
Differential Modulation of Matrix Metalloproteinases-2 and -7 in LAM/TSC Cells

 , ,
, ,  ,
,  and
and 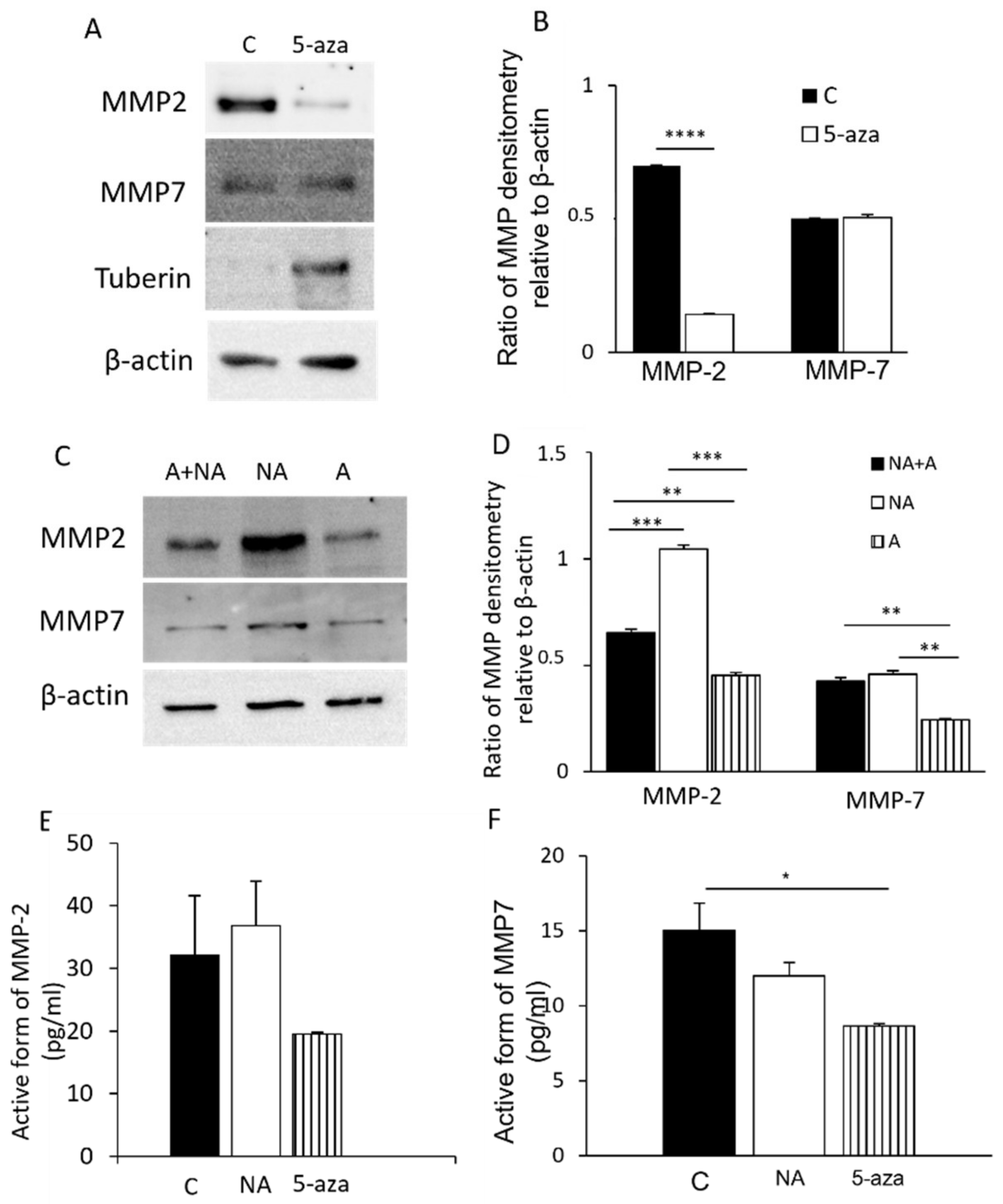{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Treatments
2.2. Western Blot Analysis
2.3. ELISA
2.4. Wound Healing Assay
2.5. Flow Cytometric Analysis
2.6. Animal Experiments and Pharmacological Treatments
2.7. Immunohistochemical Analysis
2.8. MMP-2 and MMP-7 Activity Assay
2.9. Statistical Analysis
3. Results
3.1. MMP-2 and MMP-7 Expression in LAM/TSC Cells
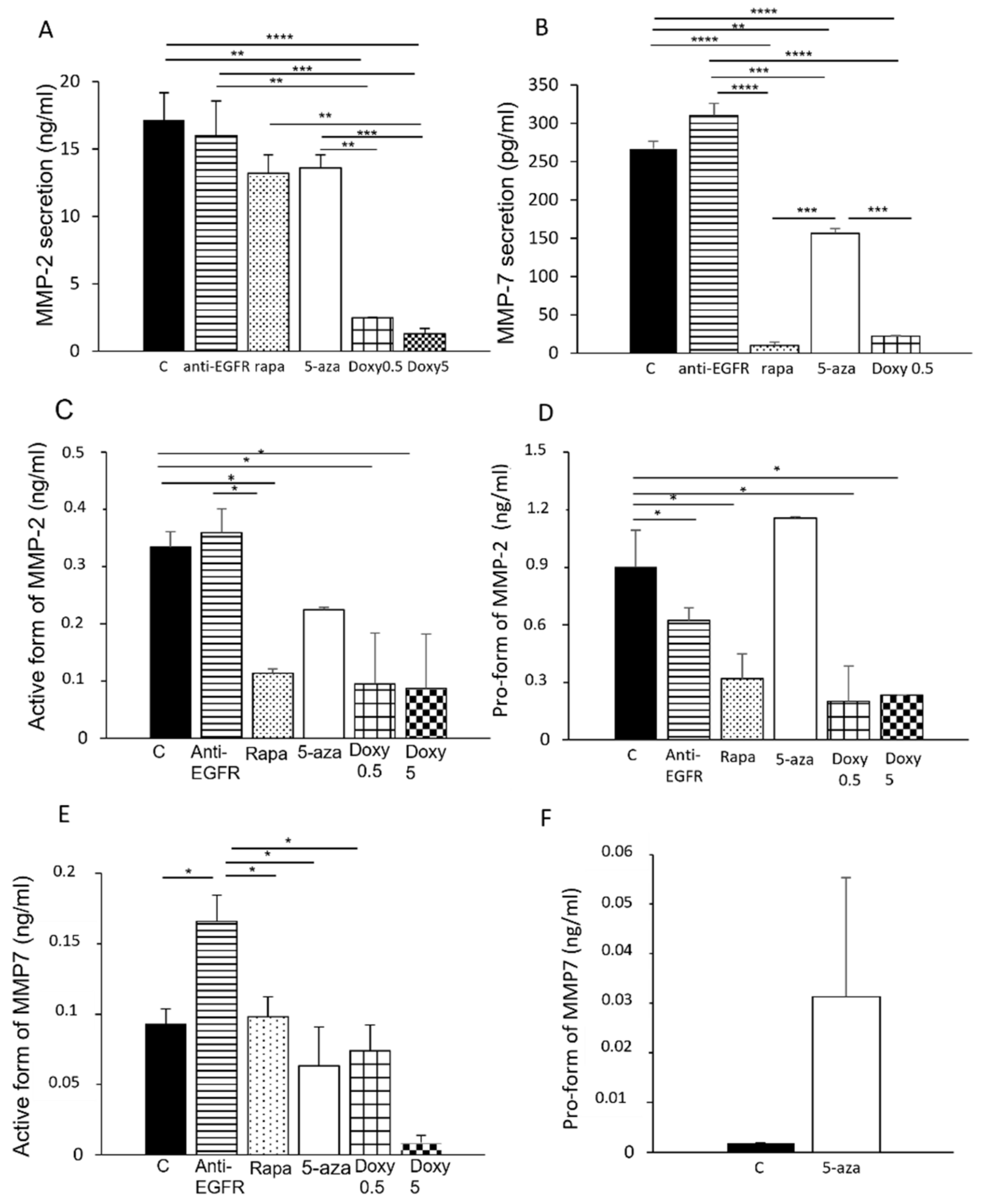
3.2. Modulation of MMP-2 and MMP-7 Expression with Drugs
3.3. Secretion of MMP-2 and MMP-7 in LAM/TSC Cells
3.4. Influence of MMP-2 and MMP-7 Secretion on LAM/TSC Cell Migration
3.5. Secretion of MMP-2 and MMP-7 during LAM/TSC Cell Migration
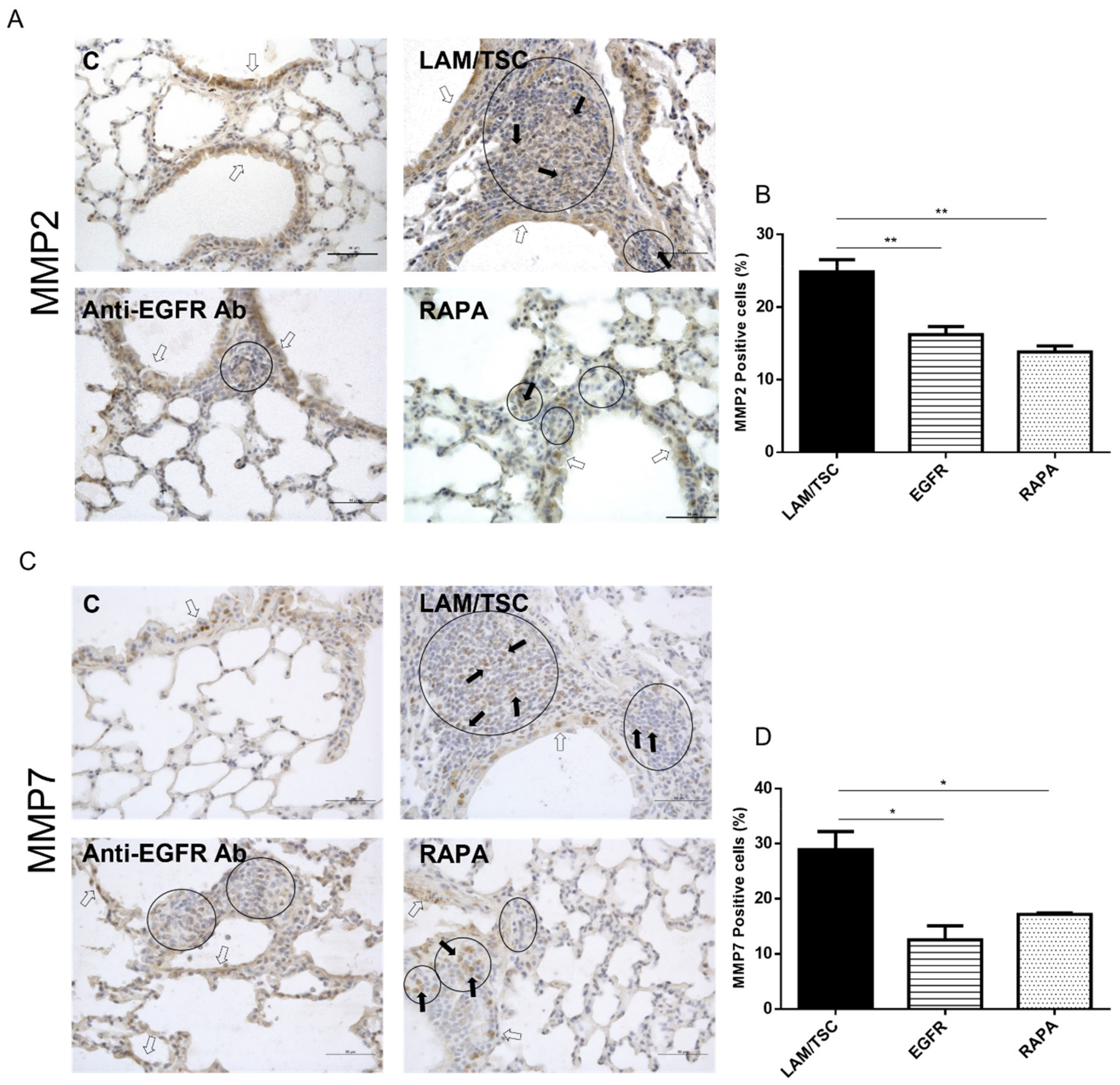
3.6. Expression of MMP-2 and MMP7 in Lungs of a Mouse LAM Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| MMP | matrix metalloproteinases |
| LAM | lymphangioleiomyomatosis |
| TSC | tuberous sclerosis complex |
| ECM | extracellular matrix |
| EGF | epidermal growth factor |
| VEGF | vascular-endothelial growth factor |
References
- Khokha, R.; Murthy, A.; Weiss, A. Metalloproteinases and their natural inhibitors in inflammation and immunity. Nat. Rev. Immunol. 2013, 13, 649–665. [Google Scholar] [CrossRef] [PubMed]
- Kessenbrock, K.; Plaks, V.; Werb, Z. Matrix metalloproteinases: Regulators of the tumor microenvironment. Cell 2010, 141, 52–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, N.; Hu, M.; Khalil, R.A. Biochemical and Biological Attributes of Matrix Metalloproteinases. Prog. Mol. Biol. Transl. Sci. 2017, 147, 1–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, D.; Vetrivel, U.; Parameswaran, S.; Subramanian, K.K. Structural insights on druggable hotspots in CD147: A bull’s eye view. Life Sci. 2019, 224, 76–87. [Google Scholar] [CrossRef]
