

Possible Processes and Mechanisms of Hexachlorobenzene Decomposition by the Selected Comamonas testosteroni Bacterial Strains




Abstract
:1. Introduction
2. Materials and Methods
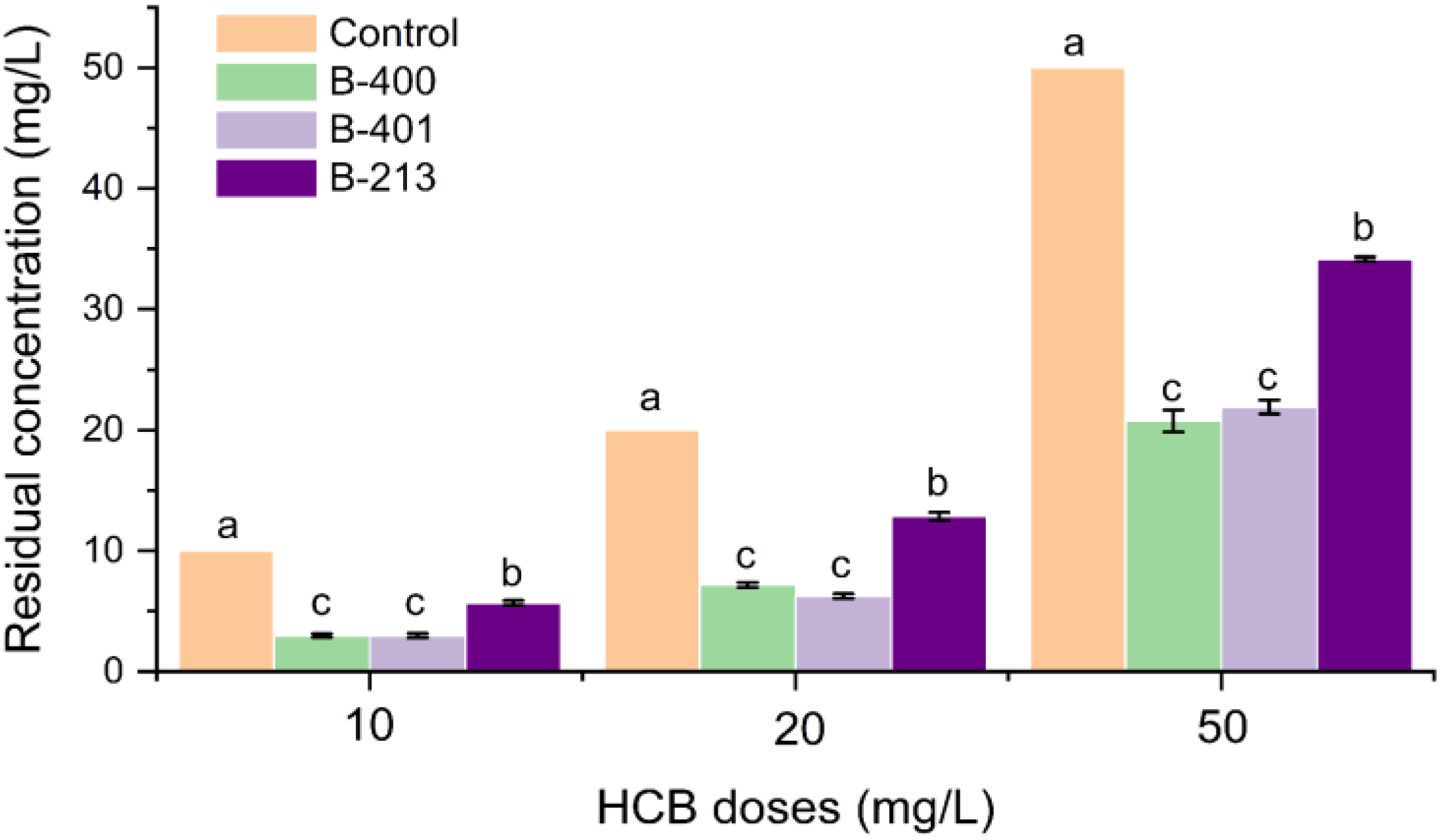
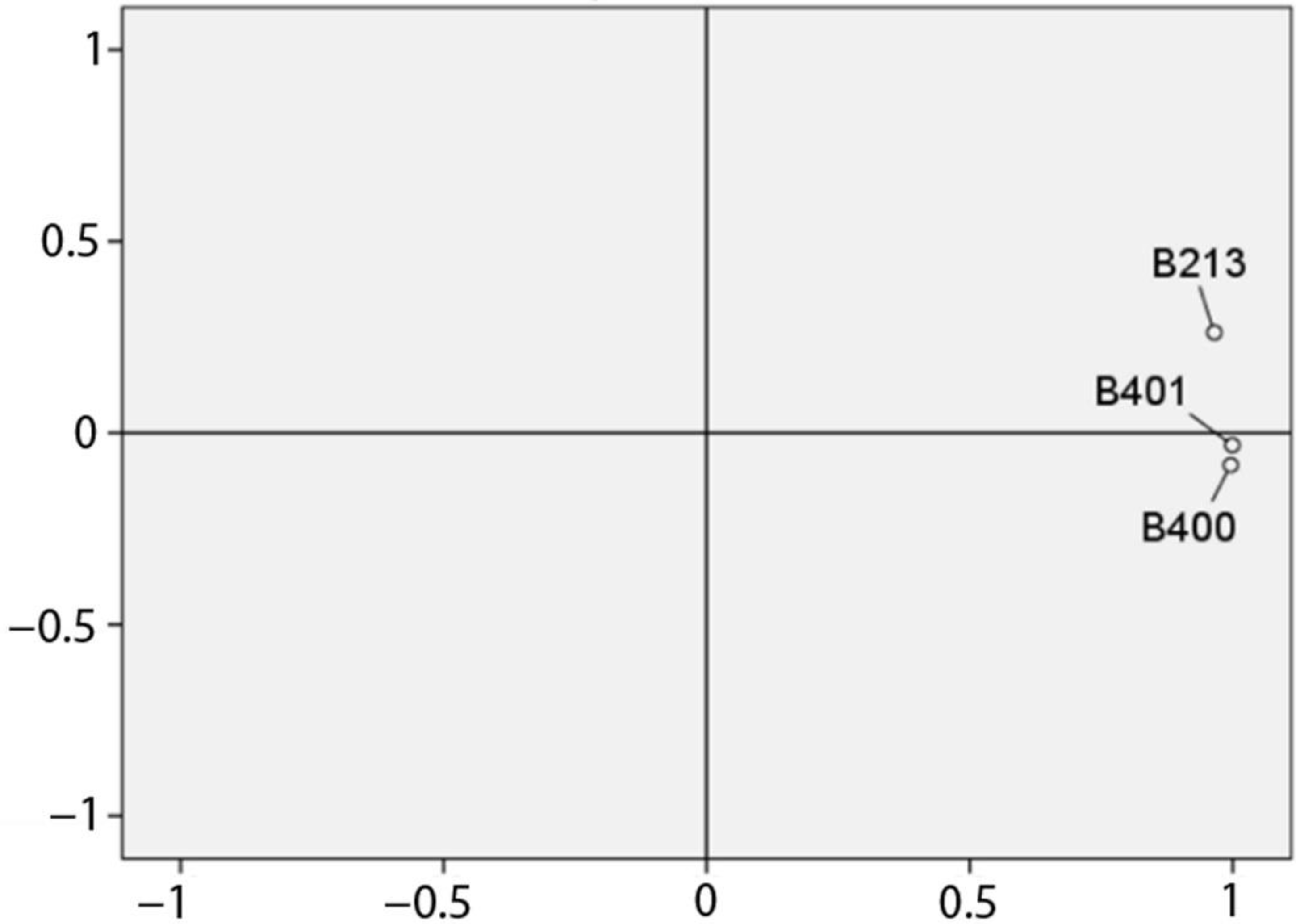
3. Results
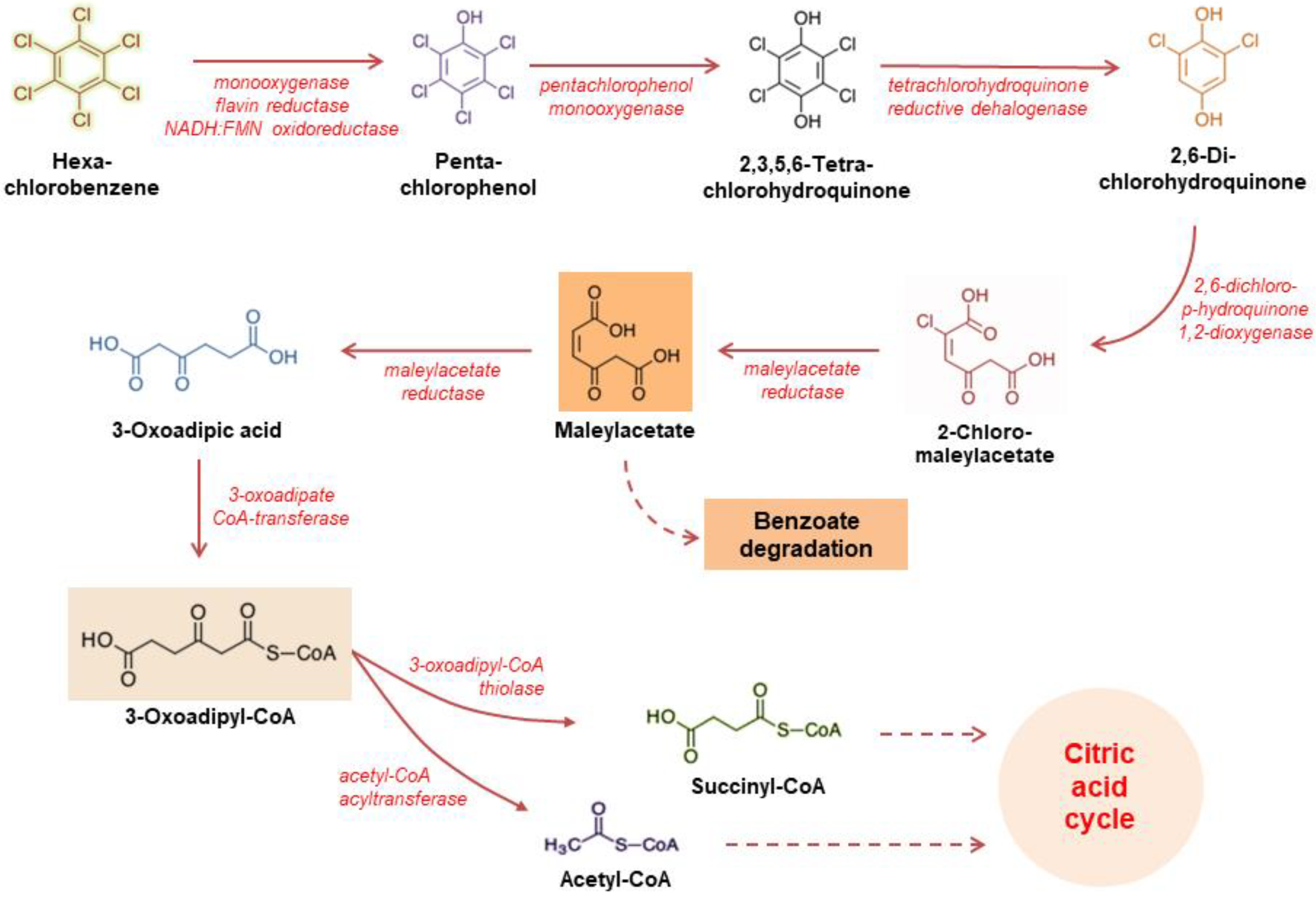
4. Discussion

5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sun, G.; Du, Y.; Yin, J.; Jiang, Y.; Zhang, D.; Jiang, B.; Li, G.; Wang, H.; Kong, F.; Su, L.; et al. Response of Microbial Communities to Different Organochlorine Pesticides (OCPs) Contamination Levels in Contaminated Soils. Chemosphere 2019, 215, 461–469. [Google Scholar] [CrossRef]
- Raffa, C.M.; Chiampo, F. Bioremediation of Agricultural Soils Polluted with Pesticides: A Review. Bioengineering 2021, 8, 92. [Google Scholar] [CrossRef] [PubMed]
- Stockholm Convention on Persistent Organic Pollutants (POPs); Secretariat of the Stockholm Convention (SSC): Stockholm, Switzerland, 2020; Volume 2020.
