
Safety and Protective Activities of Manufactured Alcohol-Free Beers




Abstract
:1. Introduction
2. Materials and Methods
2.1. Samples Preparation and Single Compounds
2.2. Model System Used
2.2.1. In Vivo: D. melanogaster
- mwh/mwh: holds the recessive mutation mwh (multiple wing hairs) that produces multiple tricomas per cell instead of one when homozygous [39].
- flr3/In (3LR) TM3, rippsep bx34eesBdS: the flr3(flare) marker is a homozygous recessive lethal mutation that causes deformed tricomas to appear, but once larvae begin development it is viable in homozygous somatic cells [40].
2.2.2. In Vitro: HL-60 Promyelocytic Human Leukaemia Cells
2.3. Safety In Vivo Assays
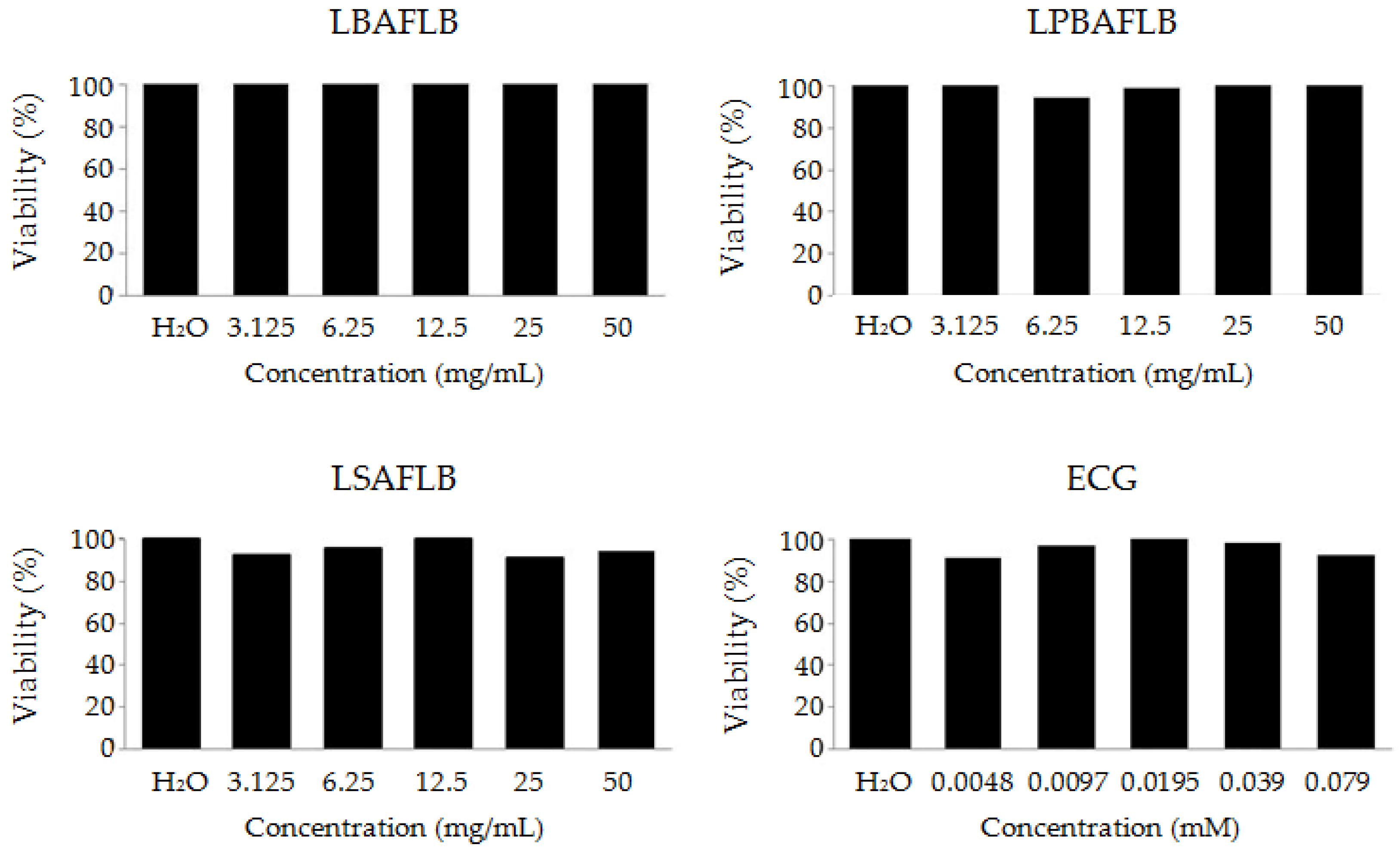
2.3.1. Toxicity
2.3.2. Genotoxicity
2.4. Protection In Vivo Tests
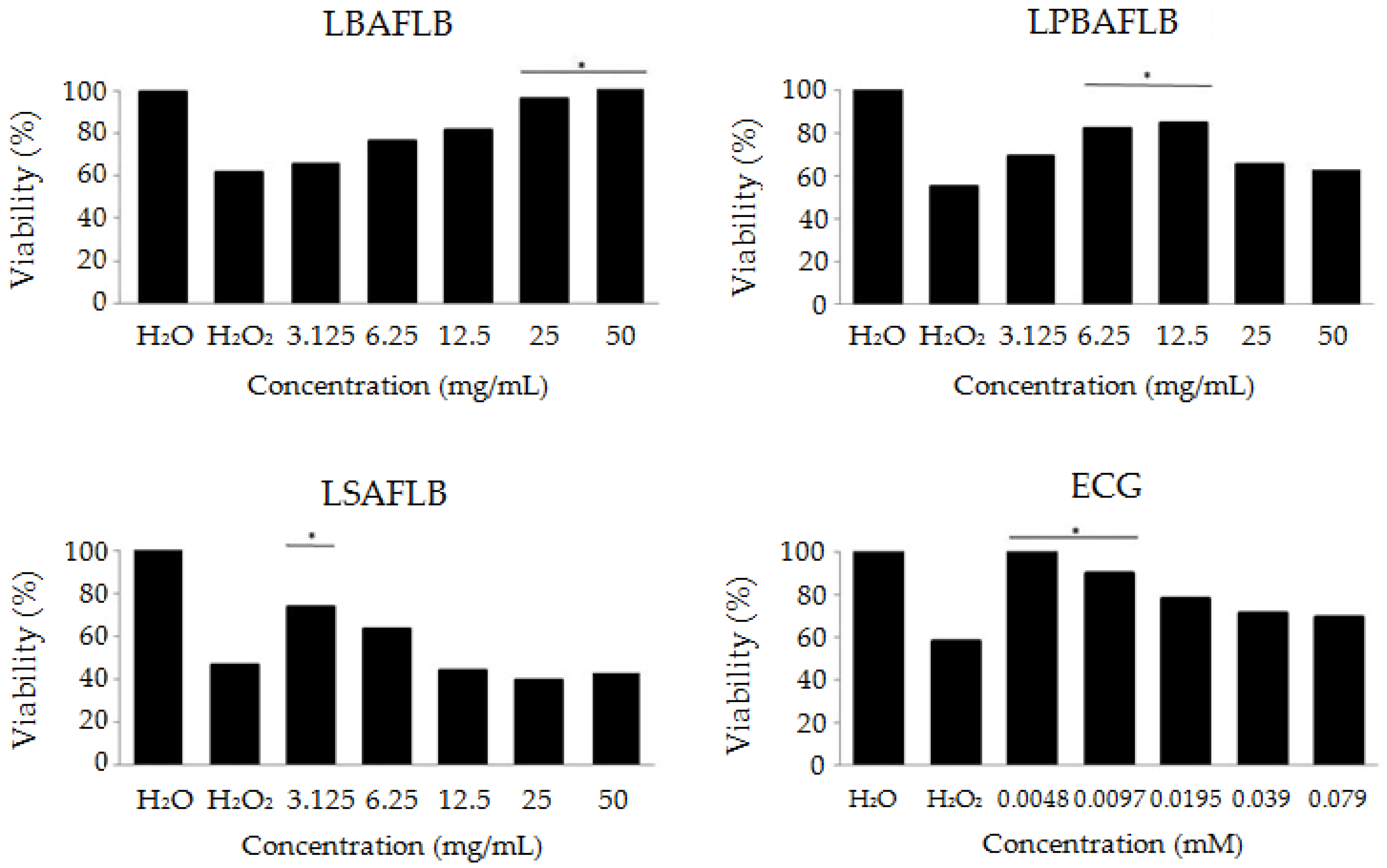
2.4.1. Antitoxicity
2.4.2. Antigenotoxicity
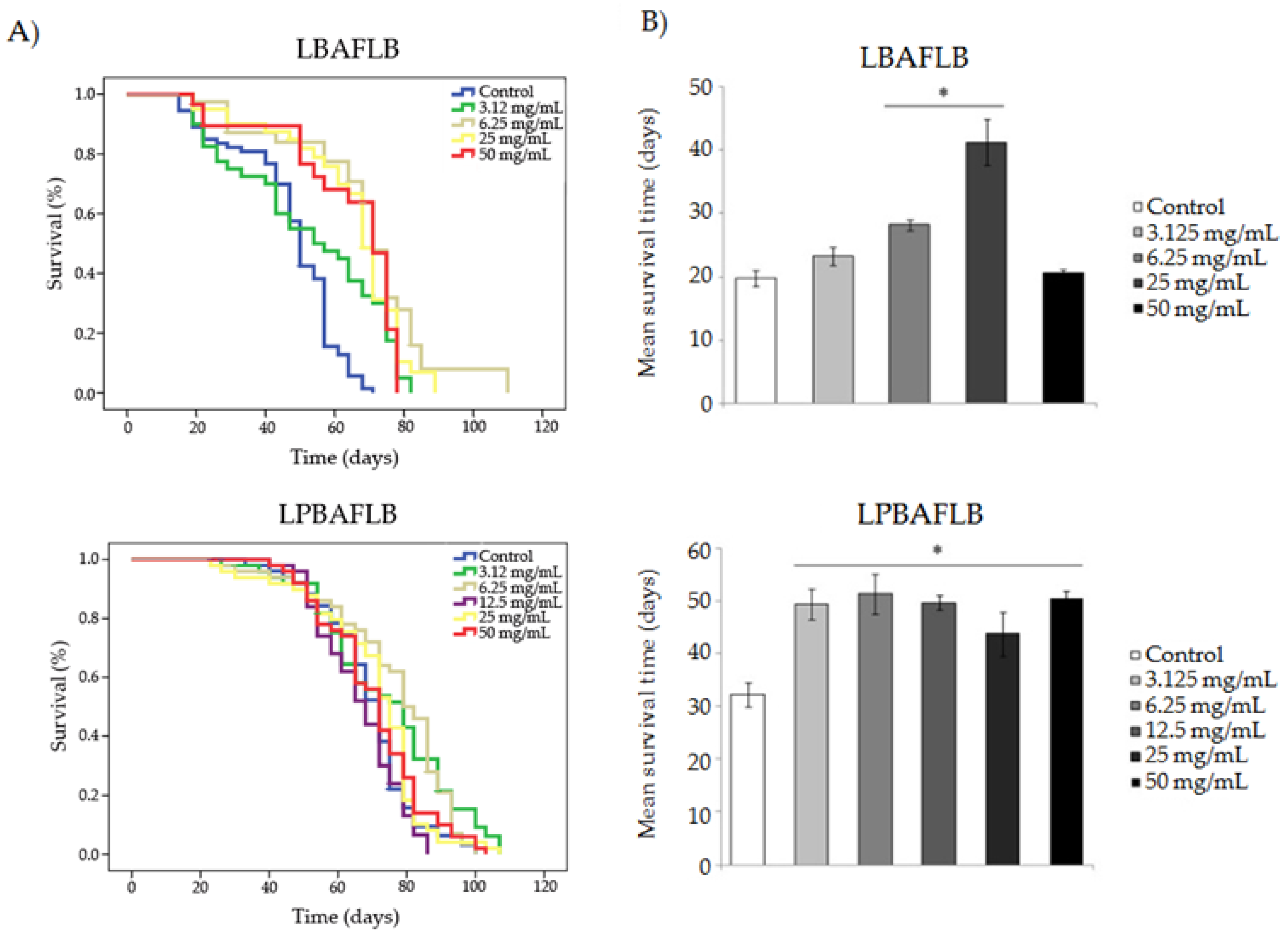
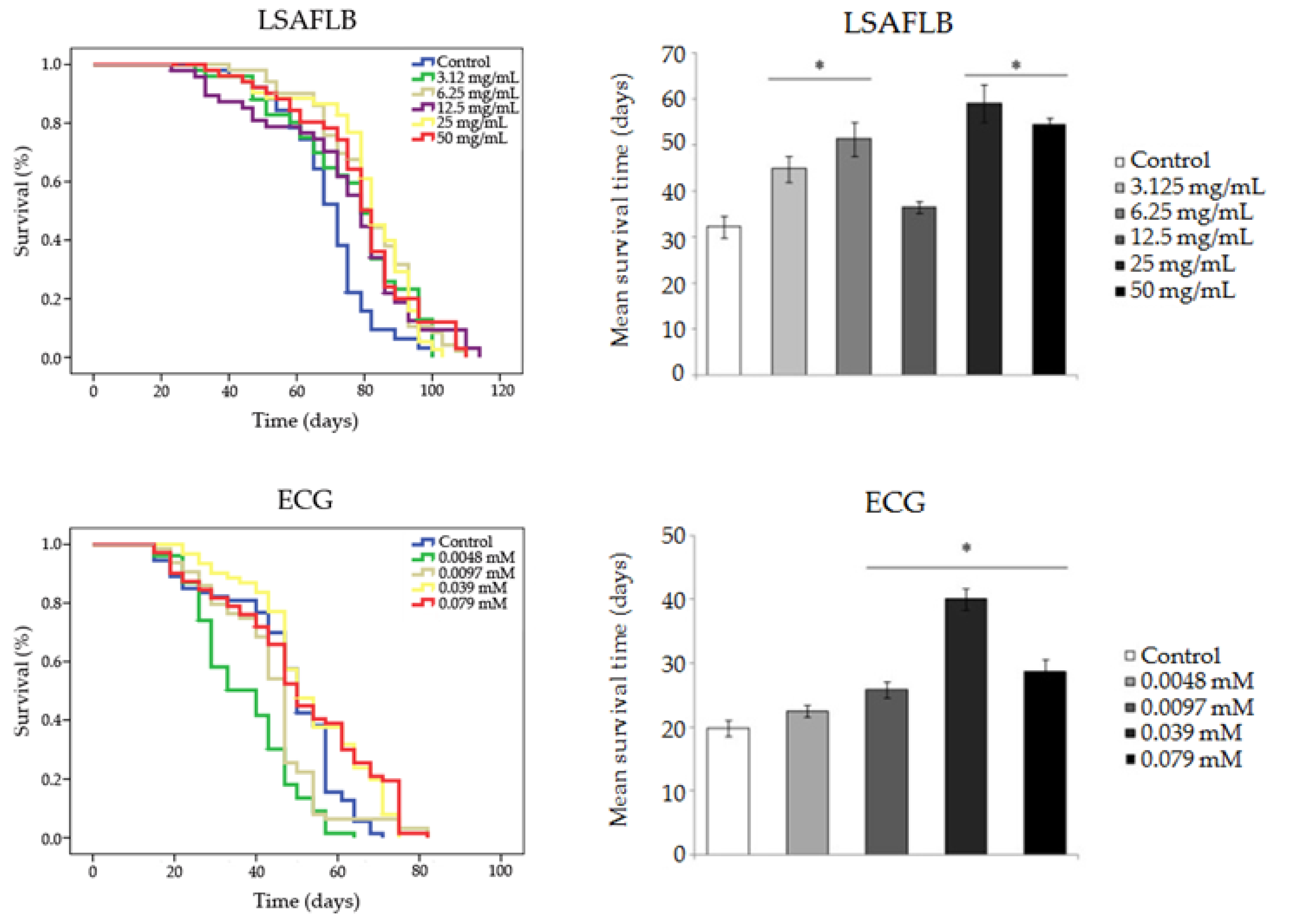
2.4.3. Longevity
2.5. Protection In Vitro Tests
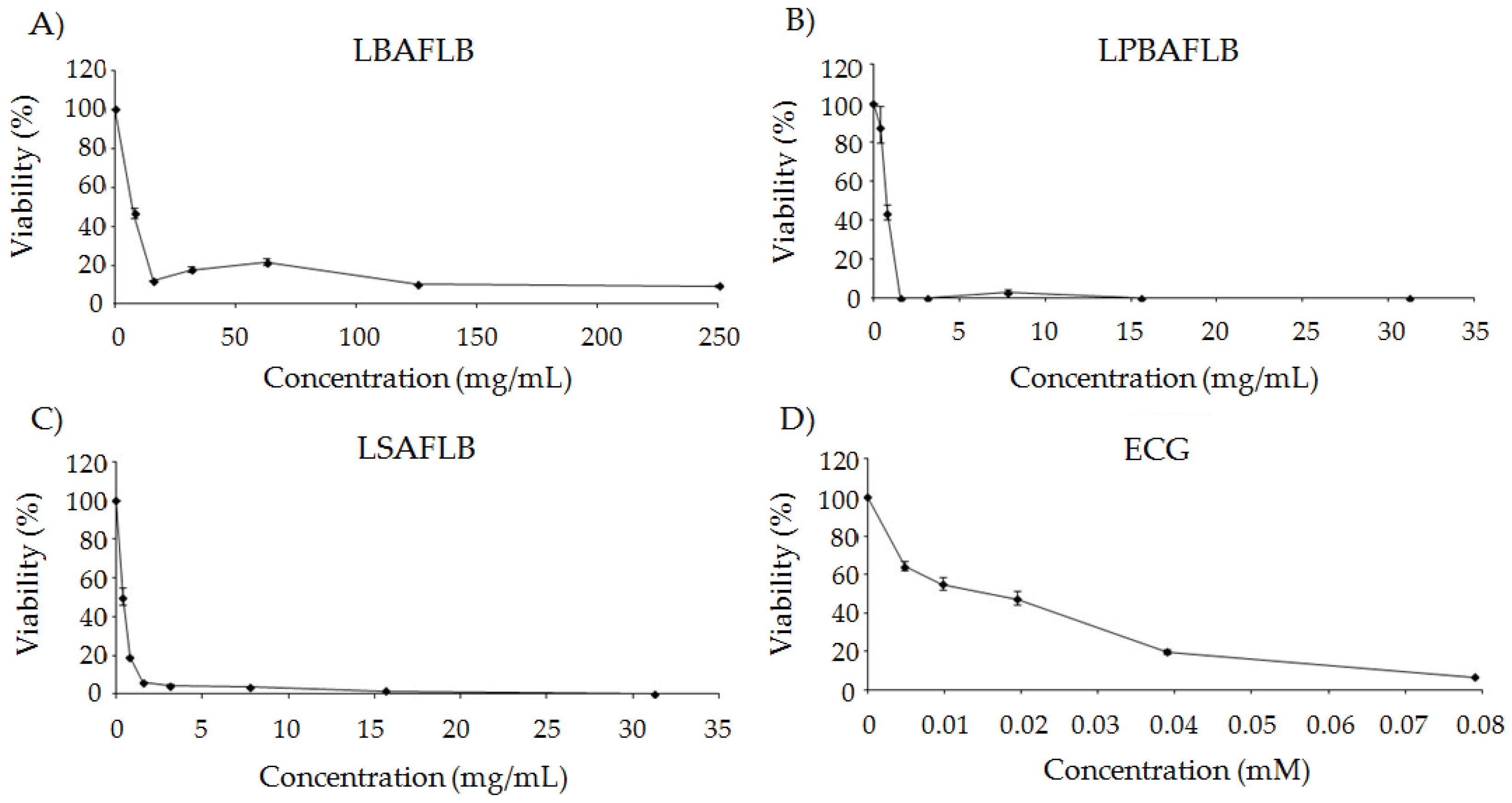
2.5.1. Cytotoxicity Assay
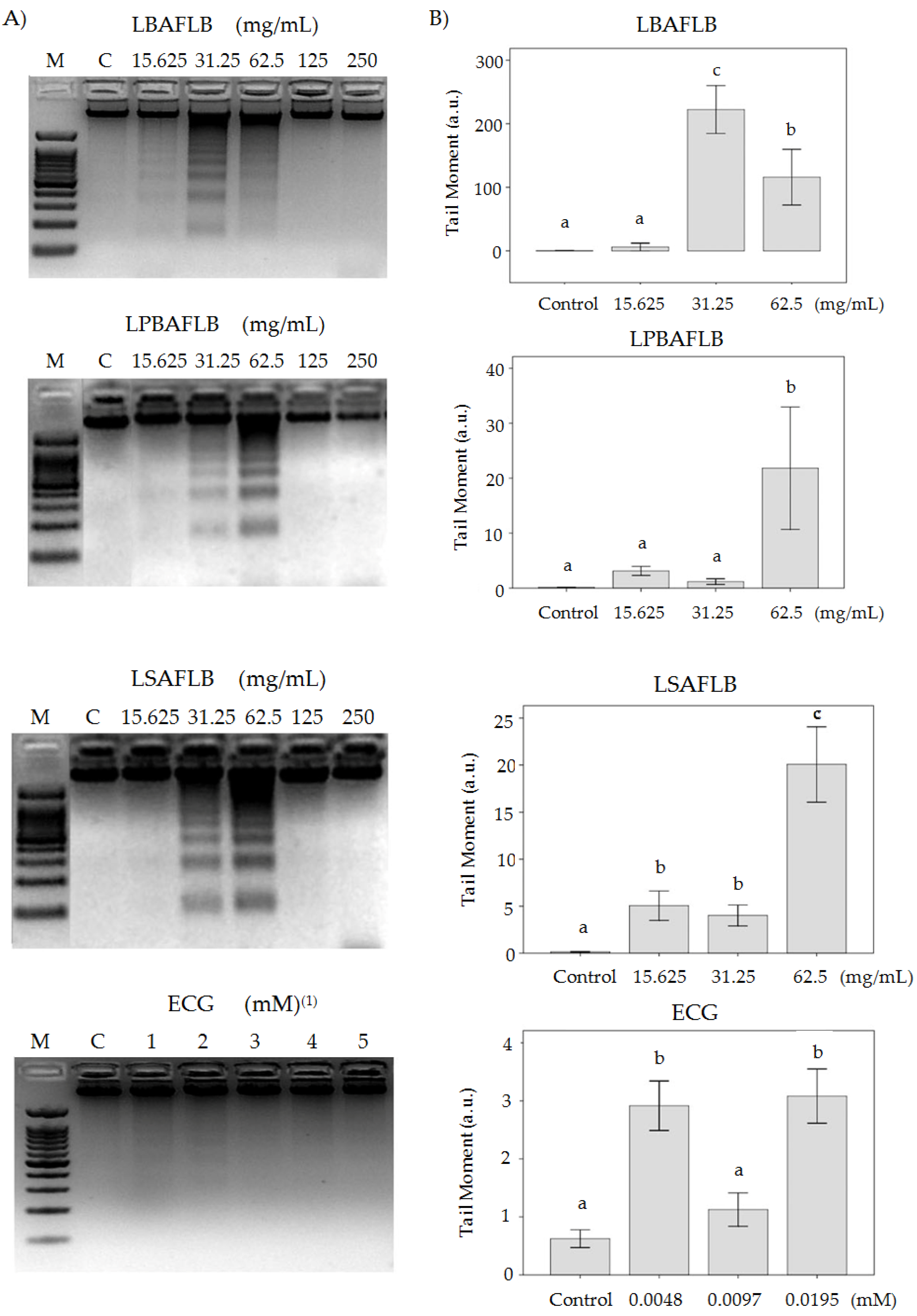
2.5.2. Tumour Cells’ DNA Damage Evaluation
DNA Fragmentation
Comet Assay
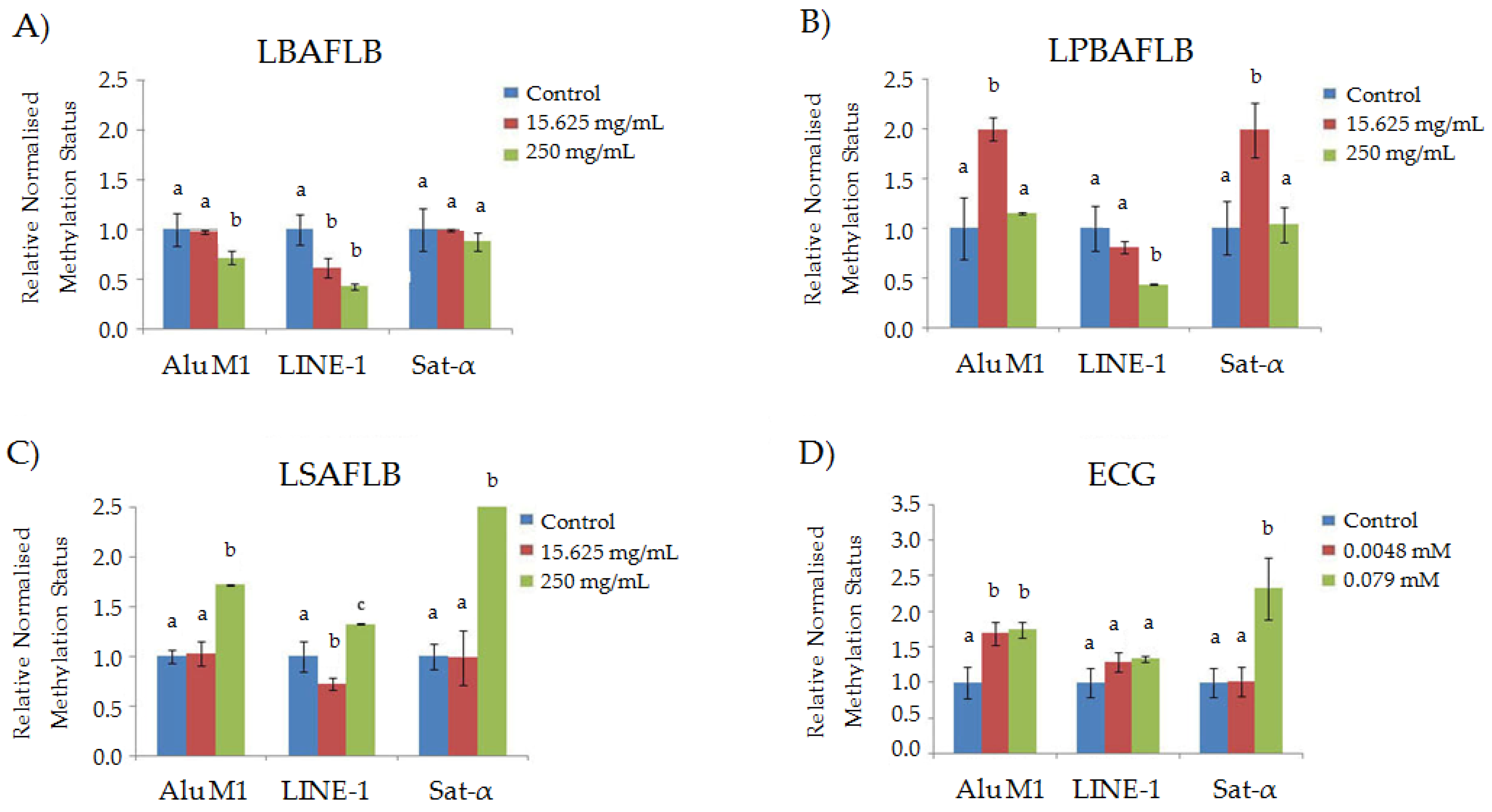
Methylation Status
3. Results and Discussion
3.1. Safety: Toxicity and Genotoxicity
3.2. Protection: Antitoxicity, Antigenotoxicity and Longevity
3.3. Chemoprotection: Cytotoxicity and DNA Damage
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Muller, R.; Walker, S.; Brauer, J.; Junquera, M. Does beer contain compounds that might interfere with cholesterol metabolism? J. Inst. Brew. 2007, 113, 102–109. [Google Scholar] [CrossRef]
- Bassus, S.; Mahnel, R.; Scholz, T.; Wegert, W.; Westrup, D.; Kirchmaier, C.M. Effect of dealcoholized beer (Bitburger Drive) consumption on hemostasis in humans. Alcohol. Clin. Exp. Res. 2004, 28, 786–791. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Becker, T.; Qian, F.; Ring, J. Beer and beer compounds: Physiological effects on skin health. J. Eur. Acad. Derm. Venereol. 2013, 28, 142–150. [Google Scholar] [CrossRef] [PubMed]
- Piazzon, A.; Forte, M.; Nardini, M. Characterization of phenolics content and antioxidant activity of different beer types. J. Agric. Food Chem. 2010, 58, 10677–10683. [Google Scholar] [CrossRef] [PubMed]
- Hornsey, I.S. A History of Beer and Brewing; Royal Society of Chemistry: London, UK, 2003; Volume 34. [Google Scholar]
- Gerhäuser, C. Beer constituents as potential cancer chemopreventive agents. Eur. J. Cancer 2005, 41, 1941–1954. [Google Scholar] [CrossRef]
- Merinas-Amo, T.; Tasset-Cuevas, I.; Díaz-Carretero, A.M.; Alonso-Moraga, Á.; Calahorro, F. In vivo and in vitro studies of the role of lyophilised blond Lager beer and some bioactive components in the modulation of degenerative processes. J. Funct. Foods 2016, 27, 274–294. [Google Scholar] [CrossRef]
- Merinas-Amo, T.; Merinas-Amo, R.; Font, R.; del Río Celestino, M.; Alonso-Moraga, Á. Toxicological and Epigenetic Studies of Two Types of Ale Beer, Tyrosol and Iso-Alpha Humulone. Processes 2021, 9, 485. [Google Scholar] [CrossRef]
- Stevens, J.F.; Miranda, C.L.; Wolthers, K.R.; Schimerlik, M.; Deinzer, M.L.; Buhler, D.R. Identification and in vitro biological activities of hop proanthocyanidins: Inhibition of nNOS activity and scavenging of reactive nitrogen species. J. Agric. Food Chem. 2002, 50, 3435–3443. [Google Scholar] [CrossRef]
- Gerhäuser, C.; Becker, H. Phenolic compounds in beer. In Beer in Health and Disease Prevention; Elsevier: London, UK, 2009; pp. 124–144. [Google Scholar]
- Cui, L.; Liu, Y.; Liu, T.; Yuan, Y.; Yue, T.; Cai, R.; Wang, Z. Extraction of Epigallocatechin Gallate and Epicatechin Gallate from Tea Leaves Using β-Cyclodextrin. J. Food Sci. 2017, 82, 394–400. [Google Scholar] [CrossRef]
