
MicroRNAs as Critical Biomarkers of Major Depressive Disorder: A Comprehensive Perspective

 ,
, 
 , ,
, ,  ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. A General Overview of Micro RNAs
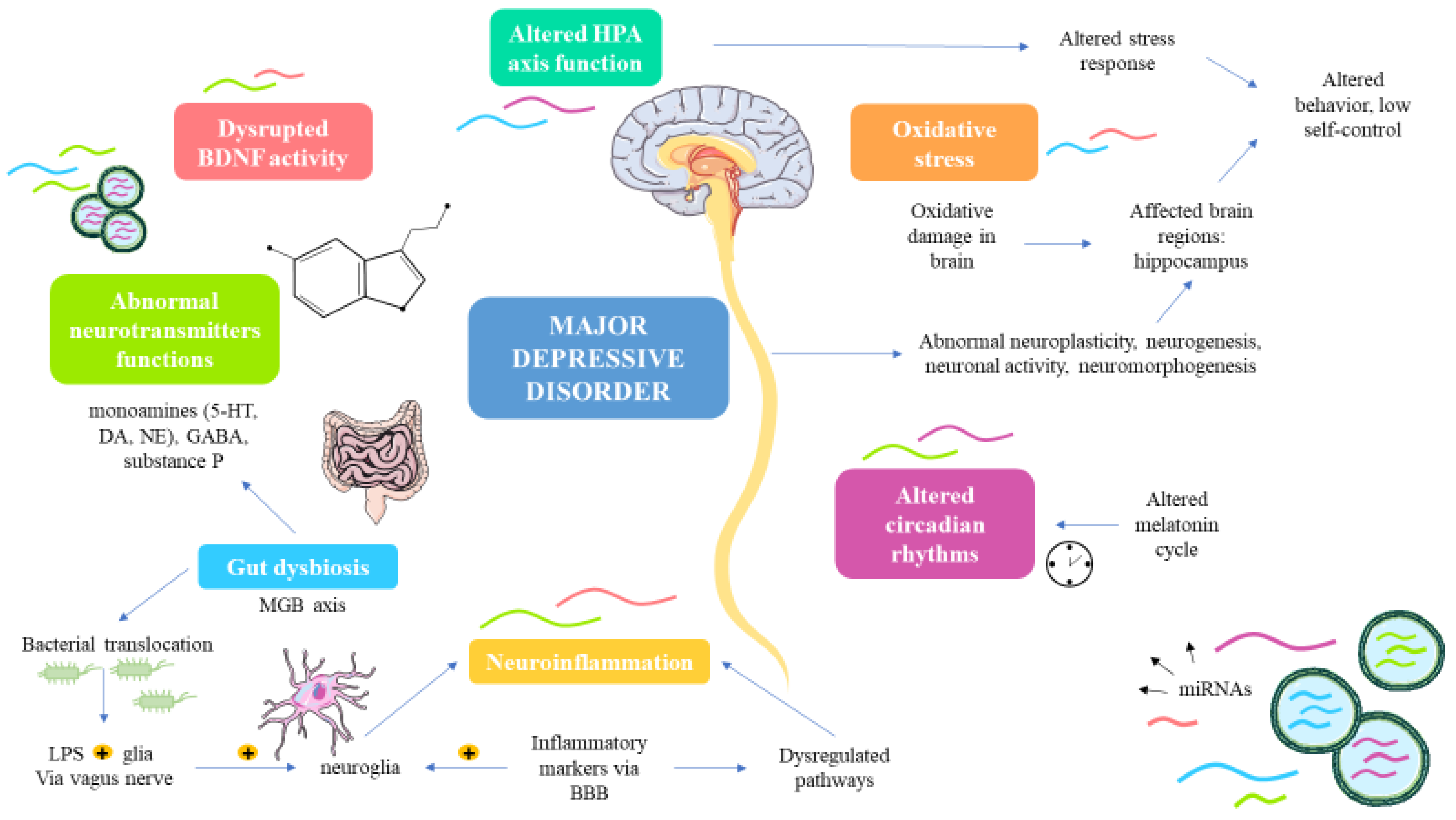
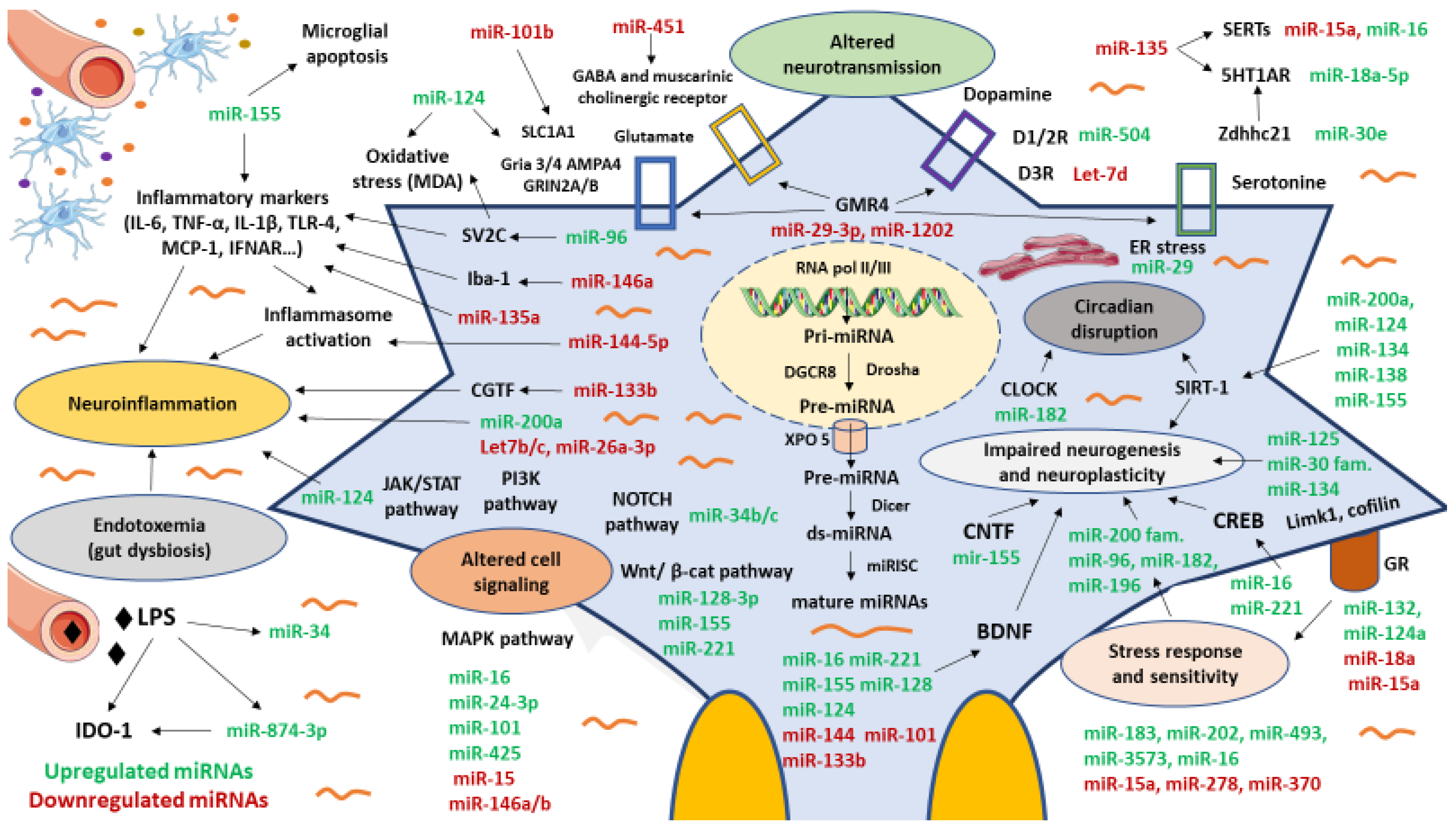
3. miRNAs Involved in the Pathogenesis of MDD
3.1. miRNAs Involved in Neurotransmitters and Neuropeptide Dysregulation
3.2. miRNAs Related to Stress and Structural, Functional and Molecular Changes in the Brain
3.3. miRNAs Associated to Neuroinflammation and MGB Axis
4. miRNAs with Diagnostic, Prognostic and Predictive Value
4.1. miRNAs with Diagnostic Value
4.2. miRNAs with Prognostic Value
4.3. miRNAs with Predictive Value
5. miRNAs as Promising Therapeutic Targets of MDD
6. Future Directions: miRNAs Modulation through Lifestyle Interventions
7. Conclusions

{kind=link}
{kind=link}
| miRNA | Upregulated/ Downregulated | Mechanism Involved/Effects | Translational Applications | References |
|---|---|---|---|---|
| Downregulated | Let-7d targets D3R in the hippocampus Let7b and let7c regulates PI3k-Akt-mTOR pathway IL-6 lead to a dysregulation of Let-7 family in depressive rat models | Diagnostic: rs10877887 and rs13293512 polymorphisms affecting Let-7 family are associated with increased MDD susceptibility Prognostic biomarker: Lower expression of Let-7 family correlates with higher degree of severity and treatment resistance Therapeutic: Let-7 family dysregulation could be ameliorated by epigenetic interventions such as physical activity Lentiviral-mediated Let-7d overexpression is associated with anxiolytic and anti-depressant-like action | [75,105,133,177,207,238] |
| Upregulated | Increased levels of miR-9 affect intrinsic amygdala functional connectivity, related to depressive severity and childhood maltreatment | Prognostic: To classify MDD severity Therapeutic: Animal models show: an inhibitor may block neuronal apoptosis activating Notch 1 signaling pathway, which is key for neural development and brain homeostasis (neuronal connectivity, synaptic plasticity and learning/memory). Notch signaling pathway is crucial in early neurodevelopment and late-life neurodegeneration. | [208,239,240,241,262] |
| Downregulated in the amygdala | Post-transcriptional regulation of SERT with miR-16. Reduced levels of amygdalar miR-15 but elevated circulating levels in peripheral blood increase anxiety-like behaviors. | Predictive: significant up-regulation at 3–6 h post-treatment with dexamethasone | [65,111,225] |
| Upregulated | Post-transcriptional regulation of SERT with miR-15a Depression induced by maternal deprivation but not CUPS was significantly associated with miR-16 upregulation, probably targeting BDNF in the hippocampus MiR-16 regulate apoptosis and autophagy and could account for some part of the therapeutic effect of SSRIs | Diagnostic: CSF levels of miR-16 is downregulated in patients with MDD Predictive: By reducing miR-16, serotonergic functions in noradrenergic neurons improve when combined with antidepressants Therapeutic: Although inconsistencies within different animal models cannot conclude if miR-16 is involved in affective disorders, it is clear that it interferes with antidepressant treatments, antagonizing them. | [65,66,98,110,190,222] |
| Downregulated | Implied in resilience to stress, miR-18a-5p acts as a negative epigenetic regulator of 5-HT1AR, altering the serotonergic homeostatic balance and functioning of the hippocampus. The upregulation of miR-18a not only in hippocampus, but also in prefrontal cortex, is related to downregulation of GRs and depressive-like behaviors | Therapeutic: Possible new antidepressant strategies aiming to restore this biological processes. | [118,119] |
| Upregulated | miR-22 might be involved in cerebral microvascular impairment associated to depression | Diagnostic: miR-22 levels are potential indicators of post-stroke depression | [180] |
| Upregulated | MiR-24 targets oxytocin, which may be related to MDD etiopathogenesis MiR-24-3p is associated with the regulation of Wnt and MAPK signaling pathways, | Diagnostic: Elevated miR-24 serum levels is an indicator of MDD Predictive: Serum levels of miR-24 is a robust blood marker of antidepressants response | [93,94,136,263] |
| Downregulated | MiR-26a-3p is a critical regulator of PTEN/PI3K/Akt pathway, regulating neuronal autophagy, synaptic plasticity, and survival in the dentate gyrus of a rat model of depression | Diagnostic: serum miR-26 is altered in depressed mice Predictive: miR-26a-2 levels are significantly upregulated in the dorsal raphe nucleus following antidepressant therapy and mir-26a KO shows poorer antidepressant response. Therapeutic: miR-26a-2 functions as an endogenous antidepressant by targeting HTR1A in serotonergic neurons. | [134,178,179] |
| Downregulated | MiR-29 is essential for neuronal survival in the brain, targeting VDAC1 MiR-29 is highly produced by astrocytes and their dysregulation in MDD may be an indicator of abnormal functioning of these cellsMiR-29b-3p is critical for inhibiting GRM4 expression in the prefrontal cortex of depressive-like rats MiR-29 is a major regulator of endoplasmic reticulum stress in neurons, a crucial pathophysiological event in the neurons related to MDD | Predictive: Overexpression of miR-29 acts as a critical mediator of ketamine’s antidepressant effect in depressive rats | [55,79,101,102,264] |
| Upregulated | miR-30 family miRNAs mediate chronic stress-induced depression-like phenotype by altering hippocampal neurogenesis, neuroplasticity, epigenetic and transcriptional regulators such as Mll3 and Runx1 Socs3, Ppp3r1, Gpr125, and Nrp1. MiR-30a regulates neural nutrient signaling pathway, axon guidance, insulin and other signaling pathways The polymorphism miR-30e ss178077483 is associated with P300 latency and the individuals with the C/T genotype have a longer P300 latency than those carrying the C/C genotype | Diagnostic: polymorphic miR-30e variant ss178077483 appears to increase MDD susceptibility miR-30a-5p is a diagnostic marker of PSD miR-30d is associated with late life depression Prognostic: Elevated levels of miR-30e was detected in post-mortem brain samples of patients with MDD died by suicide Therapeutic: The use of lurasidone during adolescence was able to prevent the up-regulation of miR-30a and normalized the expression of its target genes in response to prenatal stress exposure. | [69,113,181,182,183,184] |
| Upregulated | LPS and stress, both related to MDD induce the expression of miR-34a MiR-34a targets synaptotagmin-1 and Bcl-2 associated with neuronal spine damage MiR-34a induce depressive-like behavior and impact in the serotoninergic activity in the raphe nuclei of mice Patients with MDD show a significant decrease in NOTCH signaling components, inversely related to miR-34b/c expression | Diagnostic: rs4938723/rs28757623 polymorphisms affecting miR-34b/c genes are potential risk factors for suffering from MDD MiR-34a is differentially expressed in MDD, bipolar disorder and schizophrenia in the anterior cingulate cortex. Prognostic: There is a significant association between rs4938723 and negative life events in relation to MDD risk. The polymorphism rs2187473 affecting miR-34c is a potential genetic risk factor for cognitive decline in MDD Peripheral blood leukocytes levels of miR-34b and miR-34c were related to suicide idea and cognitive function Therapeutic: antagomiR-34a activated TrkB/MEK1/ERK signaling and improved spine morphology in the hippocampus., exerting antidepressant effects Predictive: Responder patients show decreased serum miR-34a-5p, a miRNA directly correlated with HAM-D score | [64,168,170,171,214,265] |
| Upregulated | miR-96 targets SV2C in the CA1 area of the hippocampus, leading to a depressive-like behavior and memory impairment | Therapeutic The use of miR-96 antagonists led to a reduced pro-inflammatory and pathogenic markers related with MDD | [141] |
| Downregulated | CUMS rats present reduced miR-101 in the ventrolateral orbital cortex with increased dual specificity phosphatase 1 (DUSP1) expression and reduced ERK-1 and BDNF Flinders sensitive line rats presented decreased miR-101b expression in the prefrontal cortex targeting the neuronal glutamate transporter SLC1A1. | Diagnostic: miR-101 in combination with miR-93 are potential reference biomarkers of MDD Therapeutic: miR-101 mimics reversed depressive-like behaviors in CUMS rats | [80,120,204] |
| Upregulated | miR-124-3p is a master regulator of multiple processes targeting SOX-9, JAG-Notch, CREB-1, SIRT-1 BDNF, Ezh2, SCP1, GR and neuroinflammatory components. | Diagnosis: miR-124-3p is upregulated in patients with MDD Predictive: miR-124 reduced levels in plasma could be an indicator of favorable antidepressant response Therapeutic: In vivo inhibition of miR-124 conducted in different animal models and brain regions show some promising results | [81,89,103,104,123,145,266,267] |
| upregulated | MiR-128-3p is involved in Wnt downregulation in the amygdala of learned helpless rats and patients with MDD Patients with type 2 diabetes mellitus and depression show increased levels of miR-128, cortisol while reduced BDNF and shortened telomeres | Therapy: Potential posttranscriptional switching mechanism in the amygdala targeting miR-128 may benefit Wnt restoring | [125,138] |
| Upregulated | MeCP2, regulation of BDNF hippocampal levels Higher miR-132 levels are associated with both lower fractional amplitude of low frequency fluctuations and lower grey matter volume in fronto-limbic network; as well as poorer cognitive performance in attention and executive function Currently considered a neurimiRs | Diagnosis: There is a direct relationship between miR-132 expression levels and self-rating depression scale and HAM-D scale Prognostic: miR-132 may play a role in the coexistence of cardiovascular disease and MDD. Increased miR-132 expression levels were associated with visual memory deficits, correlated with anxiety symptoms Therapeutic: miR-132 inhibition increased BNDF levels in vivo | [126,209,210,242,243,266,268] |
| Downregulated | CGTF suppression | Predictive: miR-133b increase in the hippocampus of mice after fluoxetine treatment Therapeutic: miR-133b augmentation was associated with decreased apoptosis, repressed inflammatory reaction, and increased expression of GFAP, BDNF and neurotransmitters in hippocampal tissues of depression rats | [122,228] |
| Upregulated, | UCMS exposure significantly increased the expression of miR-134 within the ventromedial prefrontal cortex, leading to a decrease in Limk1 and cofilin. | Diagnostic: miR-134 is downregulated in patients with MDD in comparison to healthy controls (79% sensitivity and 84% specificity), bipolar and schizophrenic patients (79% sensitivity and 76.5 specificity). Therapeutic: Blocking miR-134 in vivo ameliorated neuronal structural abnormalities, biochemical changes and depression-like behaviors. Inhibition of miR-134-5p together with the use of a GR antagonist and SIRT-1 agonist depression susceptibility induced by prenatal dexamethasone exposure (PDE) in offspring rats could be prevented | [90,114,191] |
| Downregulated | MiR-135 exert anti-inflammatory actions in the brain targeting multiple inflammatory mediators in the hippocampus (IL-1β, IL-6 TNF-α and TLR-4) MiR-135 is critical for serotonin regulation in the raphe nuclei | Diagnostic: Downregulated in patients with MDD Predictive: circulating miR-135 levels could be an indicator of favorable response to antidepressants. Therapeutic: Combined use of miR-135a mimic plus antidepressants (fluoxetine) induced a significant decrease of pro-inflammatory markers in CUMS mice | [71,72,146,203,269] |
| Probably downregulated | miR-137 levels are significantly diminished in the brain in post-stroke depression rats, miR-137 is related to anxiety behaviors in mice | Prognostic: miR-137 levels are substantially down-regulated by 25% in the postmortem prefrontal cortex of depressed patients with suicidal behavior Potential diagnostic, therapeutic and prognostic factor still unexplored | [198,270,271] |