- EPA—United States Environmental Protection Agency 2022. Available online: https://www.epa.gov/ (accessed on 26 September 2022).
- Olisah, C.; Okoh, O.O.; Okoh, A.I. Occurrence of Organochlorine Pesticide Residues in Biological and Environmental Matrices in Africa: A Two-Decade Review. Heliyon 2020, 6, e03518. [Google Scholar] [CrossRef]
- Garbou, A.M.; Liu, M.; Zou, S.; Yestrebsky, C.L. Degradation Kinetics of Hexachlorobenzene over Zero-Valent Magnesium/Graphite in Protic Solvent System and Modeling of Degradation Pathways Using Density Functional Theory. Chemosphere 2019, 222, 195–204. [Google Scholar] [CrossRef]
- Kumar, M.; Dinesh, K.; Kubendran, D.; Kalaichelvan, P.T. Hexachlorobenzene—Sources, Remediation and Future Prospects. Int. J. Curr. Res. Rev. 2013, 2013, 1–12. [Google Scholar]
- Pouch, A.; Zaborska, A.; Pazdro, K. The History of Hexachlorobenzene Accumulation in Svalbard Fjords. Env. Monit Assess 2018, 190, 360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lovecka, P.; Pacovska, I.; Stursa, P.; Vrchotova, B.; Kochankova, L.; Demnerova, K. Organochlorinated Pesticide Degrading Microorganisms Isolated from Contaminated Soil. New Biotechnol. 2015, 32, 26–31. [Google Scholar] [CrossRef]
- Wang, H.; Cao, X.; Li, L.; Fang, Z.; Li, X. Augmenting Atrazine and Hexachlorobenzene Degradation under Different Soil Redox Conditions in a Bioelectrochemistry System and an Analysis of the Relevant Microorganisms. Ecotoxicol. Environ. Saf. 2018, 147, 735–741. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Claeys, L.; van der Ha, D.; Verstraete, W.; Boon, N. Effects of Chemically and Electrochemically Dosed Chlorine on Escherichia Coli and Legionella Beliardensis Assessed by Flow Cytometry. Appl. Microbiol. Biotechnol. 2010, 87, 331–341. [Google Scholar] [CrossRef]
- Andersen, M.S.; Fuglei, E.; König, M.; Lipasti, I.; Pedersen, Å.Ø.; Polder, A.; Yoccoz, N.G.; Routti, H. Levels and Temporal Trends of Persistent Organic Pollutants (POPs) in Arctic Foxes (Vulpes Lagopus) from Svalbard in Relation to Dietary Habits and Food Availability. Sci. Total Environ. 2015, 511, 112–122. [Google Scholar] [CrossRef]
- Corsolini, S. Industrial Contaminants in Antarctic Biota. J. Chromatogr. A 2009, 1216, 598–612. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Li, Y.; Zhang, W.; Wang, F.; Bian, Y.; Boughner, L.A.; Jiang, X. Novel Biochar-Plant Tandem Approach for Remediating Hexachlorobenzene Contaminated Soils: Proof-of-Concept and New Insight into the Rhizosphere. J. Agric. Food Chem. 2016, 64, 5464–5471. [Google Scholar] [CrossRef]
- Jayaraj, R.; Megha, P.; Sreedev, P. Review Article. Organochlorine Pesticides, Their Toxic Effects on Living Organisms and Their Fate in the Environment. Interdiscip. Toxicol. 2016, 9, 90–100. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.A. Persistent Organic Pollutants (POPs): A Global Issue, a Global Challenge. Env. Sci. Pollut. Res. 2017, 24, 4223–4227. [Google Scholar] [CrossRef]
- Chávez-Almazán, L.A.; Diaz-Ortiz, J.; Alarcón-Romero, M.; Dávila-Vazquez, G.; Saldarriaga-Noreña, H.; Waliszewski, S.M. Organochlorine Pesticide Levels in Breast Milk in Guerrero, Mexico. Bull. Env. Contam. Toxicol. 2014, 93, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Jin, Y.; Cheng, Y.; Leaderer, B.; Lin, S.; Holford, T.R.; Qiu, J.; Zhang, Y.; Shi, K.; Zhu, Y.; et al. Prenatal Exposure to Organochlorine Pesticides and Infant Birth Weight in China. Chemosphere 2014, 110, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Cheng, Y.; Tang, Q.; Lin, S.; Li, Y.; Hu, X.; Nian, J.; Gu, H.; Lu, Y.; Tang, H.; et al. The Association between Prenatal Exposure to Organochlorine Pesticides and Thyroid Hormone Levels in Newborns in Yancheng, China. Environ. Res. 2014, 129, 47–51. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, B.; Dai, X.; Li, S.; Lu, G.; Zhou, Y. Structure and Function of the Bacterial Communities during Rhizoremediation of Hexachlorobenzene in Constructed Wetlands. Env. Sci. Pollut. Res. 2017, 24, 11483–11492. [Google Scholar] [CrossRef] [PubMed]
