
Reproducibility of Aerobic Granules in Treating Low-Strength and Low-C/N-Ratio Wastewater and Associated Microbial Community Structure


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup and Operation
2.2. Medium
2.3. Analytical Method
2.4. Microbiological Analysis of Granular Sludge
3. Results and Discussion
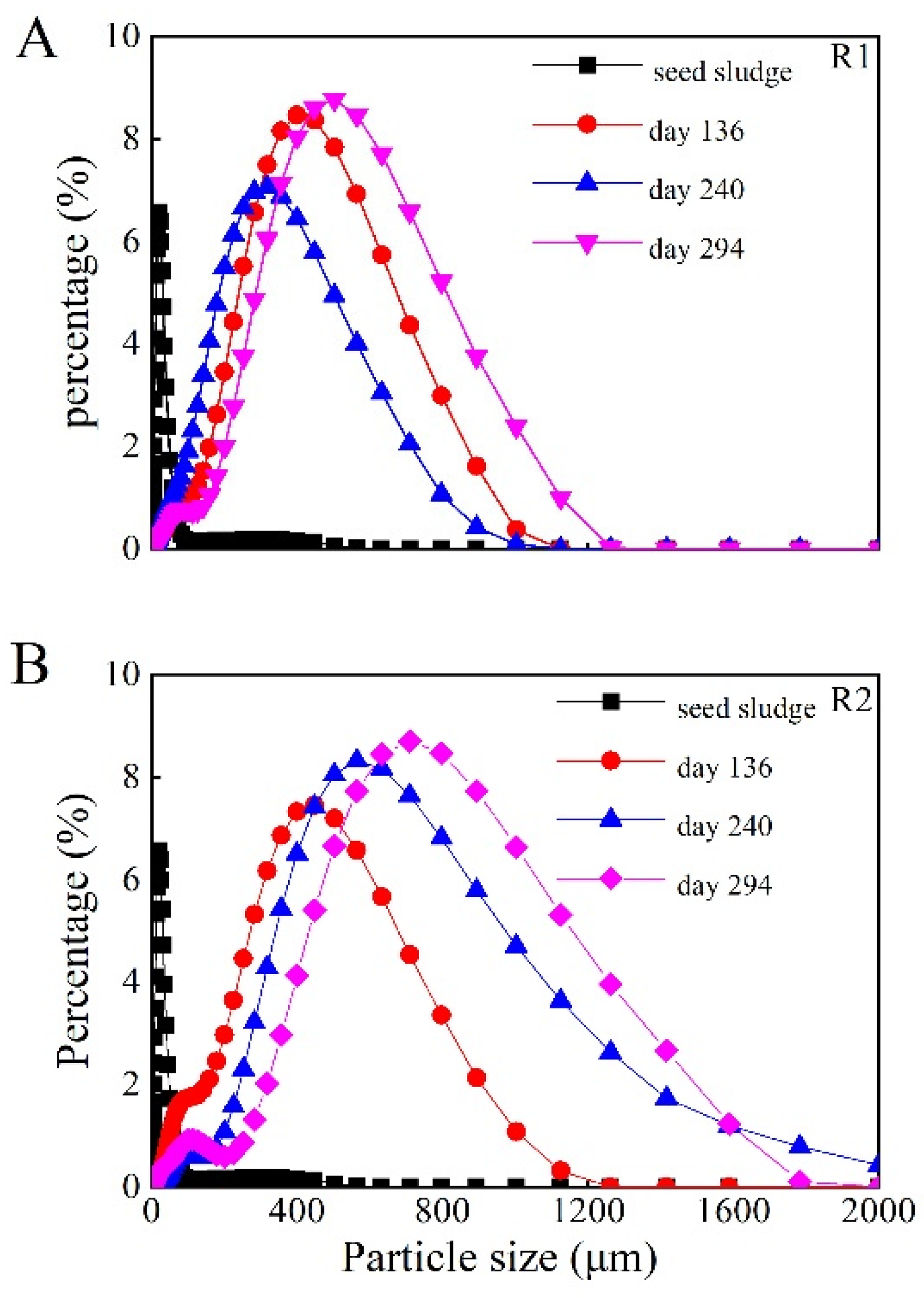
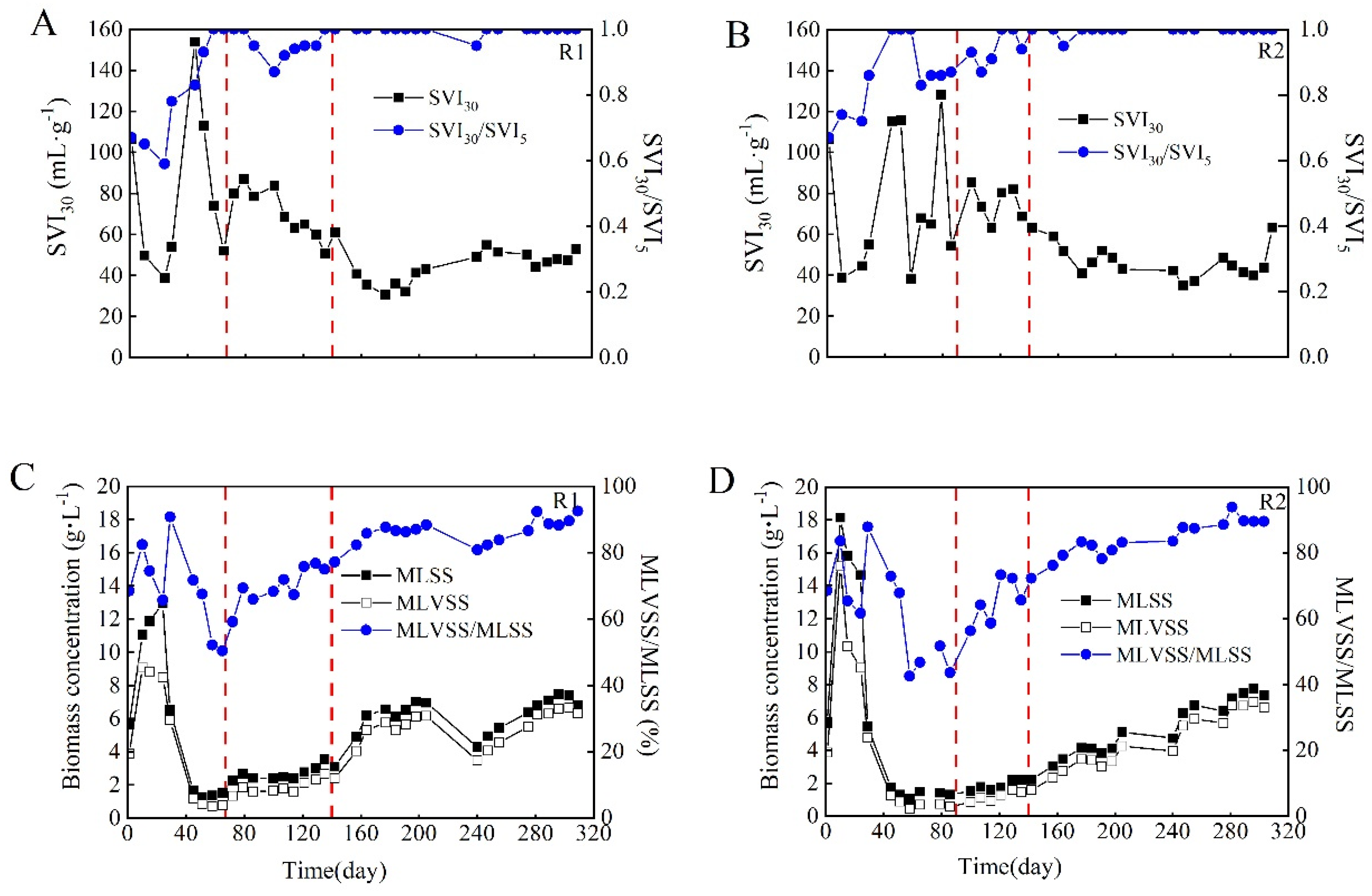
3.1. Characteristics of Aerobic Granular Sludge during Long-Term Operation
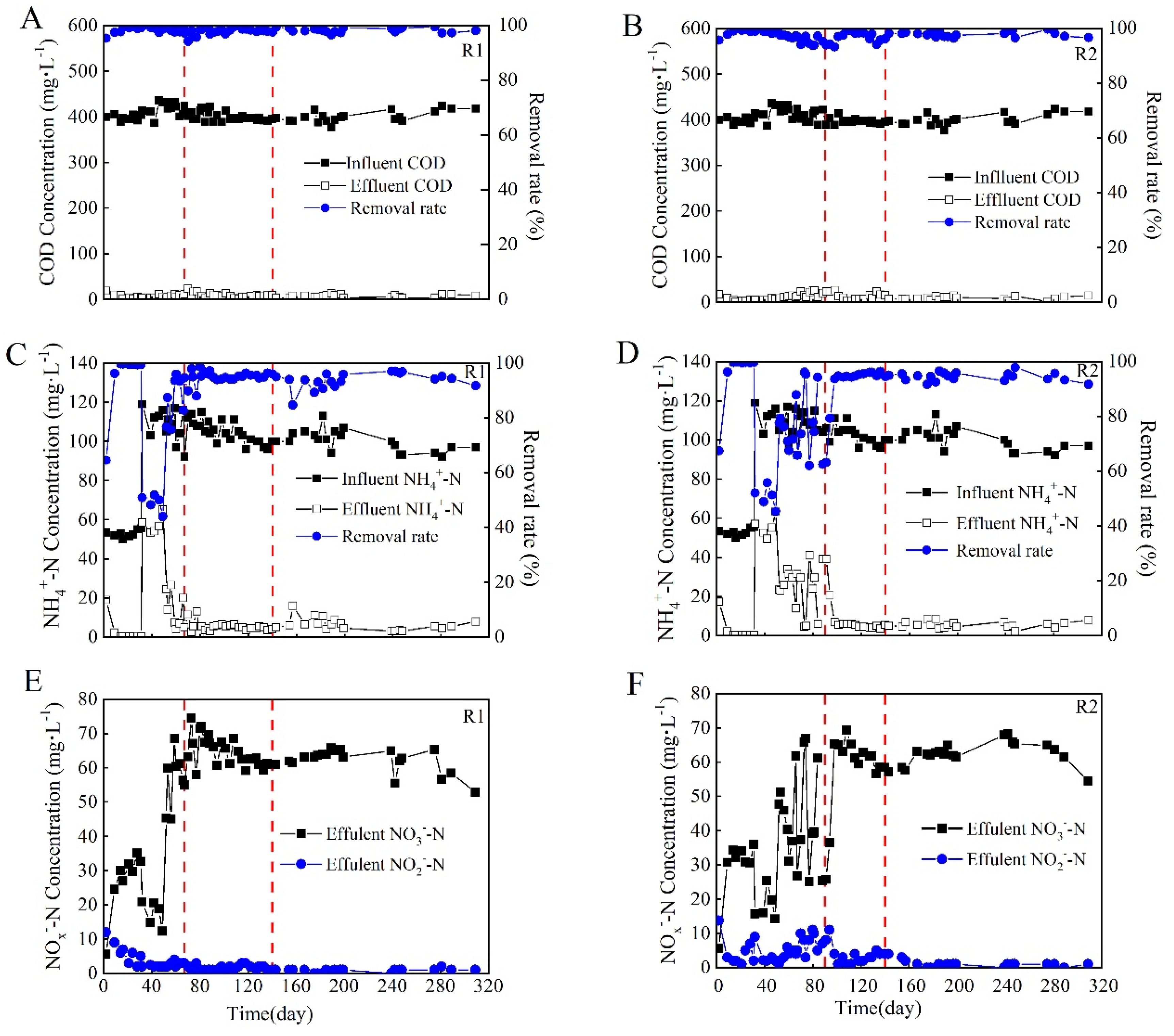
3.2. Performance of the Aerobic Granular Sludge during the Long-Term Operation Period
3.3. The Richness and Diversity of Microbial Community in the Aerobic Granular Sludge
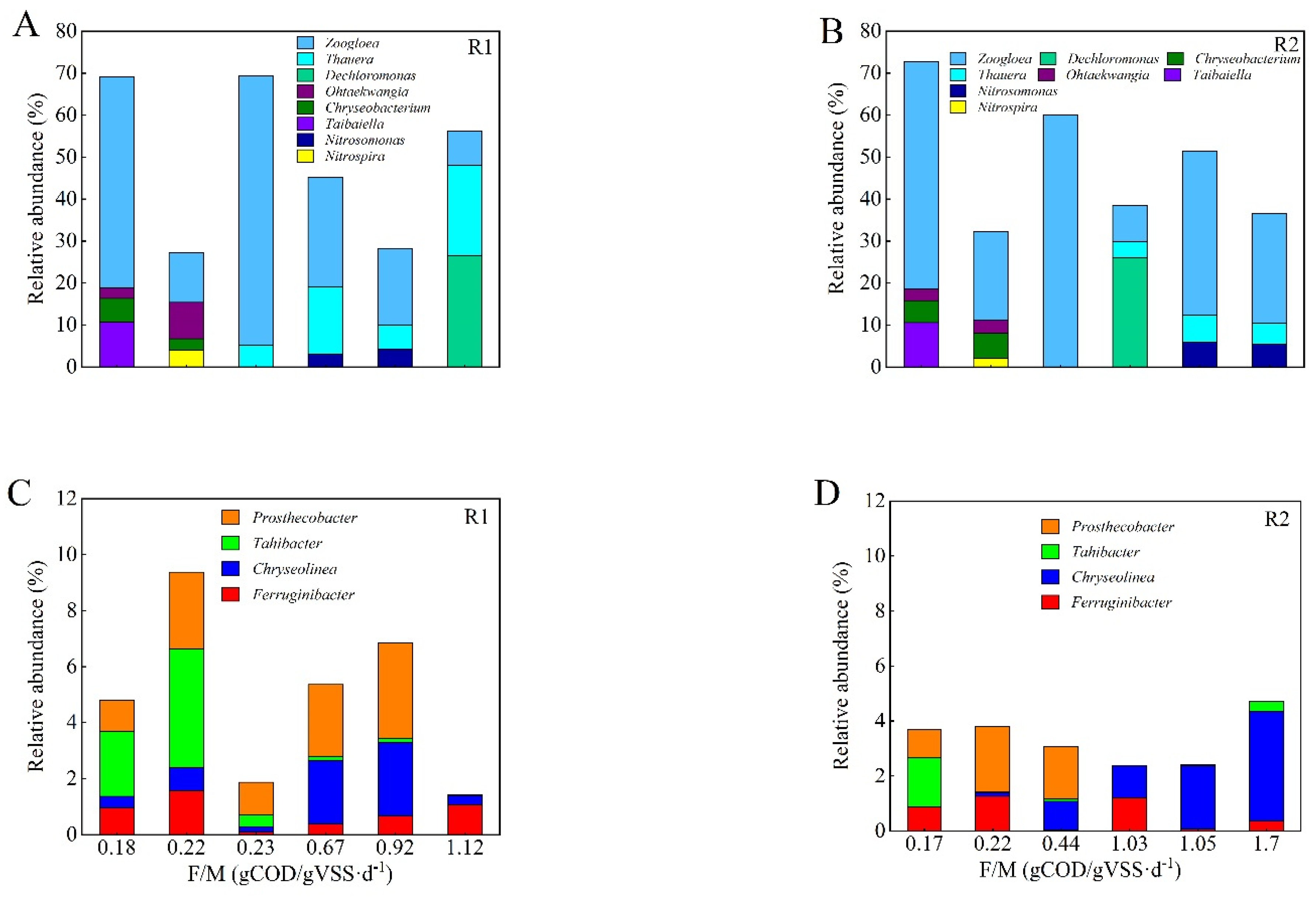
