
Intravesical High Dose BCG Tokyo and Low Dose BCG Tokyo with GMCSF+IFN α Induce Systemic Immunity in a Murine Orthotopic Bladder Cancer Model

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plasmids and BCG
2.2. Effect of BCG Strains on Immune Cells In Vitro
2.3. In Vitro Stimulation of Tumor Cells and Macrophages with BCG Tokyo and IFNα
2.4. Orthotopic Tumor Model
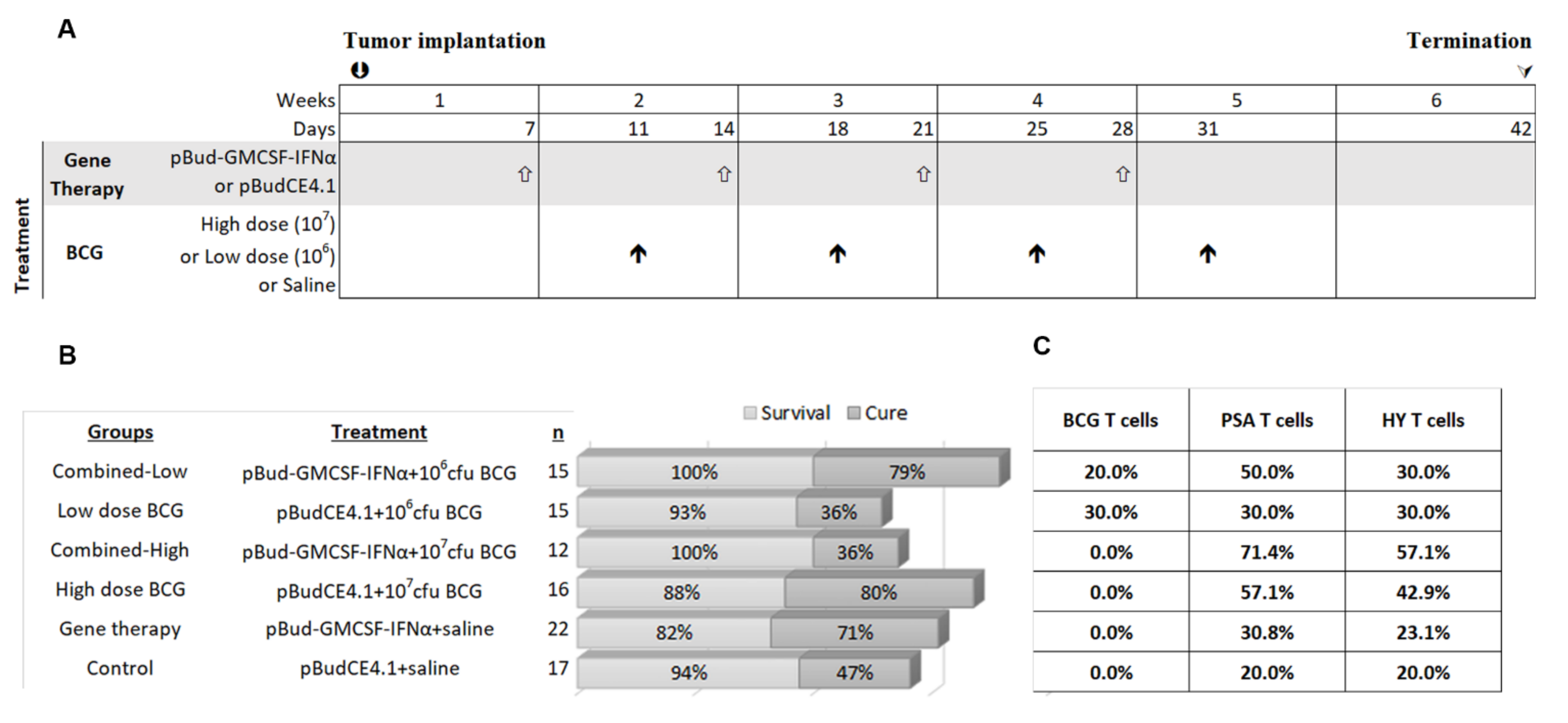
2.5. Treatment Groups
2.6. Flow Cytometry
2.7. Real-time PCR Analysis and RT-PCR
2.8. Statistical Analysis
3. Results
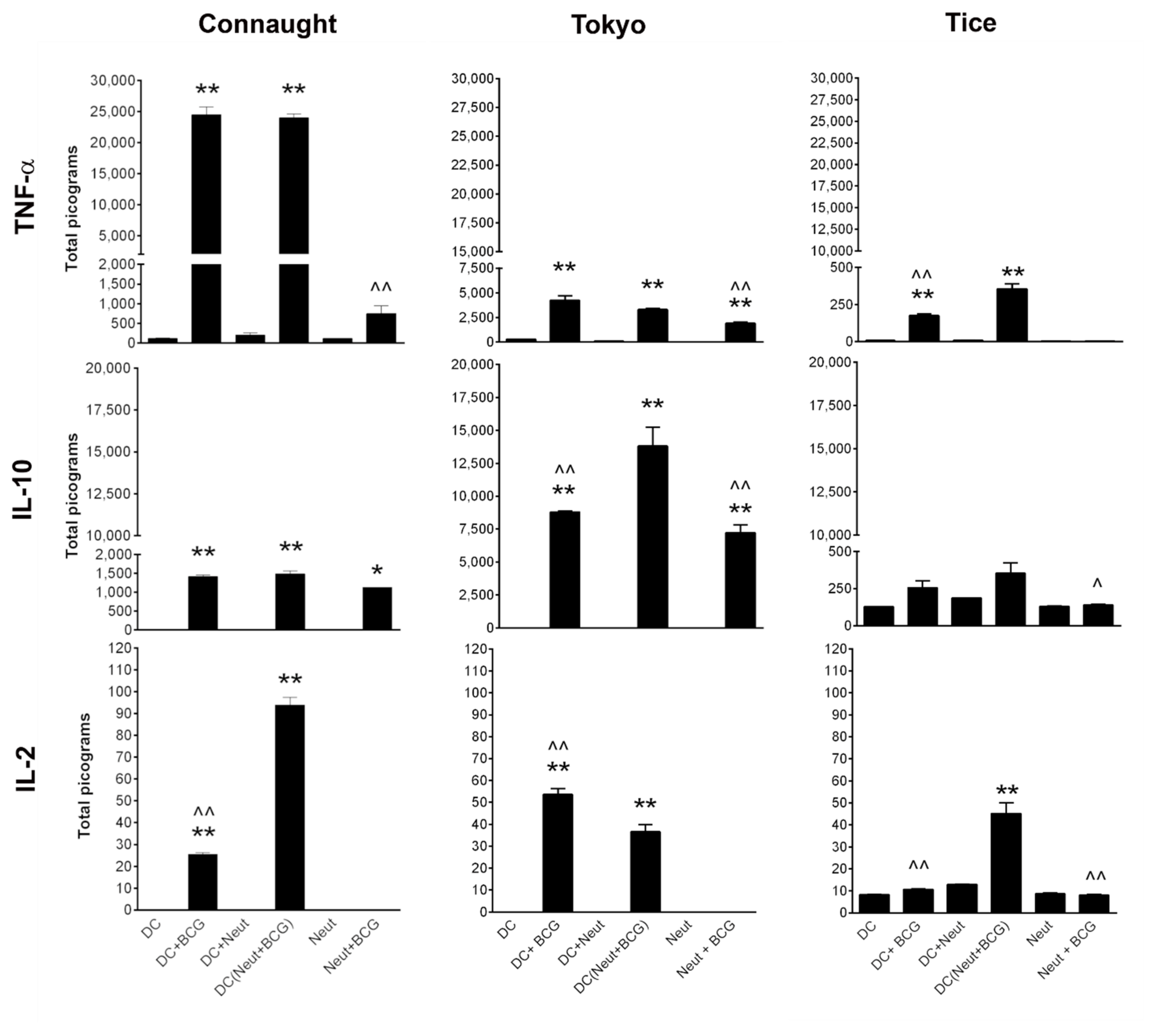
3.1. Comparison of Cytokine Induction by the Three BCG Strains
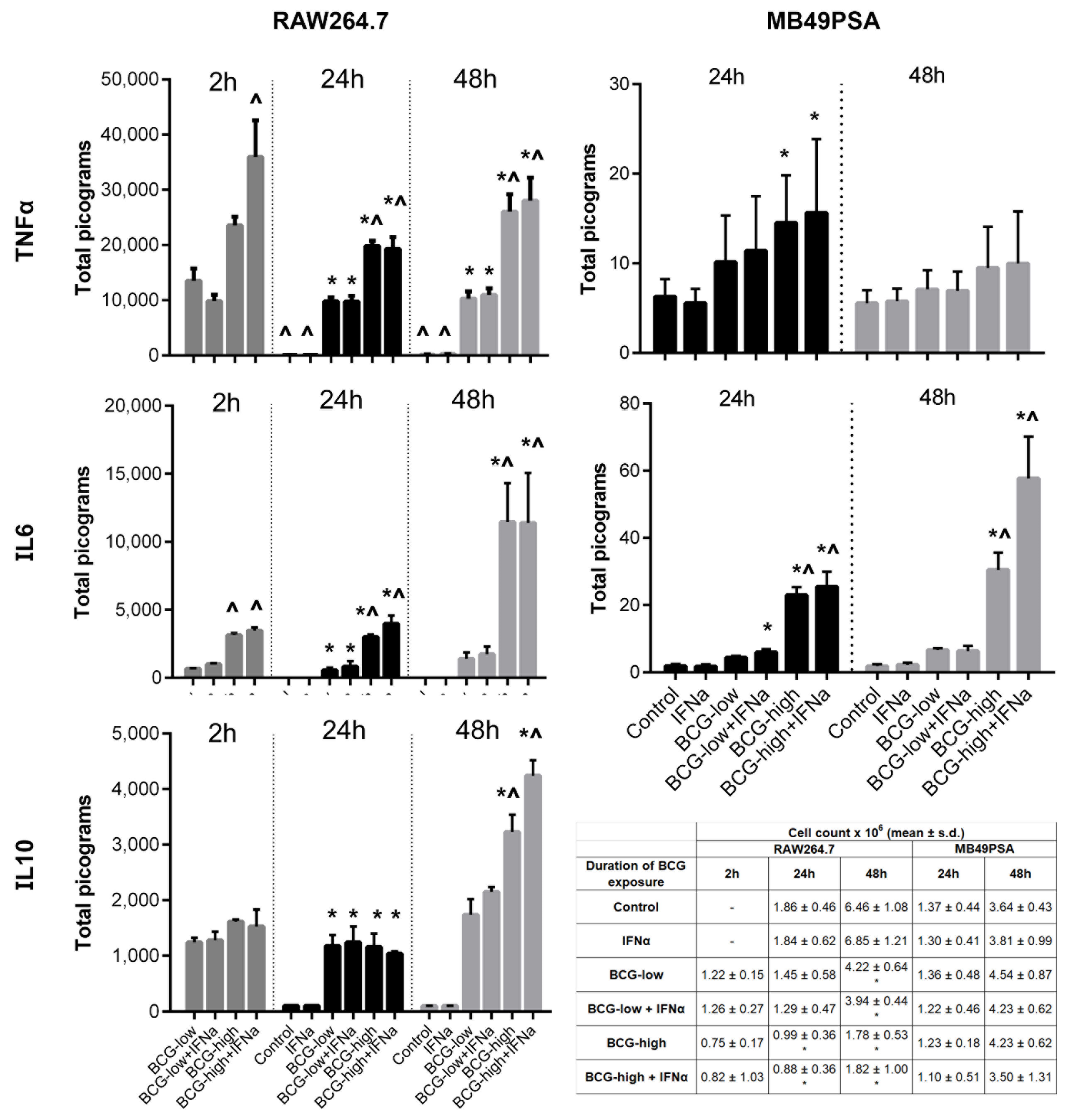
3.2. Dose Response of BCG Tokyo on Cytokine Production and Cell Proliferation
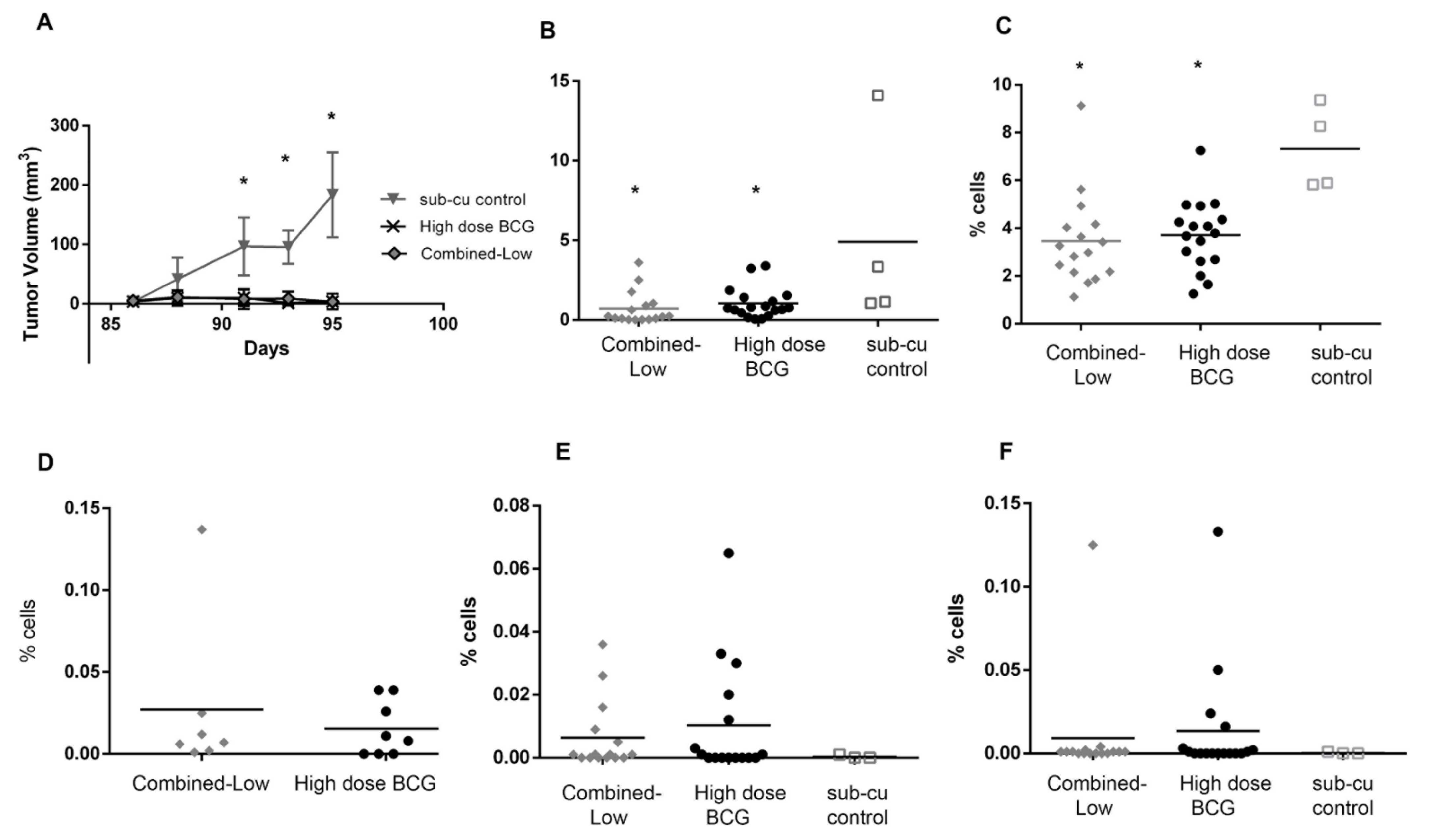
3.3. Impact of High and Low Dose BCG Tokyo Alone or Combined with Cytokine Gene Therapy on Orthotopic Bladder Tumors in Mice
3.4. Gene Expression in the Local Environment in Response to Therapy
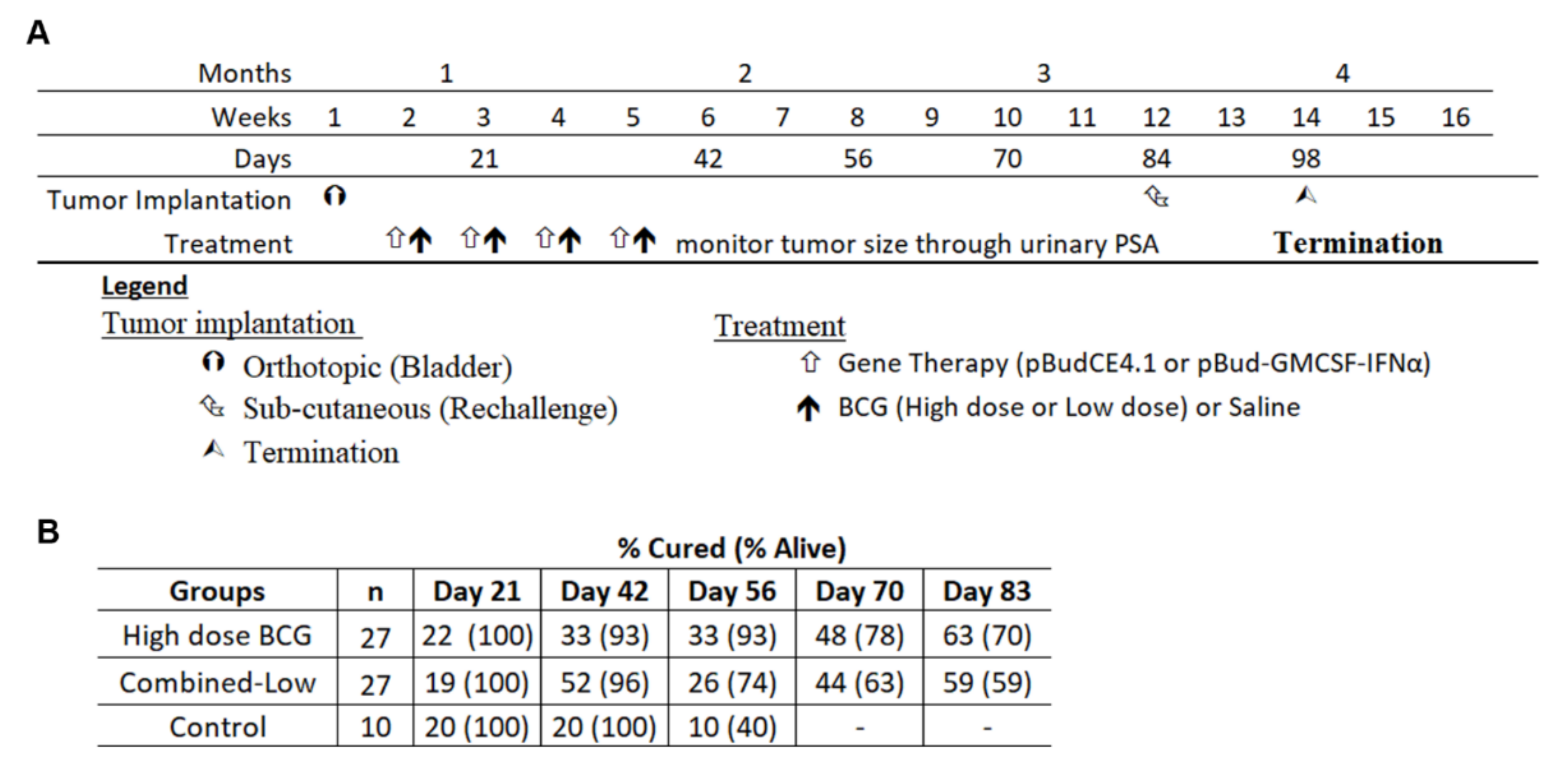
3.5. Re-Challenge of Cured Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Lamm, D.L. Efficacy and safety of bacille Calmette-Guerin immunotherapy in superficial bladder cancer. Clin. Infect. Dis. 2000, 31, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhang, Y.; Zheng, H.; Pan, Y.; Liu, H.; Du, P. Genome sequencing and analysis of BCG vaccine strains. PLoS ONE 2013, 8, e71243. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Gu, X.; Li, Y.; Wu, Q. Mechanisms of BCG in the treatment of bladder cancer-current understanding and the prospect. Biomed. Pharmacother. 2020, 129, 110393. [Google Scholar] [CrossRef] [PubMed]
- Secanella-Fandos, S.; Luquin, M.; Julian, E. Connaught and Russian strains showed the highest direct antitumor effects of different Bacillus Calmette-Guerin substrains. J. Urol. 2013, 189, 711–718. [Google Scholar] [CrossRef] [PubMed]
- Noon, A.P.; Kulkarni, G.S. All bacillus Calmette-Guerin (BCG) strains are equal, but some BCG strains are more equal than others. Eur. Urol. 2014, 66, 689–691. [Google Scholar] [CrossRef] [PubMed]