- Zaveri, N.T. Green tea and its polyphenolic catechins: Medicinal uses in cancer and noncancer applications. Life Sci. 2006, 78, 2073–2080. [Google Scholar] [CrossRef]
- Bartolomé, B.; Pena-Neira, A.; Gómez-Cordovés, C. Phenolics and related substances in alcohol-free beers. Eur. Food Res. Technol. 2000, 210, 419–423. [Google Scholar] [CrossRef]
- McMurrough, I.; Baert, T. Identification of proanthocyanidins in beer and their direct measurement with a dual electrode electrochemical detector. J. Inst. Brew. 1994, 100, 409–416. [Google Scholar] [CrossRef]
- Granato, D.; Branco, G.F.; Faria, J.d.A.F.; Cruz, A.G. Characterization of Brazilian lager and brown ale beers based on color, phenolic compounds, and antioxidant activity using chemometrics. J. Sci. Food Agric. 2011, 91, 563–571. [Google Scholar] [CrossRef]
- De Keukeleire, D. Fundamentals of beer and hop chemistry. Quim. Nova 2000, 23, 108–112. [Google Scholar] [CrossRef]
- Kodama, Y.; Kielland-Brandt, M.C.; Hansen, J. Lager brewing yeast. In Comparative Genomics; Springer: Berlin/Heidelberg, Germany, 2006; pp. 145–164. [Google Scholar]
- Cantrell, I.; Griggs, D. Malt: Its role in oxidation. Tech. Q.-Master Brew. Assoc. Am. 1996, 33, 82–86. [Google Scholar]
- Woffenden, H.M.; Ames, J.M.; Chandra, S. Relationships between antioxidant activity, color, and flavor compounds of crystal malt extracts. J. Agric. Food Chem. 2001, 49, 5524–5530. [Google Scholar] [CrossRef] [PubMed]
- Coghe, S.; Vanderhaegen, B.; Pelgrims, B.; Basteyns, A.-V.; Delvaux, F.R. Characterization of dark specialty malts: New insights in color evaluation and pro-and antioxidative activity. J. Am. Soc. Brew. Chem. 2003, 61, 125–132. [Google Scholar] [CrossRef]
- Brányik, T.; Silva, D.P.; Baszczyňski, M.; Lehnert, R.; e Silva, J.B.A. A review of methods of low alcohol and alcohol-free beer production. J. Food Eng. 2012, 108, 493–506. [Google Scholar] [CrossRef]
- Etuk, B.; Murray, K. Mechanism for the removal of alcohol from beer by emulsion liquid membranes. Food Bioprod. Processing 1991, 69, 27–34. [Google Scholar]
- Matson, S.L. Production of Low-Ethanol Beverages by Membrane Extraction. Patent WO1987002380, 30 June 1988. [Google Scholar]
- Anglerot, D. Process of Making Alcohol-Free Beer and Beer Aroma Concentrates. U.S. Patent US5308631A, 3 May 1994. [Google Scholar]
- Von Hodenberg, G. Production of alcoholfree beers using reverse osmosis. Brauwelt Int. (Ger. F.R.) 1991, 2, 145–149. [Google Scholar]
- Richardson, H.E.; Willoughby, L.; Humbert, P.O. Screening for Anti-cancer Drugs in Drosophila. eLS 2015, 2, 97–103. [Google Scholar]
- Graf, U.; Wurgler, F.E.; Katz, A.J.; Frei, H.; Juon, H.; Hall, C.B.; Kale, P.G. Somatic mutation and recombination test in Drosophila melanogaster. Environ. Mutagenesis 1984, 6, 153–188. [Google Scholar] [CrossRef] [PubMed]
- De Gaetano, G.; Costanzo, S.; Di Castelnuovo, A.; Badimon, L.; Bejko, D.; Alkerwi, A.a.; Chiva-Blanch, G.; Estruch, R.; La Vecchia, C.; Panico, S. Effects of moderate beer consumption on health and disease: A consensus document. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 443–467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavsler, A.; Buiatti, S. Lager beer. In Beer in Health and Disease Prevention; Elsevier: Amsterdam, The Netherlands, 2009; pp. 31–43. [Google Scholar]
- Kirin. Globar Beer Consumption by Country in 2011. Available online: https://www.kirinholdings.com/en/newsroom/release/2019/1224_01.html (accessed on 24 December 2019).
- Ja, W.W.; Carvalho, G.B.; Mak, E.M.; de la Rosa, N.N.; Fang, A.Y.; Liong, J.C.; Brummel, T.; Benzer, S. Prandiology of Drosophila and the CAFE assay. Proc. Natl. Acad. Sci. USA 2007, 104, 8253–8256. [Google Scholar] [CrossRef] [Green Version]