- Feng, Q.; Liang, S.; Jia, H.; Stadlmayr, A.; Tang, L.; Lan, Z.; Zhang, D.; Xia, H.; Xu, X.; Jie, Z.; et al. Gut Microbiome Development along the Colorectal Adenoma–Carcinoma Sequence. Nat. Commun. 2015, 6, 6528. [Google Scholar] [CrossRef] [Green Version]
- Kushkevych, I.; Dordević, D.; Vítězová, M. Toxicity of Hydrogen Sulfide toward Sulfate-Reducing Bacteria Desulfovibrio Piger Vib-7. Arch Microbiol 2019, 201, 389–397. [Google Scholar] [CrossRef]
- Kushkevych, I.; Dordević, D.; Kollar, P.; Vítězová, M.; Drago, L. Hydrogen Sulfide as a Toxic Product in the Small–Large Intestine Axis and Its Role in IBD Development. JCM 2019, 8, 1054. [Google Scholar] [CrossRef] [PubMed]
- Kushkevych, I.; Dordević, D.; Vítězová, M. Possible Synergy Effect of Hydrogen Sulfide and Acetate Produced by Sulfate-Reducing Bacteria on Inflammatory Bowel Disease Development. J. Adv. Res. 2020, 27, 71–78. [Google Scholar] [CrossRef] [PubMed]
- Dordević, D.; Jančíková, S.; Vítězová, M.; Kushkevych, I. Hydrogen Sulfide Toxicity in the Gut Environment: Meta-Analysis of Sulfate-Reducing and Lactic Acid Bacteria in Inflammatory Processes. J. Adv. Res. 2020, 27, 55–69. [Google Scholar] [CrossRef]
- Prashar, P.; Shah, S. Impact of Fertilizers and Pesticides on Soil Microflora in Agriculture. In Sustainable Agriculture Reviews; Lichtfouse, E., Ed.; Sustainable Agriculture Reviews; Springer International Publishing: Cham, Switzerland, 2016; Volume 19, pp. 331–361. ISBN 978-3-319-26776-0. [Google Scholar]
- Hussain, S.; Siddique, T.; Saleem, M.; Arshad, M.; Khalid, A. Chapter 5 Impact of Pesticides on Soil Microbial Diversity, Enzymes, and Biochemical Reactions. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2009; Volume 102, pp. 159–200. ISBN 978-0-12-374818-8. [Google Scholar]
- Imfeld, G.; Vuilleumier, S. Measuring the Effects of Pesticides on Bacterial Communities in Soil: A Critical Review. Eur. J. Soil Biol. 2012, 49, 22–30. [Google Scholar] [CrossRef]
- Merlin, C.; Devers, M.; Béguet, J.; Boggio, B.; Rouard, N.; Martin-Laurent, F. Evaluation of the Ecotoxicological Impact of the Organochlorine Chlordecone on Soil Microbial Community Structure, Abundance, and Function. Env. Sci. Pollut. Res. 2016, 23, 4185–4198. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Ishida, T.; Oh, K.; Sudo, R. Biodegradation of Chlordane and Hexachlorobenzenes in River Sediment. Chemosphere 2007, 67, 428–434. [Google Scholar] [CrossRef]
- Yan, D.-Z.; Mao, L.-Q.; Li, C.-Z.; Liu, J. Biodegradation of Hexachlorobenzene by a Constructed Microbial Consortium. World J Microbiol Biotechnol 2015, 31, 371–377. [Google Scholar] [CrossRef]
- Jayachandran, G.; Gorisch, H.; Adrian, L. Dehalorespiration with Hexachlorobenzene and Pentachlorobenzene by Dehalococcoides sp. Strain CBDB1. Arch. Microbiol. 2003, 180, 411–416. [Google Scholar] [CrossRef]
- Regulation (EU) 2019/1021 of the European Parliament and of the Council of 20 June 2019 on Persistent Organic Pollutants (POPs) 2021. Available online: https://lexparency.org/eu/32019R1021/ (accessed on 26 September 2022).