- Sengiku, A.; Ito, M.; Miyazaki, Y.; Sawazaki, H.; Takahashi, T.; Ogura, K. A prospective comparative study of intravesical bacillus Calmette-Guerin therapy with the Tokyo or Connaught strain for nonmuscle invasive bladder cancer. J. Urol. 2013, 190, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Taniguchi, K.; Koga, S.; Nishikido, M.; Yamashita, S.; Sakuragi, T.; Kanetake, H. Systemic immune response after intravesical instillation of bacille Calmette-Guerin (BCG) for superficial bladder cancer. Clin. Exp. Immunol. 1999, 115, 131–135. [Google Scholar] [CrossRef]
- Kamat, A.M.; Briggman, J.; Urbauer, D.L.; Svatek, R.; Nogueras Gonzalez, G.M.; Anderson, R. Cytokine Panel for Response to Intravesical Therapy (CyPRIT): Nomogram of Changes in Urinary Cytokine Levels Predicts Patient Response to Bacillus Calmette-Guerin. Eur. Urol. 2016, 69, 197–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redelman-Sidi, G.; Glickman, M.S.; Bochner, B.H. The mechanism of action of BCG therapy for bladder cancer—A current perspective. Nat. Rev. Urol. 2014, 11, 153–162. [Google Scholar] [CrossRef]
- Luo, Y.; Yamada, H.; Evanoff, D.P.; Chen, X. Role of Th1-stimulating cytokines in bacillus Calmette-Guerin (BCG)-induced macrophage cytotoxicity against mouse bladder cancer MBT-2 cells. Clin. Exp. Immunol. 2006, 146, 181–188. [Google Scholar] [CrossRef]
- Luo, Y.; Han, R.; Evanoff, D.P.; Chen, X. Interleukin-10 inhibits Mycobacterium bovis bacillus Calmette-Guerin (BCG)-induced macrophage cytotoxicity against bladder cancer cells. Clin. Exp. Immunol. 2010, 160, 359–368. [Google Scholar] [CrossRef]
- Joudi, F.N.; Smith, B.J.; O’Donnell, M.A. Final results from a national multicenter phase II trial of combination bacillus Calmette-Guerin plus interferon alpha-2B for reducing recurrence of superficial bladder cancer. Urol. Oncol. 2006, 24, 344–348. [Google Scholar] [CrossRef]
- Smith, S.G.; Zaharoff, D.A. Future directions in bladder cancer immunotherapy: Towards adaptive immunity. Immunotherapy 2016, 8, 351–365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, C.J.; Nguyen, P.H.D.; Wasser, M.; Kumar, P.; Lee, Y.H.; Nasir, N.J.M. Immunological Hallmarks for Clinical Response to BCG in Bladder Cancer. Front. Immunol. 2020, 11, 615091. [Google Scholar] [CrossRef] [PubMed]