- Bier, E. Drosophila, the golden bug, emerges as a tool for human genetics. Nat. Rev. Genet. 2005, 6, 9–23. [Google Scholar] [CrossRef]
- Lloyd, T.E.; Taylor, J.P. Flightless flies: Drosophila models of neuromuscular disease. Ann. N. Y. Acad. Sci. 2010, 1184, E1–E20. [Google Scholar] [CrossRef] [Green Version]
- Bhargav, D.; Singh, M.P.; Murthy, R.C.; Mathur, N.; Misra, D.; Saxena, D.K.; Chowdhuri, D.K. Toxic potential of municipal solid waste leachates in transgenic Drosophila melanogaster (hsp70-lacZ): Hsp70 as a marker of cellular damage. Ecotoxicol. Environ. Saf. 2008, 69, 233–245. [Google Scholar] [CrossRef]
- Coulom, H.; Birman, S. Chronic exposure to rotenone models sporadic Parkinson’s disease in Drosophila melanogaster. J. Neurosci. 2004, 24, 10993–10998. [Google Scholar] [CrossRef] [Green Version]
- Dean, B.J. Recent findings on the genetic toxicology of benzene, toluene, xylenes and phenols. Mutat. Res./Rev. Genet. Toxicol. 1985, 154, 153–181. [Google Scholar] [CrossRef]
- Hosamani, R. Acute exposure of Drosophila melanogaster to paraquat causes oxidative stress and mitochondrial dysfunction. Arch. Insect Biochem. Physiol. 2013, 83, 25–40. [Google Scholar] [CrossRef]
- Siddique, Y.H.; Fatima, A.; Jyoti, S.; Naz, F.; Khan, W.; Singh, B.R.; Naqvi, A.H. Evaluation of the toxic potential of graphene copper nanocomposite (GCNC) in the third instar larvae of transgenic Drosophila melanogaster (hsp70-lacZ) Bg 9. PLoS ONE 2013, 8, e80944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, J.; Huen, D.; Morely, T.; Johnson, G.; Gubb, D.; Roote, J.; Adler, P.N. The multiple-wing-hairs gene encodes a novel GBD-FH3 domain-containing protein that functions both prior to and after wing hair initiation. Genetics 2008, 180, 219–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, N.; Charlton, J.; Adler, P.N. The flare gene, which encodes the AIP1 protein of Drosophila, functions to regulate F-actin disassembly in pupal epidermal cells. Genetics 2007, 176, 2223–2234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, S.J. The HL-60 promyelocytic leukemia cell line: Proliferation, differentiation, and cellular oncogene expression. Blood 1987, 70, 1233–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gallagher, R.; Collins, S.; Trujillo, J.; McCredie, K.; Ahearn, M.; Tsai, S.; Metzgar, R.; Aulakh, G.; Ting, R.; Ruscetti, F.; et al. Characterization of the continuous, differentiating myeloid cell line (HL-60) from a patient with acute promyelocytic leukemia. Blood 1979, 54, 713–733. [Google Scholar] [CrossRef] [Green Version]
- Frei, H.; Wurgler, F.E. Statistical methods to decide whether mutagenicity test data from Drosophila assays indicate a positive, negative, or inconclusive result. Mutat. Res. 1988, 203, 297–308. [Google Scholar] [CrossRef]
- Frei, H.; Wurgler, F.E. Optimal experimental design and sample size for the statistical evaluation of data from somatic mutation and recombination tests (SMART) in Drosophila. Mutat. Res. 1995, 334, 247–258. [Google Scholar] [CrossRef]
- Valadares, B.; Graf, U.; Spanó, M. Inhibitory effects of water extract of propolis on doxorubicin-induced somatic mutation and recombination in Drosophila melanogaster. Food Chem. Toxicol. 2008, 46, 1103–1110. [Google Scholar] [CrossRef]
- Tasset-Cuevas, I.; Fernandez-Bedmar, Z.; Lozano-Baena, M.D.; Campos-Sanchez, J.; de Haro-Bailon, A.; Munoz-Serrano, A.; Alonso-Moraga, A. Protective effect of borage seed oil and gamma linolenic acid on DNA: In vivo and in vitro studies. PLoS ONE 2013, 8, e56986. [Google Scholar] [CrossRef] [Green Version]
