
Disease-Modifying Potential of Metformin and Alendronate in an Experimental Mouse Model of Osteoarthritis



 ,
,  ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Model of Collagenase-Induced Osteoarthritis (CIOA) and Treatment Regimen
- (1)
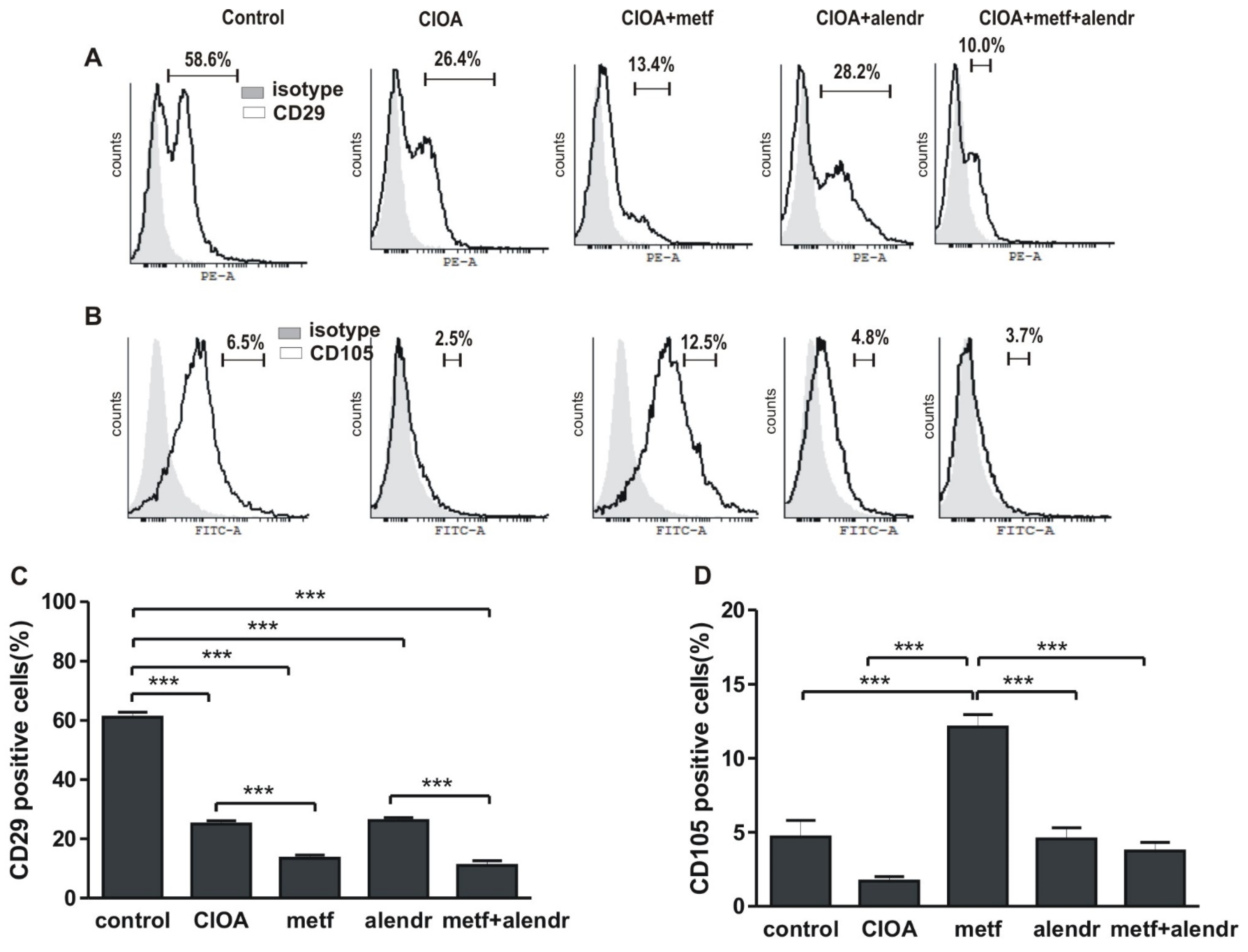
- cultivated in supplemented RPMI-1640 medium for 12 days and then subjected to FACS analysis for the determination of CD29+ and CD105+ populations;
- (2)
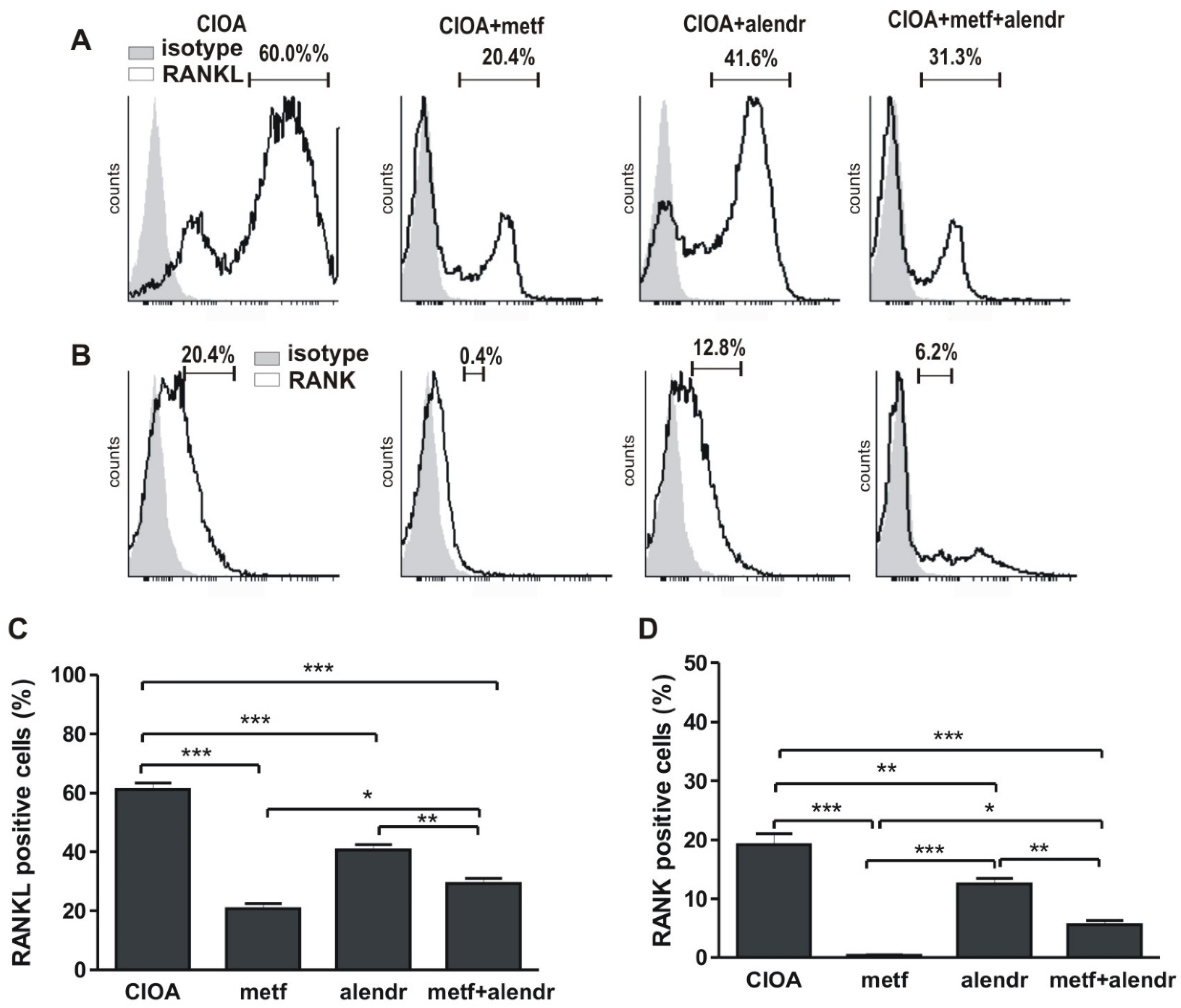
- for osteoclast differentiation, adherent cells were cultivated in supplemented RPMI-1640 medium in the presence of 10−7 M vitamin D3 (1α, 25-Dihydroxychilecalciferol, Sigma-Aldrich, Darmstadt, Germany) for 6 days and RANKL expression was determined;
- (3)
- for osteoblast differentiation, adherent cells were cultivated in supplemented α-minimal essential medium (α-MEM), containing L-ascorbic acid (50 μg/mL) and 5 mM β-glycerophosphate (Sigma-Aldrich, Darmstadt, Germany) for 14 days (mineralization medium) which was half changed every 3 days and then RANK+ cells were counted.