- Chow, S.J.; Lorah, M.M.; Wadhawan, A.R.; Durant, N.D.; Bouwer, E.J. Sequential Biodegradation of 1,2,4-Trichlorobenzene at Oxic-Anoxic Groundwater Interfaces in Model Laboratory Columns. J. Contam. Hydrol. 2020, 231, 103639. [Google Scholar] [CrossRef]
- Wiltschka, K.; Neumann, L.; Werheid, M.; Bunge, M.; Düring, R.-A.; Mackenzie, K.; Böhm, L. Hydrodechlorination of Hexachlorobenzene in a Miniaturized Nano-Pd(0) Reaction System Combined with the Simultaneous Extraction of All Dechlorination Products. Appl. Catal. B Environ. 2020, 275, 119100. [Google Scholar] [CrossRef]
- Ito, K.; Takagi, K.; Matsushima, Y.; Iwasaki, A.; Tanaka, N.; Kanesaki, Y.; Martin-Laurent, F.; Igimi, S. Identification of the Novel HcbB Operon Catalyzing the Dechlorination of Pentachlorophenol in the Gram-Positive Bacterium Nocardioides sp. Strain PD653. J. Pestic. Sci. 2018, 43, 124–131. [Google Scholar] [CrossRef]
- Takagi, K.; Iwasaki, A.; Kamei, I.; Satsuma, K.; Yoshioka, Y.; Harada, N. Aerobic Mineralization of Hexachlorobenzene by Newly Isolated Pentachloronitrobenzene-Degrading Nocardioides sp. Strain PD653. Appl. Env. Microbiol. 2009, 75, 4452–4458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dimova, M.; Dankevych, L.; Yamborko, N.; Iutynska, G. Polyphasic Taxonomy Analyse of Comamonas Testosteroni Resistant to Hexachlorobenzene. J. Microb. Biotech. Food Sci. 2022, 11, e4711. [Google Scholar] [CrossRef]
- Arai, H.; Akahira, S.; Ohishi, T.; Maeda, M.; Kudo, T. Adaptation of Cornamonas Testosteroni TAM1 to Utilize Phenol: Organization and Regulation of the Genes Involved in Phenol Degradatio. Microbiology 1998, 144, 2895–2903. [Google Scholar] [CrossRef]
- Yap, L.F.; Lee, Y.K.; Poh, C.L. Mechanism for Phenol Tolerance in Phenol-Degrading Comamonas Testosteroni Strain. Appl. Microbiol. Biotechnol. 1999, 51, 833–840. [Google Scholar] [CrossRef]
- Liu, T.; Chen, Z.; Shen, Y. Aerobic Biodegradation of Hexachlorobenzene by an Acclimated Microbial Community. IJEP 2009, 37, 235. [Google Scholar] [CrossRef]
- Uhlik, O.; Strejcek, M.; Vondracek, J.; Musilova, L.; Ridl, J.; Lovecka, P.; Macek, T. Bacterial Acquisition of Hexachlorobenzene-Derived Carbon in Contaminated Soil. Chemosphere 2014, 113, 141–145. [Google Scholar] [CrossRef]
- Jones, J.P.; O’Hare, E.J.; Wong, L.-L. Oxidation of Polychlorinated Benzenes by Genetically Engineered CYP101 (Cytochrome P450cam): Oxidation of Polychlorinated Benzenes by CYP101. Eur. J. Biochem. 2001, 268, 1460–1467. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Christopher, A.; Jones, J.P.; Bell, S.G.; Guo, Q.; Xu, F.; Rao, Z.; Wong, L.-L. Crystal Structure of the F87W/Y96F/V247L Mutant of Cytochrome P-450cam with 1,3,5-Trichlorobenzene Bound and Further Protein Engineering for the Oxidation of Pentachlorobenzene and Hexachlorobenzene. J. Biol. Chem. 2002, 277, 37519–37526. [Google Scholar] [CrossRef] [Green Version]
