Proteomic Characterization of Two Extracellular Vesicle Subtypes Isolated from Human Glioblastoma Stem Cell Secretome by Sequential Centrifugal Ultrafiltration


 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Cell Cultures
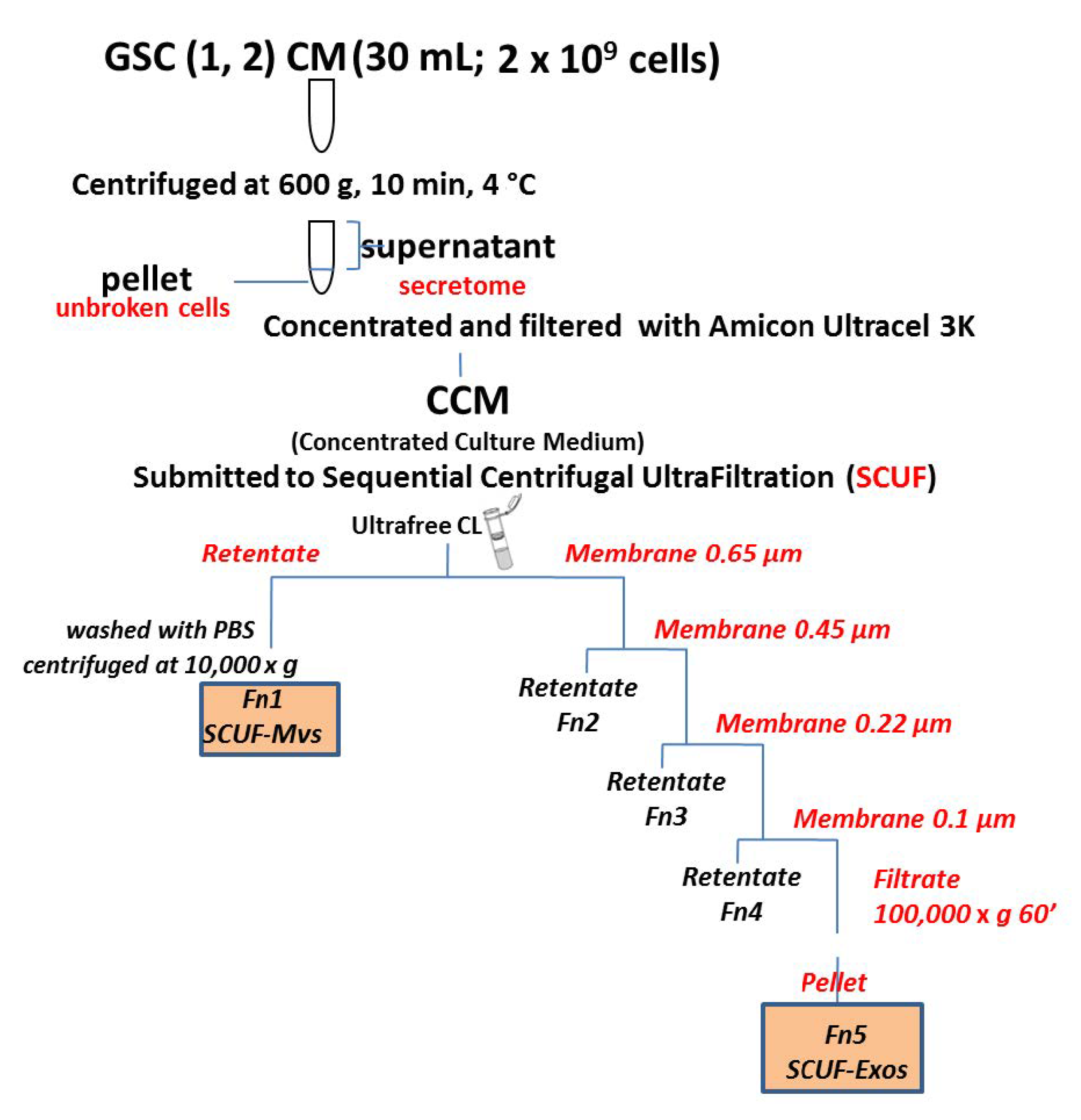
2.3. EV Isolation by Sequential Centrifugal Ultra-Filtration
- centrifugation of 2 mL of CCM at 3000× g combined with the use of 0.65 μm ultrafilters;
- transfer of the non-filtered fraction into a microtube with 0.5 mL of PBS and centrifugation at 10,000× g for 30 min. The particles isolated from this step were then characterized as MVs or Fn1 fraction;
- sequential filtration of the fraction obtained from the passage 1 through 0.45, 0.22, and 0.1 μm filters;
- ultrafiltration of the last filtered fraction at 100,000× g for 1 h. The particles isolated from this step were then characterized as Exos or Fn5 fraction;
- re-suspension of all preparations with 500 μL of phosphate buffered saline (PBS) and subsequent protein lysis of each fraction to perform two-dimensional electrophoretic analysis.
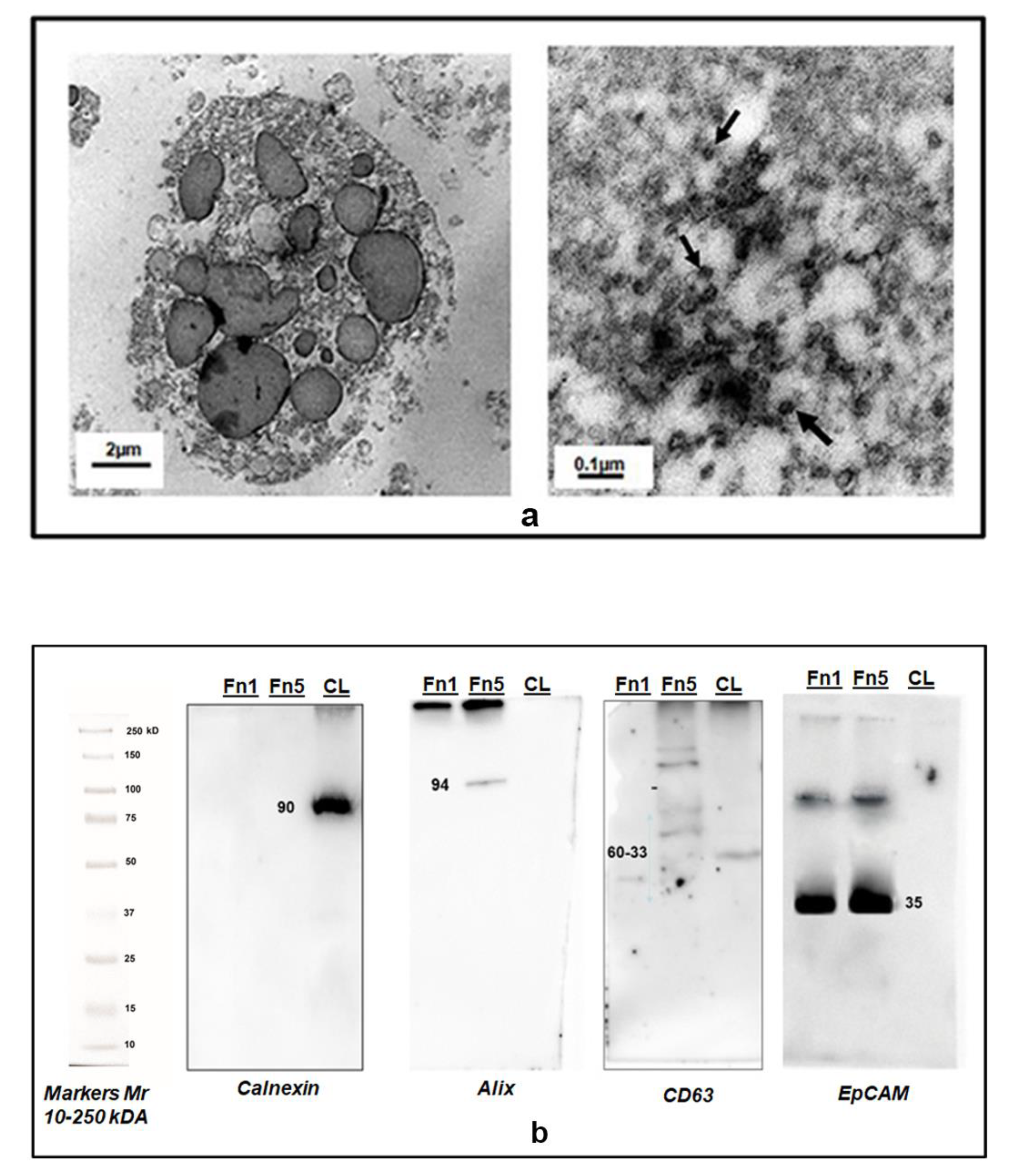
2.4. Electron Microscopy of Isolated EVs
2.5. Western Blot Analysis
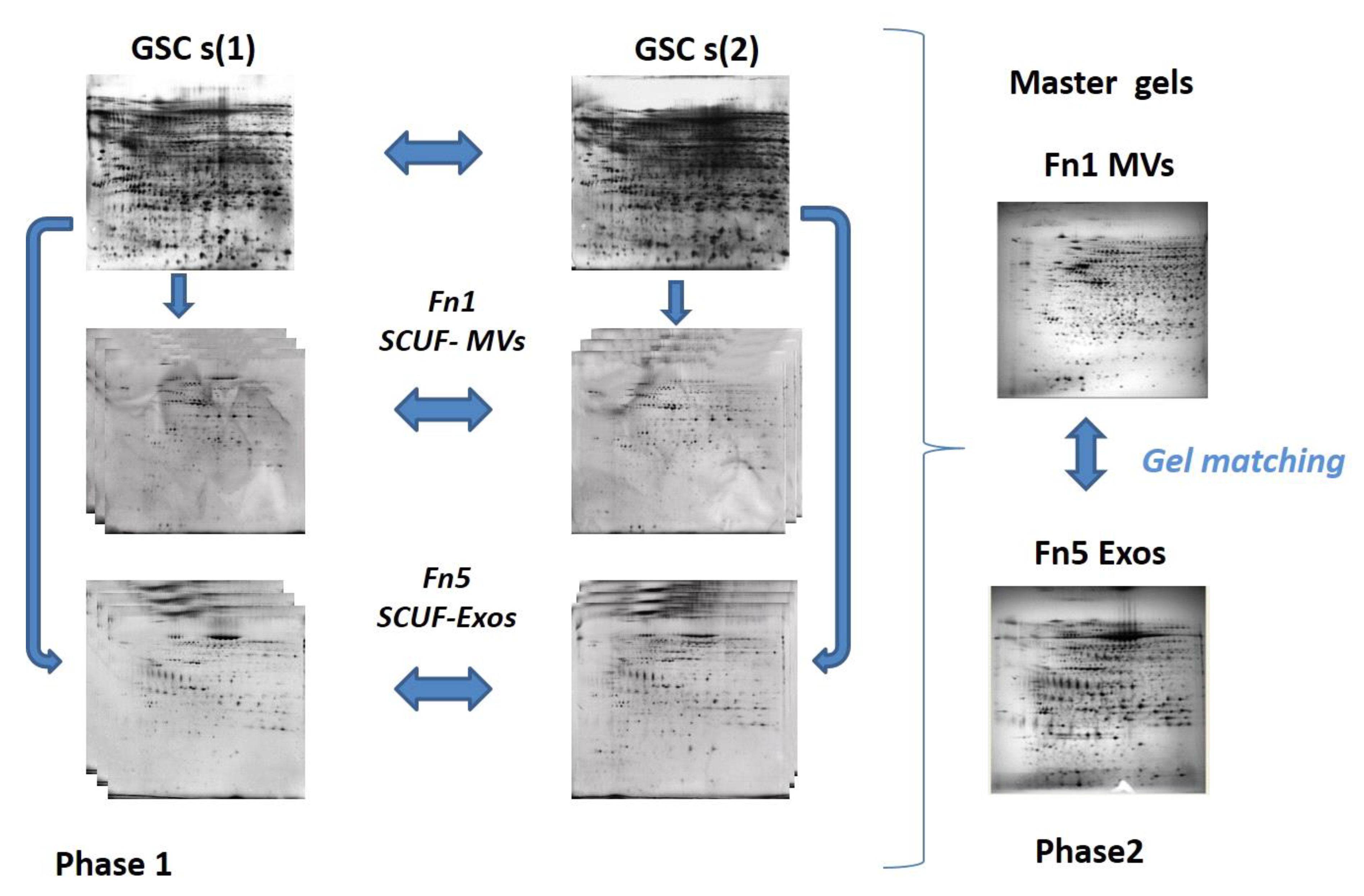
2.6. 2DE Analysis
2.7. Protein Digestion and MALDI TOF MS/MS Analysis
2.8. Bioinformatic Analysis
2.9. Data Analysis
3. Results
3.1. Isolation of Two EV Subtypes by Sequential Centrifugal Ultrafiltration (SCUF) Technique and Their Characterization by Transmission Electron Microscopy (TEM) and Western Blot Analysis
3.2. Proteomic Analysis of Exo and MV Content
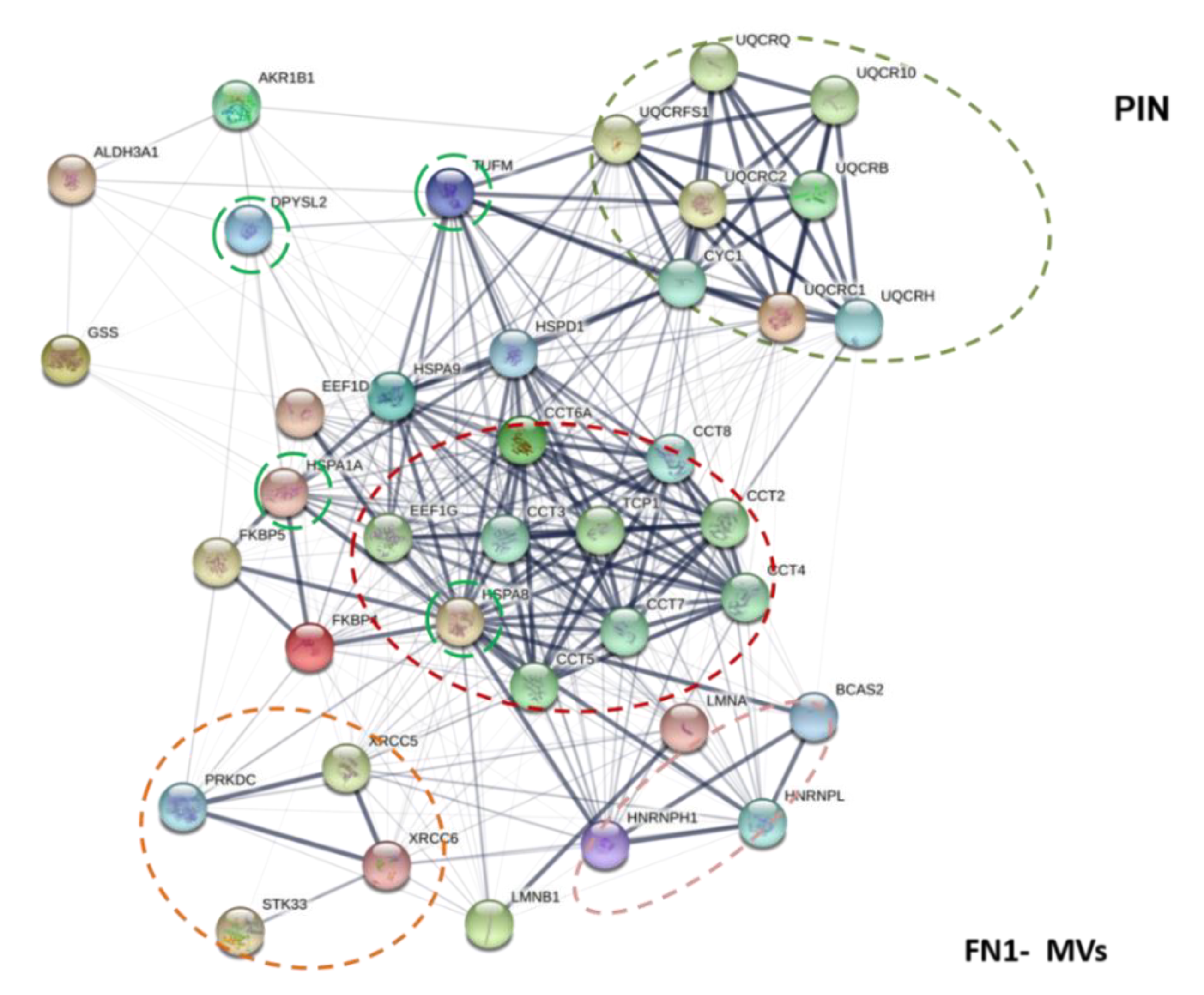
3.3. Identification of Some Proteins Exclusively Present in MV Fraction
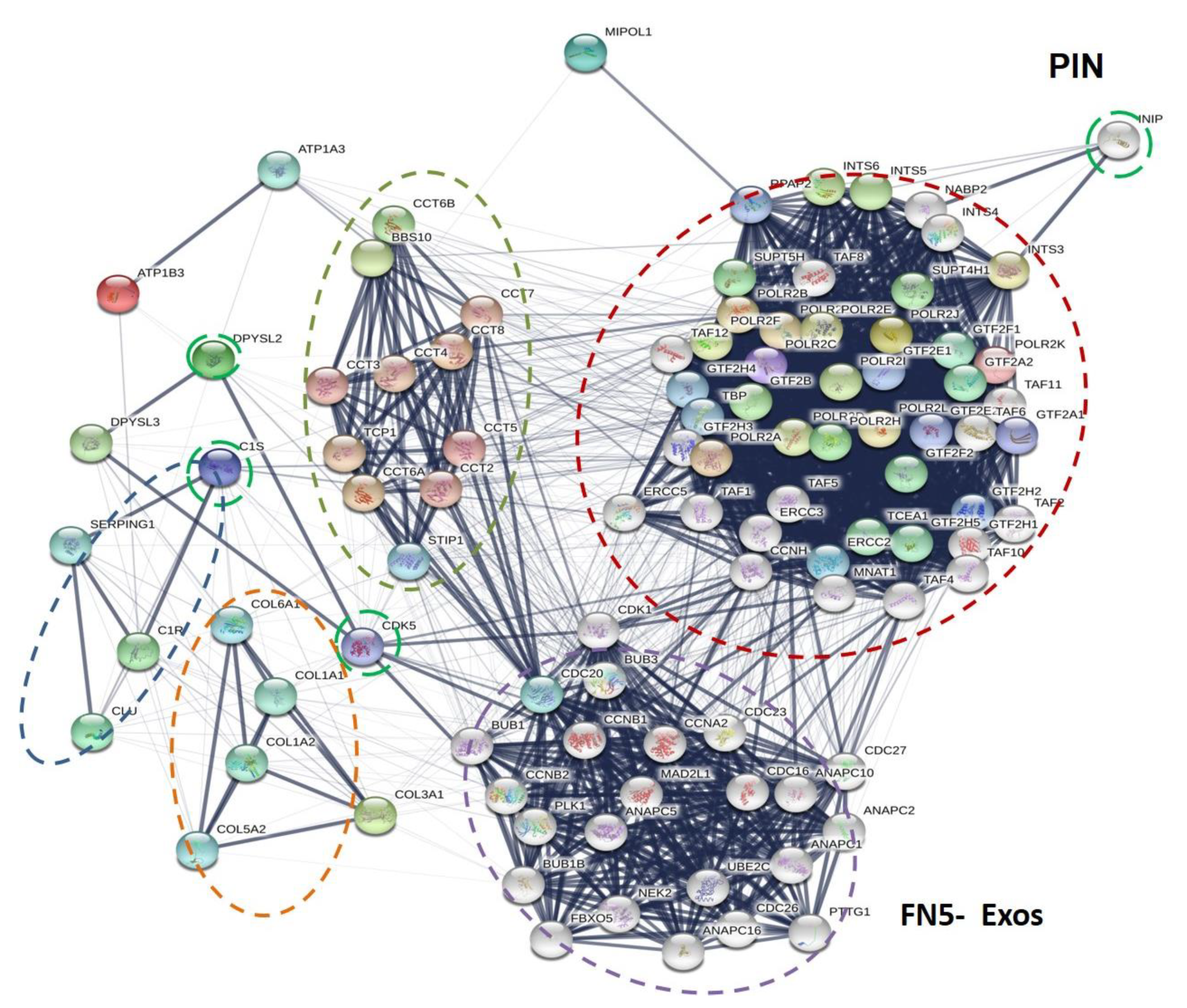
3.4. Identification of Some Proteins Exclusively Present in the Exo Fraction
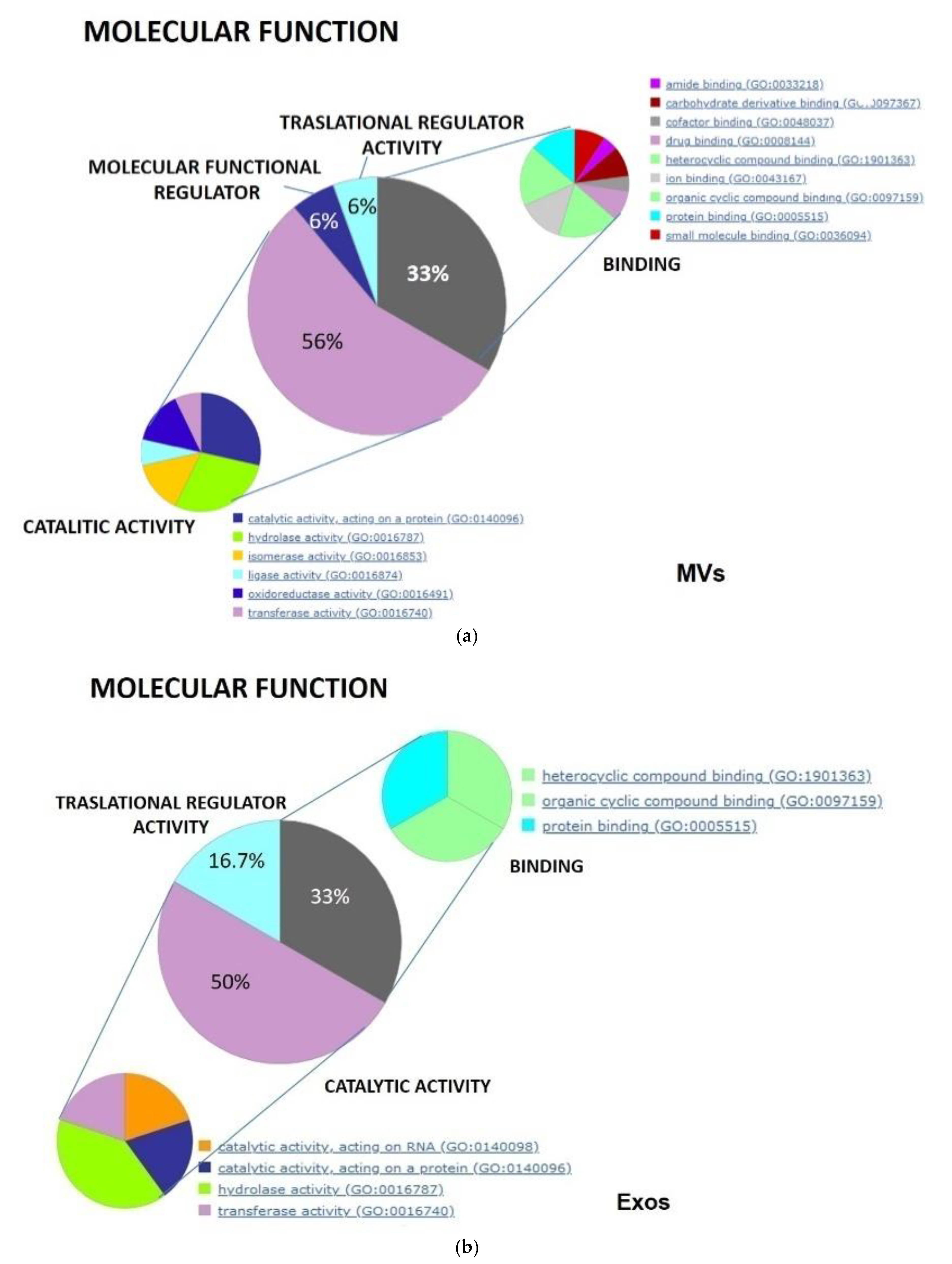
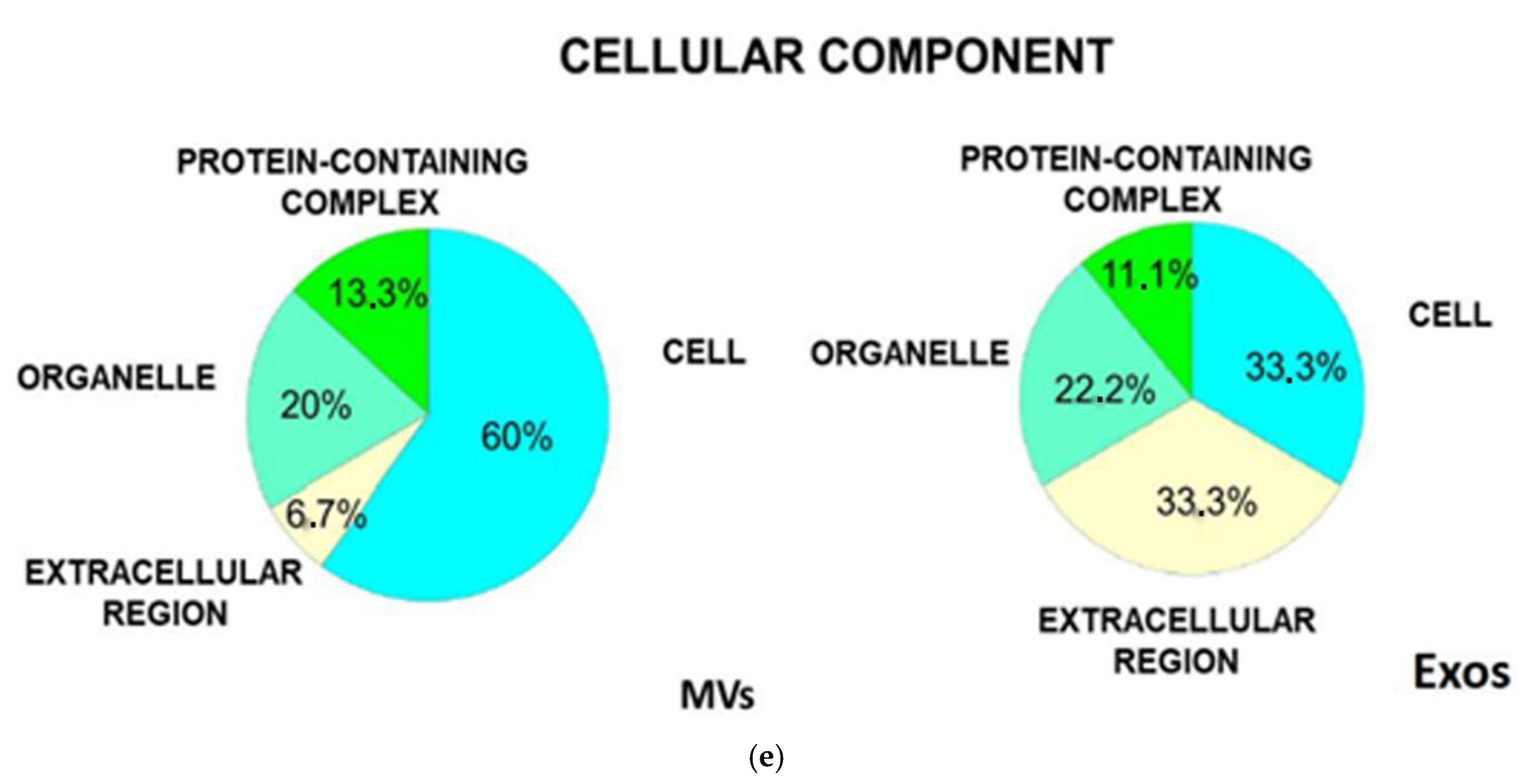
3.5. Functional and Biological Analysis of the Proteome of Isolated Exo and MV Fractions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yáñez-Mó, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borràs, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.; Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell. Vesicles 2015, 4, 27066. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, R.; Sedger, L.M. Intercellular Vesicular Transfer by Exosomes, Microparticles and Oncosomes—Implications for Cancer Biology and Treatments. Front. Oncol. 2019, 9, 125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akers, J.C.; Gonda, D.; Kim, R.; Carter, B.S.; Chen, C.C. Biogenesis of extracellular vesicles (EV): Exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J. Neurooncol. 2013, 113, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minciacchi, V.R.; Freeman, M.R.; Di Vizio, D. Extracellular vesicles in cancer: Exosomes, microvescicles and the emerging role of large oncosomes. Semin. Cell Dev. Biol. 2015, 40, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciardiello, C.; Migliorino, R.; Leone, A.; Budillon, A. Large extracellular vesicles: Size matters in tumor progression. Cytokine Growth Factor Rev. 2020, 51, 69–74. [Google Scholar] [CrossRef]
- Nakano, I.; Garnier, D.; Minata, M.; Rak, J. Extracellular vesicles in the biology of brain tumour stem cells-Implications for inter-cellular communication, therapy and biomarker development. Semin. Cell Dev. Biol. 2015, 40, 17–26. [Google Scholar] [CrossRef]
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996. [Google Scholar] [CrossRef]
- Urbańska, K.; Sokołowska, J.; Szmidt, M.; Sysa, P. Glioblastoma multiforme, an overview. Contemp. Oncol. 2014, 18, 307–312. [Google Scholar] [CrossRef]