2.2. Flow Cytometry
2.3. ELISA Assays
2.4. Histopathological Analysis
2.5. Statistical Analysis
3. Results
3.1. Inlfuence of Metformin and Alendronate on Bone Marrow Cell Differentiation
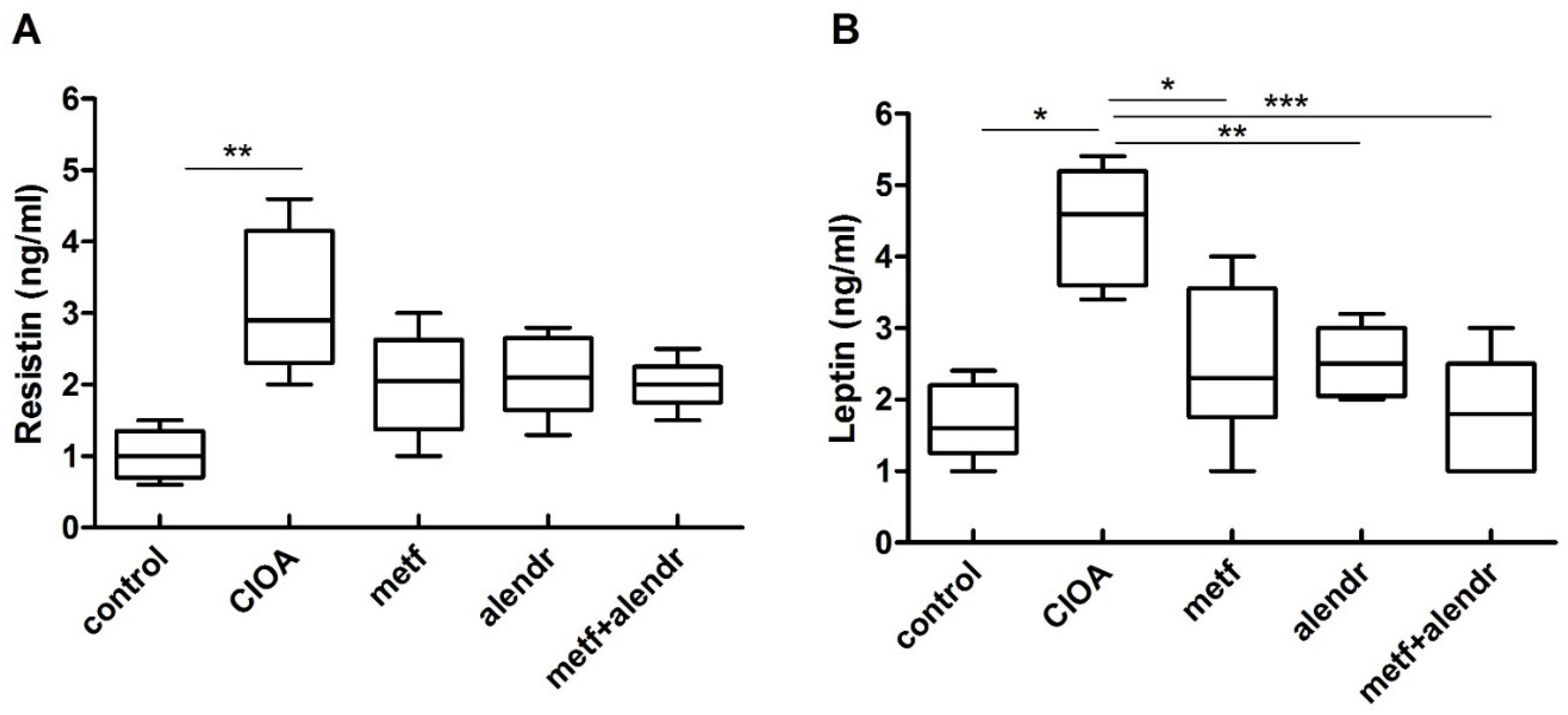
3.2. Influence of Metformin and Alendronate on Serum Resistin and Leptin Levels
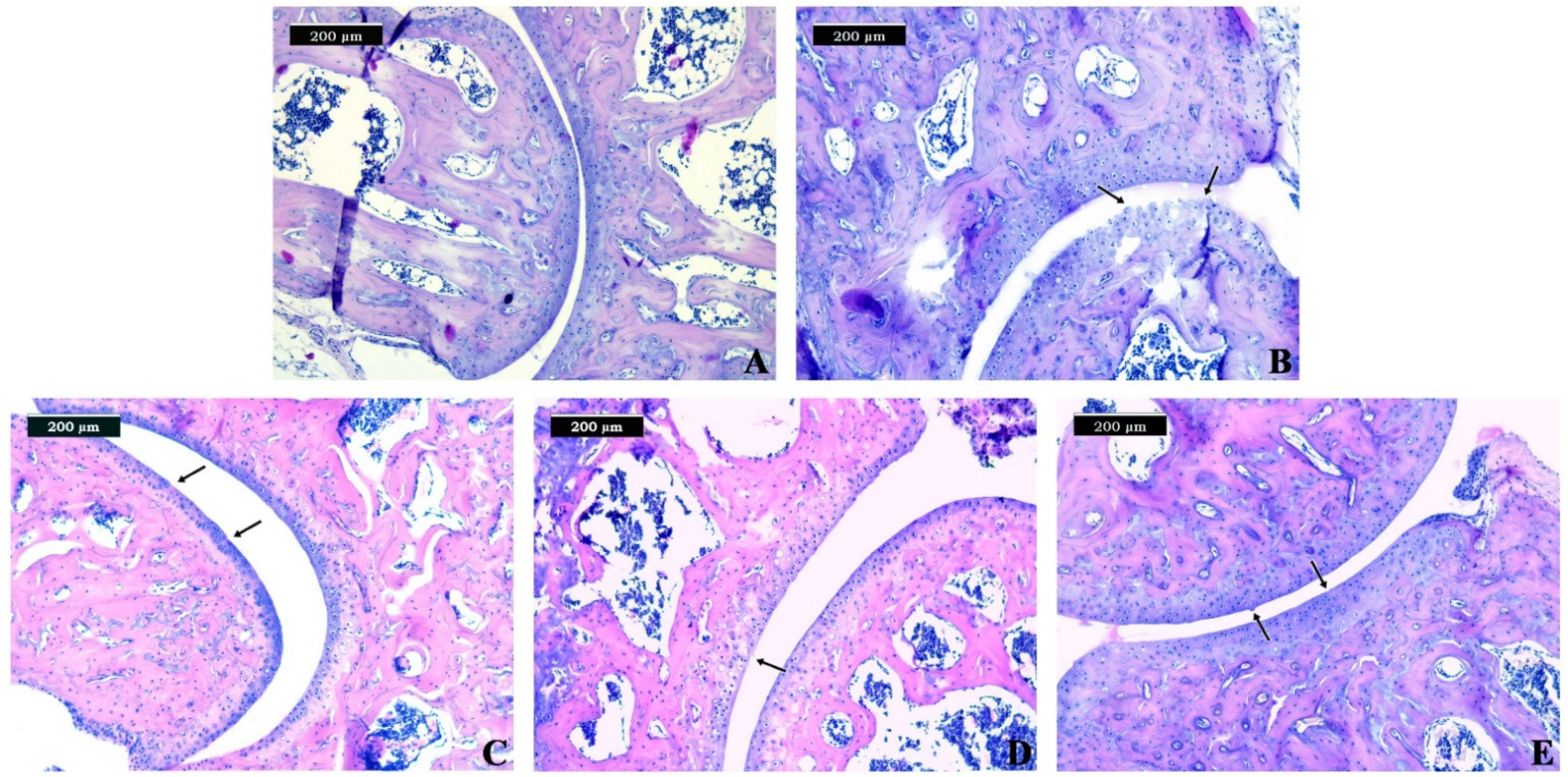
3.3. Histopathological Examination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vina, E.R.; Kwoh, C.K. Epidemiology of osteoarthritis: Literature update. Curr. Opin. Rheumatol. 2018, 30, 160–167. [Google Scholar] [CrossRef]
- Dieppe, P. Developments in osteoarthritis. Rheumatology 2011, 50, 245–247. [Google Scholar] [CrossRef] [Green Version]
- Robinson, W.H.; Lepus, C.M.; Wang, Q.; Raghu, H.; Mao, R.; Lindstrom, T.M.; Sokolove, J. Low-grade inflammation as a key mediator of the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 580–592. [Google Scholar] [CrossRef]
- Berenbaum, F. Osteoarthritis as an inflammatory disease (osteoarthritis is not osteoarthrosis!). Osteoarthr. Cartil. 2013, 21, 16–21. [Google Scholar] [CrossRef] [Green Version]
- Abramoff, B.; Caldera, F.E. Osteoarthritis: Pathology, Diagnosis, and Treatment Options. Med. Clin. N. Am. 2020, 104, 293–311. [Google Scholar] [CrossRef]
- Khorasani, M.S.; Diko, S.; Hsia, A.W.; Anderson, M.J.; Genetos, D.C.; Haudenschild, D.R.; Christiansen, B.A. Effect of alendronate on post-traumatic osteoarthritis induced by anterior cruciate ligament rupture in mice. Arthritis Res. Ther. 2015, 17, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, M.J.; Crockett, J.C.; Coxon, F.P.; Mönkkönen, J. Biochemical and molecular mechanisms of action of bisphosphonates. Bone 2011, 49, 34–41. [Google Scholar] [CrossRef]
- Shirai, T.; Kobayashi, M.; Nishitani, K.; Satake, T.; Kuroki, H.; Nakagawa, Y.; Nakamura, T. Chondroprotective effect of alendronate in a rabbit model of osteoarthritis. J. Orthop. Res. Off. Publ. Orthop. Res. Soc. 2011, 29, 1572–1577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, G.; Myers, R.; Li, Y.; Chen, Y.; Shen, X.; Fenyk-Melody, J.; Wu, M.; Ventre, J.; Doebber, T.; Fujii, N.; et al. Role of AMP-activated protein kinase in mechanism of metformin action. J. Clin. Investig. 