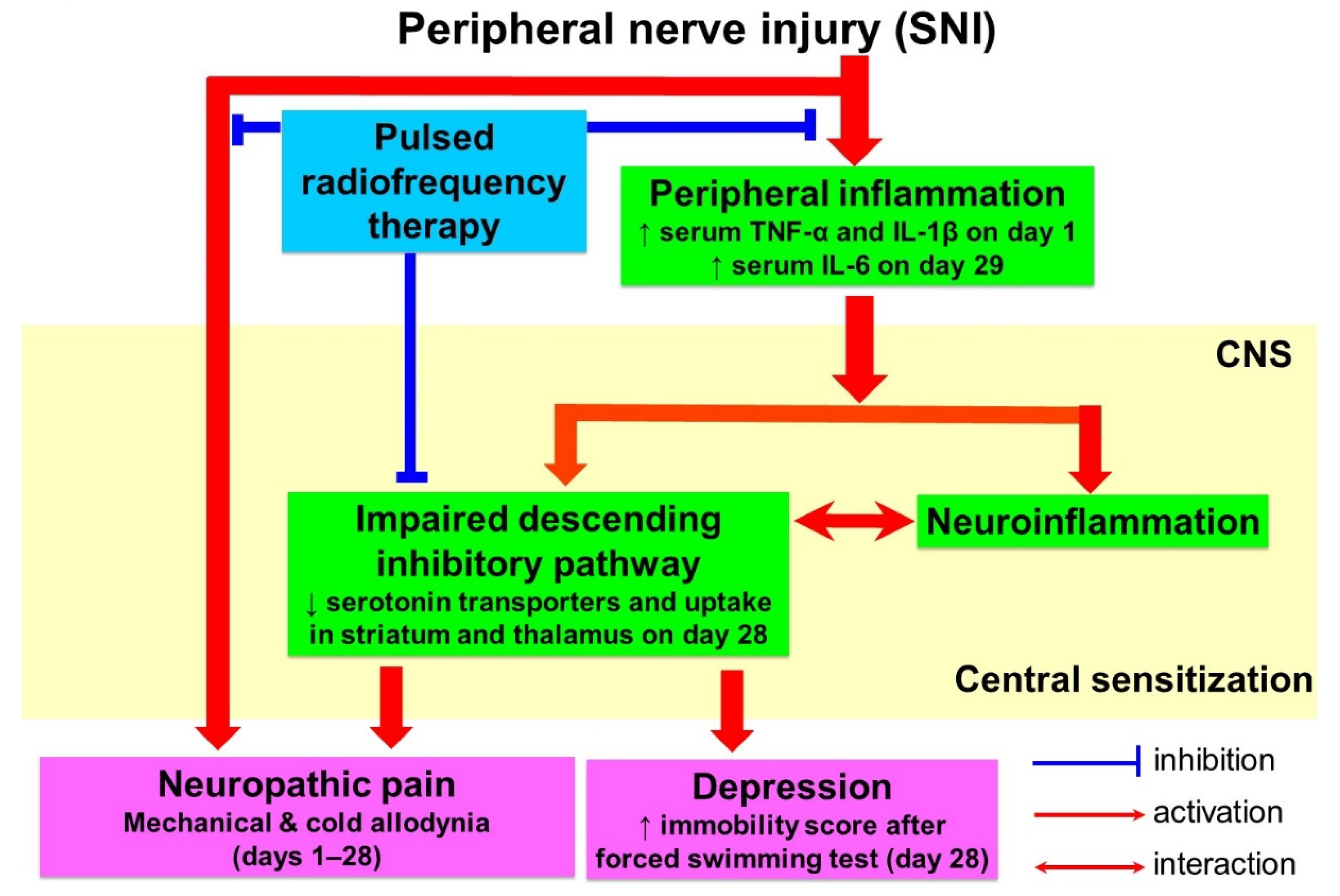
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Declaration of Transparency and Scientific Rigor
References
- Corallo, F.; De Salvo, S.; Floridia, D.; Bonanno, L.; Muscarà, N.; Cerra, F.; Cannistraci, C.; Di Cara, M.; Buono, V.L.; Bramanti, P. Assessment of spinal cord stimulation and radiofrequency: Chronic pain and psychological impact. Medicine 2020, 99, e18633. [Google Scholar] [CrossRef] [PubMed]
- Ji, R.R.; Xu, Z.Z.; Wang, X.; Lo, E.H. Matrix metalloprotease regulation of neuropathic pain. Trends Pharmacol. Sci. 2009, 30, 336–340. [Google Scholar] [CrossRef] [Green Version]
- Felger, J.C.; Lotrich, F.E. Inflammatory cytokines in depression: Neurobiological mechanisms and therapeutic implications. Neuroscience 2013, 246, 199–229. [Google Scholar] [CrossRef] [Green Version]
- Üçeyler, N.; Schäfers, M.; Sommer, C. Mode of action of cytokines on nociceptive neurons. Exp. Brain Res. 2009, 196, 67–78. [Google Scholar] [CrossRef]
- Moalem, G.; Tracey, D.J. Immune and inflammatory mechanisms in neuropathic pain. BrainRes. Rev. 2006, 51, 240–264. [Google Scholar] [CrossRef]
- Scholz, J.; Woolf, C.J. The neuropathic pain triad: Neurons, immune cells and glia. Nat. Neurosci. 2007, 10, 1361–1368. [Google Scholar] [CrossRef]
- Mann, J.J.; Huang, Y.Y.; Underwood, M.D.; Kassir, S.A.; Oppenheim, S.; Kelly, T.M.; Dwork, A.J.; Arango, V. A serotonin transporter gene promoter polymorphism (5-HTTLPR) and prefrontal cortical binding in major depression and suicide. Arch. Gen. Psychiatry 2000, 57, 729–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hains, B.C.; Everhart, A.W.; Fullwood, S.D.; Hulsebosch, C.E. Changes in serotonin, serotonin transporter expression and serotonin denervation supersensitivity: Involvement in chronic central pain after spinal hemisection in the rat. Exp. Neurol. 2002, 175, 347–362. [Google Scholar] [CrossRef] [PubMed]
- Yoon, C.; Wook, Y.Y.; Sik, N.H.; Ho, K.S.; Mo, C.J. Behavioral signs of ongoing pain and cold allodynia in a rat model of neuropathic pain. Pain 1994, 59, 369–376. [Google Scholar] [CrossRef]
- Weng, S.J.; Li, I.H.; Huang, Y.S.; Chueh, S.H.; Chou, T.K.; Huang, S.Y.; Shiue, C.Y.; Cheng, C.Y.; Ma, K.H. KA-bridged transplantation of mesencephalic tissue and olfactory ensheathing cells in a Parkinsonian rat model. J. Tissue Eng. Regen. Med. 2017, 11, 2024–2033. [Google Scholar] [CrossRef] [PubMed]
- Weng, S.J.; Shiue, C.Y.; Huang, W.S.; Cheng, C.Y.; Huang, S.Y.; Li, I.H.; Tao, C.C.; Chou, T.K.; Liao, M.H.; Chang, Y.P. PET imaging of serotonin transporters with 4-[18F]-ADAM in a Parkinsonian rat model. Cell Transplant. 2013, 22, 1295–1305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagiwara, S.; Iwasaka, H.; Takeshima, N.; Noguchi, T. Mechanisms of analgesic action of pulsed radiofrequency on adjuvant-induced pain in the rat: Roles of descending adrenergic and serotonergic systems. Eur. J. Pain 2009, 13, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Decosterd, I.; Woolf, C.J. Spared nerve injury: An animal model of persistent peripheral neuropathic pain. Pain 2000, 87, 149–158. [Google Scholar] [CrossRef]