- Kates, M.; Matoso, A.; Choi, W.; Baras, A.S.; Daniels, M.J.; Lombardo, K. Adaptive Immune Resistance to Intravesical BCG in Non-Muscle Invasive Bladder Cancer: Implications for Prospective BCG-Unresponsive Trials. Clin. Cancer Res. 2020, 26, 882–891. [Google Scholar] [CrossRef] [PubMed]
- Elsäßer, J.; Janssen, M.W.; Becker, F.; Suttmann, H.; Schmitt, K.; Sester, U. Antigen-specific CD4 T cells are induced after intravesical BCG-instillation therapy in patients with bladder cancer and show similar cytokine profiles as in active tuberculosis. PLoS ONE 2013, 8, e69892. [Google Scholar] [CrossRef] [PubMed]
- Kates, M.; Nirschl, T.; Sopko, N.A.; Matsui, H.; Kochel, C.M.; Reis, L.O. Intravesical BCG Induces CD4(+) T-Cell Expansion in an Immune Competent Model of Bladder Cancer. Cancer Immunol. Res. 2017, 5, 594–603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antonelli, A.C.; Binyamin, A.; Hohl, T.M.; Glickman, M.S.; Redelman-Sidi, G. Bacterial immunotherapy for cancer induces CD4-dependent tumor-specific immunity through tumor-intrinsic interferon-γ signaling. Proc. Natl. Acad. Sci. USA 2020, 117, 18627–18637. [Google Scholar] [CrossRef]
- Rentsch, C.A.; Birkhauser, F.D.; Biot, C.; Gsponer, J.R.; Bisiaux, A.; Wetterauer, C. Bacillus Calmette-Guerin Strain Differences Have an Impact on Clinical Outcome in Bladder Cancer Immunotherapy. Eur. Urol. 2014, 66, 677–688. [Google Scholar] [CrossRef] [PubMed]
- Biot, C.; Rentsch, C.A.; Gsponer, J.R.; Birkhauser, F.D.; Jusforgues-Saklani, H.; Lemaitre, F. Preexisting BCG-specific T cells improve intravesical immunotherapy for bladder cancer. Sci. Transl. Med. 2012, 4, 137ra72. [Google Scholar] [CrossRef]
- Lamm, D.L.; Blumenstein, B.A.; Crissman, J.D.; Montie, J.E.; Gottesman, J.E.; Lowe, B.A. Maintenance bacillus Calmette-Guerin immunotherapy for recurrent TA, T1 and carcinoma in situ transitional cell carcinoma of the bladder: A randomized Southwest Oncology Group Study. J. Urol. 2000, 163, 1124–1129. [Google Scholar] [CrossRef]
- Martínez-Piñeiro, J.A.; Flores, N.; Isorna, S.; Solsona, E.; Sebastián, J.L.; Pertusa, C. Long-term follow-up of a randomized prospective trial comparing a standard 81 mg dose of intravesical bacille Calmette-Guérin with a reduced dose of 27 mg in superficial bladder cancer. BJU Int. 2002, 89, 671–680. [Google Scholar] [CrossRef] [PubMed]