- Graf, U.; Abraham, S.K.; Guzman-Rincon, J.; Wurgler, F.E. Antigenotoxicity studies in Drosophila melanogaster. Mutat. Res. 1998, 402, 203–209. [Google Scholar] [CrossRef]
- Abraham, S.K. Antigenotoxicity of coffee in the Drosophila assay for somatic mutation and recombination. Mutagenesis 1994, 9, 383–386. [Google Scholar] [CrossRef] [PubMed]
- Soh, J.W.; Hotic, S.; Arking, R. Dietary restriction in Drosophila is dependent on mitochondrial efficiency and constrained by pre-existing extended longevity. Mech. Ageing Dev. 2007, 128, 581–593. [Google Scholar] [CrossRef] [PubMed]
- Mateo-Fernández, M.; Merinas-Amo, T.; Moreno-Millán, M.; Alonso-Moraga, Á.; Demyda-Peyrás, S. In vivo and in vitro genotoxic and epigenetic effects of two types of cola beverages and caffeine: A multi-assay approach. BioMed Res. Int. 2016, 2016, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deininger, P.L.; Moran, J.V.; Batzer, M.A.; Kazazian, H.H., Jr. Mobile elements and mammalian genome evolution. Curr. Opin. Genet. Dev. 2003, 13, 651–658. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.; Wevrick, R.; Fisher, R.B.; Ferguson-Smith, M.A.; Lin, C.C. Human centromeric DNAs. Hum. Genet. 1997, 100, 291–304. [Google Scholar] [CrossRef] [PubMed]
- Ehrlich, M. DNA methylation in cancer: Too much, but also too little. Oncogene 2002, 21, 5400–5413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiner, A.M. SINEs and LINEs: The art of biting the hand that feeds you. Curr. Opin. Cell Biol. 2002, 14, 343–350. [Google Scholar] [CrossRef]
- Weisenberger, D.J.; Campan, M.; Long, T.I.; Kim, M.; Woods, C.; Fiala, E.; Ehrlich, M.; Laird, P.W. Analysis of repetitive element DNA methylation by MethyLight. Nucleic Acids Res. 2005, 33, 6823–6836. [Google Scholar] [CrossRef] [Green Version]
- Nikolaidis, G.; Raji, O.Y.; Markopoulou, S.; Gosney, J.R.; Bryan, J.; Warburton, C.; Walshaw, M.; Sheard, J.; Field, J.K.; Liloglou, T. DNA methylation biomarkers offer improved diagnostic efficiency in lung cancer. Cancer Res. 2012, 72, 5692–5701. [Google Scholar] [CrossRef] [Green Version]
- Liloglou, T.; Bediaga, N.G.; Brown, B.R.; Field, J.K.; Davies, M.P. Epigenetic biomarkers in lung cancer. Cancer Lett. 2014, 342, 200–212. [Google Scholar] [CrossRef]
- Anter, J.; Romero-Jimenez, M.; Fernandez-Bedmar, Z.; Villatoro-Pulido, M.; Analla, M.; Alonso-Moraga, A.; Munoz-Serrano, A. Antigenotoxicity, cytotoxicity, and apoptosis induction by apigenin, bisabolol, and protocatechuic acid. J. Med. Food 2011, 14, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Romero-Jimenez, M.; Campos-Sanchez, J.; Analla, M.; Munoz-Serrano, A.; Alonso-Moraga, A. Genotoxicity and anti-genotoxicity of some traditional medicinal herbs. Mutat. Res. 2005, 585, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Psomiadou, E.; Tsimidou, M. Stability of virgin olive oil. 1. Autoxidation studies. J. Agric. Food Chem. 2002, 50, 716–721. [Google Scholar] [CrossRef] [PubMed]
- Anter, J.; Campos-Sanchez, J.; Hamss, R.E.; Rojas-Molina, M.; Munoz-Serrano, A.; Analla, M.; Alonso-Moraga, A. Modulation of genotoxicity by extra-virgin olive oil and some of its distinctive components assessed by use of the Drosophila wing-spot test. Mutat. Res. 2010, 703, 137–142. [Google Scholar] [CrossRef]
- Preedy, V.R. Beer in Health and Disease Prevention; Academic Press: London, UK, 2011. [Google Scholar]
- Arimoto-Kobayashi, S.; Ishida, R.; Nakai, Y.; Idei, C.; Takata, J.; Takahashi, E.; Okamoto, K.; Negishi, T.; Konuma, T. Inhibitory effects of beer on mutation in the Ames test and DNA adduct formation in mouse organs induced by 2-Amino-1-methyl-6-phenylimidazo [4, 5-b] pyridine (PhIP). Biol. Pharm. Bull. 2006, 29, 67–70. [Google Scholar] [CrossRef] [Green Version]