3.4. Microbial Population Dynamics and the Predominant Functional Groups
3.5. Food to Microbial Biomass Ratio (F/M) and its Relationship with the Predominant Genus in the Aerobic Granules
4. Conclusions
- The SVI30 before and during granulation was different in the two reactors, although it reached similar values after granule maturation. In addition, the mean size and size distribution of the sludge were quite different in the two reactors, although both reactors were operated under exactly the same conditions. These differences indicate some degree of randomness in granule formation and size development even under identical conditions. However, the similarity of the physicochemical and microbial properties of the granules, as well as the performance of the wastewater treatment after granule maturation, indicate that the operating conditions can produce consistent results, implying predictability of stable operation in practice.
- A high F/M value promotes the formation of aerobic granules, while a low F/M value in the range of 0.2–0.4 g COD/g VSS·d facilitates the long-term stability of aerobic granules.
- The richness of the microbial population of the granules was much lower than that of the inoculum and the flocs with bulking, although the richness may increase slightly during the long-term operation period. The diversity of the microbial structure decreased over time. However, the reduction in the richness and diversity of the microbial population due to the conversion of flocs to granules did not affect the wastewater treatment performance and long-term stability of the sludge, indicating the robustness of different microbial structures to achieve the same function in wastewater treatment.
- Among the dominant genera of the sludge, Zoogloea played a key role in maintaining the stable structure of aerobic granules throughout the operation period, while Thauera is an important genus for the formation and maturation of aerobic granules, but not for long-term maintenance. In addition, Ohtaekwangia, Chryseobacterium, Taibaiella, and Tahibacter can play an important role in the long-term stability of aerobic granules.