- Kalmar, B.; Greensmith, L.; Malcangio, M.; McMahon, S.; Csermely, P.; Burnstock, G. The effect of treatment with BRX-220, a co-inducer of heat shock proteins, on sensory fibers of the rat following peripheral nerve injury. Exp. Neurol. 2003, 184, 636–647. [Google Scholar] [CrossRef]
- Lever, I.; Cunningham, J.; Grist, J.; Yip, P.K.; Malcangio, M. Release of BDNF and GABA in the dorsal horn of neuropathic rats. Eur. J. Neurosci. 2003, 18, 1169–1174. [Google Scholar] [CrossRef]
- Loeser, J.D.; Treede, R.D. The Kyoto protocol of IASP basic pain Terminology. Pain 2008, 137, 473–477. [Google Scholar] [CrossRef]
- Dowdall, T.; Robinson, I.; Meert, T.F. Comparison of five different rat models of peripheral nerve injury. Pharmacol. Biochem. Behav. 2005, 80, 93–108. [Google Scholar] [CrossRef]
- Yankelevitch-Yahav, R.; Franko, M.; Huly, A.; Doron, R. The forced swim test as a model of depressive-like behavior. J. Vis. Exp. JoVE 2015, 97, 52587. [Google Scholar] [CrossRef]
- Peng, C.J.; Huang, Y.Y.; Huang, W.S.; Shiue, C.Y. An automated synthesis of N, N-dimethyl-2-(2-amino-4-[18F] fluorophenylthio) benzylamine (4-[18F]-ADAM) for imaging serotonin transporters. Appl. Radiat. Isot. 2008, 66, 625–631. [Google Scholar] [CrossRef]
- Li, I.H.; Huang, W.S.; Shiue, C.Y.; Huang, Y.Y.; Liu, R.S.; Chyueh, S.C.; Hu, S.H.; Liao, M.H.; Shen, L.H.; Liu, J.C. Study on the neuroprotective effect of fluoxetine against MDMA-induced neurotoxicity on the serotonin transporter in rat brain using micro-PET. Neuroimage 2010, 49, 1259–1270. [Google Scholar] [CrossRef]
- Ma, K.H.; Huang, W.S.; Kuo, Y.Y.; Peng, C.J.; Liou, N.H.; Liu, R.S.; Hwang, J.J.; Liu, J.C.; Chen, H.J.; Shiue, C.Y. Validation of 4-[18F]-ADAM as a SERT imaging agent using micro-PET and autoradiography. Neuroimage 2009, 45, 687–693. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates: Hard Cover Edition; Elsevier: Amsterdam, The Netherlands, 2006. [Google Scholar]
- Wang, J.; Goffer, Y.; Xu, D.; Tukey, D.S.; Shamir, D.; Eberle, S.E.; Zou, A.H.; Blanck, T.J.; Ziff, E.B. A single subanesthetic dose of ketamine relieves depression-like behaviors induced by neuropathic pain in rats. J. Am. Soc. Anesthesiol. 2011, 115, 812–821. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humo, M.; Lu, H.; Yalcin, I. The molecular neurobiology of chronic pain–induced depression. Cell Tissue Res. 2019, 377, 21–43. [Google Scholar] [CrossRef] [PubMed]
- Norman, G.; Karelina, K.; Zhang, N.; Walton, J.; Morris, J.; Devries, A. Stress and IL-1β contribute to the development of depressive-like behavior following peripheral nerve injury. Mol. Psychiatry 2010, 15, 404–414. [Google Scholar] [CrossRef] [Green Version]