- Yi, Y.; Hsieh, I.Y.; Huang, X.; Li, J.; Zhao, W. Glioblastoma Stem-Like Cells: Characteristics, Microenvironment, and Therapy. Front. Pharm. 2016, 7, 477. [Google Scholar] [CrossRef] [Green Version]
- Guerra-Rebollo, M.; Garrido, C.; Sánchez-Cid, L.; Soler-Botija, C.; Meca-Cortés, O.; Rubio, N.; Blanco, J. Targeting of replicating CD133 and OCT4/SOX2 expressing glioma stem cells selects a cell population that reinitiates tumors upon release of therapeutic pressure. Sci. Rep. 2019, 9, 9549. [Google Scholar] [CrossRef]
- Li, A.; Walling, J.; Kotliarov, Y.; Center, A.; Steed, M.E.; Ahn, S.J.; Rosenblum, M.; Mikkelsen, T.; Zenklusen, J.C.; Fine, H.A. Genomic changes and gene expression profiles reveal that established glioma cell lines are poorly representative of primary human gliomas. Mol. Cancer Res. 2008, 6, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Kotliarova, S.; Kotliarov, Y.; Li, A.; Su, Q.; Donin, N.M.; Pastorino, S.; Purow, B.W.; Christopher, N.; Zhang, W.; et al. Tumor stem cells derived from glioblastomas cultured in bFGF and EGF more closely mirror the phenotype and genotype of primary tumors than do serum-cultured cell lines. Cancer Cell 2006, 9, 391–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wee, B.; Charles, N.; Holland, E.C. Animal models to study cancer-initiating cells from glioblastoma. Front. Biosci. 2011, 16, 2243–2258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redzic, J.S.; Ung, T.H.; Graner, M.W. Glioblastoma extracellular vesicles: Reservoirs of potential biomarkers. Pharmgenom. Pers. Med. 2014, 7, 65–77. [Google Scholar] [CrossRef] [Green Version]
- Mallawaaratchy, D.M.; Hallal, S.; Russell, B.; Ly, L.; Ebrahimkhani, S.; Wei, H.; Christopherson, R.I.; Buckland, M.E.; Kaufman, K.L. Comprehensive proteome profiling of glioblastoma-derived extracellular vesicles identifies markers for more aggressive disease. J. Neurooncol. 2017, 131, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Indira Chandran, V.; Welinder, C.; Gonçalves de Oliveira, K.; Cerezo-Magaña, M.; Månsson, A.S.; Johansson, M.C.; Marko-Varga, G.; Belting, M. Global extracellular vesicle proteomic signature defines U87-MG glioma cell hypoxic status with potential implications for non-invasive diagnostics. J. Neurooncol. 2019, 144, 477–488. [Google Scholar] [CrossRef] [Green Version]
- García-Romero, N.; Carrión-Navarro, J.; Esteban-Rubio, S.; Lázaro-Ibáñez, E.; Peris-Celda, M.; Alonso, M.M.; Guzmán-De-Villoria, J.; Fernández-Carballal, C.; de Mendivil, A.O.; García-Duque, S.; et al. DNA sequences within glioma-derived extracellular vesicles can cross the intact blood-brain barrier and be detected in peripheral blood of patients. Oncotarget 2017, 8, 1416–1428. [Google Scholar] [CrossRef] [Green Version]
- Godlewski, J.; Ferrer-Luna, R.; Rooj, A.K.; Mineo, M.; Ricklefs, F.; Takeda, Y.S.; Nowicki, M.O.; Salińska, E.; Nakano, I.; Lee, H.; et al. MicroRNA Signatures and Molecular Subtypes of Glioblastoma: The Role of Extracellular Transfer. Stem. Cell Rep. 2017, 8, 1497–1505. [Google Scholar] [CrossRef] [Green Version]
- Spinelli, C.; Montermini, L.; Meehan, B.; Brisson, A.R.; Tan, S.; Choi, D.; Nakano, I.; Rak, J. Molecular subtypes and differentiation programmes of glioma stem cells as determinants of extracellular vesicle profiles and endothelial cell-stimulating activities. J. Extracell. Vesicles 2018, 7, 1490144. [Google Scholar] [CrossRef]
- Cumba Garcia, L.M.; Peterson, T.E.; Cepeda, M.A.; Johnson, A.J.; Parney, I.F. Isolation and Analysis of Plasma-Derived Exosomes in Patients with Glioma. Front. Oncol. 2019, 9, 651. [Google Scholar] [CrossRef] [Green Version]