| Upregulated | MiR-138 regulate depression-like behavior by targeting by SIRT1 expression in the hippocampus. | Not studied yet | [91] |
| Downregulated | Low MiR-144-5p is associated with multiple neuroinflammatory proteins MiR-144 targets PTP1B activating the TrkB/BDNF signaling in the hippocampus | Diagnostic: Circulating miR-144-5p levels are inversely correlated with depressive symptoms and Montgomery-Åsberg Depression Rating Scale (MADRS-S) Therapeutic: Lentiviral injection of miR-144 exerted antidepressant roles in the hippocampus of chronic unpredictable mild stress rats. Mood stabilizers (lithium and sodium valproate) increases miR-144 levels in animal models. | [121,148,199,247] |
| Downregulated | MiR-146a reduces neuroinflammation and depressive behavior in mice models through targeting Iba-1, iNOS, IL-1β, TNF-α, interleukin 1 receptor associated kinase 1 (IRAK1), TNF receptor-associated factor 6 (TRAF6) and phosphorylated NF-κB p65 MiR-146 is related to Wnt signaling, Cancer, Endocytosis, Axon guidance and MAPK signaling | Diagnostic: miR-146a expression before treatment is inversely correlated with HAM-D score Therapeutic: miR-146a mimic treatment inhibited TNF-α, IL-1β, IRAK1 and TRAF6 expression in BV-2 cells Predictive: miR-146a may be a promising marker to predict fluoroxetine and antidepressant response in patients with MDD. | [136,149,200,207,227] |
| Upregulated | miR-155 inhibits MyD88 gene alleviating depressive-like behaviors miR-155 is involved in neuroinflammatory pathways (IL-6 and TNF-α raise) and ciliary neurotrophic factor expression. MiR-155 is associated with increased apoptosis in the hippocampus by targeting Wnt/β-catenin signalling and microglia | Therapeutic: miR-155 deletion reduces anxiety and depressive-like behavior in vivo; Citalopram and other antidepressants appears to downregulate miR-155 expression miR-155 has been proposed as an interesting approach in treatment-resistant depression. Diagnostic: Cellular and CSF levels of miR-155 are upregulated in patients with MDD However there is still controversy regarding serum levels of miR-155 miR-155 before treatment was directly correlated with severity of depression. | [92,127,137,142,143,207,272] |
| Probably upregulated | MiR-181a appears to be associated with stress-induced upregulation of the noradrenergic phenotype, observed in depressive patients, also regulating GRs; PI3K/Akt/mTOR pathway | Diagnostic: miR-181a together with 32 additional miRNAs were altered in patients with MDD in comparison to controls. | [201,202] |
| Upregulated | Abnormal processing of miR-182 in the dentate gyrus of the hippocampus in individuals carrying appears to contribute to the dysregulation of circadian rhythms in MDD patients with insomnia targeting CLOCK gene miR-182 is inversely correlated with BDNF levels | Diagnostic: T allele of the rs76481776 polymorphism may be related to clinic insomnia; miR-182 is inversely correlated with self-rating depression score Therapeutic: miR-182 inhibition in the hippocampus may exert antidepressant effects in vivo. | [84,126,244,273] |
| Upregulated | Inhibition of BDNF activity in the brain. MiR-183 is upregulated in the hippocampus of chronic unpredictable mild stressed rats. | Predictive: miR-183 is increased after 4 weeks of antidepressant (escitalopram) therapy | [115,229] |
| Downregulated | MiR-184 targets PDE4B NCOR2 in the anterior cingulate cortex of depressed patients KO models of flies demonstrate that reduced levels of miR-184 is associated with a depressive behavior, more prominent in older flies | Diagnostic: MiR-184 is differentially expressed in MDD patients in comparison to bipolar or either bipolar and depressed patients | [192,274] |
| Upregulated | Upregulated in the hippocampus of rodents affected by maternal separation and inescapable shock targeting Zeb1 and Zeb2 MiR-200a appears to be involved in altered neurogenesis, inflammatory activation, lipid metabolism, disturbed circadian rhythm, and insulin secretion in the co-existence of pain and depression | Diagnostic: Depressed patients present reduced levels of blood miR-200a, miR-200b and miR-200c in comparison to healthy controls. | [106,193,275] |
| Upregulated | Regulation of a plethora of neuronal signaling pathways and morphogenesis Both acute and chronic stress exposure lead to miR-212 augmentation in the hippocampus and the amygdala, related to anxiety behaviors | Predictive: Electroconvulsive stimulation raises miR-212 levels either in the blood or dentate gyrus both after acute and chronic administration MiR-212 is also proposed as a promising predictor of serotonin reuptake inhibitors | [220,223,224,243] |
| Downregulated | miR-218 acts as a molecular switch that may determine susceptibility vs. resilience to chronic stress acting through corticosterone-related networks | Diagnostic: Plasma levels of miR-218 appears to be diminished in patients with MDD | [116,196,276] |
| Upregulated | MiR-221 is positively associated with MDD pathogenesis in rats with chronic unpredictable mild stress by targeting Wnt2/CREB/BDNF pathway | Diagnostic: Serum and cerebrospinal fluid of depressed patients show increased levels of miR-221; elevated serum levels of miR-221-3p can be used as an indicator for depressed mood in perioperative patients; Increased serum and CSF levels is a major risk factor of PSD, directly correlated with HAM-D score, IL-6 and TNF-α levels. Predictive: Paroxetine may lead to decreased serum levels of miR-221-3p, which is related to decreased HAM-D score Therapy: ERK pathway regulates miR-221 expression | [124,187,194,195,246,277] |
| Downregulated | MiR-323 appears to be diminished in depression animal models, but upregulated in the brain of newborns after prenatal stress Targets and affected regions: Erb-b2 receptor tyrosine kinase 4 (ERBB4), neuregulin pathway, hippocampus, anterior cingulate cortex and habenula | Therapeutic: In vivo knock-down and upregulation of miR-323 was associated with increased anxiety and augmented emotionality. | [245,278,279] |
| Upregulated | A shift in the expression of miR-376c-3p, miR-455-3p and miR-337-3p in the synaptic fraction over total fraction was found in MDD subjects compared with healthy controls. MiR-376b-5p probably targets a number of genes relevant to stress signaling pathways and neuronal regulation, being downregulated after augmented maternal care (Kcnh5 ↓, Sh3rf2↓, Acox2↓, Otx2↓, Myof↓, Frrs1↓, Dio2↓, Acss3) miR-376b and miR-208 increased whereas miR-9-1 decreased under acute and chronic stress conditions, | Not studied yet. | [97,105,117,280] |
| Upregulated | miR-429 appears to be upregulated after acute and repetitive stress exposure miR-429 is upregulated in the fibroblasts of depressed patients, affecting to the expression of 842 genes in which 9 are known to be involved in MDD pathogenesis. | Not studied yet. | [85,281,282,283] |
| Downregulated | Maternal deprivation induced downregulation of miR-451 in the hippocampus. MiR-451 appears to regulate some critical genes associated to MDD including CREB pathway, GABAergic and cholinergic neurotransmission | Diagnostic: Prior works have denoted either increased or decreased serum levels of miR-451 in depressed patients Prognostic: There is an inverse relationship between miR-451 levels and HAMD score Predictive: A positive relationship between paroxetine and fluoexetine effectiveness and miR-451 was found, although the effect of ketamine in miR-451 is still controversial | [83,133,187,194,230] |
| Downregulated | miR-494 is downregulated in the dorsolateral prefrontal cortex depressed suicide patients | Diagnostic: miR-494 is augmented in the peripheral blood of depressed patients in comparison to healthy controls. | [197,198,276] |
| Upregulated | miR-504 inhibits dopamine D1 and D2 receptor gene (DRD1/2) miR-504 is increased in the nucleus accumbens of animals with maternal deprivation, leading to chronic unpredictable stress and higher sensitivity to stress in the adulthood | Diagnostic: rs686 polymorphism affecting DRD1 miR-504 regulation is associated with higher depression scales | [76,77,189] |
| Upregulated | MiR-874-3p influence IDO1 activity and its effect on lipopolysaccharide (LPS)-induced depression-like behavior in mice | Not studied yet. | [155] |
| Downregulation | Regulation of glutamatergic, dopaminergic, GABAergic, and serotonergic neurotransmission | Diagnosis: miR-1202 is reduced in patients with MDD Predictive: miR-1202 predicts citalopram treatment response Responder patients displayed lower baseline miR-1202 levels than nonresponders. Prognosis: Probable association with increased risk of suicide. | [78,82,203,226] |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chand, S.P.; Arif, H. Depression; StatPearls: Treasure Island, FL, USA, 2021. [Google Scholar]
- Gutiérrez-Rojas, L.; Porras-Segovia, A.; Dunne, H.; Andrade-González, N.; Cervilla, J.A. Prevalence and Correlates of Major Depressive Disorder: A Systematic Review. Rev. Bras. Psiquiatr. 2020, 42, 657–672. [Google Scholar] [CrossRef]
- Kessler, R.C.; Bromet, E.J. The Epidemiology of Depression across Cultures. Annu. Rev. Public Health 2013, 34, 119. [Google Scholar] [CrossRef] [Green Version]
- Greenberg, P.E.; Fournier, A.A.; Sisitsky, T.; Pike, C.T.; Kessler, R.C. The Economic Burden of Adults with Major Depressive Disorder in the United States (2005 and 2010). J. Clin. Psychiatry 2015, 76, 155–162. [Google Scholar] [CrossRef] [Green Version]
- Lépine, J.P.; Briley, M. The Increasing Burden of Depression. Neuropsychiatr. Dis. Treat. 2011, 7, 3–7. [Google Scholar] [CrossRef] [Green Version]
- König, H.; König, H.H.; Konnopka, A. The Excess Costs of Depression: A Systematic Review and Meta-Analysis. Epidemiol. Psychiatr. Sci. 2019, 29, e30. [Google Scholar] [CrossRef] [Green Version]
- Orsolini, L.; Latini, R.; Pompili, M.; Serafini, G.; Volpe, U.; Vellante, F.; Fornaro, M.; Valchera, A.; Tomasetti, C.; Fraticelli, S.; et al. Understanding the Complex of Suicide in Depression: From Research to Clinics. Psychiatry Investig. 2020, 17, 207. [Google Scholar] [CrossRef] [Green Version]
- Sheline, Y.I.; Liston, C.; McEwen, B.S. Parsing the Hippocampus in Depression: Chronic Stress, Hippocampal Volume, and Major Depressive Disorder. Biol. Psychiatry 2019, 85, 436–438. [Google Scholar] [CrossRef]
- Raison, C.L.; Miller, A.H. The Evolutionary Significance of Depression in Pathogen Host Defense (PATHOS-D). Mol. Psychiatry 2013, 18, 15–37. [Google Scholar] [CrossRef]
- Inserra, A.; Rogers, G.B.; Licinio, J.; Wong, M.L. The Microbiota-Inflammasome Hypothesis of Major Depression. BioEssays: News Rev. Mol. Cell. Dev. Biol. 2018, 40, 1800027. [Google Scholar] [CrossRef] [Green Version]
- Troubat, R.; Barone, P.; Leman, S.; Desmidt, T.; Cressant, A.; Atanasova, B.; Brizard, B.; El Hage, W.; Surget, A.; Belzung, C.; et al. Neuroinflammation and Depression: A Review. Eur. J. Neurosci. 2021, 53, 151–171. [Google Scholar] [CrossRef]
- Gautam, S.; Jain, A.; Gautam, M.; Vahia, V.N.; Grover, S. Clinical Practice Guidelines for the Management of Depression. Indian J. Psychiatry 2017, 59, S34. [Google Scholar] [CrossRef]
- Alvarez-Mon, M.A.; Ortega, M.A.; García-Montero, C.; Fraile-Martinez, O.; Monserrat, J.; Lahera, G.; Mora, F.; Rodriguez-Quiroga, A.; Fernandez-Rojo, S.; Quintero, J.; et al. Exploring the Role of Nutraceuticals in Major Depressive Disorder (MDD): Rationale, State of the Art and Future Prospects. Pharmaceuticals 2021, 14, 821. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Li, X.H.; Zheng, W.; Xu, Z.Y.; Ng, C.H.; Ungvari, G.S.; Yuan, Z.; Xiang, Y.T. Mindfulness-Based Interventions for Major Depressive Disorder: A Comprehensive Meta-Analysis of Randomized Controlled Trials. J. Affect. Disord. 2018, 229, 429–436. [Google Scholar] [CrossRef]