- Vanderhaegen, B.; Neven, H.; Verstrepen, K.J.; Delvaux, F.R.; Verachtert, H.; Derdelinckx, G. Influence of the brewing process on furfuryl ethyl ether formation during beer aging. J. Agric. Food Chem. 2004, 52, 6755–6764. [Google Scholar] [CrossRef]
- Coghe, S.; Martens, E.; D’Hollander, H.; Dirinck, P.J.; Delvaux, F.R. Sensory and instrumental flavour analysis of wort brewed with dark specialty malts. J. Inst. Brew. 2004, 110, 94–103. [Google Scholar] [CrossRef]
- Liégeois, C.; Lermusieau, G.; Collin, S. Measuring antioxidant efficiency of wort, malt, and hops against the 2,2′-azobis (2-amidinopropane) dihydrochloride-induced oxidation of an aqueous dispersion of linoleic acid. J. Agric. Food Chem. 2000, 48, 1129–1134. [Google Scholar] [CrossRef]
- Tafulo, P.A.R.; Queirós, R.B.; Delerue-Matos, C.M.; Sales, M.G.F. Control and comparison of the antioxidant capacity of beers. Food Res. Int. 2010, 43, 1702–1709. [Google Scholar] [CrossRef] [Green Version]
- Rizvi, S.I.; Zaid, M.A.; Anis, R.; Mishra, N. Protective role of tea catechins against oxidation-induced damage of type 2 diabetic erythrocytes. Clin. Exp. Pharmacol. Physiol. 2005, 32, 70–75. [Google Scholar] [CrossRef]
- Siddique, Y.H.; Jyoti, S.; Naz, F. Effect of epicatechin gallate dietary supplementation on transgenic Drosophila model of Parkinson’s disease. J. Diet. Suppl. 2014, 11, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Cuvelier, M.-E.; Richard, H.; Berset, C. Comparison of the antioxidative activity of some acid-phenols: Structure-activity relationship. Biosci. Biotechnol. Biochem. 1992, 56, 324–325. [Google Scholar] [CrossRef] [Green Version]
- Srinivasan, M.; Sudheer, A.R.; Menon, V.P. Ferulic acid: Therapeutic potential through its antioxidant property. J. Clin. Biochem. Nutr. 2007, 40, 92–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-Moreno, C.; Larrauri, J.A.; Saura-Calixto, F. A procedure to measure the antiradical efficiency of polyphenols. J. Sci. Food Agric. 1998, 76, 270–276. [Google Scholar] [CrossRef]
- Żołnierczyk, A.K.; Mączka, W.K.; Grabarczyk, M.; Wińska, K.; Woźniak, E.; Anioł, M. Isoxanthohumol—Biologically active hop flavonoid. Fitoterapia 2015, 103, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.-Y.; Juan, C.-C.; Ho, L.-T.; Hsu, Y.-P.; Hwang, L.S. Effect of green tea supplementation on insulin sensitivity in Sprague-Dawley rats. J. Agric. Food Chem. 2004, 52, 643–648. [Google Scholar] [CrossRef]
- Miura, Y.; Chiba, T.; Miura, S.; Tomita, I.; Umegaki, K.; Ikeda, M.; Tomita, T. Green tea polyphenols (flavan 3-ols) prevent oxidative modification of low density lipoproteins: An ex vivo study in humans. J. Nutr. Biochem. 2000, 11, 216–222. [Google Scholar] [CrossRef]
- Kao, Y.-h.; Hiipakka, R.A.; Liao, S. Modulation of obesity by a green tea catechin. Am. J. Clin. Nutr. 2000, 72, 1232–1233. [Google Scholar] [CrossRef] [Green Version]
- Tian, W.-X. Inhibition of fatty acid synthase by polyphenols. Curr. Med. Chem. 2006, 13, 967–977. [Google Scholar] [CrossRef]
- Zhang, R.; Xiao, W.; Wang, X.; Wu, X.; Tian, W. Novel inhibitors of fatty-acid synthase from green tea (Camellia sinensis Xihu Longjing) with high activity and a new reacting site. Biotechnol. Appl. Biochem. 2006, 43, 1–7. [Google Scholar]
- Gorinstein, S.; Zemser, M.; Berliner, M.; Goldstein, R.; Libman, I.; Trakhtenberg, S.; Caspi, A. Moderate beer consumption and positive biochemical changes in patients with coronary atherosclerosis. J. Intern. Med. 1997, 242, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Gorinstein, S.; Zemser, M.; Lichman, I.; Berebi, A.; Kleipfish, A.; Libman, I.; Trakhtenberg, S.; Caspi, A. Moderate beer consumption and the blood coagulation in patients with coronary artery disease. J. Intern. Med. 1997, 241, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Nie, G.; Jin, C.; Cao, Y.; Shen, S.; Zhao, B. Distinct effects of tea catechins on 6-hydroxydopamine-induced apoptosis in PC12 cells. Arch. Biochem. Biophys. 2002, 397, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Mandel, S.; Maor, G.; Youdim, M.B. Iron and α-synuclein in the substantia nigra of MPTP-treated mice. J. Mol. Neurosci. 2004, 24, 401–416. [Google Scholar] [CrossRef]