- Oddens, J.; Brausi, M.; Sylvester, R.; Bono, A.; van de Beek, C.; van Andel, G. Final results of an EORTC-GU cancers group randomized study of maintenance bacillus Calmette-Guerin in intermediate- and high-risk Ta, T1 papillary carcinoma of the urinary bladder: One-third dose versus full dose and 1 year versus 3 years of maintenance. Eur. Urol. 2013, 63, 462–472. [Google Scholar] [CrossRef]
- D’Andrea, D.; Gontero, P.; Shariat, S.F.; Soria, F. Intravesical bacillus Calmette-Guérin for bladder cancer: Are all the strains equal? Transl. Androl. Urol. 2019, 8, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Liu, X.; Niu, H.; Lv, W.; Han, X.; Zhang, Y.; Zhu, B. Persistent stimulation with Mycobacterium tuberculosis antigen impairs the proliferation and transcriptional program of hematopoietic cells in bone marrow. Mol. Immunol. 2019, 112, 115–122. [Google Scholar] [CrossRef]
- Ray, D.; Yung, R. Immune senescence, epigenetics and autoimmunity. Clin. Immunol. 2018, 196, 59–63. [Google Scholar] [CrossRef] [PubMed]
- Zlotta, A.R.; van Vooren, J.P.; Huygen, K.; Drowart, A.; Decock, M.; Pirson, M.; Schulman, C.C. What is the optimal regimen for BCG intravesical therapy? Are six weekly instillations necessary? Eur. Urol. 2000, 37, 470–477. [Google Scholar] [CrossRef]
- De Boer, E.C.; Rooyakkers, S.J.; Schamhart, D.H.; de Reijke, T.M.; Kurth, K.H. BCG dose reduction by decreasing the instillation frequency: Effects on local Th1/Th2 cytokine responses in a mouse model. Eur. Urol. 2005, 48, 333–338. [Google Scholar] [CrossRef]
- Rentsch, C.A.; Biot, C.; Gsponer, J.R.; Bachmann, A.; Albert, M.L.; Breban, R. BCG-mediated bladder cancer immunotherapy: Identifying determinants of treatment response using a calibrated mathematical model. PLoS ONE 2013, 8, e56327. [Google Scholar]
- Shah, G.; Zhang, G.; Chen, F.; Cao, Y.; Kalyanaraman, B.; See, W.A. The Dose-Response Relationship of bacillus Calmette-Guerin and Urothelial Carcinoma Cell Biology. J. Urol. 2016, 195, 1903–1910. [Google Scholar] [CrossRef] [PubMed]