- Hu, X.; Su, J.; Zhou, Y.; Xie, X.; Peng, C.; Yuan, Z.; Chen, X. Repressing CD147 is a novel therapeutic strategy for malignant melanoma. Oncotarget 2017, 8, 25806–25813. [Google Scholar] [CrossRef] [Green Version]
- Greenlee, K.J.; Werb, Z.; Kheradmand, F. Matrix metalloproteinases in lung: Multiple, multifarious, and multifaceted. Physiol. Rev. 2007, 87, 69–98. [Google Scholar] [CrossRef]
- Hayashi, T.; Fleming, M.V.; Stetler-Stevenson, W.G.; Liotta, L.A.; Moss, J.; Ferrans, V.J.; Travis, W.D. Immunohistochemical study of matrix metalloproteinases (MMPs) and their tissue inhibitors (TIMPs) in pulmonary lymphangioleiomyomatosis (LAM). Hum. Pathol. 1997, 28, 1071–1078. [Google Scholar] [CrossRef]
- Juvet, S.C.; McCormack, F.X.; Kwiatkowski, D.J.; Downey, G.P. Molecular pathogenesis of lymphangioleiomyomatosis: Lessons learned from orphans. Am. J. Respir. Cell Mol. Biol. 2007, 36, 398–408. [Google Scholar] [CrossRef]
- Di Marco, F.; Terraneo, S.; Imeri, G.; Palumbo, G.; La Briola, F.; Tresoldi, S.; Volpi, A.; Gualandri, L.; Ghelma, F.; Alfano, R.M.; et al. Women with TSC: Relationship between Clinical, Lung Function and Radiological Features in a Genotyped Population Investigated for Lymphangioleiomyomatosis. PLoS ONE 2016, 11, e0155331. [Google Scholar] [CrossRef]
- McCormack, F.X.; Travis, W.D.; Colby, T.V.; Henske, E.P.; Moss, J. Lymphangioleiomyomatosis Calling It What It Is: A Low-Grade, Destructive, Metastasizing Neoplasm. Am. J. Respir. Crit. Care Med. 2012, 188, 1210–1212. [Google Scholar] [CrossRef]
- Johnson, S.R.; Taveira-DaSilva, A.M.; Moss, J. Lymphangioleiomyomatosis. Clin. Chest Med. 2016, 37, 389–403. [Google Scholar] [CrossRef] [Green Version]