- Murri, M.B.; Ekkekakis, P.; Magagnoli, M.; Zampogna, D.; Cattedra, S.; Capobianco, L.; Serafini, G.; Calcagno, P.; Zanetidou, S.; Amore, M. Physical Exercise in Major Depression: Reducing the Mortality Gap While Improving Clinical Outcomes. Front. Psychiatry 2018, 9, 762. [Google Scholar] [CrossRef] [Green Version]
- Gee, B.; Orchard, F.; Clarke, E.; Joy, A.; Clarke, T.; Reynolds, S. The Effect of Non-Pharmacological Sleep Interventions on Depression Symptoms: A Meta-Analysis of Randomised Controlled Trials. Sleep Med. Rev. 2019, 43, 118–128. [Google Scholar] [CrossRef]
- Sato, S.; Yeh, T.L. Challenges in Treating Patients with Major Depressive Disorder: The Impact of Biological and Social Factors. CNS Drugs 2013, 27 (Suppl. 1), 5–10. [Google Scholar] [CrossRef] [Green Version]
- Nemeroff, C.B. The Burden of Severe Depression: A Review of Diagnostic Challenges and Treatment Alternatives. J. Psychiatr. Res. 2007, 41, 189–206. [Google Scholar] [CrossRef]
- Kraus, C.; Kadriu, B.; Lanzenberger, R.; Zarate, C.A., Jr.; Kasper, S. Prognosis and Improved Outcomes in Major Depression: A Review. Focus (Am. Psychiatr. Publ.) 2020, 18, 220–235. [Google Scholar] [CrossRef]
- Subramanian, S.; Steer, C.J. Special Issue: MicroRNA Regulation in Health and Disease. Genes 2019, 10, 457. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Lu, Q.; Chang, C. Epigenetics in Health and Disease. Adv. Exp. Med. Biol. 2020, 1253, 3–55. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.X.; Rothenberg, M.E. MicroRNA. J. Allergy Clin. Immunol. 2018, 141, 1202. [Google Scholar] [CrossRef] [Green Version]
- Hendrickson, D.G.; Hogan, D.J.; McCullough, H.L.; Myers, J.W.; Herschlag, D.; Ferrell, J.E.; Brown, P.O. Concordant Regulation of Translation and MRNA Abundance for Hundreds of Targets of a Human MicroRNA. PLoS Biol. 2009, 7, e100238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jonas, S.; Izaurralde, E. Towards a Molecular Understanding of MicroRNA-Mediated Gene Silencing. Nat. Rev. Genet. 2015, 16, 421–433. [Google Scholar] [CrossRef]
- Ortega, M.A.; Fraile-Martínez, O.; Guijarro, L.G.; Casanova, C.; Coca, S.; Álvarez-Mon, M.; Buján, J.; García-Honduvilla, N.; Asúnsolo, Á. The Regulatory Role of Mitochondrial MicroRNAs (MitomiRs) in Breast Cancer: Translational Implications Present and Future. Cancers 2020, 12, 2443. [Google Scholar] [CrossRef] [PubMed]
- Borchert, G.M.; Lanier, W.; Davidson, B.L. RNA Polymerase III Transcribes Human MicroRNAs. Nat. Struct. Mol. Biol. 2006, 13, 1097–1101. [Google Scholar] [CrossRef]
- Lee, Y.; Kim, M.; Han, J.; Yeom, K.H.; Lee, S.; Baek, S.H.; Kim, V.N. MicroRNA Genes Are Transcribed by RNA Polymerase II. EMBO J. 2004, 23, 4051–4060. [Google Scholar] [CrossRef]
- Gregory, R.I.; Yan, K.P.; Amuthan, G.; Chendrimada, T.; Doratotaj, B.; Cooch, N.; Shiekhattar, R. The Microprocessor Complex Mediates the Genesis of MicroRNAs. Nature 2004, 432, 235–240. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, J.; Hayder, H.; Zayed, Y.; Peng, C. Overview of MicroRNA Biogenesis, Mechanisms of Actions, and Circulation. Front. Endocrinol. 2018, 9, 402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukla, G.C.; Singh, J.; Barik, S. MicroRNAs: Processing, Maturation, Target Recognition and Regulatory Functions. Mol. Cell. Pharmacol. 2011, 3, 83. [Google Scholar]
- Ha, M.; Kim, V.N. Regulation of MicroRNA Biogenesis. Nat. Reviews. Mol. Cell Biol. 2014, 15, 509–524. [Google Scholar] [CrossRef]
- Abdelfattah, A.M.; Park, C.; Choi, M.Y. Update on Non-Canonical MicroRNAs. Biomol. Concepts 2014, 5, 275. [Google Scholar] [CrossRef] [Green Version]
- Stavast, C.J.; Erkeland, S.J. The Non-Canonical Aspects of MicroRNAs: Many Roads to Gene Regulation. Cells 2019, 8, 1465. [Google Scholar] [CrossRef] [Green Version]
- Kosik, K.S. MicroRNAs and Cellular Phenotypy. Cell 2010, 143, 21–26. [Google Scholar] [CrossRef] [Green Version]
- Turchinovich, A.; Samatov, T.R.; Tonevitsky, A.G.; Burwinkel, B. Circulating MiRNAs: Cell–Cell Communication Function? Front. Genet. 2013, 4, 119. [Google Scholar] [CrossRef] [Green Version]
- Weber, J.A.; Baxter, D.H.; Zhang, S.; Huang, D.Y.; Huang, K.H.; Lee, M.J.; Galas, D.J.; Wang, K. The MicroRNA Spectrum in 12 Body Fluids. Clin. Chem. 2010, 56, 1733. [Google Scholar] [CrossRef]
- Turchinovich, A.; Weiz, L.; Burwinkel, B. Extracellular MiRNAs: The Mystery of Their Origin and Function. Trends Biochem. Sci. 2012, 37, 460–465. [Google Scholar] [CrossRef]
- Brown, R.A.M.; Epis, M.R.; Horsham, J.L.; Kabir, T.D.; Richardson, K.L.; Leedman, P.J. Total RNA Extraction from Tissues for MicroRNA and Target Gene Expression Analysis: Not All Kits Are Created Equal. BMC Biotechnol. 2018, 18, 16. [Google Scholar] [CrossRef] [Green Version]
- Desvignes, T.; Batzel, P.; Berezikov, E.; Eilbeck, K.; Eppig, J.T.; McAndrews, M.S.; Singer, A.; Postlethwait, J.H. MicroRNA Nomenclature: A View Incorporating Genetic Origins, Biosynthetic Pathways, and Sequence Variants. Trends Genet. TIG 2015, 31, 613. [Google Scholar] [CrossRef] [Green Version]
- Budak, H.; Bulut, R.; Kantar, M.; Alptekin, B. MicroRNA Nomenclature and the Need for a Revised Naming Prescription. Brief. Funct. Genom. 2016, 15, 65–71. [Google Scholar] [CrossRef]
- Camkurt, M.A.; Güneş, S.; Coşkun, S.; Fındıklı, E. Peripheral Signatures of Psychiatric Disorders: MicroRNAs. Clin. Psychopharmacol. Neurosci. 2017, 15, 313. [Google Scholar] [CrossRef] [Green Version]
- Luoni, A.; Riva, M.A. MicroRNAs and Psychiatric Disorders: From Aetiology to Treatment. Pharmacol. Ther. 2016, 167, 13–27. [Google Scholar] [CrossRef]
- Issler, O.; Chen, A. Determining the Role of MicroRNAs in Psychiatric Disorders. Nat. Rev. Neurosci. 2015, 16, 201–212. [Google Scholar] [CrossRef]
- Narahari, A.; Hussain, M.; Sreeram, V. MicroRNAs as Biomarkers for Psychiatric Conditions: A Review of Current Research. Innov. Clin. Neurosci. 2017, 14, 53. [Google Scholar]
- Roy, B.; Yoshino, Y.; Allen, L.; Prall, K.; Schell, G.; Dwivedi, Y. Exploiting Circulating MicroRNAs as Biomarkers in Psychiatric Disorders. Mol. Diagn. Ther. 2020, 24, 279–298. [Google Scholar] [CrossRef]
- Elhwuegi, A.S. Central Monoamines and Their Role in Major Depression. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2004, 28, 435–451. [Google Scholar] [CrossRef]
- Mulinari, S. Monoamine Theories of Depression: Historical Impact on Biomedical Research. J. Hist. Neurosci. 2012, 21, 366–392. [Google Scholar] [CrossRef]
- Hirschfeld, R.M. History and evolution of the monoamine hypothesis of depression. J. Clin. Psychiatry 2000, 61 (Suppl. 6), 4–6. [Google Scholar]
- Rot, M.A.H.; Mathew, S.J.; Charney, D.S. Neurobiological Mechanisms in Major Depressive Disorder. Can. Med. Assoc. J. 2009, 180, 305–313. [Google Scholar]
- A Quera Salva, M.; Hartley, S.; Barbot, F.; C Alvarez, J.; Lofaso, F.; Guilleminault, C. Circadian Rhythms, Melatonin and Depression. Curr. Pharm. Des. 2011, 17, 1459–1470. [Google Scholar] [CrossRef]
- Boku, S.; Nakagawa, S.; Toda, H.; Hishimoto, A. Neural Basis of Major Depressive Disorder: Beyond Monoamine Hypothesis. Psychiatry Clin. Neurosci. 2018, 72, 3–12. [Google Scholar] [CrossRef]
- Pandya, M.; Altinay, M.; Malone, D.A.; Anand, A. Where in the Brain Is Depression? Curr. Psychiatry Rep. 2012, 14, 634–642. [Google Scholar] [CrossRef] [Green Version]
- Bourin, M. Neurogenesis and Neuroplasticity in Major Depression: Its Therapeutic Implication. Adv. Exp. Med. Biol. 2021, 1305, 157–173. [Google Scholar] [CrossRef]
- Dwivedi, Y. Brain-Derived Neurotrophic Factor: Role in Depression and Suicide. Neuropsychiatr. Dis. Treat. 2009, 5, 433. [Google Scholar] [CrossRef] [Green Version]
- Benatti, C.; Blom, J.M.; Rigillo, G.; Alboni, S.; Zizzi, F.; Torta, R.; Brunello, N.; Tascedda, F. Disease-Induced Neuroinflammation and Depression. CNS Neurol Disord Drug Targets 2016, 15, 414–433. [Google Scholar] [CrossRef]
- Alvarez-Mon, M.A.; Gómez, A.M.; Orozco, A.; Lahera, G.; Sosa, M.D.; Diaz, D.; Auba, E.; Albillos, A.; Monserrat, J.; Alvarez-Mon, M. Abnormal Distribution and Function of Circulating Monocytes and Enhanced Bacterial Translocation in Major Depressive Disorder. Front. Psychiatry 2019, 10, 812. [Google Scholar] [CrossRef]
- Alvarez-Mon, M.A.; Gomez-Lahoz, A.M.; Orozco, A.; Lahera, G.; Sosa-Reina, M.D.; Diaz, D.; Albillos, A.; Quintero, J.; Molero, P.; Monser-rat, J.; et al. Blunted Expansion of Regulatory T Lymphocytes Is Associated With Increased Bacterial Translocation in Patients With Major Depressive Disorder. Front. Psychiatry 2021, 11, 591962. [Google Scholar] [CrossRef]
- Appleton, J. The Gut-Brain Axis: Influence of Microbiota on Mood and Mental Health. Integr. Med. A Clin. J. 2018, 17, 28. [Google Scholar]
- Jiménez-Fernández, S.; Gurpegui, M.; Díaz-Atienza, F.; Pérez-Costillas, L.; Gerstenberg, M.; Correll, C.U. Oxidative Stress and Antioxidant Parameters in Patients with Major Depressive Disorder Compared to Healthy Controls before and after Antidepressant Treatment: Results from a Meta-Analysis. J. Clin. Psychiatry 2015, 76, 1658–1667. [Google Scholar] [CrossRef]
- Bajpai, A.; Verma, A.K.; Srivastava, M.; Srivastava, R. Oxidative Stress and Major Depression. J. Clin. Diagn. Res. 2014, 8, CC04–CC07. [Google Scholar] [CrossRef]
- Duman, R.S.; Voleti, B. Signaling Pathways Underlying the Pathophysiology and Treatment of Depression: Novel Mechanisms for Rapid-Acting Agents. Trends Neurosci. 2012, 35, 47. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, Y. MicroRNAs as Biomarker in Depression Pathogenesis. Ann. Psychiatry Ment. Health 2013, 1, 1003. [Google Scholar]
- Martinetz, S. MicroRNA’s Impact on Neurotransmitter and Neuropeptide Systems: Small but Mighty Mediators of Anxiety. Pflug. Arch. Eur. J. Physiol. 2016, 468, 1061–1069. [Google Scholar] [CrossRef]
- Millan, M.J. MicroRNA in the Regulation and Expression of Serotonergic Transmission in the Brain and Other Tissues. Curr. Opin. Pharmacol. 2011, 11, 11–22. [Google Scholar] [CrossRef]
- Moya, P.R.; Wendland, J.R.; Salemme, J.; Fried, R.L.; Murphy, D.L. MiR-15a and MiR-16 Regulate Serotonin Transporter Expression in Human Placental and Rat Brain Raphe Cells. Int. J. Neuropsychopharmacol. 2013, 16, 621–629. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Hu, Z.; Du, X.; Davies, H.; Huo, X.; Fang, M. MiR-16 and Fluoxetine Both Reverse Autophagic and Apoptotic Change in Chronic Unpredictable Mild Stress Model Rats. Front. Neurosci. 2017, 11, 428. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wang, Z.C.; Zhu, M.X.; Fan, G.B.; Xu, G.S.; Zhao, T.Y.; Zhao, A.Y.; Ning, S.W.; Qi, S.H. Network and Pathway-Based Integrated Analysis Identified a Novel “Rs28457673-MiR-15/16/195/424/497 Family-IGF1R-MAPK Signaling Pathway” Axis Associated With Post-Stroke Depression. Front. Cell Dev. Biol. 2021, 8, 1723. [Google Scholar] [CrossRef] [PubMed]
- Towfighi, A.; Ovbiagele, B.; El Husseini, N.; Hackett, M.L.; Jorge, R.E.; Kissela, B.M.; Mitchell, P.H.; Skolarus, L.E.; Whooley, M.A.; Williams, L.S. Poststroke Depression: A Scientific Statement for Healthcare Professionals From the American Heart Association/American Stroke Association. Stroke 2017, 48, e30–e43. [Google Scholar] [CrossRef] [PubMed]
- Gorinski, N.; Bijata, M.; Prasad, S.; Wirth, A.; Abdel Galil, D.; Zeug, A.; Bazovkina, D.; Kondaurova, E.; Kulikova, E.; Ilchibaeva, T.; et al. Attenuated Palmitoylation of Serotonin Receptor 5-HT1A Affects Receptor Function and Contributes to Depression-like Behaviors. Nat. Commun. 2019, 10, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Lo Iacono, L.; Ielpo, D.; Accoto, A.; di Segni, M.; Babicola, L.; D’Addario, S.L.; Ferlazzo, F.; Pascucci, T.; Ventura, R.; Andolina, D. MicroRNA-34a Regulates the Depression-like Behavior in Mice by Modulating the Expression of Target Genes in the Dorsal Raphè. Mol. Neurobiol. 2019, 57, 823–836. [Google Scholar] [CrossRef] [PubMed]
- Bray, N. MiR-135: A Marker of Mood. Nat. Rev. Neurosci. 2014, 15, 495. [Google Scholar] [CrossRef]