- Seminowicz, D.A.; Laferriere, A.L.; Millecamps, M.; Jon, S.; Coderre, T.J.; Bushnell, M.C. MRI structural brain changes associated with sensory and emotional function in a rat model of long-term neuropathic pain. Neuroimage 2009, 47, 1007–1014. [Google Scholar] [CrossRef] [Green Version]
- Fang, X.; Xu, X.; Lin, X.; Liu, R. Downregulated spinal IRF8 and BDNF in NAC are involved in neuropathic pain-induced depression relief via pulsed radiofrequency on dorsal root ganglion in rat SNI model. Brain Res. Bull. 2019, 146, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Rathbun, A.M.; Stuart, E.A.; Shardell, M.; Yau, M.S.; Baumgarten, M.; Hochberg, M.C. Dynamic effects of depressive symptoms on osteoarthritis knee pain. Arthritis Care Res. 2018, 70, 80–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doan, L.; Manders, T.; Wang, J. Neuroplasticity underlying the comorbidity of pain and depression. Neural Plast 2015, 2015, 504691. [Google Scholar] [CrossRef]
- Santana Pineda, M.M.; Vanlinthout, L.E.; Santana Ramirez, S.; vanZundert, J.; Novalbos Ruiz, J.P. A Randomized Controlled Trial to Compare Analgesia and Functional Improvement After Continuous Neuroablative and Pulsed Neuromodulative Radiofrequency Treatment of the Genicular Nerves in Patients with Knee Osteoarthritis up to One Year After the Intervention. Pain Med. 2021, 22, 637–652. [Google Scholar] [CrossRef]
- Zhang, G.F.; Wang, J.; Han, J.F.; Guo, J.; Xie, Z.M.; Pan, W.; Yang, J.J.; Sun, K.J. Acute single dose of ketamine relieves mechanical allodynia and consequent depression-like behaviors in a rat model. Neurosci. Lett. 2016, 631, 7–12. [Google Scholar] [CrossRef]
- Walker, A.K.; Kavelaars, A.; Heijnen, C.; Dantzer, R. Neuroinflammation and comorbidity of pain and depression. Pharmacol. Rev. 2014, 66, 80–101. [Google Scholar] [CrossRef] [Green Version]
- Selvaraj, S.; Murthy, N.V.; Bhagwagar, Z.; Bose, S.K.; Hinz, R.; Grasby, P.M.; Cowen, P.J. Diminished brain 5-HT transporter binding in major depression: A positron emission tomography study with [11 C] DASB. Psychopharmacology 2011, 213, 555–562. [Google Scholar] [CrossRef]
- Malison, R.T.; Price, L.H.; Berman, R.; Van Dyck, C.H.; Pelton, G.H.; Carpenter, L.; Sanacora, G.; Owens, M.J.; Nemeroff, C.B.; Rajeevan, N. Reduced brain serotonin transporter availability in major depression as measured by [123I]-2β-carbomethoxy-3β-(4-iodophenyl) tropane and single photon emission computed tomography. Biol. Psychiatry 1998, 44, 1090–1098. [Google Scholar] [CrossRef]
- Kong, E.; Sucic, S.; Monje, F.J.; Reisinger, S.N.; Savalli, G.; Diao, W.; Khan, D.; Ronovsky, M.; Cabatic, M.; Koban, F. STAT3 controls IL6-dependent regulation of serotonin transporter function and depression-like behavior. Sci. Rep. 2015, 5, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, K.D.; Taylor, K.S.; Anastakis, D.J. Nerve injury triggers changes in the brain. Neuroscientist 2011, 17, 407–422. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.L.; Lin, B.F.; Cherng, C.H.; Huh, B.K.; Ma, H.I.; Lin, S.L.; Wong, C.S.; Yeh, C.C. Pulsed radiofrequency therapy for relieving neuropathic bone pain in cancer patients. J. Med. Sci. 2014, 34, 84. [Google Scholar]