- Bissler, J.J.; Kingswood, J.C.; Radzikowska, E.; Zonnenberg, B.A.; Frost, M.; Belousova, E.; Sauter, M.; Nonomura, N.; Brakemeier, S.; de Vries, P.J.; et al. Everolimus for angiomyolipoma associated with tuberous sclerosis complex or sporadic lymphangioleiomyomatosis (EXIST-2): A multicentre, randomised, double-blind, placebo-controlled trial. Lancet 2013, 381, 817–824. [Google Scholar] [CrossRef]
- Lesma, E.; Ancona, S.; Sirchia, S.M.; Orpianesi, E.; Grande, V.; Colapietro, P.; Chiaramonte, E.; Di Giulio, A.M.; Gorio, A. TSC2 epigenetic defect in primary LAM cells. Evidence of an anchorage-independent survival. J. Cell. Mol. Med. 2014, 18, 766–779. [Google Scholar] [CrossRef]
- Lesma, E.; Grande, V.; Carelli, S.; Brancaccio, D.; Canevini, M.P.; Alfano, R.M.; Coggi, G.; Di Giulio, A.M.; Gorio, A. Isolation and growth of smooth muscle-like cells derived from tuberous sclerosis complex-2 human renal angiomyolipoma: Epidermal growth factor is the required growth factor. Am. J. Pathol. 2005, 167, 1093–1103. [Google Scholar] [CrossRef]
- Lesma, E.; Sirchia, S.M.; Ancona, S.; Carelli, S.; Bosari, S.; Ghelma, F.; Montanari, E.; Di Giulio, A.M.; Gorio, A. The methylation of the TSC2 promoter underlies the abnormal growth of TSC2 angiomyolipoma-derived smooth muscle cells. Am. J. Pathol. 2009, 174, 2150–2159. [Google Scholar] [CrossRef] [Green Version]
- Lesma, E.; Chiaramonte, E.; Ancona, S.; Orpianesi, E.; Di Giulio, A.M.; Gorio, A. Anti-EGFR antibody reduces lung nodules by inhibition of EGFR-pathway in a model of lymphangioleiomyomatosis. Biomed. Res. Int. 2015, 2015, 315240. [Google Scholar] [CrossRef]
- Prizant, H.; Hammes, S.R. Minireview: Lymphangioleiomyomatosis (LAM): The “Other” Steroid-Sensitive Cancer. Endocrinology 2016, 157, 3374–3383. [Google Scholar] [CrossRef]