- D’Alimonte, I.; Nargi, E.; Zuccarini, M.; Lanuti, P.; Di Iorio, P.; Giuliani, P.; Ricci-Vitiani, L.; Pallini, R.; Caciagli, F.; Ciccarelli, R. Potentiation of temozolomide antitumor effect by purine receptor ligands able to restrain the in vitro growth of human glioblastoma stemcells. Purinergic Signal. 2015, 11, 331–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ziberi, S.; Zuccarini, M.; Carluccio, M.; Giuliani, P.; Ricci-Vitiani, L.; Pallini, R.; Caciagli, F.; Di Iorio, P.; Ciccarelli, R. Upregulation of Epithelial-to-Mesenchymal Transition markers and P2X7 receptors is associated to increased invasiveness caused by P2X7 receptor stimulation in human glioblastoma stem cells. Cells 2019, 9, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Alessandris, Q.G.; Biffoni, M.; Martini, M.; Runci, D.; Buccarelli, M.; Cenci, T.; Signore, M.; Stancato, L.; Olivi, A.; De Maria, R.; et al. The clinicalvalue of patient-derived glioblastoma tumorspheres in predicting treatment response. Neuro-Oncology 2017, 19, 1097–1108. [Google Scholar] [CrossRef] [Green Version]
- Marziali, G.; Signore, M.; Buccarelli, M.; Grande, S.; Palma, A.; Biffoni, M.; Rosi, A.; D’Alessandris, Q.G.; Martini, M.; Larocca, L.M.; et al. Metabolic/Proteomic Signature Defines Two Glioblastoma Subtypes with Different Clinical Outcome. Sci. Rep. 2016, 6, 21557. [Google Scholar] [CrossRef]
- Ricci-Vitiani, L.; Pallini, R.; Larocca, L.M.; Lombardi, D.G.; Signore, M.; Pierconti, F.; Petrucci, G.; Montano, N.; Maira, G.; De Maria, R. Mesenchymal differentiation of glioblastoma stem cells. Cell Death Differ. 2008, 15, 1491–1498. [Google Scholar] [CrossRef] [Green Version]
- Pallini, R.; Ricci-Vitiani, L.; Banna, G.L.; Signore, M.; Lombardi, D.; Todaro, M.; Stassi, G.; Martini, M.; Maira, G.; Larocca, L.M.; et al. Cancer stem cell analysis and clinical outcome in patients with glioblastoma multiforme. Clin. Cancer Res. 2008, 14, 8205–8212. [Google Scholar] [CrossRef] [Green Version]
- Eramo, A.; Ricci-Vitiani, L.; Zeuner, A.; Pallini, R.; Lotti, F.; Sette, G.; Pilozzi, E.; Larocca, L.M.; Peschle, C.; De Maria, R. Chemotherapy resistance of glioblastoma stem cells. Cell Death Differ. 2006, 13, 1238–1241. [Google Scholar] [CrossRef] [Green Version]
- Xu, R.; Greening, D.W.; Rai, A.; Ji, H.; Simpson, R.J. Highly-purified exosomes and shed microvesicles isolated from the human colon cancer cell line LIM1863 by sequential centrifugal ultrafiltration are biochemically and functionally distinct. Methods 2015, 87, 11–25. [Google Scholar] [CrossRef]
- Brown, R.E.; Jarvis, K.L.; Hyland, K.J. Protein measurement using bicinchoninic acid: Elimination of interfering substances. Anal. Biochem. 1989, 180, 136–139. [Google Scholar] [CrossRef]
- Giampietro, F.; Sancilio, S.; Tiboni, G.M.; Rana, R.A.; Di Pietro, R. Levels of apoptosis in human granulosa cells seem to be comparable after therapy with a gonadotropin-releasing hormone agonist or antagonist. Fertil. Steril. 2006, 85, 412–419. [Google Scholar] [CrossRef]
- Mun, Y.S.; Kim, D.; Kim, I.T. Electrochemical Performance of FeSb₂-P@C Composites as Anode Materials for Lithium-Ion Storage. J. Nanosci. Nanotechnol. 2018, 18, 1343–1346. [Google Scholar] [CrossRef] [PubMed]