- Kim, K.; Jo, D.; Kim, E. Pulsed radiofrequency to the dorsal root ganglion in acute herpes zoster and postherpetic neuralgia. Pain Physician 2017, 20, E411–E418. [Google Scholar] [CrossRef]
- Van Boxem, K.; de Meij, N.; Kessels, A.; van Kleef, M.; Van Zundert, J. Pulsed radiofrequency for chronic intractable lumbosacral radicular pain: A six-month cohort study. Pain Med. 2015, 16, 1155–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaggi, A.S.; Singh, N. Role of different brain areas in peripheral nerve injury-induced neuropathic pain. Brain Res. 2011, 1381, 187–201. [Google Scholar] [CrossRef]
- Saade, N.; Al Amin, H.; Baki, S.A.; Chalouhi, S.; Jabbur, S.; Atweh, S. Reversible attenuation of neuropathic-like manifestations in rats by lesions or local blocks of the intralaminar or the medial thalamic nuclei. Exp. Neurol. 2007, 204, 205–219. [Google Scholar] [CrossRef]
- Jeanmonod, D.; Magnin, M.; Morel, A. Low–threshold calcium spike bursts in the human thalamus: Common physiopathology for sensory, motor and limbic positive symptoms. Brain 1996, 119, 363–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ansah, O.B.; Leite-Almeida, H.; Wei, H.; Pertovaara, A. Striatal dopamine D2 receptors attenuate neuropathic hypersensitivity in the rat. Exp. Neurol. 2007, 205, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Huo, B.B.; Shen, J.; Hua, X.Y.; Zheng, M.X.; Lu, Y.C.; Wu, J.J.; Shan, C.L.; Xu, J.G. Alteration of metabolic connectivity in a rat model of deafferentation pain: A 18F-FDG PET/CT study. J. Neurosurg. 2019, 132, 1295–1303. [Google Scholar] [CrossRef]
- Pandya, M.; Altinay, M.; Malone, D.A.; Anand, A. Where in the brain is depression? Curr. Psychiatry Rep. 2012, 14, 634–642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janicak, P.G.; Dokucu, M.E. Transcranial magnetic stimulation for the treatment of major depression. Neuropsychiatr. Dis. Treat. 2015, 11, 1549. [Google Scholar] [CrossRef] [PubMed]
- Dandekar, M.; Fenoy, A.; Carvalho, A.; Soares, J.; Quevedo, J. Deep brain stimulation for treatment-resistant depression: An integrative review of preclinical and clinical findings and translational implications. Mol. Psychiatry 2018, 23, 1094–1112. [Google Scholar] [CrossRef]
- van Zundert, J.; de Louw, A.J.A.; Joosten, E.A.J.; Kessels, A.G.H.; Honig, W.; Dederen, P.J.W.C.; Veening, J.; Vles, J.S.H.; van Kleef, M. Pulsed and continuous radiofrequency current adjacent to the cervical dorsal root ganglion of the rat induce late cellular activity in the dorsal horn. Anesthesiology 2005, 102, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chua, N.H.L.; Vissers, K.C.; Sluijter, M.E. Pulsed radiofrequency treatment in interventional pain management: Mechanisms and potential indications-A review. Acta Neurochir. Wien. 2011, 153, 763–771. [Google Scholar] [CrossRef] [Green Version]