Author Contributions
Funding
Informed Consent Statement
Conflicts of Interest
References
- Wang, L.; Liu, X.; Lee, D.-J.; Tay, J.-H.; Zhang, Y.; Wan, C.-L.; Chen, X.-F. Recent advances on biosorption by aerobic granular sludge. J. Hazard. Mater. 2018, 357, 253–270. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.-J.; Chen, Y.-Y.; Show, K.-Y.; Whiteley, C.G.; Tay, J.-H. Advances in aerobic granule formation and granule stability in the course of storage and reactor operation. Biotechnol. Adv. 2010, 28, 919–934. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, Q.S. Causes and control of filamentous growth in aerobic granular sludge sequencing batch reactors. Biotechnol. Adv. 2006, 24, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Bengtsson, S.; de Blois, M.; Wilen, B.-M.; Gustavsson, D. Treatment of municipal wastewater with aerobic granular sludge. Crit. Rev. Environ. Sci. Technol. 2018, 48, 119–166. [Google Scholar] [CrossRef]
- Adav, S.S.; Lee, D.-J.; Show, K.-Y.; Tay, J.-H. Aerobic granular sludge: Recent advances. Biotechnol. Adv. 2008, 26, 411–423. [Google Scholar] [CrossRef]
- Cydzik-Kwiatkowska, A.; Wojnowska-Barya, I. Nitrogen-converting communities in aerobic granules at different hydraulic retention times (HRTs) and operational modes. World J. Microbiol. Biotechnol. 2015, 31, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Swiatczak, P.; Cydzik-Kwiatkowska, A. Performance and microbial characteristics of biomass in a full-scale aerobic granular sludge wastewater treatment plant. Environ. Sci. Pollut. Res. 2018, 25, 1655–1669. [Google Scholar] [CrossRef] [Green Version]
- Cha, L.; Liu, Y.-Q.; Duan, W.; Sternberg, C.E.W.; Yuan, Q.; Chen, F. Fluctuation and Re-Establishment of Aerobic Granules Properties during the Long-Term Operation Period with Low-Strength and Low C/N Ratio Wastewater. Process 2021, 9, 1290. [Google Scholar] [CrossRef]
- Pronk, M.; de Kreuk, M.K.; de Bruin, B.; Kamminga, P.; Kleerebezem, R.; van Loosdrecht, M.C.M. Full scale performance of the aerobic granular sludge process for sewage treatment. Water Res. 2015, 84, 207–217. [Google Scholar] [CrossRef]
- Yang, S.F.; Tay, J.H.; Liu, Y. Effect of substrate nitrogen/chemical oxygen demand ratio on the formation of aerobic granules. J. Environ. Eng. 2005, 131, 86–92. [Google Scholar] [CrossRef]
- de Kreuk, M.K.; Kishida, N.; Tsuneda, S.; van Loosdrecht, M.C.M. Behavior of polymeric substrates in an aerobic granular sludge system. Water Res. 2010, 44, 5929–5938. [Google Scholar] [CrossRef] [PubMed]
- Layer, M.; Adler, A.; Reynaert, E.; Hernandez, A.; Pagni, M.; Morgenroth, E.; Holliger, C.; Derlon, N. Organic substrate diffusibility governs microbial community composition, nutrient removal performance and kinetics of granulation of aerobic granular sludge. Water Res. X 2019, 4, 100033. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.Q.; Liu, Y.; Tay, J.H. Relationship between size and mass transfer resistance in aerobic granules. Lett. Appl. Microbiol. 2005, 40, 312–315. [Google Scholar] [CrossRef] [PubMed]
- APHA, Standard Methods for the Examination of Water and Wastewater, 23rd ed.; American Public Health Association, American Water Works Association, and Water Environmental Federation: Washington, DC, USA, 2017.