2001, 108, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.Y.; Kim, Y.K.; Yi, H.; Kim, J.; Jung, H.R.; Kim, I.J.; Cho, J.H.; Park, S.H.; Kim, H.Y.; Ju, J.H. Metformin downregulates Th17 cells differentiation and attenuates murine autoimmune arthritis. Int. Immunopharmacol. 2013, 16, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Nath, N.; Khan, M.; Paintlia, M.K.; Singh, I.; Hoda, M.N.; Giri, S. Metformin attenuated the autoimmune disease of the central nervous system in animal models of multiple sclerosis. J. Immunol. 2009, 182, 8005–8014. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Hussain, S.M.; Wluka, A.E.; Lim, Y.Z.; Abram, F.; Pelletier, J.P.; Martel-Pelletier, J.; Cicuttini, F.M. Association between metformin use and disease progression in obese people with knee osteoarthritis: Data from the Osteoarthritis Initiative-a prospective cohort study. Arthritis Res. Ther. 2019, 21, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.; Li, H.; Guo, Z. Mesenchymal stem cell-like properties in fibroblasts. Cell Physiol. Biochem. 2014, 34, 703–714. [Google Scholar] [CrossRef]
- Soundararajan, M.; Kannan, S. Fibroblasts and mesenchymal stem cells: Two sides of the same coin? J. Cell. Physiol. 2018, 233, 9099–9109. [Google Scholar] [CrossRef] [PubMed]
- Park, M.J.; Moon, S.J.; Baek, J.A.; Lee, E.J.; Jung, K.A.; Kim, E.K.; Kim, D.S.; Lee, J.H.; Kwok, S.K.; Min, J.K.; et al. Metformin Augments Anti-Inflammatory and Chondroprotective Properties of Mesenchymal Stem Cells in Experimental Osteoarthritis. J. Immunol. 2019, 203, 127–136. [Google Scholar] [CrossRef] [PubMed]
- Carrión, M.; Frommer, K.W.; Pérez-García, S.; Müller-Ladner, U.; Gomariz, R.P.; Neumann, E. The Adipokine Network in Rheumatic Joint Diseases. Int. J. Mol. Sci. 2019, 20, 4091. [Google Scholar] [CrossRef] [Green Version]
- Azamar-Llamas, D.; Hernandez-Molina, G.; Ramos-Avalos, B.; Furuzawa-Carballeda, J. Adipokine Contribution to the Pathogenesis of Osteoarthritis. Mediat. Inflamm. 2017, 2017, 5468023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vuolteenaho, K.; Koskinen, A.; Moilanen, E. Leptin—A Link between Obesity and Osteoarthritis. Applications for Prevention and Treatment. Basic Clin. Pharmacol. Toxicol. 2014, 114, 103–108. [Google Scholar] [CrossRef]