- Issler, O.; Haramati, S.; Paul, E.D.; Maeno, H.; Navon, I.; Zwang, R.; Gil, S.; Mayberg, H.S.; Dunlop, B.W.; Menke, A.; et al. MicroRNA 135 Is Essential for Chronic Stress Resiliency, Antidepressant Efficacy, and Intact Serotonergic Activity. Neuron 2014, 83, 344–360. [Google Scholar] [CrossRef] [Green Version]
- Belujon, P.; Grace, A.A. Dopamine System Dysregulation in Major Depressive Disorders. Int. J. Neuropsychopharmacol. 2017, 20, 1036. [Google Scholar] [CrossRef] [Green Version]
- Simpson, E.H.; Winiger, V.; Biezonski, D.K.; Haq, I.; Kandel, E.R.; Kellendonk, C. Selective Overexpression of Dopamine D3 Receptors in the Striatum Disrupts Motivation but Not Cognition. Biol. Psychiatry 2014, 76, 823. [Google Scholar] [CrossRef] [Green Version]
- Bahi, A.; Dreyer, J.L. Lentiviral-Mediated Let-7d MicroRNA Overexpression Induced Anxiolytic- and Anti-Depressant-like Behaviors and Impaired Dopamine D3 Receptor Expression. Eur. Neuropsychopharmacol. J. Eur. Coll. Neuropsychopharmacol. 2018, 28, 1394–1404. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.; Li, M.D. Differential Allelic Expression of Dopamine D1 Receptor Gene (DRD1) Is Modulated by MicroRNA MiR-504. Biol. Psychiatry 2009, 65, 702. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Zhu, X.; Bai, M.; Zhang, L.; Xue, L.; Yi, J. Maternal Deprivation Enhances Behavioral Vulnerability to Stress Associated with MiR-504 Expression in Nucleus Accumbens of Rats. PLoS ONE 2013, 8, e69934. [Google Scholar] [CrossRef] [Green Version]
- Lopez, J.P.; Lim, R.; Cruceanu, C.; Crapper, L.; Fasano, C.; Labonte, B.; Maussion, G.; Yang, J.P.; Yerko, V.; Vigneault, E.; et al. MiR-1202: A Primate Specific and Brain Enriched MiRNA Involved in Major Depression and Antidepressant Treatment. Nat. Med. 2014, 20, 764. [Google Scholar] [CrossRef]
- Wan, Y.Q.; Feng, J.G.; Li, M.; Wang, M.Z.; Liu, L.; Liu, X.; Duan, X.X.; Zhang, C.X.; Wang, X.B. Prefrontal Cortex MiR-29b-3p Plays a Key Role in the Antidepressant-like Effect of Ketamine in Rats. Exp. Mol. Med. 2018, 50, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Wei, Y.B.; Melas, P.A.; Villaescusa, J.C.; Liu, J.J.; Xu, N.; Christiansen, S.H.; Elbrønd-Bek, H.; Woldbye, D.P.; Wegener, G.; Mathé, A.A.; et al. MicroRNA 101b Is Downregulated in the Prefrontal Cortex of a Genetic Model of Depression and Targets the Glutamate Transporter SLC1A1 (EAAT3) in Vitro. Int. J. Neuropsychopharmacol. 2016, 19, PYW069. [Google Scholar] [CrossRef] [Green Version]
- Roy, B.; Dunbar, M.; Shelton, R.C.; Dwivedi, Y. Identification of MicroRNA-124-3p as a Putative Epigenetic Signature of Major Depressive Disorder. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2017, 42, 864–875. [Google Scholar] [CrossRef] [Green Version]
- Gray, A.L.; Hyde, T.M.; Deep-Soboslay, A.; Kleinman, J.E.; Sodhi, M.S. Sex Differences in Glutamate Receptor Gene Expression in Major Depression and Suicide. Mol. Psychiatry 2015, 20, 1057–1068. [Google Scholar] [CrossRef]
- O’Connor, R.M.; Grenham, S.; Dinan, T.G.; Cryan, J.F. MicroRNAs as Novel Antidepressant Targets: Converging Effects of Ketamine and Electroconvulsive Shock Therapy in the Rat Hippocampus. Int. J. Neuropsychopharmacol. 2013, 16, 1885–1892. [Google Scholar] [CrossRef] [Green Version]
- Saus, E.; Soria, V.; Escaramís, G.; Vivarelli, F.; Crespo, J.M.; Kagerbauer, B.; Menchón, J.M.; Urretavizcaya, M.; Gratacòs, M.; Estivill, X. Genetic Variants and Abnormal Processing of Pre-MiR-182, a Circadian Clock Modulator, in Major Depression Patients with Late Insomnia. Hum. Mol. Genet. 2010, 19, 4017–4025. [Google Scholar] [CrossRef] [Green Version]
- Satyanarayanan, S.K.; Shih, Y.H.; Wen, Y.R.; Palani, M.; Lin, Y.W.; Su, H.; Gałecki, P.; Su, K.P. MiR-200a-3p Modulates Gene Expression in Comorbid Pain and Depression: Molecular Implication for Central Sensitization. Brain Behav. Immun. 2019, 82, 230–238. [Google Scholar] [CrossRef]
- Finkel, T.; Deng, C.X.; Mostoslavsky, R. Recent Progress in the Biology and Physiology of Sirtuins. Nature 2009, 460, 587–591. [Google Scholar] [CrossRef] [Green Version]
- Lu, G.; Li, J.; Zhang, H.; Zhao, X.; Yan, L.-J.; Yang, X. Role and Possible Mechanisms of Sirt1 in Depression. Oxidative Med. Cell. Longev. 2018, 2018, 8596903. [Google Scholar] [CrossRef] [Green Version]
- Nakahata, Y.; Kaluzova, M.; Grimaldi, B.; Sahar, S.; Hirayama, J.; Chen, D.; Guarente, L.P.; Sassone-Corsi, P. The NAD+-Dependent Deacetylase SIRT1 Modulates CLOCK-Mediated Chromatin Remodeling and Circadian Control. Cell 2008, 134, 329–340. [Google Scholar] [CrossRef] [Green Version]
- Gu, Z.; Pan, J.; Chen, L. MiR-124 Suppression in the Prefrontal Cortex Reduces Depression-like Behavior in Mice. Biosci. Rep. 2019, 39, BSR20190186. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Hu, S.; Dai, S.; Yi, Y.; Wang, T.; Li, X.; Luo, M.; Li, K.; Chen, L.; Wang, H.; et al. Programming Changes of Hippocampal MiR-134-5p/SOX2 Signal Mediate the Susceptibility to Depression in Prenatal Dexamethasone-Exposed Female Offspring. Cell Biol. Toxicol. 2021. [Google Scholar] [CrossRef]
- Li, C.; Wang, F.; Miao, P.; Yan, L.; Liu, S.; Wang, X.; Jin, Z.; Gu, Z. MiR-138 Increases Depressive-Like Behaviors by Targeting SIRT1 in Hippocampus. Neuropsychiatr. Dis. Treat. 2020, 16, 949. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Wang, B.; Zhao, J.; Liu, C.; Qu, X.; Li, Y. MiR-155 Is Involved in Major Depression Disorder and Antidepressant Treatment via Targeting SIRT1. Biosci. Rep. 2018, 38, BSR20181139. [Google Scholar] [CrossRef] [Green Version]
- Slattery, D.A.; Neumann, I.D. Oxytocin and Major Depressive Disorder: Experimental and Clinical Evidence for Links to Aetiology and Possible Treatment. Pharmaceuticals 2010, 3, 702. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.W.; Kang, S.M.; Lee, Y.; Hong, S.H.; Sanek, N.A.; Young, W.S.; Lee, H.J. MicroRNA Profiling in the Mouse Hypothalamus Reveals Oxytocin-Regulating MicroRNA. J. Neurochem. 2013, 126, 331–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tafet, G.E.; Nemeroff, C.B. The Links between Stress and Depression: Psychoneuroendocrinological, Genetic, and Environmental Interactions. J. Neuropsychiatry Clin. Neurosci. 2015, 28, 77–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, A.K.L.; Sharp, P.A. MicroRNA Functions in Stress Responses. Mol. Cell 2010, 40, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, L.; Dwivedi, Y. MicroRNA Mediators of Early Life Stress Vulnerability to Depression and Suicidal Behavior. Mol. Psychiatry 2020, 25, 308. [Google Scholar] [CrossRef] [Green Version]
- Bai, M.; Zhu, X.; Zhang, Y.; Zhang, S.; Zhang, L.; Xue, L.; Yi, J.; Yao, S.; Zhang, X. Abnormal Hippocampal BDNF and MiR-16 Expression Is Associated with Depression-like Behaviors Induced by Stress during Early Life. PLoS ONE 2012, 7, e0046921. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, D.; Xu, J.; Jiang, H.; Pan, F. Early Adolescent Stress-Induced Changes in Prefrontal Cortex MiRNA-135a and Hippocampal MiRNA-16 in Male Rats. Dev. Psychobiol. 2017, 59, 958–969. [Google Scholar] [CrossRef] [PubMed]
- Antoniuk, S.; Bijata, M.; Ponimaskin, E.; Wlodarczyk, J. Chronic Unpredictable Mild Stress for Modeling Depression in Rodents: Meta-Analysis of Model Reliability. Neurosci. Biobehav. Rev. 2019, 99, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Nolan, K.; Walter, F.; Tuffy, L.P.; Poeschel, S.; Gallagher, R.; Haunsberger, S.; Bray, I.; Stallings, R.L.; Concannon, C.G.; Prehn, J.H. Endoplasmic Reticulum Stress-Mediated Upregulation of MiR-29a Enhances Sensitivity to Neuronal Apoptosis. Eur. J. Neurosci. 2016, 43, 640–652. [Google Scholar] [CrossRef]
- Mao, J.; Hu, Y.; Ruan, L.; Ji, Y.; Lou, Z. Role of Endoplasmic Reticulum Stress in Depression. Mol. Med. Rep. 2019, 20, 4774. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.S.; Mu, R.H.; Li, C.F.; Dong, S.Q.; Geng, D.; Liu, Q.; Yi, L.T. MicroRNA-124 Targets Glucocorticoid Receptor and Is Involved in Depression-like Behaviors. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2017, 79, 417–425. [Google Scholar] [CrossRef]
- Dwivedi, Y. MicroRNA-124: A Putative Therapeutic Target and Biomarker for Major Depression. Expert Opin. Ther. Targets 2017, 21, 653–656. [Google Scholar] [CrossRef] [Green Version]
- Dwivedi, Y. Emerging Role of MicroRNAs in Major Depressive Disorder: Diagnosis and Therapeutic Implications. Dialogues Clin. Neurosci. 2014, 16, 43. [Google Scholar] [CrossRef]
- Dwivedi, Y. Evidence Demonstrating Role of MicroRNAs in the Etiopathology of Major Depression. J. Chem. Neuroanat. 2011, 42, 142. [Google Scholar] [CrossRef] [Green Version]
- Pittenger, C.; Duman, R.S. Stress, Depression, and Neuroplasticity: A Convergence of Mechanisms. Neuropsychopharmacology 2007, 33, 88–109. [Google Scholar] [CrossRef]
- Schoenfeld, T.; Gould, E. Stress, Stress Hormones, and Adult Neurogenesis. Exp. Neurol. 2012, 233, 12. [Google Scholar] [CrossRef] [Green Version]
- Surget, A.; Belzung, C. Adult Hippocampal Neurogenesis Shapes Adaptation and Improves Stress Response: A Mechanistic and Integrative Perspective. Mol. Psychiatry 2021, 1–19. [Google Scholar] [CrossRef]
- Martins, H.C.; Schratt, G. MicroRNA-Dependent Control of Neuroplasticity in Affective Disorders. Transl. Psychiatry 2021, 11, 1–16. [Google Scholar] [CrossRef]
- Volk, N.; Pape, J.C.; Engel, M.; Zannas, A.S.; Cattane, N.; Cattaneo, A.; Binder, E.B.; Chen, A. Amygdalar MicroRNA-15a Is Essential for Coping with Chronic Stress. Cell Rep. 2016, 17, 1882. [Google Scholar] [CrossRef] [Green Version]
- Lekman, M.; Laje, G.; Charney, D.; Rush, A.J.; Wilson, A.F.; Sorant, A.J.M.; Lipsky, R.; Wisniewski, S.R.; Manji, H.; McMahon, F.J.; et al. The FKBP5-Gene in Depression and Treatment Response—An Association Study in the STAR*D-Cohort. Biol. Psychiatry 2008, 63, 1103. [Google Scholar] [CrossRef] [Green Version]
- Khandelwal, N.; Dey, S.K.; Chakravarty, S.; Kumar, A. MiR-30 Family MiRNAs Mediate the Effect of Chronic Social Defeat Stress on Hippocampal Neurogenesis in Mouse Depression Model. Front. Mol. Neurosci. 2019, 12, 188. [Google Scholar] [CrossRef] [Green Version]
- Fan, C.; Zhu, X.; Song, Q.; Wang, P.; Liu, Z.; Yu, S. MiR-134 Modulates Chronic Stress-Induced Structural Plasticity and Depression-like Behaviors via Downregulation of Limk1/Cofilin Signaling in Rats. Neuropharmacology 2018, 131, 364–376. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, M.; Wang, X.; Liu, K.; Wan, Y.; Li, M.; Liu, L.; Zhang, C. Abnormal Expression of MicroRNAs Induced by Chronic Unpredictable Mild Stress in Rat Hippocampal Tissues. Mol. Neurobiol. 2017, 55, 917–935. [Google Scholar] [CrossRef]
- Torres-Berrío, A.; Nouel, D.; Cuesta, S.; Parise, E.M.; Restrepo-Lozano, J.M.; Larochelle, P.; Nestler, E.J.; Flores, C. MiR-218: A Molecular Switch and Potential Biomarker of Susceptibility to Stress. Mol. Psychiatry 2019, 25, 951–964. [Google Scholar] [CrossRef]
- Yoshino, Y.; Roy, B.; Dwivedi, Y. Differential and Unique Patterns of Synaptic MiRNA Expression in Dorsolateral Prefrontal Cortex of Depressed Subjects. Neuropsychopharmacology 2020, 46, 900–910. [Google Scholar] [CrossRef]
- Zurawek, D.; Gruca, P.; Antkiewicz-Michaluk, L.; Dziedzicka-Wasylewska, M. Resilient Phenotype in Chronic Mild Stress Paradigm Is Associated with Altered Expression Levels of MiR-18a-5p and Serotonin 5-HT1a Receptor in Dorsal Part of the Hippocampus. Mol. Neurobiol. 2019, 56, 7680–7693. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Wang, R.; Liu, Y.; Wang, W.; Liu, D.; Jiang, H.; Pan, F. Short- and Long-Term Alterations of FKBP5-GR and Specific MicroRNAs in the Prefrontal Cortex and Hippocampus of Male Rats Induced by Adolescent Stress Contribute to Depression Susceptibility. Psychoneuroendocrinology 2019, 101, 204–215. [Google Scholar] [CrossRef]
- Zhao, Y.; Wang, S.; Chu, Z.; Dang, Y.; Zhu, J.; Su, X. MicroRNA-101 in the Ventrolateral Orbital Cortex (VLO) Modulates Depressive-like Behaviors in Rats and Targets Dual-Specificity Phosphatase 1 (DUSP1). Brain Res. 2017, 1669, 55–62. [Google Scholar] [CrossRef]