- Cai, M.; Xun, L. Organization and Regulation of Pentachlorophenol-Degrading Genes in Sphingobium Chlorophenolicum ATCC 39723. J. Bacteriol. 2002, 184, 4672–4680. [Google Scholar] [CrossRef] [Green Version]
- Liu, L.; Jiang, C.-Y.; Liu, X.-Y.; Wu, J.-F.; Han, J.-G.; Liu, S.-J. Plant-Microbe Association for Rhizoremediation of Chloronitroaromatic Pollutants with Comamonas sp. Strain CNB-1. Env. Microbiol. 2007, 9, 465–473. [Google Scholar] [CrossRef]
- Wu, J.; Jiang, C.; Wang, B.; Ma, Y.; Liu, Z.; Liu, S. Novel Partial Reductive Pathway for 4-Chloronitrobenzene and Nitrobenzene Degradation in Comamonas sp. Strain CNB-1. Appl. Env. Microbiol. 2006, 72, 1759–1765. [Google Scholar] [CrossRef] [Green Version]
- Dimova, M.; Tugai, A.; Tugai, T.; Iutynska, G.; Dordevic, D.; Kushkevych, I. Molecular Research of Lipid Peroxidation and Antioxidant Enzyme Activity of Comamonas Testosteroni Bacterial Cells under the Hexachlorobenzene Impact. IJMS 2022, 23, 11415. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wu, J.-F.; Zeyer, J.; Meng, B.; Liu, L.; Jiang, C.-Y.; Liu, S.-Q.; Liu, S.-J. Proteomic and Molecular Investigation on the Physiological Adaptation of Comamonas sp. Strain CNB-1 Growing on 4-Chloronitrobenzene. Biodegradation 2009, 20, 55–66. [Google Scholar] [CrossRef]
- Yong, Y.-C.; Zhong, J.-J. Recent Advances in Biodegradation in China: New Microorganisms and Pathways, Biodegradation Engineering, and Bioenergy from Pollutant Biodegradation. Process. Biochem. 2010, 45, 1937–1943. [Google Scholar] [CrossRef]
- Aráoz, B.; Viale, A.A. Microbial Dehalogenation of Polychlorinated Biphenyls in Aerobic Conditions. Rev. Argent. Microbiol. 2004, 36, 47–51. [Google Scholar]
- Tan, W.A.; Parales, R.E. Hydrocarbon Degradation by Betaproteobacteria. In Taxonomy, Genomics and Ecophysiology of Hydrocarbon-Degrading Microbes; McGenity, T.J., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 125–141. ISBN 978-3-030-14795-2. [Google Scholar]
- Vítková, M.; Dercová, K.; Molnárová, J.; Tóthová, L.; Polek, B.; Godočíková, J. The Effect of Lignite and Comamonas Testosteroni on Pentachlorophenol Biodegradation and Soil Ecotoxicity. Water Air Soil Pollut. 2011, 218, 145–155. [Google Scholar] [CrossRef]
- Tobajas, M.; Monsalvo, V.M.; Mohedano, A.F.; Rodriguez, J.J. Enhancement of Cometabolic Biodegradation of 4-Chlorophenol Induced with Phenol and Glucose as Carbon Sources by Comamonas Testosteroni. J. Environ. Manag. 2012, 95, S116–S121. [Google Scholar] [CrossRef]