- Hill, T.C.J.; Walsh, K.A.; Harris, J.A.; Moffett, B.F. Using ecological diversity measures with bacterial communities. FEMS Microbiol. Ecol. 2003, 43, 1–11. [Google Scholar]
- Westphal, C.; Bommarco, R.; Carré, G.; Lamborn, E.; Morison, N.; Petanidou, T.; Potts, S.G.; Roberts, S.P.M.; Szentgyörgyi, H.; Tscheulin, T.; et al. Measuring bee diversity in different European habitats and biogeographical regions. Ecol. Monogr. 2008, 78, 653–671. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.-Q.; Tay, J.-H. The competition between flocculent sludge and aerobic granules during the long-term operation period of granular sludge sequencing batch reactor. Environ. Technol. 2012, 33, 2619–2626. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Kong, Y.H.; Zhang, R.; Zhang, X.; Wong, F.S.; Tay, J.H.; Zhu, J.R.; Jiang, W.J.; Liu, W.T. Microbial population dynamics of granular aerobic sequencing batch reactors during start-up and steady state periods. Water Sci. Technol. 2010, 62, 1281–1287. [Google Scholar] [CrossRef]
- Wang, L.; Zhan, H.; Wang, Q.; Wu, G.; Cui, D. Enhanced aerobic granulation by inoculating dewatered activated sludge under short settling time in a sequencing batch reactor. Bioresour. Technol. 2019, 286, 121386. [Google Scholar] [CrossRef]
- He, Q.; Chen, L.; Zhang, S.; Chen, R.; Wang, H. Hydrodynamic shear force shaped the microbial community and function in the aerobic granular sequencing batch reactors for low carbon to nitrogen (C/N) municipal wastewater treatment. Bioresour. Technol. 2019, 271, 48–58. [Google Scholar] [CrossRef]
- Yoon, J.-H.; Kang, S.-J.; Oh, T.-K. Chryseobacterium daeguense sp nov., isolated from wastewater of a textile dye works. Int. J. Syst. Evol. Microbiol. 2007, 57, 1355–1359. [Google Scholar] [CrossRef] [Green Version]
- Makk, J.; Homonnay, Z.G.; Keki, Z.; Lejtovicz, Z.; Marialigeti, K.; Sproeer, C.; Schumann, P.; Toth, E.M. Tahibacter aquaticus gen. nov., sp nov., a new gammaproteobacterium isolated from the drinking water supply system of Budapest (Hungary). Syst. Appl. Microbiol. 2011, 34, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Chhetri, G.; Kim, I.; Kim, J.; Kang, M.; Seo, T. Taibaiella lutea sp. nov., Isolated from Ubiquitous Weedy Grass. Curr. Microbiol. 2021, 78, 2799–2805. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Gil, G.; Holliger, C. Dynamics of microbial community structure of and enhanced biological phosphorus removal by aerobic granules cultivated on propionate or acetate. Appl. Environ. Microbiol. 2011, 77, 8041–8051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehsani, E.; Jauregui, R.; Geffers, R.; Jarek, M.; Boon, N.; Pieper, D.H.; Vilchez-Vargas, R. First Draft Genome Sequence of the Acidovorax caeni sp. nov. Type Strain R-24608 (DSM 19327). Genome Announc. 2015, 3, e01378-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, S.; Tan, C.H.; Constancias, F.; Kohli, G.S.; Cohen, Y.; Rice, S.A. Predation by Bdellovibrio bacteriovorus significantly reduces viability and alters the microbial community composition of activated sludge flocs and granules. FEMS Microbiol. Ecol. 2017, 93, fix020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, X.; Zhou, Z.; Mei, X.; Ma, Y.; Xie, Z. Influence of fermentation liquid from waste activated sludge on anoxic/oxic-membrane bioreactor performance: Nitrogen removal, membrane fouling and microbial community. Bioresour. Technol. 2018, 250, 699–707. [Google Scholar] [CrossRef]