- Li, Y.; Wang, N.; Pan, J.; Wang, X.; Zhao, Y.; Guo, Z. Hippocampal MiRNA-144 Modulates Depressive-Like Behaviors in Rats by Targeting PTP1B. Neuropsychiatr. Dis. Treat. 2021, 17, 389. [Google Scholar] [CrossRef]
- Pei, G.; Xu, L.; Huang, W.; Yin, J. The Protective Role of MicroRNA-133b in Restricting Hippocampal Neurons Apoptosis and Inflammatory Injury in Rats with Depression by Suppressing CTGF. Int. Immunopharmacol. 2020, 78. [Google Scholar] [CrossRef]
- Yang, W.; Liu, M.; Zhang, Q.; Zhang, J.; Chen, J.; Chen, Q.; Suo, L. Knockdown of MiR-124 Reduces Depression-like Behavior by Targeting CREB1 and BDNF. Curr. Neurovascular Res. 2020, 17, 196. [Google Scholar] [CrossRef] [PubMed]
- Lian, N.; Niu, Q.; Lei, Y.; Li, X.; Li, Y.; Song, X. MiR-221 Is Involved in Depression by Regulating Wnt2/CREB/BDNF Axis in Hippocampal Neurons. Cell Cycle 2018, 17, 2745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prabu, P.; Poongothai, S.; Shanthirani, C.S.; Anjana, R.M.; Mohan, V.; Balasubramanyam, M. Altered Circulatory Levels of MiR-128, BDNF, Cortisol and Shortened Telomeres in Patients with Type 2 Diabetes and Depression. Acta Diabetol. 2020, 57, 799–807. [Google Scholar] [CrossRef]
- Li, Y.J.; Xu, M.; Gao, Z.H.; Wang, Y.Q.; Yue, Z.; Zhang, Y.X.; Li, X.X.; Zhang, C.; Xie, S.Y.; Wang, P.Y. Alterations of Serum Levels of BDNF-Related MiRNAs in Patients with Depression. PLoS ONE 2013, 8, e0063648. [Google Scholar] [CrossRef] [Green Version]
- Fonken, L.K.; Gaudet, A.D.; Gaier, K.R.; Nelson, R.J.; Popovich, P.G. MicroRNA-155 Deletion Reduces Anxiety- and Depressive-like Behaviors in Mice. Psychoneuroendocrinology 2016, 63, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Peruga, I.; Hartwig, S.; Merkler, D.; Thöne, J.; Hovemann, B.; Juckel, G.; Gold, R.; Linker, R.A. Endogenous Ciliary Neurotrophic Factor Modulates Anxiety and Depressive-like Behavior. Behav. Brain Res. 2012, 229, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Papanota, A.-M.; Karousi, P.; Kontos, C.K.; Ntanasis-Stathopoulos, I.; Scorilas, A.; Terpos, E. Multiple Myeloma Bone Disease: Implication of MicroRNAs in Its Molecular Background. Int. J. Mol. Sci. 2021, 22, 2375. [Google Scholar] [CrossRef]
- Artemaki, P.I.; Letsos, P.A.; Zoupa, I.C.; Katsaraki, K.; Karousi, P.; Papageorgiou, S.G.; Pappa, V.; Scorilas, A.; Kontos, C.K. The Multifaceted Role and Utility of MicroRNAs in Indolent B-Cell Non-Hodgkin Lymphomas. Biomedicines 2021, 9, 333. [Google Scholar] [CrossRef]
- Katsaraki, K.; Karousi, P.; Artemaki, P.I.; Scorilas, A.; Pappa, V.; Kontos, C.K.; Papageorgiou, S.G. MicroRNAs: Tiny Regulators of Gene Expression with Pivotal Roles in Normal B-Cell Development and B-Cell Chronic Lymphocytic Leukemia. Cancers 2021, 13, 593. [Google Scholar] [CrossRef]
- Ferrúa, C.P.; Giorgi, R.; da Rosa, L.C.; do Amaral, C.C.; Ghisleni, G.C.; Pinheiro, R.T.; Nedel, F. MicroRNAs Expressed in Depression and Their Associated Pathways: A Systematic Review and a Bioinformatics Analysis. J. Chem. Neuroanat. 2019, 100, 101650. [Google Scholar] [CrossRef] [PubMed]
- Gururajan, A.; Naughton, M.E.; Scott, K.A.; O’Connor, R.M.; Moloney, G.; Clarke, G.; Dowling, J.; Walsh, A.; Ismail, F.; Shorten, G.; et al. MicroRNAs as Biomarkers for Major Depression: A Role for Let-7b and Let-7c. Transl. Psychiatry 2016, 6, e862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Fan, C.; Wang, L.; Lan, T.; Gao, R.; Wang, W.; Yu, S.Y. MicroRNA-26a-3p Rescues Depression-like Behaviors in Male Rats via Preventing Hippocampal Neuronal Anomalies. J. Clin. Investig. 2021, 131. [Google Scholar] [CrossRef]
- Matsuda, S.; Ikeda, Y.; Murakami, M.; Nakagawa, Y.; Tsuji, A.; Kitagishi, Y. Roles of PI3K/AKT/GSK3 Pathway Involved in Psychiatric Illnesses. Diseases 2019, 7, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, J.P.; Fiori, L.M.; Cruceanu, C.; Lin, R.; Labonte, B.; Cates, H.M.; Heller, E.A.; Vialou, V.; Ku, S.M.; Gerald, C.; et al. MicroRNAs 146a/b-5 and 425-3p and 24-3p Are Markers of Antidepressant Response and Regulate MAPK/Wnt-System Genes. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Pan, J.Y.; Liao, N.; Shi, J.; Zeng, Q.; Huang, L.; Chen, L.P. Influence of MiR-155 on Behaviors of Depression Mice through Regulating Wnt/β-Catenin Signaling Pathway. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 1398–1407. [Google Scholar] [CrossRef]
- Roy, B.; Dunbar, M.; Agrawal, J.; Allen, L.; Dwivedi, Y. Amygdala-Based Altered MiRNome and Epigenetic Contribution of MiR-128-3p in Conferring Susceptibility to Depression-Like Behavior via Wnt Signaling. Int. J. Neuropsychopharmacol. 2020, 23, 165–177. [Google Scholar] [CrossRef]
- Felger, J.C. Role of Inflammation in Depression and Treatment Implications. Handb. Exp. Pharmacol. 2019, 250, 255–286. [Google Scholar] [CrossRef]
- Alvarez-Mon, M.A.; Gómez-Lahoz, A.M.; Orozco, A.; Lahera, G.; Diaz, D.; Ortega, M.A.; Albillos, A.; Quintero, J.; Aubá, E.; Monserrat, J.; et al. Expansion of CD4 T Lymphocytes Expressing Interleukin 17 and Tumor Necrosis Factor in Patients with Major Depressive Disorder. J. Pers. Med. 2021, 11, 220. [Google Scholar] [CrossRef]
- Sun, L.; Bai, D.; Lin, M.; Eerdenidalai Zhang, L.; Wang, F.; Jin, S. MiR-96 Inhibits SV2C to Promote Depression-Like Behavior and Memory Disorders in Mice. Front. Behav. Neurosci. 2021, 14, 281. [Google Scholar] [CrossRef]
- Sun, X.H.; Song, M.F.; Song, H.D.; Wang, Y.W.; Luo, M.J.; Yin, L.M. MiR-155 Mediates Inflammatory Injury of Hippocampal Neuronal Cells via the Activation of Microglia. Mol. Med. Rep. 2019, 19, 2627–2635. [Google Scholar] [CrossRef]
- Boxberger, N.; Hecker, M.; Zettl, U.K. Dysregulation of Inflammasome Priming and Activation by MicroRNAs in Human Immune-Mediated Diseases. J. Immunol. 2019, 202, 2177–2187. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, F.N.; Costa, A.P.; Ghisleni, G.; Diaz, A.P.; Rodrigues, A.L.S.; Peluffo, H.; Kaster, M.P. NLRP3 Inflammasome-Driven Pathways in Depression: Clinical and Preclinical Findings. Brain Behav. Immun. 2017, 64, 367–383. [Google Scholar] [CrossRef] [PubMed]
- Lou, D.; Wang, J.; Wang, X. MiR-124 Ameliorates Depressive-like Behavior by Targeting STAT3 to Regulate Microglial Activation. Mol. Cell. Probes 2019, 48, 101470. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Zhong, M.; Qiu, B.; Liu, C.; Wang, J.; Liang, J. Abnormal Expression of MiR-135a in Patients with Depression and Its Possible Involvement in the Pathogenesis of the Condition. Exp. Ther. Med. 2021, 22, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Buisman-Pijlman, F.; Hutchinson, M.R. Toll-like Receptor 4: Innate Immune Regulator of Neuroimmune and Neuroendocrine Interactions in Stress and Major Depressive Disorder. Front. Neurosci. 2014, 8, 309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sundquist, K.; Memon, A.A.; Palmér, K.; Sundquist, J.; Wang, X. Inflammatory Proteins and MiRNA-144-5p in Patients with Depression, Anxiety, or Stress- and Adjustment Disorders after Psychological Treatment. Cytokine 2021, 146, 155646. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.P.; Zhong, M.; Sun, J.X.; He, J.; Gao, Y.; Qin, F.X. MiR-146a Reduces Depressive Behavior by Inhibiting Microglial Activation. Mol. Med. Rep. 2021, 23. [Google Scholar] [CrossRef]
- Merchak, A.; Gaultier, A. Microbial Metabolites and Immune Regulation: New Targets for Major Depressive Disorder. Brain Behav. Immun. Health 2020, 9, 100169. [Google Scholar] [CrossRef]
- Hoban, A.E.; Stilling, R.M.; MMoloney, G.; Moloney, R.D.; Shanahan, F.; Dinan, T.G.; Cryan, J.F.; Clarke, G. Microbial Regulation of MicroRNA Expression in the Amygdala and Prefrontal Cortex. Microbiome 2017, 5, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Wang, H.; Yu, Y.; Zeng, B.; Rao, X.; Chen, J.; Zhou, C.; Zheng, P.; Pu, J.; Yang, L.; et al. Microbial Regulation of a LincRNA–MiRNA–MRNA Network in the Mouse Hippocampus. Epigenomics 2020, 12, 1377–1383. [Google Scholar] [CrossRef]
- Chen, J.J.; Zeng, B.H.; Li, W.W.; Zhou, C.J.; Fan, S.H.; Cheng, K.; Zeng, L.; Zheng, P.; Fang, L.; Wei, H.; et al. Effects of Gut Microbiota on the MicroRNA and MRNA Expression in the Hippocampus of Mice. Behav. Brain Res. 2017, 322, 34–41. [Google Scholar] [CrossRef]
- O’Connor, J.C.; Lawson, M.A.; André, C.; Moreau, M.; Lestage, J.; Castanon, N.; Kelley, K.W.; Dantzer, R. Lipopolysaccharide-Induced Depressive-like Behavior Is Mediated by Indoleamine 2,3-Dioxygenase Activation in Mice. Mol. Psychiatry 2009, 14, 511. [Google Scholar] [CrossRef] [Green Version]
- Suento, W.J.; Kunisawa, K.; Wulaer, B.; Kosuge, A.; Iida, T.; Fujigaki, S.; Fujigaki, H.; Yamamoto, Y.; Tanra, A.J.; Saito, K.; et al. Prefrontal Cortex MiR-874-3p Prevents Lipopolysaccharide-Induced Depression-like Behavior through Inhibition of Indoleamine 2,3-Dioxygenase 1 Expression in Mice. J. Neurochem. 2021, 157, 1963–1978. [Google Scholar] [CrossRef]
- Karen, C.; Shyu, D.J.H.; Rajan, K.E. Lactobacillus Paracasei Supplementation Prevents Early Life Stress-Induced Anxiety and Depressive-Like Behavior in Maternal Separation Model-Possible Involvement of Microbiota-Gut-Brain Axis in Differential Regulation of MicroRNA124a/132 and Glutamate Receptors. Front. Neurosci. 2021, 1115. [Google Scholar] [CrossRef]
- DeVries, L.D.; Carlson, C.D. Consumption of B. Longnum Changes Expression of MiR-652 in the Brains of Mice. FASEB J. 2020, 34, 1. [Google Scholar] [CrossRef]
- García-Montero, C.; Fraile-Martínez, O.; Gómez-Lahoz, A.M.; Pekarek, L.; Castellanos, A.J.; Noguerales-Fraguas, F.; Coca, S.; Guijarro, L.G.; García-Honduvilla, N.; Asúnsolo, A.; et al. Nutritional Components in Western Diet versus Mediterranean Diet at the Gut Microbiota-Immune System Interplay. Implications for Health and Disease. Nutrients 2021, 13, 699. [Google Scholar] [CrossRef]
- Díez-Sainz, E.; Lorente-Cebrián, S.; Aranaz, P.; Riezu-Boj, J.I.; Martínez, J.A.; Milagro, F.I. Potential Mechanisms Linking Food-Derived MicroRNAs, Gut Microbiota and Intestinal Barrier Functions in the Context of Nutrition and Human Health. Front. Nutr. 2021, 8, 85. [Google Scholar] [CrossRef]
- Schatzberg, A.F. Scientific Issues Relevant to Improving the Diagnosis, Risk Assessment, and Treatment of Major Depression. Am. J. Psychiatry 2019, 176, 342–347. [Google Scholar] [CrossRef]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders (DSM-5®); American Psychiatric Association: Arlington, TX, USA, 2013. [Google Scholar]
- Tolentino, J.C.; Schmidt, S.L. DSM-5 Criteria and Depression Severity: Implications for Clinical Practice. Front. Psychiatry 2018, 9, 450. [Google Scholar] [CrossRef] [Green Version]
- Hamilton, M. A Rating Scale for Depression. J. Neurol. Neurosurg. Psychiatry 1960, 23, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Zimmerman, M.; Martinez, J.H.; Young, D.; Chelminski, I.; Dalrymple, K. Severity Classification on the Hamilton Depression Rating Scale. J. Affect. Disord. 2013, 150, 384–388. [Google Scholar] [CrossRef]
- Rohan, K.J.; Rough, J.N.; Evans, M.; Ho, S.Y.; Meyerhoff, J.; Roberts, L.M.; Vacek, P.M. A Protocol for the Hamilton Rating Scale for Depression: Item Scoring Rules, Rater Training, and Outcome Accuracy with Data on Its Application in a Clinical Trial. J. Affect. Disord. 2016, 200, 111–118. [Google Scholar] [CrossRef] [Green Version]
- Carrozzino, D.; Patierno, C.; Fava, G.A.; Guidi, J. The Hamilton Rating Scales for Depression: A Critical Review of Clinimetric Properties of Different Versions. Psychother. Psychosom. 2020, 89, 133–150. [Google Scholar] [CrossRef]
- Ng, C.W.M.; How, C.H.; Ng, Y.P. Major Depression in Primary Care: Making the Diagnosis. Singap. Med. J. 2016, 57, 591. [Google Scholar] [CrossRef] [Green Version]
- InformedHealth.org. Cologne, Germany: Institute for Quality and Efficiency in Health Care (IQWiG); 2006-. Depression: How Effective Are Antidepressants. Available online: https://wwwncbinlmnihgov/books/NBK361016/ (accessed on 25 October 2021).