- Matsui, K.; Takeda, K.; Yu, Z.X.; Travis, W.D.; Moss, J.; Ferrans, V.J. Role for activation of matrix metalloproteinases in the pathogenesis of pulmonary lymphangioleiomyomatosis. Arch. Pathol. Lab. Med. 2000, 124, 267–275. [Google Scholar] [CrossRef]
- Odajima, N.; Betsuyaku, T.; Nasuhara, Y.; Inoue, H.; Seyama, K.; Nishimura, M. Matrix metalloproteinases in blood from patients with LAM. Respir. Med. 2009, 103, 124–129. [Google Scholar] [CrossRef] [Green Version]
- Terraneo, S.; Lesma, E.; Ancona, S.; Imeri, G.; Palumbo, G.; Torre, O.; Giuliani, L.; Centanni, S.; Peron, A.; Tresoldi, S.; et al. Exploring the Role of Matrix Metalloproteinases as Biomarkers in Sporadic Lymphangioleiomyomatosis and Tuberous Sclerosis Complex. A Pilot Study. Front. Med. 2021, 8, 605909. [Google Scholar] [CrossRef]
- Moses, M.A.; Harper, J.; Folkman, J. Doxycycline treatment for lymphangioleiomyomatosis with urinary monitoring for MMPs. N. Engl. J. Med. 2006, 354, 2621–2622. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.S.; Tsang, S.W.; Moses, M.A.; Trayes-Gibson, Z.; Hsiao, L.L.; Jensen, R.; Squillace, R.; Kwiatkowski, D.J. Rapamycin-insensitive up-regulation of MMP2 and other genes in tuberous sclerosis complex 2-deficient lymphangioleiomyomatosis-like cells. Am. J. Respir. Cell Mol. Biol. 2010, 42, 227–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomes, C.; Cunha, C.; Nascimento, F.; Ribeiro, J.A.; Vaz, A.R.; Brites, D. Cortical Neurotoxic Astrocytes with Early ALS Pathology and miR-146a Deficit Replicate Gliosis Markers of Symptomatic SOD1G93A Mouse Model. Mol. Neurobiol. 2019, 56, 2137–2158. [Google Scholar] [CrossRef] [PubMed]