- Ummalyma, S.B.; Gnansounou, E.; Sukumaran, R.K.; Sindhu, R.; Pandey, A.; Sahoo, D. Bioflocculation: An alternative strategy for harvesting of microalgae—An overview. Bioresour. Technol. 2017, 242, 227–235. [Google Scholar] [CrossRef]
- Yu, H.; Meng, W.; Song, Y.; Tian, Z. Understanding bacterial communities of partial nitritation and nitratation reactors at ambient and low temperature. Chem. Eng. J. 2017, 337, 755–763. [Google Scholar] [CrossRef]
- Staley, J.T.; Bouzek, H.; Jenkins, C. Eukaryotic signature proteins of Prosthecobacter dejongeii and Gemmata sp Wa-1 as revealed by in silico analysis. FEMS Microbiol. Lett. 2005, 243, 9–14. [Google Scholar] [CrossRef] [Green Version]
- Baek, K.; Choi, A. Complete Genome Sequence of Phreatobacter sp. Strain NMCR1094, a Formate-Utilizing Bacterium Isolated from a Freshwater Stream. Microbiol. Resour. Announc. 2019, 8, e00860-19. [Google Scholar] [CrossRef] [Green Version]
- Raittz, R.T.; de Pierri, C.R.; Maluk, M.; Batista, M.B.; Carmona, M.; Junghare, M.; Faoro, H.; Cruz, L.M.; Battistoni, F.; Souza, E.D.; et al. Article comparative genomics provides insights into the taxonomy of Azoarcus and reveals separate origins of nif genes in the proposed Azoarcus and Aromatoleum genera. Genes 2021, 12, 71. [Google Scholar] [CrossRef] [PubMed]
- Xi, L.; Liu, D.; Huang, W. Effect of acetate and propionate on the production and characterization of soluble microbial products (SMP) in aerobic granular sludge system. Toxicol. Environ. Chem. 2018, 100, 175–190. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Operation Day | ||||||
|---|---|---|---|---|---|---|---|
| 1 | 43 | 74 | 105 | 161 | 273 | 294 | |
| Alkanindiges | 6.88% | ||||||
| Dechloromonas | 4.57% | 26.43% | 1.95% | 0.45% | 0.15% | 0.05% | 0.11% |
| Acidovorax | 1.52% | 0.31% | 0.66% | 0.86% | 1.53% | ||
| Stenotrophomonas | 2.72% | 0.03% | |||||
| Propionivibrio | 1.94% | 0.04% | |||||
| Nitrosomonas | 1.63% | 4.25% | 3.09% | 0.15% | 0.23% | ||
| Flavobacterium | 1.61% | 0.42% | 0.09% | 0.1% | 0.06% | 0.22% | 0.05% |
| Thauera | 0.13% | 21.6% | 5.69% | 16.06% | 5.20% | 0.04% | 0.12% |
| Zoogloea | 0.38% | 8.15% | 18.32% | 26.00% | 64.26% | 11.9% | 50.13% |
| Sideroxydans | 3.99% | 0.02% | 0.14% | ||||
| Bdellovibrio | 0.11% | 1.69% | 0.96% | 0.26% | 0.08% | 0.10% | |
| Ferruginibacter | 0.26% | 1.07% | 0.66% | 0.39% | 0.11% | 1.58% | 0.95% |
| Simplicispira | 0.05% | 0.64% | 0.05% | 0.01% | 0.03% | ||
| Phreatobacter | 0.12% | 0.14% | 0.06% | 0.01% | |||
| Prosthecobacter | 3.43% | 2.57% | 1.16% | 2.76% | 1.1% | ||
| Aquimonas | 0.83% | 1.24% | 0.37% | 0.05% | |||
| Sphingopyxis | 1.8% | 1.02% | 0.04% | 0.01% | |||
| Chryseolinea | 0.05% | 0.33% | 2.64% | 2.26% | 0.18% | 0.83% | 0.42% |
| Terrimonas | 0.45% | 0.57% | 0.75% | 0.3% | 0.32% | 1.03% | 0.48% |
| Lacibacter | 0.05% | 0.06% | 0.04% | 0.01% | |||
| Azoarcus | 0.06% | 0.74% | 2.08% | 0.03% | 0.72% | 0.09% | |
| Phaeodactylibacter | 0.04% | 4.83% | 0.12% | ||||
| Ohtaekwangia | 0.02% | 0.11% | 0.55% | 1.09% | 8.67% | 2.61% | |
| Aggregicoccus | 0.11% | 0.01% | 0.02% | 0.04% | 0.13% | 0.04% | |
| Chryseobacterium | 0.04% | 0.26% | 0.01% | 0.36% | 2.65% | 5.63% | |
| Nitrospira | 0.47% | 0.01% | 0.01% | 0.41% | 4.06% | 0.76% | |