- Angelucci, S.; Marchisio, M.; Di Giuseppe, F.; Pierdomenico, L.; Sulpizio, M.; Eleuterio, E.; Lanuti, P.; Sabatino, G.; Miscia, S.; Di Ilio, C. Proteome analysis of human Wharton’s jelly cells during in vitro expansion. Proteome Sci. 2010, 8, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minciacchi, V.R.; You, S.; Spinelli, C.; Morley, S.; Zandian, M.; Aspuria, P.J.; Cavallini, L.; Ciardiello, C.; ReisSobreiro, M.; Morello, M.; et al. Large oncosomes contain distinct protein cargo and represent functional class of tumor-derived extracellular vesicles. Oncotarget 2015, 6, 11327–11341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dittmer, T.A.; Misteli, T. The lamin protein family. Genome Biol. 2011, 12, 222. [Google Scholar] [CrossRef] [Green Version]
- Geuens, T.; Bouhy, D.; Timmerman, V. The hnRNP family: Insights into their role in health and disease. Hum. Genet. 2016, 135, 851–867. [Google Scholar] [CrossRef] [Green Version]
- Sasikumar, A.N.; Perez, W.B.; Kinzy, T.G. The many roles of the eukaryotic elongation factor 1 complex. Wiley Interdiscip. Rev. RNA 2012, 3, 543–555. [Google Scholar] [CrossRef] [Green Version]
- Parmeggiani, A.; Nissen, P. Elongation factor Tu-targeted antibiotics: Four different structures, two mechanisms of action. FEBS Lett. 2006, 580, 4576–4581. [Google Scholar] [CrossRef] [Green Version]
- Kaul, S.C.; Deocaris, C.C.; Wadhwa, R. Three faces of mortalin: A housekeeper, guardian and killer. Exp. Gerontol. 2007, 42, 263–274. [Google Scholar] [CrossRef]
- Haura, E.B.; Sacco, R.; Li, J.; Muller, A.C.; Grebien, F.; Superti-Furga, G.; Bennet, K.L. Optimization of downscaled tandem affinity purifications to identify core protein complexes. J. Integr. OMICS 2012, 2, 55–68. [Google Scholar] [CrossRef] [Green Version]
- Jang, J.; Kim, M.R.; Kim, T.K.; Lee, W.R.; Kim, J.H.; Heo, K.; Lee, S. CLEC14a-HSP70-1A interaction regulates HSP70-1A-induced angiogenesis. Sci. Rep. 2017, 7, 10666. [Google Scholar] [CrossRef] [Green Version]
- Haziza, S.; Magnani, R.; Lan, D.; Keinan, O.; Saada, A.; Hershkovitz, E.; Yanay, N.; Cohen, Y.; Nevo, Y.; Houtz, R.L.; et al. Calmodulin Methyltransferase Is Required for Growth, Muscle Strength, Somatosensory Development and Brain Function. PLoS Genet. 2015, 11, e1005388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, W.J.; Stagos, D.; Marchitti, S.A.; Nebert, D.W.; Tipton, K.F.; Bairoch, A.; Vasiliou, V. Human aldehyde dehydrogenase genes: Alternative spliced transcriptional variants and their suggested nomenclature. Pharm. Genom. 2009, 19, 893–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hyndman, D.; Bauman, D.R.; Heredia, V.V.; Penning, T.M. The aldo-keto reductase superfamily homepage. Chem. Biol. Interact. 2003, 143–144, 621–631. [Google Scholar] [CrossRef]
- Han, N.; Li, W.; Zhang, M. The function of the RNA-binding protein hnRNP in cancer metastasis. J. Cancer Res. 2013, 9, S129–S134. [Google Scholar] [CrossRef]
- Cheng, D.-d.; Li, S.; Zhu, B.; Zhou, S.-m.; Yang, Q.-C. EEF1D overexpression promotes osteosarcoma cell proliferation by facilitating Akt-mTOR and Akt-bad signaling. J. Exp. Clin. Cancer Res. 2018, 35, 50. [Google Scholar] [CrossRef]
- Li, W.; Wubulikasimu, G.; Zhao, X.; Wang, C.; Liu, R.; Wang, L.; Zhu, X.; Chen, Z. UQCRC1 downregulation is correlated with lymph node metastasis and poor prognosis in CRC. Eur. J. Surg. Oncol. 2019, 45, 1005–1110. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, Y.; Cui, M.; Wang, X.; Lin, Z. Mortalin contributes to colorectal cancer by promoting proliferation and epithelial-mesenchymal transition. IUBMB Life 2020, 72, 771–781. [Google Scholar] [CrossRef]
- Clucas, J.; Valderrama, F. ERM proteins in cancer progression. J. Cell Sci. 2014, 127, 267–275. [Google Scholar] [CrossRef] [Green Version]
- Satelli, A.; Li, S. Vimentin in cancer and its potential as a molecular target for cancer therapy. Cell. Mol. Life Sci. 2011, 68, 3033–3046. [Google Scholar] [CrossRef] [Green Version]