- Fields, G.B. The Rebirth of Matrix Metalloproteinase Inhibitors: Moving Beyond the Dogma. Cells 2019, 8, 984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moir, L.M.; Ng, H.Y.; Poniris, M.H.; Santa, T.; Burgess, J.K.; Oliver, B.G.G.; Krymskaya, V.P.; Black, J.L. Doxycycline inhibits matrix metalloproteinase-2 secretion from TSC2-null mouse embryonic fibroblasts and lymphangioleiomyomatosis cells. Br. J. Pharmacol. 2011, 164, 83–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pimenta, S.P.; Baldi, B.G.; Kairalla, R.A.; Carvalho, C.R. Doxycycline use in patients with lymphangioleiomyomatosis: Biomarkers and pulmonary function response. J. Bras. Pneumol. 2013, 39, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.Y.; Cane, J.L.; Kumaran, M.; Lewis, S.; Tattersfield, A.E.; Johnson, S.R. A two year randomised placebo controlled trial of doxycycline for lymphangioleiomyomatosis. Eur. Respir. J. 2014, 43, 1114–1123. [Google Scholar] [CrossRef] [Green Version]
- McCormack, F.X.; Gupta, N.; Finlay, G.R.; Young, L.R.; Taveira-DaSilva, A.M.; Glasgow, C.G.; Steagall, W.K.; Johnson, S.R.; Sahn, S.A.; Ryu, J.H.; et al. ATS/JRS Committee on Lymphangioleiomyomatosis Official American Thoracic Society/Japanese Respiratory Society Clinical Practice Guidelines: Lymphangioleiomyomatosis Diagnosis and Management. Am. J. Respir. Crit. Care Med. 2016, 194, 748–761. [Google Scholar] [CrossRef] [Green Version]
- Parsons, S.L.; Watson, S.A.; Brown, P.D.; Collins, H.M.; Steele, R.J. Matrix metalloproteinases. Br. J. Surg. 1997, 84, 160–166. [Google Scholar]
- Ii, M.; Yamamoto, H.; Adachi, Y.; Maruyama, Y.; Shinomura, Y. Role of matrix metalloproteinase-7 (matrilysin) in human cancer invasion, apoptosis, growth, and angiogenesis. Exp. Biol. Med. 2006, 231, 20–27. [Google Scholar] [CrossRef]