- Riboli, E.; Cornée, J.; Macquart-Moulin, G.; Kaaks, R.; Casagrande, C.; Guyader, M. Cancer and polyps of the colorectum and lifetime consumption of beer and other alcoholic beverages. Am. J. Epidemiol. 1991, 134, 157–166. [Google Scholar] [CrossRef]
- Kato, I.; Tominaga, S.; Ikari, A. A Case-Control Study of Male Colorectal Cancer in Aichi Prefecture, Japan: With Special Reference to Occupational Activity Level, Drinking Habits and Family History. Jpn. J. Cancer Res. 1990, 81, 115–121. [Google Scholar] [CrossRef]
- Potter, J.D.; Sellers, T.A.; Folsom, A.R.; McGovern, P.G. Alcohol, beer, and lung cancer in postmenopausal women the iowa women’s health study. Ann. Epidemiol. 1992, 2, 587–595. [Google Scholar] [CrossRef]
- Swanson, C.A.; Wilbanks, G.D.; Twiggs, L.B.; Monet, R.; Berman, M.L.; Barrett, R.J.; Brintonl, L.A. Moderate alcohol consumption and the risk of endometrial cancer. Epidemiology 1993, 4, 530–536. [Google Scholar] [CrossRef]
- Hevia, D.; Mayo, J.C.; Quiros, I.; Sainz, R.M. Beer constituents inhibit prostate cancer cells proliferation. EJC Suppl. 2008, 6, 142. [Google Scholar] [CrossRef]
- Hong, J.; Smith, T.J.; Ho, C.-T.; August, D.A.; Yang, C.S. Effects of purified green and black tea polyphenols on cyclooxygenase-and lipoxygenase-dependent metabolism of arachidonic acid in human colon mucosa and colon tumor tissues. Biochem. Pharmacol. 2001, 62, 1175–1183. [Google Scholar] [CrossRef]
- Weyant, M.J.; Carothers, A.M.; Dannenberg, A.J.; Bertagnolli, M.M. (+)-Catechin inhibits intestinal tumor formation and suppresses focal adhesion kinase activation in the min/+ mouse. Cancer Res. 2001, 61, 118–125. [Google Scholar] [PubMed]
- Baek, S.J.; Wilson, L.C.; Eling, T.E. Resveratrol enhances the expression of non-steroidal anti-inflammatory drug-activated gene (NAG-1) by increasing the expression of p53. Carcinogenesis 2002, 23, 425–432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, S.; Umekita, Y.; Guo, J.; Kokontis, J.M.; Hiipakka, R.A. Growth inhibition and regression of human prostate and breast tumors in athymic mice by tea epigallocatechin gallate. Cancer Lett. 1995, 96, 239–243. [Google Scholar] [CrossRef]
- Bigelow, R.; Cardelli, J. The green tea catechins,(−)-epigallocatechin-3-gallate (EGCG) and (−)-epicatechin-3-gallate (ECG), inhibit HGF/Met signaling in immortalized and tumorigenic breast epithelial cells. Oncogene 2006, 25, 1922–1930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, N.; Feyes, D.K.; Agarwal, R.; Mukhtar, H.; Nieminen, A.-L. Green tea constituent epigallocatechin-3-gallate and induction of apoptosis and cell cycle arrest in human carcinoma cells. J. Natl. Cancer Inst. 1997, 89, 1881–1886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, L.; Cheung, T.; Kong, S.; Fung, K.; Choy, Y.; Chan, Z.; Kwok, T. Induction of apoptosis by green tea catechins in human prostate cancer DU145 cells. Life Sci. 2001, 68, 1207–1214. [Google Scholar] [CrossRef]