- Izdebska, M.; Gagat, M.; Grzanka, A. Overexpression of lamin B1 induces mitotic catastrophe in colon cancer LoVo cells and is associated with worse clinical outcomes. Int. J. Oncol. 2018, 52, 89–102. [Google Scholar] [CrossRef]
- Huo, Y.; Zheng, Z.; Chen, Y.; Wang, Q.; Zhang, Z.; Deng, H. Downregulation of vimentin expression increased drug resistance in ovarian cancer cells. Oncotarget 2016, 7, 45876–45888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaffer, J.R.; Orlova, E.; Lee, M.K.; Leslie, E.J.; Raffensperger, Z.D.; Heike, C.L.; Cunningham, M.L.; Hecht, J.T.; Kau, C.H.; Nidey, N.L.; et al. Genome-Wide Association Study Reveals Multiple Loci Influencing Normal Human Facial Morphology. PLoS Genet. 2016, 12, e1006149. [Google Scholar] [CrossRef] [PubMed]
- Futai, M.; Omote, H.; Sambongi, Y.; Wada, Y. Synthase (H+ ATPase): Coupling between catalysis, mechanical work, and proton translocation. Biochim. Biophys. Acta 2000, 1458, s0005–s2728. [Google Scholar] [CrossRef] [Green Version]
- Normanno, N.; De Luca, A.; Bianco, C.; Strizzi, L.; Mancino, M.; Maiello, M.R.; Carotenuto, A.; De Feo, G.; Caponigro, F.; Salomon, D.S. Epidermal growth factor receptor (EGFR) signaling in cancer. Gene 2006, 366, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Yue, F.; Wang, L.S.; Xia, L.; Wang, X.L.; Feng, B.; Lu, A.G.; Chen, G.Q.; Zheng, M.H. Modulated T-complex protein 1 ζ and peptidyl-prolyl cis-trans isomerase B are two novel indicators for evaluating lymph node metastasis in colorectal cancer: Evidence from proteomics and bioinformatics. Proteom. Clin. Appl. 2009, 3, 1225–1235. [Google Scholar] [CrossRef]
- Ma, I.; Allan, A.L. The role of human aldehyde dehydrogenase in normal and cancer stem cells. Stem. Cell Rev. 2011, 7, 292–306. [Google Scholar] [CrossRef]
- Khayami, R.; Hashemi, S.R.; Kerachian, M.A. Role of aldo-keto reductase family 1 member B1 (AKR1B1) in the cancer process and its therapeutic potential. J. Cell. Mol. Med. 2020, 1–13. [Google Scholar] [CrossRef]
- Garcia-Aranda, M.; Téllez, T.; Munoz, M.; Redondo, M. Clusterin inhibition mediates sensitivity to chemotherapy and radiotherapy in human cancer. Anticancer Drugs 2017, 28, 702–716. [Google Scholar] [CrossRef]
- Santala, M.; Simojoki, M.; Risteli, J.; Risteli, L.; Kauppila, A. Type I and III collagen metabolites as predictors of clinical outcome in epithelial ovarian cancer. Clin. Cancer Res. 1999, 5, 4091–4096. [Google Scholar]
- Su, B.; Zhao, W.; Shi, B.; Zhang, Z.; Yu, X.; Xie, F.; Guo, Z.; Zhang, X.; Liu, J.; Shen, Q.; et al. Let-7d suppresses growth, metastasis, and tumor macrophage infiltration in renal cell carcinoma by targeting COL3A1 and CCL7. Mol. Cancer 2014, 13, 206. [Google Scholar] [CrossRef] [Green Version]
- Chang, I.W.; Lin, V.C.; Wu, W.J.; Liang, P.I.; Li, W.M.; Yeh, B.W.; He, H.L.; Liao, A.C.; Chan, T.C.; Li, C.F. Complement Component 1, s Subcomponent Overexpression is an Independent Poor Prognostic Indicator in Patients with Urothelial Carcinomas of the Upper Urinary Tract and Urinary Bladder. J. Cancer 2016, 7, 1396–1405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leong, M.M.L.; Cheung, A.K.L.; Kwok, T.C.T.; Lung, M.L. Functional characterization of a candidate tumor suppressor gene, Mirror Image Polydactyly 1, in nasopharyngeal carcinoma. Int. J. Cancer 2020, 146, 2891–2900. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.J.; Song, Q.F.; Jiang, S.S.; Song, Q.; Wang, W.; Zhang, G.H.; Kan, B.; Chen, L.J.; Yang, J.L.; Luo, F.; et al. Identification of ATP synthase beta subunit (ATPB) on the cell surface as a non-small cell lung cancer (NSCLC) associated antigen. BMC Cancer 2009, 9, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Li, Y.; Yu, T.-S.; McKay, R.M.; Burns, D.K.; Kernie, S.G.; Parada, L.F. A restricted cell population propagates glioblastoma growth after chemotherapy. Nature 2012, 488, 522–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montano, N.; Cenci, T.; Martini, M.; D’Alessandris, Q.G.; Pelacchi, F.; Ricci-Vitiani, L.; Maira, G.; de Maria, R.; Larocca, L.M.; Pallini, R. Expression of EGFRvIII in glioblastoma: Prognostic significance revisited. Neoplasia 2011, 13, 1113–1121. [Google Scholar] [CrossRef] [PubMed]
- Hessvik, N.P.; Llorente, A. Current knowledge on exosome biogenesis and release. Cell. Mol. Life Sci. 2018, 75, 193–208. [Google Scholar] [CrossRef] [Green Version]
- Choi, D.; Montermini, L.; Kim, D.K.; Meehan, B.; Roth, F.P.; Rak, J. The Impact of Oncogenic EGFRvIII on the Proteome of Extracellular Vesicles Released from Glioblastoma Cells. Mol. Cell. Proteom. 2018, 17, 1948–1964. [Google Scholar] [CrossRef] [Green Version]
- Lane, R.; Simon, T.; Vintu, M.; Solkin, B.; Koch, B.; Stewart, N.; Benstead-Hume, G.; Pearl, F.M.G.; Critchley, G.; Stebbing, J.; et al. Cell-derived extracellular vesicles can be used as a biomarker reservoir for glioblastoma tumor subtyping. Commun. Biol. 2019, 2, 315. [Google Scholar] [CrossRef] [Green Version]
- Zottel, A.; Jovčevska, I.; Šamec, N.; Mlakar, J.; Šribar, J.; Križaj, I.; Skoblar Vidmar, M.; Komel, R. Anti-vimentin, anti-TUFM, anti-NAP1L1 and anti-DPYSL2 nanobodies display cytotoxic effect and reduce glioblastoma cell migration. Adv. Med. Oncol. 2020, 12, 1758835920915302. [Google Scholar] [CrossRef]
- Wang, Q.; Lu, X.; Zhao, S.; Pang, M.; Wu, X.; Wu, H.; Hoffman, R.M.; Yang, Z.; Zhang, Y. Moesin Expression Is Associated with Glioblastoma Cell Proliferation and Invasion. Anticancer Res. 2017, 37, 2211–2218. [Google Scholar] [CrossRef] [Green Version]
- Ott, M.; Litzenburger, U.M.; Rauschenbach, K.J.; Bunse, L.; Ochs, K.; Sahm, F.; Pusch, S.; Opitz, C.A.; Blaes, J.; von Deimling, A.; et al. Suppression of TDO-mediated tryptophan catabolism in glioblastoma cells by a steroid-responsive FKBP52-dependent pathway. Glia 2015, 63, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Graner, M.W.; Cumming, R.I.; Bigner, D.D. The heat shock response and chaperones/heat shock proteins in brain tumors: Surface expression, release, and possible immune consequences. J. Neurosci. 2007, 27, 11214–11227. [Google Scholar] [CrossRef] [PubMed]
- Hermisson, M.; Strik, H.; Rieger, J.; Dichgans, J.; Meyermann, R.; Weller, M. Expression and functional activity of heat shock proteins in human glioblastoma multiforme. Neurology 2000, 54, 1357–1365. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, S.G.; Ezrin, A.; Jackson, E.; Dewey, L.; Doucette, A.A. A robust strategy for proteomic identification of biomarkers of invasive phenotype complexed with extracellular heat shock proteins. Cell Stress Chaperones 2019, 24, 1197–1209. [Google Scholar] [CrossRef]
- González-Cruz, R.D.; Dahl, K.N.; Darling, E.M. The emerging role of lamin C as an important LMNA isoform in mechanophenotype. Front. Cell Dev. Biol. 2018, 6, 151. [Google Scholar] [CrossRef] [Green Version]
- Graner, M.W.; Alzate, O.; Dechkovskaia, A.M.; Keene, J.D.; Sampson, J.H.; Mitchell, D.A.; Bigner, D.D. Proteomic and immunologic analyses of brain tumor exosomes. FASEB J. 2009, 23, 1541–1557. [Google Scholar] [CrossRef] [Green Version]