- Tham, S.M.; Mahendran, R.; Chiong, E.; Wu, Q.H.; Esuvaranathan, K. Gmcsf and Ifnalpha gene therapy improves the response to BCG immunotherapy in a murine model of bladder cancer. Future Oncol. 2020, 16, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, M.; Bay, B.H.; Lee, Y.K.; Mahendran, R. Lactobacilli secreting a tumor antigen and IL15 activates neutrophils and dendritic cells and generates cytotoxic T lymphocytes against cancer cells. Cell Immunol. 2011, 271, 89–96. [Google Scholar] [CrossRef]
- Wu, Q.; Esuvaranathan, K.; Mahendran, R. Monitoring the response of orthotopic bladder tumors to granulocyte macrophage colony-stimulating factor therapy using the prostate-specific antigen gene as a reporter. Clin. Cancer Res. 2004, 10, 6977–6984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tham, S.M.; Esuvaranathan, K.; Mahendran, R. A Murine Orthotopic Bladder Tumor Model and Tumor Detection System. JoVE 2017, 119, e55078. [Google Scholar]
- Wu, Q.; Mahendran, R.; Esuvaranathan, K. Nonviral cytokine gene therapy on an orthotopic bladder cancer model. Clin. Cancer Res. 2003, 9, 4522–4528. [Google Scholar] [PubMed]
- Morel, C.; Badell, E.; Abadie, V.; Robledo, M.; Setterblad, N.; Gluckman, J.C. Mycobacterium bovis BCG-infected neutrophils and dendritic cells cooperate to induce specific T cell responses in humans and mice. Eur. J. Immunol. 2008, 38, 437–447. [Google Scholar] [CrossRef]
- Esuvaranathan, K.; Ravuru, M.; Kamaraj, R.; Ng, T.P.; Chan, Y.H.; Cheng, C.W.S.; Chia, S.J.; Ng, F.C.; Feng, L.; Mahendran, R. Long term results of double blind randomised controlled trial of interferon alpha 2b and low dose BCG in patients with high risk non-muscle invasive bladder cancer. BJU Int. 2014, 3, 1–37. [Google Scholar]
- Summerhayes, I.C.; Franks, L.M. Effects of donor age on neoplastic transformation of adult mouse bladder epithelium in vitro. J. Natl. Cancer Inst. 1979, 62, 1017–1023. [Google Scholar]
- Seow, S.W.; Cai, S.; Rahmat, J.N.; Bay, B.H.; Lee, Y.K.; Chan, Y.H. Lactobacillus rhamnosus GG induces tumor regression in mice bearing orthotopic bladder tumors. Cancer Sci. 2010, 101, 751–758. [Google Scholar] [CrossRef]