- Liu, D.; Nakano, J.; Ishikawa, S.; Yokomise, H.; Ueno, M.; Kadota, K.; Urushihara, M.; Huang, C.L. Overexpression of matrix metalloproteinase-7 (MMP-7) correlates with tumor proliferation, and a poor prognosis in non-small cell lung cancer. Lung Cancer 2007, 58, 384–391. [Google Scholar] [CrossRef]
- Barnes, E.A.; Kenerson, H.L.; Mak, B.C.; Yeung, R.S. The loss of tuberin promotes cell invasion through the ß-catenin pathway. Am. J. Respir. Cell Mol. Biol. 2010, 43, 617–627. [Google Scholar] [CrossRef] [Green Version]
- Odajima, N.; Betsuyaku, T.; Yoshida, T.; Hosokawa, T.; Nishimura, M. High levels of extracellular matrix metalloproteinase inducer are expressed in lymphangioleiomyomatosis. Hum. Pathol. 2010, 41, 935–943. [Google Scholar] [CrossRef]
- Toole, B.P. Emmprin (CD147), a cell surface regulator of matrix metalloproteinase production and function. Curr. Top. Dev. Biol. 2003, 54, 371–389. [Google Scholar] [CrossRef]
- Gabison, E.E.; Mourah, S.; Steinfels, E.; Yan, L.; Hoang-Xuan, T.; Watsky, M.A.; De Wever, B.; Calvo, F.; Mauviel, A.; Menashi, S. Differential expression of extracellular matrix metalloproteinase inducer (CD147) in normal and ulcerated corneas: Role in epithelia-stromal interactions and matrix metalloproteinase induction. Am. J. Pathol. 2005, 166, 209–219. [Google Scholar] [CrossRef]
- Frederick, J.W.; Sweeny, L.; Hartman, Y.; Zhou, T.; Rosenthal, E.L. Epidermal growth factor receptor inhibition by anti-CD147 therapy in cutaneous squamous cell carcinoma. Head Neck 2016, 38, 247–252. [Google Scholar] [CrossRef]
- Gonchorova, E.A.; Goncharov, D.A.; Fehrenbach, M.; Khavin, I.; Ducka, B.; Hino, O.; Colby, T.V.; Merrilees, M.J.; Haczku, A.; Albelda, S.M.; et al. Prevention of alveolar destruction and airspace enlargement in a mouse model of pulmonary lymphangioleiomyomatosis (LAM). Sci. Transl. Med. 2012, 4, 154ra134. [Google Scholar] [CrossRef] [Green Version]