- Rocha, C.R.; Kajitani, G.S.; Quinet, A.; Fortunato, R.S.; Menck, C.F. NRF2 and glutathione are key resistance mediators to temozolomide in glioma and melanoma cells. Oncotarget 2016, 7, 48081–48092. [Google Scholar] [CrossRef] [Green Version]
- Gross, S.R.; Kinzy, T.G. Translation elongation factor 1A is essential for regulation of the actin cytoskeleton and cell morphology. Nat. Struct. Mol. Biol. 2005, 12, 772–778. [Google Scholar] [CrossRef]
- Lei, Y.; Wen, H.; Ting, J.P. The NLR protein, NLRX1, and its partner, TUFM, reduce type I interferon, and enhance autophagy. Autophagy 2013, 9, 432–433. [Google Scholar] [CrossRef] [Green Version]
- Wells, J.; Henkler, F.; Leversha, M.; Koshy, R. A mitochondrial elongation factor-like protein is over-expressed in tumors and differentially expressed in normal tissues. FEBS Lett. 1995, 358, 119–125. [Google Scholar] [CrossRef] [Green Version]
- Lubanska, D.; Porter, L. Revisiting CDK Inhibitors for Treatment of Glioblastoma Multiforme. Drugs R D 2017, 17, 255–263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tian, Y.; Wang, C.; Chen, S.; Liu, J.; Fu, Y.; Luo, Y. Extracellular Hsp90α and clusterin synergistically promote breast cancer epithelial-to-mesenchymal transition and metastasis via LRP1. J. Cell Sci. 2019, 132, jcs228213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mu, L.; Yang, F.; Guo, D.; Li, P.; Zhang, M. Overexpression of secretory clusterin (sCLU) induces chemotherapy resistance in human gastric cancer cells by targeting miR-195-5p. Bioengineered 2020, 11, 472–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aka, J.A.; Lin, S.X. Comparison of functional proteomic analyses of human breast cancer cell lines T47D and MCF7. PLoS ONE 2012, 7, e31532. [Google Scholar] [CrossRef]
- Liang, J.; Piao, Y.; Holmes, L.; Fuller, G.N.; Henry, V.; Tiao, N.; de Groot, J.F. Neutrophils promote the malignant glioma phenotype through S100A4. Clin. Cancer Res. 2014, 20, 187–198. [Google Scholar] [CrossRef] [Green Version]
- Li, W.; Li, Y.; Li, G.; Zhou, Z.; Chang, X.; Xia, Y.; Dong, X.; Liu, Z.; Ren, B.; Liu, W.; et al. Ectopic expression of the ATP synthase β subunit on the membrane of PC-3M cells supports its potential role in prostate cancer metastasis. Int. J. Oncol. 2017, 50, 1312–1320. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| (a) Top Proteins Identified in the MV Fraction from the CM of GSCs | ||||||||
| SPOT ID | Abbr. Name | AC a Swiss/ NCBI | Protein Description | Score b | Peptide Matched | SC c % | Theoretical (pI/Mr) | p-Value |
| S53 | MOES | P26038 | Moesin | 194 | 53 | 65 | 6.08–67.89 | 0.0001 |
| S77 | GRP75 | P38646 | Stress-70 protein, mitochondrial | 111 | 49 | 55 | 5.87–73.92 | 0.0013 |
| S81 | XRCC6 | P12956 | X-ray repair cross-complementing protein 6 | 42 | 17 | 31 | 6.23–70.08 | 0.0021 |
| S84 | HSP7C | P11142 | Heat shock cognate 71 | 50 | 33 | 40 | 5.37–71.08 | 0.0011 |
| S92 | HS71A | P0DMV8 | Heat shock 70 kDa protein 1A | 66 | 21 | 36 | 5.48–70.29 | 0.0034 |
| S100 | LMNB1 | P20700 | Lamin-B1 | 76 | 26 | 36 | 5.11–66.65 | 0.0016 |
| S114 | DPYL2 | Q16555 | Dihydropyrimidinase-relatedprotein 2 | 52 | 18 | 29 | 5.95–62.71 | 0.0003 |
| S148 | VIME | P08670 | Vimentin | 129 | 55 | 76 | 5.06–53.67 | 0.0024 |
| S115 | LMNA | P02545 | Prelamin-A/C | 81 | 29 | 41 | 6.57–74.38 | 0.0008 |
| S132 | TCPZ | P40227 | T-complex protein 1 subunit zeta | 34 | 27 | 44 | 6.24–61.59 | 0.0010 |
| S153 | FKBP4 | Q02790 | Peptidyl-prolyn cis-trans isomerase | 50 | 29 | 48 | 5.35–52.05 | 0.0073 |
| S160 | AL3A1 | P30838 | Aldehyde dehydrogenase, dimeric NADP-preferring | 41 | 11 | 18 | 6.11–50.76 | 0.0001 |