- Maslej, M.M.; Furukawa, T.A.; Cipriani, A.; Andrews, P.W.; Sanches, M.; Tomlinson, A.; Volkmann, C.; McCutcheon, R.A.; Howes, O.; Guo, X.; et al. Individual Differences in Response to Antidepressants: A Meta-Analysis of Placebo-Controlled Randomized Clinical Trials. JAMA Psychiatry 2021, 78, 490–497. [Google Scholar] [CrossRef]
- Kirsch, I. Antidepressants and the Placebo Effect. Z. Fur. Psychol. 2014, 222, 128–134. [Google Scholar] [CrossRef]
- Cheung, C.P.; Thiyagarajah, M.T.; Abraha, H.Y.; Liu, C.S.; Lanctôt, K.L.; Kiss, A.J.; Saleem, M.; Juda, A.; Levitt, A.J.; Schaffer, A.; et al. The Association between Placebo Arm Inclusion and Adverse Event Rates in Antidepressant Randomized Controlled Trials: An Examination of the Nocebo Effect. J. Affect. Disord. 2021, 280, 140–147. [Google Scholar] [CrossRef]
- Smith, K.M.; Renshaw, P.F.; Bilello, J. The Diagnosis of Depression: Current and Emerging Methods. Compr. Psychiatry 2013, 54, 1. [Google Scholar] [CrossRef] [Green Version]
- Dunlop, B.W. Prediction of Treatment Outcomes in Major Depressive Disorder. Expert Rev. Clin. Pharmacol. 2015, 8, 669–672. [Google Scholar] [CrossRef]
- Shimada, M.; Otowa, T.; Miyagawa, T.; Umekage, T.; Kawamura, Y.; Bundo, M.; Iwamoto, K.; Ikegame, T.; Tochigi, M.; Kasai, K.; et al. An Epigenome-Wide Methylation Study of Healthy Individuals with or without Depressive Symptoms. J. Hum. Genet. 2018, 63, 319–326. [Google Scholar] [CrossRef]
- Belzeaux, R.; Lin, R.; Ju, C.; Chay, M.A.; Fiori, L.M.; Lutz, P.E.; Turecki, G. Transcriptomic and Epigenomic Biomarkers of Antidepressant Response. J. Affect. Disord. 2018, 233, 36–44. [Google Scholar] [CrossRef]
- Lin, E.; Tsai, S.-J. Epigenetics and Depression: An Update. Psychiatry Investig. 2019, 16, 654. [Google Scholar] [CrossRef]
- Liang, Y.; Zhao, G.; Sun, R.; Mao, Y.; Li, G.; Chen, X.; Gao, L.; Hu, Z. Genetic Variants in the Promoters of Let-7 Family Are Associated with an Increased Risk of Major Depressive Disorder. J. Affect. Disord. 2015, 183, 295–299. [Google Scholar] [CrossRef]
- Xie, L.; Chen, J.; Ding, Y.M.; Gui, X.W.; Wu, L.X.; Tian, S.; Wu, W. MicroRNA-26a-2 Maintains Stress Resiliency and Antidepressant Efficacy by Targeting the Serotonergic Autoreceptor HTR1A. Biochem. Biophys. Res. Commun. 2019, 511, 440–446. [Google Scholar] [CrossRef]
- Solich, J.; Kuśmider, M.; Faron-Górecka, A.; Pabian, P.; Dziedzicka-Wasylewska, M. Restraint Stress in Mice Alters Set of 25 MiRNAs Which Regulate Stress- and Depression-Related MRNAs. Int. J. Mol. Sci. 2020, 21, 9469. [Google Scholar] [CrossRef]
- Hu, J.; Zhou, W.; Zhou, Z.; Yang, Q.; Xu, J.; Dong, W. MiR-22 and Cerebral Microbleeds in Brainstem and Deep Area Are Associated with Depression One Month after Ischemic Stroke. Braz. J. Med Biol. Res. 2020, 53. [Google Scholar] [CrossRef]
- Xu, Y.; Liu, H.; Li, F.; Sun, N.; Ren, Y.; Liu, Z.; Cao, X.; Wang, Y.; Liu, P.; Zhang, K. A Polymorphism in the MicroRNA-30e Precursor Associated with Major Depressive Disorder Risk and P300 Waveform. J. Affect. Disord. 2010, 127, 332–336. [Google Scholar] [CrossRef]
- Hu, J.; Zhou, Z.; Yang, Q.; Yang, K. Differential Expression of MiR-30a-5p in Post Stroke Depression and Bioinformatics Analysis of the Possible Mechanism. Nan Fang Yi Ke Da Xue Xue Bao= J. South. Med. Univ. 2020, 40, 922–929. [Google Scholar] [CrossRef]
- Cattaneo, A.; Suderman, M.; Cattane, N.; Mazzelli, M.; Begni, V.; Maj, C.; D’Aprile, I.; Pariante, C.M.; Luoni, A.; Berry, A.; et al. Long-Term Effects of Stress Early in Life on MicroRNA-30a and Its Network: Preventive Effects of Lurasidone and Potential Implications for Depression Vulnerability. Neurobiol. Stress 2020, 13, 100271. [Google Scholar] [CrossRef]
- Lin, C.C.; Lee, C.T.; Sun, M.H.; Huang, T.L. Increased Levels of miR-30e, miR-132, miR-185, and miR-212 at Baseline and Increased Brain-derived Neurotrophic Factor Protein and mRNA Levels after Treatment in Patients with Major Depressive Disorder. Neuropsychiatry 2017, 7, 920–926. [Google Scholar]
- Xu, C.; Yang, C.; Zhang, A.; Xu, Y.; Li, X.; Liu, Z.; Liu, S.; Sun, N.; Zhang, K. The Interaction of MiR-34b/c Polymorphisms and Negative Life Events Increases Susceptibility to Major Depressive Disorder in Han Chinese Population. Neurosci. Lett. 2017, 651, 65–71. [Google Scholar] [CrossRef] [PubMed]
- Yi, L.-T.; Zhu, J.-X.; Dong, S.-Q.; Li, C.-F.; Zhang, Q.-P.; Cheng, J.; Liu, Q. MiR-34a Induces Spine Damages via Inhibiting Synaptotagmin-1 in Depression. Neurobiol. Stress 2020, 13, 100243. [Google Scholar] [CrossRef] [PubMed]
- Kuang, W.-H.; Dong, Z.-Q.; Tian, L.-T.; Li, J. MicroRNA-451a, MicroRNA-34a-5p, and MicroRNA-221-3p as Predictors of Response to Antidepressant Treatment. Braz. J. Med Biol. Res. 2018, 51. [Google Scholar] [CrossRef] [Green Version]
- Sun, N.; Yang, C.; He, X.; Liu, Z.; Liu, S.; Li, X.; Wang, Y.; Jin, R.; Zhang, K. Impact of Expression and Genetic Variation of MicroRNA-34b/c on Cognitive Dysfunction in Patients with Major Depressive Disorder. Neuropsychiatr. Dis. Treat. 2020, 16, 1543–1554. [Google Scholar] [CrossRef]
- Jiménez, K.M.; Pereira-Morales, A.J.; Forero, D.A. Functional Polymorphism in the DRD1 Gene, That Modulates Its Regulation by MiR-504, Is Associated with Depressive Symptoms. Psychiatry Investig. 2018, 15, 402–406. [Google Scholar] [CrossRef] [Green Version]
- Song, M.F.; Dong, J.Z.; Wang, Y.W.; He, J.; Ju, X.; Zhang, L.; Zhang, Y.H.; Shi, J.F.; Lv, Y.Y. CSF MiR-16 Is Decreased in Major Depression Patients and Its Neutralization in Rats Induces Depression-like Behaviors via a Serotonin Transmitter System. J. Affect. Disord. 2015, 178, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.P.; Liu, X.L.; Chen, J.J.; Cheng, K.; Bai, S.J.; Zheng, P.; Zhou, C.J.; Wang, W.; Wang, H.Y.; Zhong, L.M.P. Circulating MicroRNA 134 Sheds Light on the Diagnosis of Major Depressive Disorder. Transl. Psychiatry 2020, 10, 95. [Google Scholar] [CrossRef]
- Mendes-Silva, A.P.; Fujimura, P.T.; Silva, J.R.D.C.; Teixeira, A.L.; Vieira, E.M.; Guedes, P.H.G.; Barroso, L.S.S.; Nicolau, M.S.; Ferreira, J.D.R.; Bertola, L.; et al. Brain-Enriched MicroRNA-184 Is Downregulated in Older Adults with Major Depressive Disorder: A Translational Study. J. Psychiatr. Res. 2019, 111, 110–120. [Google Scholar] [CrossRef]
- Serafini, G.; Trabucco, A.; Corsini, G.; Escelsior, A.; Amerio, A.; Aguglia, A.; Nasrallah, H.; Amore, M. The Potential of MicroRNAs as Putative Biomarkers in Major Depressive Disorder and Suicidal Behavior. Biomark. Neuropsychiatry 2021, 5, 100035. [Google Scholar] [CrossRef]
- Wan, Y.; Liu, Y.; Wang, X.; Wu, J.; Liu, K.; Zhou, J.; Liu, L.; Zhang, C. Identification of Differential MicroRNAs in Cerebrospinal Fluid and Serum of Patients with Major Depressive Disorder. PLoS ONE 2015, 10, e0121975. [Google Scholar] [CrossRef]
- Feng, J.; Wang, M.; Li, M.; Yang, J.; Jia, J.; Liu, L.; Zhou, J.; Zhang, C.; Wang, X. Serum MiR-221-3p as a New Potential Biomarker for Depressed Mood in Perioperative Patients. Brain Res. 2019, 1720, 146296. [Google Scholar] [CrossRef]
- Cui, Y.; Ma, G.; Kong, F.; Song, L. View of Diagnostic Values of MiR-221-3p in Serum and Cerebrospinal Fluid for Post-Stroke Depression and Analysis of Risk Factors. Iran J. Public Health 2021, 50, 1241–1249. [Google Scholar] [PubMed]
- Dwivedi, Y. MicroRNAs in Depression and Suicide: Recent Insights and Future Perspectives. J. Affect. Disord. 2018, 240, 146. [Google Scholar] [CrossRef] [PubMed]
- Smalheiser, N.R.; Lugli, G.; Rizavi, H.S.; Torvik, V.I.; Turecki, G.; Dwivedi, Y. MicroRNA Expression Is Down-Regulated and Reorganized in Prefrontal Cortex of Depressed Suicide Subjects. PLoS ONE 2012, 7, e0033201. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Sundquist, K.; Hedelius, A.; Palmér, K.; Memon, A.A.; Sundquist, J. Circulating MicroRNA-144-5p Is Associated with Depressive Disorders. Clin. Epigenetics 2015, 7, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Enatescu, V.R.; Papava, I.; Enatescu, I.; Antonescu, M.; Anghel, A.; Seclaman, E.; Sirbu, I.O.; Marian, C. Circulating Plasma Micro RNAs in Patients with Major Depressive Disorder Treated with Antidepressants: A Pilot Study. Psychiatry Investig. 2016, 13, 549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, M.; Tufan, T.; Raza, M.U.; Jones, T.C.; Zhu, M.-Y. MicroRNAs 29b and 181a Down-Regulate the Expression of the Norepinephrine Transporter and Glucocorticoid Receptors in PC12 Cells. J. Neurochem. 2016, 139, 197. [Google Scholar] [CrossRef] [Green Version]
- Qi, B.; Fiori, L.M.; Turecki, G.; Trakadis, Y.J. Machine Learning Analysis of Blood MicroRNA Data in Major Depression: A Case-Control Study for Biomarker Discovery. Int. J. Neuropsychopharmacol. 2020, 23, 505. [Google Scholar] [CrossRef] [PubMed]
- Gheysarzadeh, A.; Sadeghifard, N.; Afraidooni, L.; Pooyan, F.; Mofid, M.R.; Valadbeigi, H.; Bakhtiari, H.; Keikhavani, S. Serum-Based MicroRNA Biomarkers for Major Depression: MiR-16, MiR-135a, and MiR-1202. J. Res. Med. Sci. Off. J. Isfahan Univ. Med. Sci. 2018, 23, 69. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, L.; Cheng, K.; Wang, X.; Ren, G.; Xie, P. Identification of Suitable Plasma-Based Reference Genes for MiRNAome Analysis of Major Depressive Disorder. J. Affect. Disord. 2014, 163, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Serafini, G.; Pompili, M.; Hansen, K.F.; Obrietan, K.; Dwivedi, Y.; Shomron, N.; Girardi, P. The Involvement of MicroRNAs in Major Depression, Suicidal Behavior, and Related Disorders: A Focus on MiR-185 and MiR-491-3p. Cell. Mol. Neurobiol. 2014, 34, 17–30. [Google Scholar] [CrossRef]
- Kaadt, E.; Højgaard, K.; Mumm, B.; Christiansen, S.L.; Müller, H.K.; Damgaard, C.K.; Elfving, B. Dysregulation of MiR-185, MiR-193a, and MiR-450a in the Skin Are Linked to the Depressive Phenotype. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 104, 110052. [Google Scholar] [CrossRef]
- Hung, Y.-Y.; Wu, M.-K.; Tsai, M.-C.; Huang, Y.-L.; Kang, H.-Y. Aberrant Expression of Intracellular Let-7e, MiR-146a, and MiR-155 Correlates with Severity of Depression in Patients with Major Depressive Disorder and Is Ameliorated after Antidepressant Treatment. Cells 2019, 8, 647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, C.; Bai, Y.; Wang, Z.; Fan, D.; Wang, Q.; Liu, X.; Zhang, H.; Zhang, H.; Zhang, Z.; Yao, H.; et al. Identification of MicroRNA-9 Linking the Effects of Childhood Maltreatment on Depression Using Amygdala Connectivity. NeuroImage 2021, 224, 117428. [Google Scholar] [CrossRef]
