
Enzyme Immobilization and Co-Immobilization: Main Framework, Advances and Some Applications




Abstract
:1. Introduction
2. Enzyme Immobilization: Drivers, Limitations, and Metrics
3. Immobilization Methods
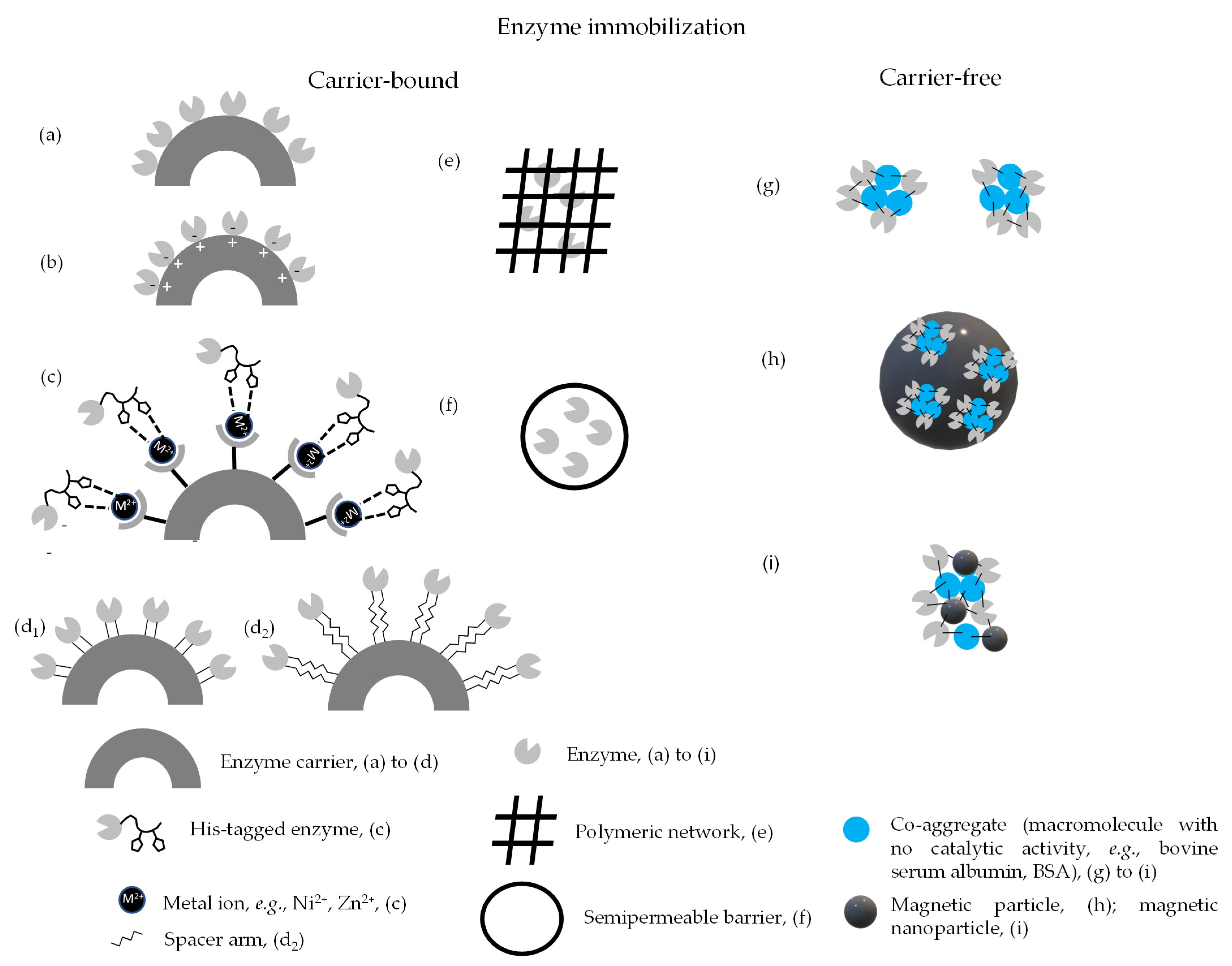
3.1. Classification of Immobilization Methods and Their Key Features
3.2. Some Significant Carrier Features and Their Impact in Biocatalyst Performance
3.2.1. Impact of the Carrier Hydrophilicity/Hydrophobicity
3.2.2. Geometry and Pore Size and Morphology of the Carrier
3.2.3. Effect of Spacer Arm
3.2.4. Effect of Carrier Loading
3.2.5. Carrier Composition and Immobilization Method
3.3. Immobilization of Multimeric Enzymes
4. Co-Immobilization of Enzymes

{kind=link}
{kind=link}
| Carrier Type | Main Features | Limitations | Immobilization Methods | Examples |
|---|---|---|---|---|
| Inorganic: |
| Limited biocompatibility, relatively low affinity to biomolecules and narrow range of geometrical shapes available or that can be created. Cross-linking agent, e.g., such as glutaraldehyde is typically needed to covalently bind the enzyme to the inorganic carrier | Adsorption/affinity/ionic binding/covalent binding to porous carriers; sol-gel entrapment/encapsulation | Carbon-based materials, ceramics (e.g., kaolinite, montmorillonite) [199], metal oxides (e.g., alumina [200], titanium oxide [201], zirconia [202]), magnetic particles (maghemite, magnetite) [203], inorganic sol-gel, e.g., silica-based sol-gel, [204] silica, silica derivatives and siliceous materials [205], porous glass [206], pumice stone [207] |
| Organic materials: | Typically display biocompatibility and affinity to biomolecules and enable the design of diverse geometry, thus tackling several shortcomings of inorganic material. Organic materials can be divided in natural polymers (biopolymers) and synthetic polymers. | |||
| Natural polymers | Exhibit high biocompatibility, biofunctionality, biostability, and biodegradability
| Shifts in chemical composition due to seasonality and/or location
| Entrapment/encapsulation most common due to easy gel formation; affinity/ionic binding/covalent binding to carriers also used | Agar-agar [208], agarose [209], albumin [210], alginate [211], carrageenan [212], cellulose, chitin/chitosan [213], collagen [214], gelatin [215], pectin [216], Sepharose (crosslinked agarose) [217], alginate/starch [217] |