- Zhang, P.; Zhong, Z.H.; Yu, H.T.; Liu, B. Significance of increased leptin expression in osteoarthritis patients. PLoS ONE 2015, 10, e0123224. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Zhang, J.; Yang, H.; Sun, Y. The role of leptin in osteoarthritis. Medicine 2018, 97, e0257. [Google Scholar] [CrossRef] [PubMed]
- Doherty, A.L.; Battaglino, R.A.; Donovan, J.; Gagnon, D.; Lazzari, A.A.; Garshick, E.; Zafonte, R.; Morse, L.R. Adiponectin is a candidate biomarker of lower extremity bone density in men with chronic spinal cord injury. J. Bone Min. Res 2014, 29, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Song, Y.Z.; Guan, J.; Wang, H.J.; Ma, W.; Li, F.; Xu, F.; Ding, L.B.; Xie, L.; Liu, B.; Liu, K.; et al. Possible Involvement of Serum and Synovial Fluid Resistin in Knee Osteoarthritis: Cartilage Damage, Clinical, and Radiological Links. J. Clin. Lab. Anal. 2016, 30, 437–443. [Google Scholar] [CrossRef]
- Koskinen, A.; Vuolteenaho, K.; Moilanen, T.; Moilanen, E. Resistin as a factor in osteoarthritis: Synovial fluid resistin concentrations correlate positively with interleukin 6 and matrix metalloproteinases MMP-1 and MMP-3. Scand. J. Rheumatol. 2014, 43, 249–253. [Google Scholar] [CrossRef]
- Cheleschi, S.; Giordano, N.; Volpi, N.; Tenti, S.; Gallo, I.; Di Meglio, M.; Giannotti, S.; Fioravanti, A. A Complex Relationship between Visfatin and Resistin and microRNA: An In Vitro Study on Human Chondrocyte Cultures. Int. J. Mol. Sci. 2018, 19, 3909. [Google Scholar] [CrossRef] [Green Version]
- Chen, W.C.; Wang, S.W.; Lin, C.Y.; Tsai, C.H.; Fong, Y.C.; Lin, T.Y.; Weng, S.L.; Huang, H.D.; Liao, K.W.; Tang, C.H. Resistin Enhances Monocyte Chemoattractant Protein-1 Production in Human Synovial Fibroblasts and Facilitates Monocyte Migration. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2019, 52, 408–420. [Google Scholar] [CrossRef]
- Zhang, Z.; Xing, X.; Hensley, G.; Chang, L.W.; Liao, W.; Abu-Amer, Y.; Sandell, L.J. Resistin induces expression of proinflammatory cytokines and chemokines in human articular chondrocytes via transcription and messenger RNA stabilization. Arthritis Rheum 2010, 62, 1993–2003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Xu, J.; Cai, J.; Zheng, S.; Yang, X.; Ding, C. Serum levels of resistin and interleukin-17 are associated with increased cartilage defects and bone marrow lesions in patients with knee osteoarthritis. Mod. Rheumatol. 2017, 27, 339–344. [Google Scholar] [CrossRef] [PubMed]