| S168 | HNRH1 | P31943 | Heterogeneous nuclear ribonucleo protein H1 | 119 | 28 | 54 | 5.89–49.48 | 0.0022 |
| S169 | FKBP5 | Q13451 | Peptidyl-prolyn cis-trans isomerase | 36 | 18 | 38 | 5.70–51.69 | 0.0016 |
| S179 | GSHB | P48637 | Glutathionesynthetase | 40 | 36 | 60 | 5.67–52.52 | 0.0002 |
| S191 | EFTU | P49411 | Elongation factor Tu, mitochondrial | 119 | 31 | 68 | 7.26–49.85 | 0.0040 |
| S181 | QCR1 | P31930 | Cytochrome b-c1 complex subunit 1, mitochondrial | 165 | 34 | 61 | 5.94–53.29 | 0.0017 |
| S184 | VIME | P08670 | Vimentin | 80 | 36 | 59 | 5.06–53.67 | 0.0037 |
| S210 | CMKMT | Q7Z624 | Calmodulin-lysine N-methyltransferase | 26 | 7 | 17 | 6.37–36.78 | 0.0056 |
| S254 | ALDR | Q9UBJ2 | Aldo-keto reductase family 1 member B1 | 118 | 21 | 52 | 6.51–36.23 | 0.0001 |
| S259 | EF1D | P29692 | Elongationfactor 1-delta | 56 | 21 | 60 | 4.90–31.27 | 0.0008 |
| (b) Top Proteins Identified in the Exo Fraction from the CM of GSCs | ||||||||
| SPOT ID | Abbr. Name | AC a Swiss/ NCBI | Protein Description | Score b | Peptide Matched | SC c % | Theoretical (pI/Mr) | p-Value |
| E23 | C1S | P09871 | Complement C1s subcomponent | 63 | 27 | 42 | 4.87–99.06 | 0.0021 |
| E46 | TCPQ | P50990 | T-complex protein 1 subunit theta | 58 | 24 | 43 | 5.26–61.26 | 0.0015 |
| E48 | DPLY2 | Q16555 | Dihydropyrimidinase-related protein 2 | 115 | 32 | 66 | 5.95–62.71 | 0.0008 |
| E51 | MIPO1 | Q8TD10 | Mirror-image polydactyly gene 1 protein | 43 | 22 | 49 | 5.55–51.84 | 0.0019 |
| E54 | ATPB | P06576 | ATP synthase subunit beta, mitochondrial | 66 | 17 | 42 | 5.26–56.52 | 0.0005 |
| E120 | S10AE | Q9HCY8 | Protein S100-A14 | 30 | 5 | 50 | 5.16–11.82 | 0.0018 |
| E130 | RPB11 | P52435 | DNA-directed RNA polymerase II subunit RPB11-a | 47 | 6 | 55 | 5.63–13.34 | 0.0043 |
| E137 | CO3A1 | P02461 | Carboxy-propeptide of alpha 1 (III) procollagen | 70 | 22 | 46 | 5.89–27.90 | 0.0005 |
| E139 | CLUS | P10909 | Clusterin | 75 | 12 | 38 | 5.89–53.03 | 0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Giuseppe, F.; Carluccio, M.; Zuccarini, M.; Giuliani, P.; Ricci-Vitiani, L.; Pallini, R.; De Sanctis, P.; Di Pietro, R.; Ciccarelli, R.; Angelucci, S. Proteomic Characterization of Two Extracellular Vesicle Subtypes Isolated from Human Glioblastoma Stem Cell Secretome by Sequential Centrifugal Ultrafiltration. Biomedicines 2021, 9, 146. https://doi.org/10.3390/biomedicines9020146
Di Giuseppe F, Carluccio M, Zuccarini M, Giuliani P, Ricci-Vitiani L, Pallini R, De Sanctis P, Di Pietro R, Ciccarelli R, Angelucci S. Proteomic Characterization of Two Extracellular Vesicle Subtypes Isolated from Human Glioblastoma Stem Cell Secretome by Sequential Centrifugal Ultrafiltration. Biomedicines. 2021; 9(2):146. https://doi.org/10.3390/biomedicines9020146
Chicago/Turabian StyleDi Giuseppe, Fabrizio, Marzia Carluccio, Mariachiara Zuccarini, Patricia Giuliani, Lucia Ricci-Vitiani, Roberto Pallini, Paolo De Sanctis, Roberta Di Pietro, Renata Ciccarelli, and Stefania Angelucci. 2021. "Proteomic Characterization of Two Extracellular Vesicle Subtypes Isolated from Human Glioblastoma Stem Cell Secretome by Sequential Centrifugal Ultrafiltration" Biomedicines 9, no. 2: 146. https://doi.org/10.3390/biomedicines9020146
APA StyleDi Giuseppe, F., Carluccio, M., Zuccarini, M., Giuliani, P., Ricci-Vitiani, L., Pallini, R., De Sanctis, P., Di Pietro, R., Ciccarelli, R., & Angelucci, S. (2021). Proteomic Characterization of Two Extracellular Vesicle Subtypes Isolated from Human Glioblastoma Stem Cell Secretome by Sequential Centrifugal Ultrafiltration. Biomedicines, 9(2), 146. https://doi.org/10.3390/biomedicines9020146