- Lim, Y.C.; Lee, S.-H.; Song, M.H.; Yamaguchi, K.; Yoon, J.-H.; Choi, E.C.; Baek, S.J. Growth inhibition and apoptosis by (−)-epicatechin gallate are mediated by cyclin D1 suppression in head and neck squamous carcinoma cells. Eur. J. Cancer 2006, 42, 3260–3266. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.-Y.; Liao, J.; Kim, K.; Yurkow, E.J.; Yang, C.S. Inhibition of growth and induction of apoptosis in human cancer cell lines by tea polyphenols. Carcinogenesis 1998, 19, 611–616. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.-C.; Fang, J.-Y.; Wu, W.-B.; Chiang, H.-S.; Wei, Y.-J.; Hung, C.-F. Protective effects of (−)-epicatechin-3-gallate on UVA-induced damage in HaCaT keratinocytes. Arch. Dermatol. Res. 2005, 296, 473–481. [Google Scholar] [CrossRef]
- Poe, B.S.; O’Neill, K.L. Caffeine modulates heat shock induced apoptosis in the human promyelocytic leukemia cell line HL-60. Cancer Lett. 1997, 121, 1–6. [Google Scholar] [CrossRef]
- Fabiani, R.; Rosignoli, P.; De Bartolomeo, A.; Fuccelli, R.; Morozzi, G. Genotoxicity of alkene epoxides in human peripheral blood mononuclear cells and HL60 leukaemia cells evaluated with the comet assay. Mutat. Res./Genet. Toxicol. Environ. Mutagenesis 2012, 747, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Babich, H.; Krupka, M.; Nissim, H.A.; Zuckerbraun, H.L. Differential in vitro cytotoxicity of (−)-epicatechin gallate (ECG) to cancer and normal cells from the human oral cavity. Toxicol. In Vitro 2005, 19, 231–242. [Google Scholar] [CrossRef] [PubMed]
- Fairbairn, D.W.; Walburger, D.K.; Fairbairn, J.J.; O’Neill, K.L. Key morphologic changes and DNA strand breaks in human lymphoid cells: Discriminating apoptosis from necrosis. Scanning 1996, 18, 407–416. [Google Scholar] [CrossRef] [PubMed]
- Fairbairn, D.W.; Olive, P.L.; O’Neill, K.L. The comet assay: A comprehensive review. Mutat. Res. 1995, 339, 37–59. [Google Scholar] [CrossRef]
- Herman, J.G.; Baylin, S.B. Gene silencing in cancer in association with promoter hypermethylation. N. Engl. J. Med. 2003, 349, 2042–2054. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. DNA methylation and cancer therapy: New developments and expectations. Curr. Opin. Oncol. 2005, 17, 55–60. [Google Scholar] [CrossRef]
- Roman-Gomez, J.; Jimenez-Velasco, A.; Agirre, X.; Castillejo, J.A.; Navarro, G.; San Jose-Eneriz, E.; Garate, L.; Cordeu, L.; Cervantes, F.; Prosper, F. Repetitive DNA hypomethylation in the advanced phase of chronic myeloid leukemia. Leuk. Res. 2008, 32, 487–490. [Google Scholar] [CrossRef]
- Fang, M.; Chen, D.; Yang, C.S. Dietary polyphenols may affect DNA methylation. J. Nutr. 2007, 137, 223S–228S. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reaction ID | GenBank Number | Amplicon Start | Amplicon End | Forward Primer Sequence 5′ to 3′ (N) | Reverse Primer Sequence 5′ to 3′ (N) | GC-Content (%) | |
|---|---|---|---|---|---|---|---|
| Forward | Reverse | ||||||
| ALU-C4 | Consensus Sequence | 1 | 98 | GGTTAGGTATAGTGGTTTATATTTGTAATTTTAGTA (36) | ATTAACTAAACTAATCTTAAACTCCTAACCTCA (33) | 25 | 27.3 |
| ALU M1 | Y07755 | 5059 | 5164 | ATTATGTTAGTTAGGATGGTTTCGATTTT (29) | CAATCGACCGAACGCGA (17) | 27.6 | 58.8 |
| LINE-1 | X52235 | 251 | 331 | GGACGTATTTGGAAAATCGGG (21) | AATCTCGCGATACGCCGTT (19) | 47.6 | 52.6 |
| Sat-α | M38468 | 139 | 260 | TGATGGAGTATTTTTAAAATATACGTTTTGTAGT (34) | AATTCTAAAAATATTCCTCTTCAATTACGTAAA (33) | 23.5 | 21.2 |
| Clones Per Wing (No. of Spots) (1) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Compound | No of Wings | Small Single Clones (1–2 Cells) m = 2 | Large Single Clones (>2 Cells) m = 5 | Twin Clones m = 5 | Total Clones m = 2 | F (2) | U-Test (3) | R (%) (4) |
| Single Treatment (mwh/flr3) | ||||||||
| H2O | 38 | 0.157 (6) | 0 | 0 | 0.157 (6) | 0.650 | 76.92 | |
| LBAFLB (mg/mL) | ||||||||
| 3.125 | 47 | 0.255 (12) | 0.0212 (1) | 0 | 0.276 (13) i | 1.134 | ∆ | |
| 50 | 45 | 0.288 (13) | 0 | 0 | 0.288 (13) i | 1.184 | ∆ | |
| LPBAFLB (mg/mL) | ||||||||
| 3.125 | 40 | 0.450 (18) | 0.15 (5) | 0 | 0.575 (23) + | 2.360 | 78.39 | |