- Gyurkovska, V.; Stefanova, T.; Dimitrova, P.; Danova, S.; Tropcheva, R.; Ivanovska, N. Tyrosine kinase inhibitor tyrphostin AG490 retards chronic joint inflammation in mice. Inflammation 2014, 37, 995–1005. [Google Scholar] [CrossRef] [PubMed]
- Glasson, S.S.; Chambers, M.G.; Van Den Berg, W.B.; Little, C.B. The OARSI histopathology initiative–recommendations for histological assessments of osteoarthritis in the mouse. Osteoarthr. Cartil. 2010, 18, S17–S23. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, N.; Laverty, S.; Kraus, V.B.; Aigner, T. Basic methods in histopathology of joint tissues. Osteoarthr. Cartil. 2010, 18, S113–S116. [Google Scholar] [CrossRef] [Green Version]
- Pritzker, K.P.; Gay, S.; Jimenez, S.A.; Ostergaard, K.; Pelletier, J.P.; Revell, P.A.; Salter, D.; van den Berg, W.B. Osteoarthritis cartilage histopathology: Grading and staging. Osteoarthr. Cartil. 2006, 14, 13–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhang, B.; Liu, W.X.; Lu, K.; Pan, H.; Wang, T.; Oh, C.D.; Yi, D.; Huang, J.; Zhao, L.; et al. Metformin limits osteoarthritis development and progression through activation of AMPK signalling. Ann. Rheum. Dis. 2020, 79, 635–645. [Google Scholar] [CrossRef] [Green Version]
- Na, H.S.; Kwon, J.Y.; Lee, S.Y.; Lee, S.H.; Lee, A.R.; Woo, J.S.; Jung, K.; Cho, K.H.; Choi, J.W.; Lee, D.H.; et al. Metformin Attenuates Monosodium-Iodoacetate-Induced Osteoarthritis via Regulation of Pain Mediators and the Autophagy-Lysosomal Pathway. Cells 2021, 10, 681. [Google Scholar] [CrossRef] [PubMed]
- Barnett, L.A.; Jordan, K.P.; Edwards, J.J.; van der Windt, D.A. Does metformin protect against osteoarthritis? An electronic health record cohort study. Prim. Health Care Res. Dev. 2017, 18, 623–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, F.; Wang, H.; Yuan, F.; Ding, Y. Platelet-Rich Plasma Combined with Alendronate Reduces Pain and Inflammation in Induced Osteoarthritis in Rats by Inhibiting the Nuclear Factor-Kappa B Signaling Pathway. BioMed Res. Int. 2020, 2020, 8070295. [Google Scholar] [CrossRef]