- Witjes, J.A.; Dalbagni, G.; Karnes, R.J.; Shariat, S.; Joniau, S.; Palou, J. The efficacy of BCG TICE and BCG Connaught in a cohort of 2099 patients with T1G3 non-muscle-invasive bladder cancer. Urol. Oncol. 2016, 34, 484.e19–484.e25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pook, S.H.; Rahmat, J.N.; Esuvaranathan, K.; Mahendran, R. Internalization of Mycobacterium bovis, Bacillus Calmette Guerin, by bladder cancer cells is cytotoxic. Oncol. Rep. 2007, 18, 1315–1320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.Y.; Kim, S.J.; Chi, B.H.; Kwon, J.K.; Chang, I.H. Modulating the internalization of bacille Calmette-Guerin by cathelicidin in bladder cancer cells. Urology 2015, 85, 964.e7–964.e12. [Google Scholar] [CrossRef]
- Sakuishi, K.; Apetoh, L.; Sullivan, J.M.; Blazar, B.R.; Kuchroo, V.K.; Anderson, A.C. Targeting Tim-3 and PD-1 pathways to reverse T cell exhaustion and restore anti-tumor immunity. J. Exp. Med. 2010, 207, 2187–2194. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.; Tatsumi, Y.; Gotoh, D.; Ohnishi, S.; Owari, T.; Iida, K. Regulatory T Cells and Tumor-Associated Macrophages in the Tumor Microenvironment in Non-Muscle Invasive Bladder Cancer Treated with Intravesical Bacille Calmette-Guérin: A Long-Term Follow-Up Study of a Japanese Cohort. Int. J. Mol. Sci. 2017, 18, 2186. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, R.; Tan, Z.; Liang, P. Identification of a novel ligand-receptor pair constitutively activated by ras oncogenes. J. Biol. Chem. 2000, 275, 24436–24443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Chen, X.; Han, R.; Chorev, M.; Dewolf, W.C.; O’Donnell, M.A. Mutated ras p21 as a target for cancer therapy in mouse transitional cell carcinoma. J. Urol. 1999, 162, 1519–1526. [Google Scholar] [CrossRef]
- Colopy, S.A.; Bjorling, D.E.; Mulligan, W.A.; Bushman, W. A population of progenitor cells in the basal and intermediate layers of the murine bladder urothelium contributes to urothelial development and regeneration. Dev. Dyn. 2014, 243, 988–998. [Google Scholar] [CrossRef] [Green Version]
- Acharya, P.; Beckel, J.; Ruiz, W.G.; Wang, E.; Rojas, R.; Birder, L. Distribution of the tight junction proteins ZO-1, occludin, and claudin-4, -8, and -12 in bladder epithelium. Am. J. Physiol. Renal Physiol. 2004, 287, F305–F318. [Google Scholar] [CrossRef]
- Korrer, M.J.; Zhang, Y.; Routes, J.M. Possible role of arginase-1 in concomitant tumor immunity. PLoS ONE 2014, 9, e91370. [Google Scholar]