- Zheng, Z.; Zeng, Y.; Huang, H.; Xu, F. MicroRNA-132 May Play a Role in Coexistence of Depression and Cardiovascular Disease: A Hypothesis. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2013, 19, 438. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Yang, X.; Zhao, L.; Zhang, J.T.; Ma, X. Increased MiR-132 Level Is Associated with Visual Memory Dysfunction in Patients with Depression. Neuropsychiatr. Dis. Treat. 2016, 12, 2905–2911. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.T.; Xu, W.; Tan, C.C.; Andrieu, S.; Suckling, J.; Evangelou, E.; Pan, A.; Zhang, C.; Jia, J.; Feng, L.; et al. Evidence-Based Prevention of Alzheimer’s Disease: Systematic Review and Meta-Analysis of 243 Observational Prospective Studies and 153 Randomised Controlled Trials. J. Neurol. Neurosurg. Psychiatry 2020, 91, 1201–1209. [Google Scholar] [CrossRef] [PubMed]
- Ownby, R.L.; Crocco, E.; Acevedo, A.; John, V.; Loewenstein, D. Depression and Risk for Alzheimer Disease: Systematic Review, Meta-Analysis, and Metaregression Analysis. Arch. Gen. Psychiatry 2006, 63, 530–538. [Google Scholar] [CrossRef] [Green Version]
- Chi, S.; Wang, C.; Jiang, T.; Zhu, X.C.; Yu, J.T.; Tan, L. The Prevalence of Depression in Alzheimer’s Disease: A Systematic Review and Meta-Analysis. Curr. Alzheimer Res. 2015, 12, 189–198. [Google Scholar] [CrossRef]
- Van den Berg, M.M.J.; Krauskopf, J.; Ramaekers, J.G.; Kleinjans, J.C.S.; Prickaerts, J.; Briedé, J.J. Circulating MicroRNAs as Potential Biomarkers for Psychiatric and Neurodegenerative Disorders. Prog. Neurobiol. 2020, 185, 101732. [Google Scholar] [CrossRef]
- Alongi, P.; Sardina, D.S.; Coppola, R.; Scalisi, S.; Puglisi, V.; Arnone, A.; Raimondo, G.D.; Munerati, E.; Alaimo, V.; Midiri, F.; et al. 18F-Florbetaben PET/CT to Assess Alzheimer’s Disease: A New Analysis Method for Regional Amyloid Quantification. J. Neuroimaging 2019, 29, 383–393. [Google Scholar] [CrossRef]
- Ciaccio, M.; Lo Sasso, B.; Scazzone, C.; Gambino, C.M.; Ciaccio, A.M.; Bivona, G.; Piccoli, T.; Giglio, R.V.; Agnello, L. COVID-19 and Alzheimer’s Disease. Brain Sci. 2021, 11, 305. [Google Scholar] [CrossRef]
- Bonanni, L.; Cagnin, A.; Agosta, F.; Babiloni, C.; Borroni, B.; Bozzali, M.; Bruni, A.C.; Filippi, M.; Galimberti, D.; Monastero, R.; et al. The Italian Dementia with Lewy Bodies Study Group (DLB-SINdem): Toward a Standardization of Clinical Procedures and Multicenter Cohort Studies Design. Neurol. Sci. Off. J. Ital. Neurol. Soc. Ital. Soc. Clin. Neurophysiol. 2017, 38, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Wingo, T.S.; Yang, J.; Fan, W.; Min Canon, S.; Gerasimov, E.S.; Lori, A.; Logsdon, B.; Yao, B.; Seyfried, N.T.; Lah, J.J.; et al. Brain MicroRNAs Associated with Late-Life Depressive Symptoms Are Also Associated with Cognitive Trajectory and Dementia. NPJ Genom. Med. 2020, 5, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Mendes-Silva, A.P.; Pereira, K.S.; Tolentino-Araujo, G.T.; Nicolau Ede, S.; Silva-Ferreira, C.M.; Teixeira, A.L.; Diniz, B.S. Shared Biologic Pathways Between Alzheimer Disease and Major Depression: A Systematic Review of MicroRNA Expression Studies. Am. J. Geriatr. Psychiatry Off. J. Am. Assoc. Geriatr. Psychiatry 2016, 24, 903–912. [Google Scholar] [CrossRef]
- Mendes-Silva, A.P.; Diniz, B.S.; Araújo, G.T.T.; de Souza Nicolau, E.; Pereira, K.S.; Ferreira, C.M.S.; Barroso, L.S. Mirnas and Their Role in the Correlation between Major Depressive Disorder, Mild Cognitive Impairment and Alzheimer’s Disease. Alzheimer’s Dement. 2017, 13, P1017–P1018. [Google Scholar] [CrossRef]
- Yi, L.-T.; Mu, R.-H.; Dong, S.-Q.; Wang, S.-S.; Li, C.-F.; Geng, D.; Liu, Q. MiR-124 Antagonizes the Antidepressant-like Effects of Standardized Gypenosides in Mice. J. Psychopharmacol. 2018, 32, 458–468. [Google Scholar] [CrossRef]
- Baudry, A.; Mouillet-Richard, S.; Schneider, B.; Launay, J.M.; Kellermann, O. MiR-16 Targets the Serotonin Transporter: A New Facet for Adaptive Responses to Antidepressants. Science 2010, 329, 1537–1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aten, S.; Page, C.E.; Kalidindi, A.; Wheaton, K.; Niraula, A.; Godbout, J.P.; Hoyt, K.R.; Obrietan, K. MiR-132/212 Is Induced by Stress and Its Dysregulation Triggers Anxiety-Related Behavior. Neuropharmacology 2019, 144, 256. [Google Scholar] [CrossRef] [PubMed]
- Ryan, K.M.; O’Donovan, S.M.; McLoughlin, D.M. Electroconvulsive Stimulation Alters Levels of BDNF-Associated MicroRNAs. Neurosci. Lett. 2013, 549, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Lopizzo, N.; Zonca, V.; Cattane, N.; Pariante, C.M.; Cattaneo, A. MiRNAs in Depression Vulnerability and Resilience: Novel Targets for Preventive Strategies. J. Neural Transm. 2019, 126, 1241–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiori, L.M.; Lopez, J.P.; Richard-Devantoy, S.; Berlim, M.; Chachamovich, E.; Jollant, F.; Foster, J.; Rotzinger, S.; Kennedy, S.H.; Turecki, G. Investigation of MiR-1202, MiR-135a, and MiR-16 in Major Depressive Disorder and Antidepressant Response. Int. J. Neuropsychopharmacol. 2017, 20, 619–623. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.K.; Tyryshkin, K.; Elmi, N.; Dharsee, M.; Evans, K.R.; Good, J.; Javadi, M.; McCormack, S.; Vaccarino, A.L.; Zhang, X.; et al. Plasma MicroRNA Expression Levels and Their Targeted Pathways in Patients with Major Depressive Disorder Who Are Responsive to Duloxetine Treatment. J. Psychiatr. Res. 2019, 110, 38–44. [Google Scholar] [CrossRef]
- Miao, N.; Jin, J.; Kim, S.-N.; Sun, T. Hippocampal MicroRNAs Respond to Administration of Antidepressant Fluoxetine in Adult Mice. Int. J. Mol. Sci. 2018, 19, 671. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.C.; Tsai, M.C.; Lee, C.T.; Sun, M.H.; Huang, T.L. Antidepressant Treatment Increased Serum MiR-183 and MiR-212 Levels in Patients with Major Depressive Disorder. Psychiatry Res. 2018, 270, 232–237. [Google Scholar] [CrossRef] [PubMed]
- Camkurt, M.A.; Acar, Ş.; Coşkun, S.; Güneş, M.; Güneş, S.; Yılmaz, M.F.; Görür, A.; Tamer, L. Comparison of Plasma MicroRNA Levels in Drug Naive, First Episode Depressed Patients and Healthy Controls. J. Psychiatr. Res. 2015, 69, 67–71. [Google Scholar] [CrossRef]
- Hansen, K.F.; Obrietan, K. MicroRNA as Therapeutic Targets for Treatment of Depression. Neuropsychiatr. Dis. Treat. 2013, 9, 1011–1021. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.A.; Moeng, S.; Sim, S.; Kuh, H.J.; Choi, S.Y.; Park, J.K. MicroRNA-Based Combinatorial Cancer Therapy: Effects of MicroRNAs on the Efficacy of Anti-Cancer Therapies. Cells 2019, 9, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laffont, B.; Rayner, K.J. MicroRNAs in the Pathobiology and Therapy of Atherosclerosis. Can. J. Cardiol. 2017, 33, 313–324. [Google Scholar] [CrossRef] [Green Version]
- Krützfeldt, J. Strategies to Use MicroRNAs as Therapeutic Targets. Best Pract. Res. Clin. Endocrinol. Metab. 2016, 30, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Raposo, G.; Stoorvogel, W. Extracellular Vesicles: Exosomes, Microvesicles, and Friends. J. Cell Biol. 2013, 200, 373–383. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; Chen, J.; Huang, Z. Recent Progress in MicroRNA-Based Delivery Systems for the Treatment of Human Disease. ExRNA 2019, 1, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Dasgupta, I.; Chatterjee, A. Recent Advances in MiRNA Delivery Systems. Methods Protoc. 2021, 4, 10. [Google Scholar] [CrossRef]
- Wei, Y.B.; Liu, J.J.; Villaescusa, J.C.; Åberg, E.; Brené, S.; Wegener, G.; Mathé, A.A.; Lavebratt, C. Elevation of Il6 Is Associated with Disturbed Let-7 Biogenesis in a Genetic Model of Depression. Transl. Psychiatry 2016, 6, e869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.Y.; Chen, F.; Xiao, P.; Zhang, X.M.; Gao, X.X. Silence of MiR-9 Protects Depression Mice through Notch Signaling Pathway. Eur. Rev. Med Pharmacol. Sci. 2019, 23, 4961–4970. [Google Scholar] [CrossRef] [PubMed]
- Alberi, L.; Hoey, S.E.; Brai, E.; Scotti, A.L.; Marathe, S. Notch Signaling in the Brain: In Good and Bad Times. Ageing Res. Rev. 2013, 12, 801–814. [Google Scholar] [CrossRef] [Green Version]
- Lasky, J.L.; Wu, H. Notch Signaling, Brain Development, and Human Disease. Pediatric Res. 2005, 57, 104–109. [Google Scholar] [CrossRef]
- Su, M.; Hong, J.; Zhao, Y.; Liu, S.; Xue, X. MeCP2 Controls Hippocampal Brain-Derived Neurotrophic Factor Expression via Homeostatic Interactions with MicroRNA-132 in Rats with Depression. Mol. Med. Rep. 2015, 12, 5399–5406. [Google Scholar] [CrossRef] [Green Version]
- Wanet, A.; Tacheny, A.; Arnould, T.; Renard, P. MiR-212/132 Expression and Functions: Within and beyond the Neuronal Compartment. Nucleic Acids Res. 2012, 40, 4742–4753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Li, S.; Yan, J.; Wang, D.; Yin, R.; Zhao, L.; Zhu, Y.; Zhu, X. MiR-182 (MicroRNA-182) Suppression in the Hippocampus Evokes Antidepressant-like Effects in Rats. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2016, 65, 96–103. [Google Scholar] [CrossRef]
- Cao, M.Q.; Chen, D.H.; Zhang, C.H.; Wu, Z.Z. Screening of Specific MicroRNA in Hippocampus of Depression Model Rats and Intervention Effect of Chaihu Shugan San. China J. Chin. Mater. Med. 2013, 1585–1589. [Google Scholar]
- Terasawa, K.; Ichimura, A.; Sato, F.; Shimizu, K.; Tsujimoto, G. Sustained Activation of ERK1/2 by NGF Induces MicroRNA-221 and 222 in PC12 Cells. FEBS J. 2009, 276, 3269–3276. [Google Scholar] [CrossRef]
- Zhou, R.; Yuan, P.; Wang, Y.; Hunsberger, J.G.; Elkahloun, A.; Wei, Y.; Damschroder-Williams, P.; Du, J.; Chen, G.; Manji, H.K. Evidence for Selective MicroRNAs and Their Effectors as Common Long-Term Targets for the Actions of Mood Stabilizers. Neuropsychopharmacol. Off. Publ. Am. Coll. Neuropsychopharmacol. 2009, 34, 1395. [Google Scholar] [CrossRef]
- Stubbs, B.; Vancampfort, D.; Rosenbaum, S.; Firth, J.; Cosco, T.; Veronese, N.; Salum, G.A.; Schuch, F.B. An Examination of the Anxiolytic Effects of Exercise for People with Anxiety and Stress-Related Disorders: A Meta-Analysis. Psychiatry Res. 2017, 249, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Qu, H.; Liu, R.; Chen, J.; Zheng, L.; Chen, R. Aerobic Exercise Inhibits CUMS-Depressed Mice Hippocampal Inflammatory Response via Activating Hippocampal MiR-223/TLR4/MyD88-NF-ΚB Pathway. Int. J. Environ. Res. Public Health 2020, 17, 2676. [Google Scholar] [CrossRef] [PubMed]