- Lofvall, H.; Newbould, H.; Karsdal, M.A.; Dziegiel, M.H.; Richter, J.; Henriksen, K.; Thudium, C.S. Osteoclasts degrade bone and cartilage knee joint compartments through different resorption processes. Arthritis Res. Ther. 2018, 20, 67. [Google Scholar] [CrossRef] [Green Version]
- Bertuglia, A.; Lacourt, M.; Girard, C.; Beauchamp, G.; Richard, H.; Laverty, S. Osteoclasts are recruited to the subchondral bone in naturally occurring post-traumatic equine carpal osteoarthritis and may contribute to cartilage degradation. Osteoarthr. Cartil. 2016, 24, 555–566. [Google Scholar] [CrossRef] [Green Version]
- Ku, J.H.; Lee, C.K.; Joo, B.S.; An, B.M.; Choi, S.H.; Wang, T.H.; Cho, H.L. Correlation of synovial fluid leptin concentrations with the severity of osteoarthritis. Clin. Rheumatol. 2009, 28, 1431–1435. [Google Scholar] [CrossRef]
- Bao, J.P.; Chen, W.P.; Feng, J.; Hu, P.F.; Shi, Z.L.; Wu, L.D. Leptin plays a catabolic role on articular cartilage. Mol. Biol. Rep. 2010, 37, 3265–3272. [Google Scholar] [CrossRef]
- Mutabaruka, M.S.; Aoulad Aissa, M.; Delalandre, A.; Lavigne, M.; Lajeunesse, D. Local leptin production in osteoarthritis subchondral osteoblasts may be responsible for their abnormal phenotypic expression. Arthritis Res. Ther. 2010, 12, R20. [Google Scholar] [CrossRef] [Green Version]
- Wyatt, L.A.; Moreton, B.J.; Mapp, P.I.; Wilson, D.; Hill, R.; Ferguson, E.; Scammell, B.E.; Walsh, D.A. Histopathological subgroups in knee osteoarthritis. Osteoarthr. Cartil. 2017, 25, 14–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Groups | Grade | Associated Criteria (Tissue Reaction) |
|---|---|---|
| Group 1 (healthy control) | Grade 0 Surface intact, cartilage morphology intact | Normal architecture; the matrix and chondrocytes are organized into superficial, mid, and deep zones. |
| Group 2 (CIOA) | Grade 3 Vertical fissures (clefts) | Vertical fissures extending into the mid zone, the matrix fibrillation extends vertically downward into the mid zone; cell death and proliferation may be observed most prominently adjacent to fissures. |
| Group 3 (CIOA + metformin) | Grade 1 Surface intact | The articular surface is uneven and can demonstrate superficial fibrillation; this may be accompanied by cell death or proliferation; the mid zone and deep zone are unaffected. |