| 50 | 40 | 0.225 (9) | 0.075 (3) | 0 | 0.300 (12) i | 1.230 | ∆ | |
| LSAFLB (mg/mL) | ||||||||
| 3.125 | 42 | 0.357 (15) | 0.095 (4) | 0 | 0.452 (19) + | 1.850 | 83.24 | |
| 50 | 32 | 0.281 (9) | 0.094 (3) | 0 | 0.375 (12) i | 1.665 | ∆ | |
| ECG (mM) | ||||||||
| 0.0048 | 38 | 0.210 (8) | 0.052 (2) | 0 | 0.263 (10) i | 1.079 | ∆ | |
| 0.079 | 40 | 0.175 (7) | 0.025 (1) | 0 | 0.200 (8) i | 0.820 | ∆ | |
| Single Treatment (mwh/TM3) | ||||||||
| H2O (5) | 80 | 0.040 (3) | 0 | 0 | 0.040 (3) | 0.150 | ||
| LPBAFLB (mg/mL) | ||||||||
| 3.125 | 40 | 0.125 (5) | 0 | 0 | 0.125 (5) | 0.510 | ||
| LSAFLB (mg/mL) | ||||||||
| 3.125 | 40 | 0.075 (3) | 0 | 0 | 0.075 (3) | 0.310 | ||
| Clones Per Wing (No. of Spots) (1) | |||||||
|---|---|---|---|---|---|---|---|
| Compound | No. of Wings | Small Single Clones (1–2 Cells) m = 2 | Large Single Clones (>2 Cells) m = 5 | Twin Clones m = 5 | Total Clones m = 2 | F (2) | IP (%) (3) |
| H2O | 38 | 0.157 (6) | 0 | 0 | 0.157 (6) | 0.650 | |
| H2O2 | 36 | 0.305 (11) | 0.083 (3) | 0 | 0.388 (14) + | 2.049 | |
| LBAFLB (mg/mL) | |||||||
| 3.125 | 39 | 0.231 (9) | 0.0512 (2) | 0 | 0.282 (11) β | 1.156 | 27.32 |
| 50 | 45 | 0.155 (7) | 0.066 (3) | 0 | 0.222 (10) β | 0.911 | 42.78 |
| LPBAFLB (mg/mL) | |||||||
| 3.125 | 44 | 0.295 (13) | 0.068 (3) | 0 | 0.363 (16) β | 1.490 | 6.44 |
| 50 | 40 | 0.025 (10) | 0.025 (1) | 0 | 0.275 (11) β | 1.127 | 29.12 |
| LSAFLB (mg/mL) | |||||||
| 3.125 | 34 | 0.235 (8) | 0.059 (2) | 0 | 0.290 (10) β | 1.205 | 25.25 |
| 50 | 40 | 0.150 (6) | 0.100 (4) | 0 | 0.250 (10) β | 1.025 | 35.56 |
| ECG (mM) | |||||||
| 0.0048 | 40 | 0.275 (11) | 0.050 (2) | 0 | 0.325 (13) β | 1.332 | 16.24 |
| 0.079 | 40 | 0.250 (10) | 0.050 (2) | 0.050 (2) | 0.350 (14) β | 1.434 | 9.79 |
| Compound | Concentration | Mean Lifespan (1) (Days) | Mean Healthspan (1) (Days) | ||
|---|---|---|---|---|---|
| LBAFLB | Control | 47.242 | 19.769 | ||
| 3.125 mg/mL | 52.900 | *** | 23.273 | ns | |
| 6.25 mg/mL | 66.380 | *** | 28.167 | *** | |
| 25 mg/mL | 64.994 | *** | 41.231 | *** | |
| 50 mg/mL | 53.488 | *** | 20.538 | ns | |
| LPBAFLB | Control | 69.324 | 32.222 | ||
| 3.125 mg/mL | 74.358 | * | 49.413 | *** | |
| 6.25 mg/mL | 76.310 | ** | 51.308 | *** | |
| 12.5 mg/mL | 69.535 | ns | 49.667 | *** | |
| 25 mg/mL | 70.490 | ns | 43.700 | * | |
| 50 mg/mL | 70.660 | ns | 50.417 | *** | |
| LSAFLB | Control | 69.324 | 32.222 | ||
| 3.125 mg/mL | 75.782 | ** | 44.786 | *** | |
| 6.25 mg/mL | 80.852 | *** | 58.675 | *** | |
| 12.5 mg/mL | 74.179 | ** | 36.500 | ns | |
| 25 mg/mL | 80.686 | *** | 59.077 | *** | |
| 50 mg/mL | 78.966 | *** | 54.577 | *** | |
| ECG | Control | 47.242 | 19.769 | ||
| 0.0048 mM | 37.187 | *** | 22.500 | ns | |
| 0.0097 mM | 43.899 | * | 25.800 | *** | |
| 0.039 mM | 53.070 | * | 39.962 | *** | |
| 0.079 mM | 50.647 | * | 28.619 | *** | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Merinas-Amo, T.; Del Río Celestino, M.; Font, R.; Alonso-Moraga, Á. Safety and Protective Activities of Manufactured Alcohol-Free Beers. Processes 2022, 10, 331. https://doi.org/10.3390/pr10020331
Merinas-Amo T, Del Río Celestino M, Font R, Alonso-Moraga Á. Safety and Protective Activities of Manufactured Alcohol-Free Beers. Processes. 2022; 10(2):331. https://doi.org/10.3390/pr10020331
Chicago/Turabian StyleMerinas-Amo, Tania, Mercedes Del Río Celestino, Rafael Font, and Ángeles Alonso-Moraga. 2022. "Safety and Protective Activities of Manufactured Alcohol-Free Beers" Processes 10, no. 2: 331. https://doi.org/10.3390/pr10020331
APA StyleMerinas-Amo, T., Del Río Celestino, M., Font, R., & Alonso-Moraga, Á. (2022). Safety and Protective Activities of Manufactured Alcohol-Free Beers. Processes, 10(2), 331. https://doi.org/10.3390/pr10020331