| Taibaiella | 0.98% | 0.36% | 2.43% | 10.68% | |||
| Tahibacter | 0.01% | 0.01% | 0.13% | 0.14% | 0.42% | 4.21% | 2.33% |
| Sediminibacterium | 0.01% | 0.13% | 0.49% | 0.27% | 0.01% | 1.32% | 0.49% |
| Luteimonas | 0.06% | 0.04% | 0.1% | 0.38% | |||
| Genus | Operation Day | ||||||
|---|---|---|---|---|---|---|---|
| 1 | 43 | 74 | 105 | 161 | 273 | 294 | |
| Alkanindiges | 6.88% | ||||||
| Dechloromonas | 4.57% | 26.01% | 2.2% | 1.11% | 0.43% | 0.02% | 0.10% |
| Acidovorax | 1.52% | 0.64% | 1.13% | 0.76% | 1.22% | ||
| Stenotrophomonas | 2.72% | 0.02% | |||||
| Propionivibrio | 1.94% | 0.03% | |||||
| Nitrosomonas | 1.63% | 5.45% | 5.84% | 0.66% | 0.24% | ||
| Flavobacterium | 1.61% | 0.22% | 0.24% | 0.45% | 0.29% | 0.76% | 0.05% |
| Thauera | 0.13% | 3.9% | 4.96% | 6.59% | 2.19% | 0.01% | 0.07% |
| Zoogloea | 0.38% | 8.53% | 26.11% | 38.93% | 60.09% | 21.13% | 54.16% |
| Sideroxydans | 3.81% | 0.01% | 0.06% | ||||
| Bdellovibrio | 0.11% | 2.09% | 0.26% | 1.10% | 0.06% | 0.13% | 0.01% |
| Ferruginibacter | 0.26% | 1.19% | 0.38% | 0.08% | 0.04% | 1.26% | 0.86% |
| Simplicispira | 0.05% | 2.11% | 0.08% | 0.01% | 0.01% | ||
| Phreatobacter | 1.53% | 0.16% | 0.16% | 0.01% | 0.02% | ||
| Prosthecobacter | 1.89% | 2.4% | 1.01% | ||||
| Aquimonas | 0.07% | 1.5% | 0.81% | 0.51% | |||
| Sphingopyxis | 0.44% | 0.10% | 0.07% | 0.07% | |||
| Chryseolinea | 0.05% | 1.17% | 3.97% | 2.30% | 1.04% | 0.14% | 0.03% |
| Terrimonas | 0.45% | 0.81% | 1.43% | 0.97% | 0.1% | 2.34% | 0.43% |
| Lacibacter | 0.05% | 3.82% | 3.64% | 0.05% | |||
| Azoarcus | 0.44% | 0.31% | 1.92% | 0.06% | 0.29% | 0.1% | |
| Phaeodactylibacter | 0.04% | 0.01% | 0.14% | 0.06% | |||
| Ohtaekwangia | 0.02% | 0.02% | 0.28% | 2.96% | 2.71% | ||
| Aggregicoccus | 0.11% | 0.17% | 0.01% | 1.04% | |||
| Chryseobacterium | 0.04% | 0.19% | 0.29% | 0.02% | 5.26% | 6.02% | |
| Nitrospira | 0.47% | 0.07% | 0.01% | 0.01% | 2.08% | 0.55% | |
| Taibaiella | 0.98% | 0.09% | 2.45% | 10.58% | |||
| Tahibacter | 0.01% | 0.03% | 0.36% | 0.03% | 0.1% | 0.01% | 1.79% |
| Sediminibacterium | 0.01% | 0.07% | 0.56% | 0.35% | 0.02% | 0.07% | 0.34% |
| Luteimonas | 0.06% | 0.06% | 2.54% | 0.38% | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Liu, Y.-Q.; Mao, S.; Steinberg, C.E.W.; Duan, W.; Chen, F. Reproducibility of Aerobic Granules in Treating Low-Strength and Low-C/N-Ratio Wastewater and Associated Microbial Community Structure. Processes 2022, 10, 444. https://doi.org/10.3390/pr10030444
Zhang H, Liu Y-Q, Mao S, Steinberg CEW, Duan W, Chen F. Reproducibility of Aerobic Granules in Treating Low-Strength and Low-C/N-Ratio Wastewater and Associated Microbial Community Structure. Processes. 2022; 10(3):444. https://doi.org/10.3390/pr10030444
Chicago/Turabian StyleZhang, Hongxing, Yong-Qiang Liu, Shichao Mao, Christain E. W. Steinberg, Wenyan Duan, and Fangyuan Chen. 2022. "Reproducibility of Aerobic Granules in Treating Low-Strength and Low-C/N-Ratio Wastewater and Associated Microbial Community Structure" Processes 10, no. 3: 444. https://doi.org/10.3390/pr10030444
APA StyleZhang, H., Liu, Y. -Q., Mao, S., Steinberg, C. E. W., Duan, W., & Chen, F. (2022). Reproducibility of Aerobic Granules in Treating Low-Strength and Low-C/N-Ratio Wastewater and Associated Microbial Community Structure. Processes, 10(3), 444. https://doi.org/10.3390/pr10030444