- Liu, Q.; Tian, Y.; Zhao, X.; Jing, H.; Xie, Q.; Li, P. NMAAP1 Expressed in BCG-Activated Macrophage Promotes M1 Macrophage Polarization. Mol. Cells 2015, 38, 886–894. [Google Scholar]
- Ye, Z.; Zeng, Z.; Shen, Y.; Yang, Q.; Chen, D.; Chen, Z. ODC1 promotes proliferation and mobility via the AKT/GSK3β/β-catenin pathway and modulation of acidotic microenvironment in human hepatocellular carcinoma. OncoTargets Ther. 2019, 12, 4081–4092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolenz, C.; Knauf, D.; John, A.; Erben, P.; Steidler, A.; Schneider, S.W. Decreased Invasion of Urothelial Carcinoma of the Bladder by Inhibition of Matrix-Metalloproteinase 7. Bladder Cancer 2018, 4, 67–75. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Zhu, H.; Gao, Z.; Li, J.; Zhuang, J.; Dong, Y. Wnt7a activates canonical Wnt signaling, promotes bladder cancer cell invasion, and is suppressed by miR-370-3p. J. Biol. Chem. 2018, 293, 6693–6706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M.; Gong, H.; Wang, J.; Tao, L.; Xu, D.; Bao, E. SENP2 regulates MMP13 expression in a bladder cancer cell line through SUMOylation of TBL1/TBLR1. Sci. Rep. 2015, 5, 13996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, L.; Liu, W.R.; Tian, M.X.; Jiang, X.F.; Wang, H.; Zhou, P.Y. CCL24 contributes to HCC malignancy via RhoB- VEGFA-VEGFR2 angiogenesis pathway and indicates poor prognosis. Oncotarget 2017, 8, 5135–5148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gan, Y.H.; Zhang, Y.; Khoo, H.E.; Esuvaranathan, K. Antitumour immunity of Bacillus Calmette-Guerin and interferon alpha in murine bladder cancer. Eur. J. Cancer 1999, 35, 1123–1129. [Google Scholar] [CrossRef]
- Joseph, M.; Enting, D. Immune Responses in Bladder Cancer-Role of Immune Cell Populations, Prognostic Factors and Therapeutic Implications. Front. Oncol. 2019, 9, 1270. [Google Scholar] [CrossRef] [Green Version]
- Pettenati, C.; Ingersoll, M.A. Mechanisms of BCG immunotherapy and its outlook for bladder cancer. Nat. Rev. Urol. 2018, 15, 615–625. [Google Scholar] [CrossRef]