| Group 4 (CIOA + alendronate) | Grade 2 Surface discontinuity | Focally fibrillation extends through the superficial zone to the superficial zone–mid zone portion; this may be accompanied by cell proliferation, decreased matrix staining, and cell death in the mid zone. |
| Group 5 (CIOA + metformin + alendronate) | Grade 1 Surface intact | The articular surface is uneven and can demonstrate superficial fibrillation; this may be accompanied by cell death or proliferation; the mid zone and deep zone are unaffected. |
| Test Groups | Stage | % Involvement (Surface, Area, Volume) | Grade | OA Score |
|---|---|---|---|---|
| Group 1 (healthy control) | Stage 0 | No OA activity seen | Grade 0 | 0 |
| Group 2 (CIOA) | Stage 3 | 25–50% | Grade 3 | 9 |
| Group 3 (CIOA + metformin) | Stage 4 | >50% | Grade 1 | 4 |
| Group 4 (CIOA + alendronate) | Stage 3 | 25–50% | Grade 2 | 6 |
| Group 5 (CIOA + metformin + alendronate) | Stage 2 | 10–25% | Grade 1 | 2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Belenska-Todorova, L.; Lambova, S.N.; Stoyanova, S.; Georgieva, E.; Batsalova, T.; Moten, D.; Kolchakova, D.; Dzhambazov, B. Disease-Modifying Potential of Metformin and Alendronate in an Experimental Mouse Model of Osteoarthritis. Biomedicines 2021, 9, 1017. https://doi.org/10.3390/biomedicines9081017
Belenska-Todorova L, Lambova SN, Stoyanova S, Georgieva E, Batsalova T, Moten D, Kolchakova D, Dzhambazov B. Disease-Modifying Potential of Metformin and Alendronate in an Experimental Mouse Model of Osteoarthritis. Biomedicines. 2021; 9(8):1017. https://doi.org/10.3390/biomedicines9081017
Chicago/Turabian StyleBelenska-Todorova, Lyudmila, Sevdalina Nikolova Lambova, Stela Stoyanova, Elenka Georgieva, Tsvetelina Batsalova, Dzhemal Moten, Desislava Kolchakova, and Balik Dzhambazov. 2021. "Disease-Modifying Potential of Metformin and Alendronate in an Experimental Mouse Model of Osteoarthritis" Biomedicines 9, no. 8: 1017. https://doi.org/10.3390/biomedicines9081017
APA StyleBelenska-Todorova, L., Lambova, S. N., Stoyanova, S., Georgieva, E., Batsalova, T., Moten, D., Kolchakova, D., & Dzhambazov, B. (2021). Disease-Modifying Potential of Metformin and Alendronate in an Experimental Mouse Model of Osteoarthritis. Biomedicines, 9(8), 1017. https://doi.org/10.3390/biomedicines9081017