- Wheelis, M.L.; Palleroni, N.J.; Stanier, R.Y. The Metabolism of Aromatic Acids by Pseudomonas Testosteroni and P. Acidovorans. Archiv. Mikrobiol. 1967, 59, 302–314. [Google Scholar] [CrossRef]
- Baumberg, S. Genetics and Biochemistry of Pseudomonas. Biochem. Educ. 1975, 3, 46. [Google Scholar] [CrossRef]
- Hegeman, G. The Mineralization of Organic Materials under Aerobic Conditions. In Bacteria in Nature; Leadbetter, E.R., Poindexter, J.S., Eds.; Springer US: Boston, MA, USA, 1985; pp. 97–112. ISBN 978-1-4615-6513-0. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
| Conditions | Cultivations | Acetone Volume (mL) | Hexachlorobenzene (mg/L) |
|---|---|---|---|
| (a) control | cultivating the strain in LB medium contained acetone volume required to dissolve | no | Pure control |
| (b) control (10) | 20 | 10 | |
| (c) control (20) | 40 | 20 | |
| (d) control (50) | 100 | 50 | |
| (e) cultivating | in LB medium contained | 20 | 10 |
| (f) cultivating | 40 | 20 | |
| (g) cultivating | 100 | 50 |
| Variants | C. testosteroni Strains | ||
|---|---|---|---|
| UCM B-400 | UCM B-401 | UCM B-213 | |
| Control | 1.50 ± 0.30 × 109 a | 1.77 ± 0.21 × 109 a | 9.03 ± 0.35 × 108 a |
| Control (10) | 7.67 ± 1.50 × 108 ab | 1.00 ± 0.10 × 109 a | 7.93 ± 0.15 × 108 ab |
| Control (20) | 4.53 ± 0.59 × 108 ab | 4.93 ± 0.61 × 108 ab | 3.80 ± 0.25 × 108 b |
| Control (50) | 2.67 ± 0.35 × 108 b | 2.70 ± 0.36 × 108 b | 8.40 ± 0.30 × 107 bc |
| 10 mg/L | 1.23 ± 0.32 × 108 b | 1.70 ± 0.20 × 108 b | 6.87 ± 0.35 × 107 bc |
| 20 mg/L | 7.43 ± 0.45 × 107 bc | 7.43 ± 0.36 × 107 bc | 4.07 ± 0.47 × 107 bc |
| 50 mg/L | 1.73 ± 0.25 × 107 c | 1.80 ± 0.30 × 107 c | 1.10 ± 0.33 × 107 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimova, M.; Iutynska, G.; Yamborko, N.; Dordevic, D.; Kushkevych, I. Possible Processes and Mechanisms of Hexachlorobenzene Decomposition by the Selected Comamonas testosteroni Bacterial Strains. Processes 2022, 10, 2170. https://doi.org/10.3390/pr10112170
Dimova M, Iutynska G, Yamborko N, Dordevic D, Kushkevych I. Possible Processes and Mechanisms of Hexachlorobenzene Decomposition by the Selected Comamonas testosteroni Bacterial Strains. Processes. 2022; 10(11):2170. https://doi.org/10.3390/pr10112170
Chicago/Turabian StyleDimova, Mariia, Galyna Iutynska, Nadiya Yamborko, Dani Dordevic, and Ivan Kushkevych. 2022. "Possible Processes and Mechanisms of Hexachlorobenzene Decomposition by the Selected Comamonas testosteroni Bacterial Strains" Processes 10, no. 11: 2170. https://doi.org/10.3390/pr10112170
APA StyleDimova, M., Iutynska, G., Yamborko, N., Dordevic, D., & Kushkevych, I. (2022). Possible Processes and Mechanisms of Hexachlorobenzene Decomposition by the Selected Comamonas testosteroni Bacterial Strains. Processes, 10(11), 2170. https://doi.org/10.3390/pr10112170