- Prizant, H.; Taya, M.; Lerman, I.; Light, A.; Sen, A.; Mitra, S.; Foster, T.H.; Hammes, S.R. Estrogen maintains myometrial tumors in a Lymphangioleiomyomatosis model. Endocr.-Relat. Cancer 2016, 23, 265–280. [Google Scholar] [CrossRef] [Green Version]
- Hadzic, S.; Wu, C.-Y.; Gredic, M.; Kojonazarov, B.; Pak, O.; Kraut, S.; Sommer, N.; Kosanovic, D.; Grimminger, F.; Schermuly, R.T.; et al. The effect of long-term doxycycline treatment in a mouse model of cigarette smoke-induced emphysema and pulmonary hypertension. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2021, 320, L903–L915. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ancona, S.; Orpianesi, E.; Bernardelli, C.; Chiaramonte, E.; Chiaramonte, R.; Terraneo, S.; Di Marco, F.; Lesma, E. Differential Modulation of Matrix Metalloproteinases-2 and -7 in LAM/TSC Cells. Biomedicines 2021, 9, 1760. https://doi.org/10.3390/biomedicines9121760
Ancona S, Orpianesi E, Bernardelli C, Chiaramonte E, Chiaramonte R, Terraneo S, Di Marco F, Lesma E. Differential Modulation of Matrix Metalloproteinases-2 and -7 in LAM/TSC Cells. Biomedicines. 2021; 9(12):1760. https://doi.org/10.3390/biomedicines9121760
Chicago/Turabian StyleAncona, Silvia, Emanuela Orpianesi, Clara Bernardelli, Eloisa Chiaramonte, Raffaella Chiaramonte, Silvia Terraneo, Fabiano Di Marco, and Elena Lesma. 2021. "Differential Modulation of Matrix Metalloproteinases-2 and -7 in LAM/TSC Cells" Biomedicines 9, no. 12: 1760. https://doi.org/10.3390/biomedicines9121760
APA StyleAncona, S., Orpianesi, E., Bernardelli, C., Chiaramonte, E., Chiaramonte, R., Terraneo, S., Di Marco, F., & Lesma, E. (2021). Differential Modulation of Matrix Metalloproteinases-2 and -7 in LAM/TSC Cells. Biomedicines, 9(12), 1760. https://doi.org/10.3390/biomedicines9121760