- Sakai, K.; Hasegawa, C.; Okura, M.; Morikawa, O.; Ueyama, T.; Shirai, Y.; Sakai, N.; Saito, N. Novel variants of murine serotonin transporter mRNA and the promoter activity of its upstream site. Neurosci. Lett. 2003, 342, 175–178. [Google Scholar] [CrossRef]
- Lanquillon, S.; Krieg, J.C.; Bening-Abu-Shach, U.; Vedder, H. Cytokine production and treatment response in major depressive disorder. Neuropsychopharmacology 2000, 22, 370–379. [Google Scholar] [CrossRef]
- Miller, A.H. Five things to know about inflammation and depression. Psychiatr. Times 2018, 35, 2018. [Google Scholar]
- Gonçalves, L.; Silva, R.; Pinto-Ribeiro, F.; Pego, J.M.; Nuno Sousa, A.P.; Almeida, A. Neuropathic pain is associated withdepressivebehaviour and induces neuroplasticity in the amygdala of the rat. Exp. Neurol. 2008, 213, 48–56. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day after SNI | SUR (Mean ± Standard Deviation) | p-Value | |||||
|---|---|---|---|---|---|---|---|
| Thalamus | Sham vs. SNI | SNI vs. SNI+PRF | Normal vs. Sham | ||||
| Normal | Sham | SNI | SNI + PRF | ||||
| Day 14 | 2.83 ± 0.21 | 2.68 ± 0.57 | 2.44 ± 0.40 | 2.55 ± 0.27 | 0.701 | 0.960 | 0.915 |
| Day 28 | 2.81 ± 0.22 | 2.80 ± 0.59 | 1.67 ± 0.52 | 2.69 ± 0.44 | 0.002 * | 0.005 * | 1.000 |
| Day after SNI | SUR (Mean ± Standard Deviation) | p-Value | |||||
|---|---|---|---|---|---|---|---|
| Striatum | Sham vs. SNI | SNI vs. SNI+PRF | Normal vs. Sham | ||||
| Normal | Sham | SNI | SNI + PRF | ||||
| Day 14 | 2.73 ± 0.10 | 2.70 ± 0.70 | 2.46 ± 0.33 | 2.21 ± 0.57 | 0.832 | 0.802 | 1.000 |
| Day 28 | 2.78 ± 0.14 | 2.57 ± 0.24 | 1.73 ± 0.19 | 2.68 ± 0.74 | 0.010 * | 0.004 * | 0.802 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, K.-H.; Cheng, C.-Y.; Chan, W.-H.; Chen, S.-Y.; Kao, L.-T.; Sung, C.-S.; Hueng, D.-Y.; Yeh, C.-C. Pulsed Radiofrequency Upregulates Serotonin Transporters and Alleviates Neuropathic Pain-Induced Depression in a Spared Nerve Injury Rat Model. Biomedicines 2021, 9, 1489. https://doi.org/10.3390/biomedicines9101489
Ma K-H, Cheng C-Y, Chan W-H, Chen S-Y, Kao L-T, Sung C-S, Hueng D-Y, Yeh C-C. Pulsed Radiofrequency Upregulates Serotonin Transporters and Alleviates Neuropathic Pain-Induced Depression in a Spared Nerve Injury Rat Model. Biomedicines. 2021; 9(10):1489. https://doi.org/10.3390/biomedicines9101489
Chicago/Turabian StyleMa, Kuo-Hsing, Cheng-Yi Cheng, Wei-Hung Chan, Shih-Yu Chen, Li-Ting Kao, Chun-Sung Sung, Dueng-Yuan Hueng, and Chun-Chang Yeh. 2021. "Pulsed Radiofrequency Upregulates Serotonin Transporters and Alleviates Neuropathic Pain-Induced Depression in a Spared Nerve Injury Rat Model" Biomedicines 9, no. 10: 1489. https://doi.org/10.3390/biomedicines9101489
APA StyleMa, K. -H., Cheng, C. -Y., Chan, W. -H., Chen, S. -Y., Kao, L. -T., Sung, C. -S., Hueng, D. -Y., & Yeh, C. -C. (2021). Pulsed Radiofrequency Upregulates Serotonin Transporters and Alleviates Neuropathic Pain-Induced Depression in a Spared Nerve Injury Rat Model. Biomedicines, 9(10), 1489. https://doi.org/10.3390/biomedicines9101489