- Eyre, H.A.; Papps, E.; Baune, B.T. Treating Depression and Depression-Like Behavior with Physical Activity: An Immune Perspective. Front. Psychiatry 2013, 4, 3. [Google Scholar] [CrossRef] [Green Version]
- Svensson, M.; Lexell, J.; Deierborg, T. Effects of Physical Exercise on Neuroinflammation, Neuroplasticity, Neurodegeneration, and Behavior: What We Can Learn From Animal Models in Clinical Settings. Neurorehabilit. Neural Repair 2014, 29, 577–589. [Google Scholar] [CrossRef]
- Soares, E.; Reis, J.; Rodrigues, M.; Ribeiro, C.F.; Pereira, F.C. Circulating Extracellular Vesicles: The Missing Link between Physical Exercise and Depression Management? Int. J. Mol. Sci. 2021, 22, 542. [Google Scholar] [CrossRef]
- Truettner, J.S.; Motti, D.; Dietrich, W.D. MicroRNA Overexpression Increases Cortical Neuronal Vulnerability to Injury. Brain Res. 2013, 1533, 122–130. [Google Scholar] [CrossRef] [Green Version]
- Brites, D.; Fernandes, A. Neuroinflammation and Depression: Microglia Activation, Extracellular Microvesicles and MicroRNA Dysregulation. Front. Cell. Neurosci. 2015, 9, 476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chieffi, S.; Messina, G.; Villano, I.; Messina, A.; Esposito, M.; Monda, V.; Valenzano, A.; Moscatelli, F.; Esposito, T.; Carotenuto, M.; et al. Exercise Influence on Hippocampal Function: Possible Involvement of Orexin-A. Front. Physiol. 2017, 8, 85. [Google Scholar] [CrossRef] [Green Version]
- Guerrera, C.S.; Furneri, G.; Grasso, M.; Caruso, G.; Castellano, S.; Drago, F.; di Nuovo, S.; Caraci, F. Antidepressant Drugs and Physical Activity: A Possible Synergism in the Treatment of Major Depression? Front. Psychol. 2020, 11, 857. [Google Scholar] [CrossRef]
- Tolahunase, M.R.; Gautam, S.; Sagar, R.; Kumar, M.; Dada, R. Yoga in Major Depressive Disorder: Molecular Mechanisms and Clinical Utility. Front. Biosci. Sch. 2021, 13, 56–81. [Google Scholar] [CrossRef]
- Wang, X.; Sundquist, K.; Palmér, K.; Hedelius, A.; Memon, A.A.; Sundquist, J. Macrophage Migration Inhibitory Factor and MicroRNA-451a in Response to Mindfulness-Based Therapy or Treatment as Usual in Patients with Depression, Anxiety, or Stress and Adjustment Disorders. Int. J. Neuropsychopharmacol. 2018, 21, 513–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashizume, S.; Nakano, M.; Kubota, K.; Kobayashi, E.; Fujimiya, M. Mindfulness Intervention Improves Cognitive Function in Older Adults by Enhancing the Level of MiRNA-29c in Neuron-Derived Exosomes. Sci. Rep. 2021, 11, 21848. [Google Scholar] [CrossRef]
- González, S. Dietary Bioactive Compounds and Human Health and Disease. Nutrients 2020, 12, 348. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Zeng, Y.; Zeng, D.; Wang, H.; Zhou, M.; Sun, N.; Xin, J.; Khalique, A.; Rajput, D.S.; Pan, K.; et al. Probiotics and MicroRNA: Their Roles in the Host–Microbe Interactions. Front. Microbiol. 2021, 11, 3363. [Google Scholar] [CrossRef] [PubMed]
- Xiao, P.; Zhang, X.; Li, Y.; Ma, Z.; Si, S.; Gao, X. MiR-9 Inhibition of Neuronal Apoptosis and Expression Levels of Apoptosis Genes Bcl-2 and Bax in Depression Model Rats through Notch Pathway. Exp. Ther. Med. 2020, 19, 551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maffioletti, E.; Cattaneo, A.; Rosso, G.; Maina, G.; Maj, C.; Gennarelli, M.; Tardito, D.; Bocchio-Chiavetto, L. Peripheral Whole Blood MicroRNA Alterations in Major Depression and Bipolar Disorder. J. Affect. Disord. 2016, 200, 250–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roshan, R.; Shridhar, S.; Sarangdhar, M.A.; Banik, A.; Chawla, M.; Garg, M.; Singh, V.P.; Pillai, B. Brain-Specific Knockdown of MiR-29 Results in Neuronal Cell Death and Ataxia in Mice. RNA 2014, 20, 1287–1297. [Google Scholar] [CrossRef] [Green Version]
- Sun, N.; Lei, L.; Wang, Y.; Yang, C.; Liu, Z.; Li, X.; Zhang, K. Preliminary Comparison of Plasma Notch-Associated MicroRNA-34b and -34c Levels in Drug Naive, First Episode Depressed Patients and Healthy Controls. J. Affect. Disord. 2016, 194, 109–114. [Google Scholar] [CrossRef]
- Fang, Y.; Qiu, Q.; Zhang, S.; Sun, L.; Li, G.; Xiao, S.; Li, X. Changes in MiRNA-132 and MiR-124 Levels in Non-Treated and Citalopram-Treated Patients with Depression. J. Affect. Disord. 2018, 227, 745–751. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Liu, X.; Jiang, K.; Peng, D.; Hong, W.; Fang, Y.; Qian, Y.; Yu, S.; Li, H. Alterations of MicroRNA-124 Expression in Peripheral Blood Mononuclear Cells in Pre- and Post-Treatment Patients with Major Depressive Disorder. J. Psychiatr. Res. 2016, 78, 65–71. [Google Scholar] [CrossRef]
- Qi, S.; Yang, X.; Zhao, L.; Calhoun, V.D.; Perrone-Bizzozero, N.; Liu, S.; Jiang, R.; Jiang, T.; Sui, J.; Ma, X. MicroRNA132 Associated Multimodal Neuroimaging Patterns in Unmedicated Major Depressive Disorder. Brain 2018, 141, 916–926. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Yu, D.; Gu, Q.H.; Yang, Y.; Tu, K.; Zhu, J.; Li, Z. MiR-191 and MiR-135 Are Required for Long-Lasting Spine Remodelling Associated with Synaptic Long-Term Depression. Nat. Commun. 2014, 5, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Li, H.; Guo, R.; Ma, T.; Hou, R.; Ma, X.; Du, Y. MiR-137, a New Target for Post-Stroke Depression? Neural Regen. Res. 2013, 8, 2441. [Google Scholar] [CrossRef] [PubMed]
- Mahmoudi, E.; Cairns, M.J. MiR-137: An Important Player in Neural Development and Neoplastic Transformation. Mol. Psychiatry 2017, 22, 44–55. [Google Scholar] [CrossRef] [Green Version]
- Woodbury, M.E.; Freilich, R.W.; Cheng, C.J.; Asai, H.; Ikezu, S.; Boucher, J.D.; Slack, F.; Ikezu, T. MiR-155 Is Essential for Inflammation-Induced Hippocampal Neurogenic Dysfunction. J. Neurosci. 2015, 35, 9764–9781. [Google Scholar] [CrossRef] [Green Version]
- Kohen, R.; Dobra, A.; Tracy, J.H.; Haugen, E. Transcriptome Profiling of Human Hippocampus Dentate Gyrus Granule Cells in Mental Illness. Transl. Psychiatry 2014, 4, e366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azevedo, J.A.; Carter, B.S.; Meng, F.; Turner, D.L.; Dai, M.; Schatzberg, A.F.; Barchas, J.D.; Jones, E.G.; Bunney, W.E.; Myers, R.M.; et al. The MicroRNA Network Is Altered in Anterior Cingulate Cortex of Patients with Unipolar and Bipolar Depression. J. Psychiatr. Res. 2016, 82, 58–67. [Google Scholar] [CrossRef] [Green Version]
- Belzeaux, R.; Bergon, A.; Jeanjean, V.; Loriod, B.; Formisano-Tréziny, C.; Verrier, L.; Loundou, A.; Baumstarck-Barrau, K.; Boyer, L.; Gall, V.; et al. Responder and Nonresponder Patients Exhibit Different Peripheral Transcriptional during Major Depressive Episode. Transl. Psychiatry 2012, 2, e185. [Google Scholar] [CrossRef]
- Oved, K.; Morag, A.; Pasmanik-Chor, M.; Oron-Karni, V.; Shomron, N.; Rehavi, M.; Stingl, J.C.; Gurwitz, D. Genome-Wide MiRNA Expression Profiling of Human Lymphoblastoid Cell Lines Identifies Tentative SSRI Antidepressant Response Biomarkers. Pharmacogenomics 2012, 13, 1129–1139. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, Y.; Roy, B.; Lugli, G.; Rizavi, H.; Zhang, H.; Smalheiser, N.R. Chronic Corticosterone-Mediated Dysregulation of MicroRNA Network in Prefrontal Cortex of Rats: Relevance to Depression Pathophysiology. Transl. Psychiatry 2015, 5, e682. [Google Scholar] [CrossRef]
- Fiori, L.M.; Kos, A.; Lin, R.; Théroux, J.F.; Lopez, J.P.; Kühne, C.; Eggert, C.; Holzapfel, M.; Huettl, R.E.; Mechawar, N.; et al. MiR-323a Regulates ERBB4 and Is Involved in Depression. Mol. Psychiatry 2020, 1–14. [Google Scholar] [CrossRef]
- Zucchi, F.C.; Yao, Y.; Ward, I.D.; Ilnytskyy, Y.; Olson, D.M.; Benzies, K.; Kovalchuk, I.; Kovalchuk, O.; Metz, G.A. Maternal Stress Induces Epigenetic Signatures of Psychiatric and Neurological Diseases in the Offspring. PLoS ONE 2013, 8, e0056967. [Google Scholar] [CrossRef] [PubMed]
- Ciernia, A.V.; Laufer, B.I.; Dunaway, K.W.; Mordaunt, C.E.; Coulson, R.L.; Totah, T.S.; Stolzenberg, D.S.; Frahm, J.C.; Singh-Taylor, A.; Baram, T.Z.; et al. Experience-Dependent Neuroplasticity of the Developing Hypothalamus: Integrative Epigenomic Approaches. Epigenetics 2018, 13, 318. [Google Scholar] [CrossRef] [Green Version]
- Smalheiser, N.R.; Lugli, G.; Rizavi, H.S.; Zhang, H.; Torvik, V.I.; Pandey, G.N.; Davis, J.M.; Dwivedi, Y. MicroRNA Expression in Rat Brain Exposed to Repeated Inescapable Shock: Differential Alterations in Learned Helplessness vs. Non-Learned Helplessness. Int. J. Neuropsychopharmacol. 2011, 14, 1315–1325. [Google Scholar] [CrossRef]
- Balakathiresan, N.S.; Chandran, R.; Bhomia, M.; Jia, M.; Li, H.; Maheshwari, R.K. Serum and Amygdala MicroRNA Signatures of Posttraumatic Stress: Fear Correlation and Biomarker Potential. J. Psychiatr. Res. 2014, 57, 65–73. [Google Scholar] [CrossRef]
- Garbett, K.A.; Vereczkei, A.; Kálmán, S.; Brown, J.A.; Taylor, W.D.; Faludi, G.; Korade, Ž.; Shelton, R.C.; Mirnics, K. Coordinated Messenger RNA/MicroRNA Changes in Fibroblasts of Patients with Major Depression. Biol. Psychiatry 2015, 77, 256–265. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortega, M.A.; Alvarez-Mon, M.A.; García-Montero, C.; Fraile-Martinez, O.; Lahera, G.; Monserrat, J.; Muñoz-Merida, L.; Mora, F.; Rodríguez-Jiménez, R.; Fernandez-Rojo, S.; et al. MicroRNAs as Critical Biomarkers of Major Depressive Disorder: A Comprehensive Perspective. Biomedicines 2021, 9, 1659. https://doi.org/10.3390/biomedicines9111659
Ortega MA, Alvarez-Mon MA, García-Montero C, Fraile-Martinez O, Lahera G, Monserrat J, Muñoz-Merida L, Mora F, Rodríguez-Jiménez R, Fernandez-Rojo S, et al. MicroRNAs as Critical Biomarkers of Major Depressive Disorder: A Comprehensive Perspective. Biomedicines. 2021; 9(11):1659. https://doi.org/10.3390/biomedicines9111659
Chicago/Turabian StyleOrtega, Miguel A., Miguel Angel Alvarez-Mon, Cielo García-Montero, Oscar Fraile-Martinez, Guillermo Lahera, Jorge Monserrat, Luis Muñoz-Merida, Fernando Mora, Roberto Rodríguez-Jiménez, Sonia Fernandez-Rojo, and et al. 2021. "MicroRNAs as Critical Biomarkers of Major Depressive Disorder: A Comprehensive Perspective" Biomedicines 9, no. 11: 1659. https://doi.org/10.3390/biomedicines9111659
APA StyleOrtega, M. A., Alvarez-Mon, M. A., García-Montero, C., Fraile-Martinez, O., Lahera, G., Monserrat, J., Muñoz-Merida, L., Mora, F., Rodríguez-Jiménez, R., Fernandez-Rojo, S., Quintero, J., & Álvarez-Mon, M. (2021). MicroRNAs as Critical Biomarkers of Major Depressive Disorder: A Comprehensive Perspective. Biomedicines, 9(11), 1659. https://doi.org/10.3390/biomedicines9111659