- Kurashige, T.; Noguchi, Y.; Saika, T.; Ono, T.; Nagata, Y.; Jungbluth, A. Ny-ESO-1 expression and immunogenicity associated with transitional cell carcinoma: Correlation with tumor grade. Cancer Res. 2001, 61, 4671–4674. [Google Scholar]
- Sharma, P.; Gnjatic, S.; Jungbluth, A.A.; Williamson, B.; Herr, H.; Stockert, E. Frequency of NY-ESO-1 and LAGE-1 expression in bladder cancer and evidence of a new NY-ESO-1 T-cell epitope in a patient with bladder cancer. Cancer Immun. 2003, 3, 19. [Google Scholar] [CrossRef]
- Horn, T.; Grab, J.; Schusdziarra, J.; Schmid, S.; Maurer, T.; Nawroth, R. Antitumor T cell responses in bladder cancer are directed against a limited set of antigens and are modulated by regulatory T cells and routine treatment approaches. Int. J. Cancer 2013, 133, 2145–2156. [Google Scholar] [CrossRef] [PubMed]
- Botteman, M.F.; Pashos, C.L.; Redaelli, A.; Laskin, B.; Hauser, R. The health economics of bladder cancer: A comprehensive review of the published literature. Pharmacoeconomics 2003, 21, 1315–1330. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tham, S.M.; Rahmat, J.N.; Chiong, E.; Wu, Q.; Esuvaranathan, K.; Mahendran, R. Intravesical High Dose BCG Tokyo and Low Dose BCG Tokyo with GMCSF+IFN α Induce Systemic Immunity in a Murine Orthotopic Bladder Cancer Model. Biomedicines 2021, 9, 1766. https://doi.org/10.3390/biomedicines9121766
Tham SM, Rahmat JN, Chiong E, Wu Q, Esuvaranathan K, Mahendran R. Intravesical High Dose BCG Tokyo and Low Dose BCG Tokyo with GMCSF+IFN α Induce Systemic Immunity in a Murine Orthotopic Bladder Cancer Model. Biomedicines. 2021; 9(12):1766. https://doi.org/10.3390/biomedicines9121766
Chicago/Turabian StyleTham, Sin Mun, Juwita N. Rahmat, Edmund Chiong, Qinghui Wu, Kesavan Esuvaranathan, and Ratha Mahendran. 2021. "Intravesical High Dose BCG Tokyo and Low Dose BCG Tokyo with GMCSF+IFN α Induce Systemic Immunity in a Murine Orthotopic Bladder Cancer Model" Biomedicines 9, no. 12: 1766. https://doi.org/10.3390/biomedicines9121766
APA StyleTham, S. M., Rahmat, J. N., Chiong, E., Wu, Q., Esuvaranathan, K., & Mahendran, R. (2021). Intravesical High Dose BCG Tokyo and Low Dose BCG Tokyo with GMCSF+IFN α Induce Systemic Immunity in a Murine Orthotopic Bladder Cancer Model. Biomedicines, 9(12), 1766. https://doi.org/10.3390/biomedicines9121766