| Sinthetic polymers | Polymeric chain assembled using selected monomers to fit the requirements of enzyme and process where the catalyst will be used
| Cost and complexity of carrier development | Entrapment/encapsulation most common due to easy gel formation; affinity/ionic binding/covalent binding to carriers also used | Amberlite [218], polyacrylamide (PAA) [219], polyamide (PA) [220] polyaniline [221], polystyrene (PS) [222], polyurethane (PU) [223], polyvinyl alcohol (PVA) [224] |
| Hybrid/composite materials | Combine organic and inorganic parts to maximize their benefits and minimize the shortcomings
| Complexity and cost New technology, stil with many unknowns | Immobilization methods: adsorption/affinity/ionic binding/covalent binding to porous carriers; sol-gel entrapment/encapsulation | Calcium phosphate/gold [225], metal organic frameworks [151] (e.g., zeolitic imidazolate based structures [226]), silica/carbon nanotubes [227] silica/titania/graphite [228] |
5. Microfluidics and (Co-)Immobilization
6. Some Applications
6.1. Co-Immobilization of Pectinases
6.1.1. Pectinases: General Aspects, Classification and Applications
6.1.2. Pectinases: Sources and Production Methods
6.1.3. Immobilized Pectinase Formulations: Some Relevant Examples
- (i)
- a-Amylase, pectinase, and cellulase in iron oxide magnetic nanoparticles functionalized with 3-aminopropyltriethoxysilane (APTES) followed by cross-linking with glutaraldehyde, which was used for the clarification of apple, pineapple, and grapes juices. Immobilization increased stability, namely the half-life increased by 2.4-fold as compared to the free enzymes within 50 to 70 °C. The immobilized form was also more stable than the free enzymes when incubated in acidic environment (pH 3.0 to 6.0). Under optimized glutaraldehyde concentration, the activity recovery of immobilized enzymes peaked (α-amylase 89%, pectinase 87%, and cellulase 84%) and encouraging reusability data was observed, as an average residual activity of 77% was obtained after eight successive batch runs [256]. The shelf life of free α-amylase, cellulase, and pectinase was found to be 54, 61 and 68%, whereas the immobilized form retained 93, 91, and 89% residual activity, respectively (in 20 days). The increased stability of the immobilized form was ascribed to the stabilization of the tertiary structure of the enzymes as an outcome of the cross-linking action of glutaraldehyde [258].
- (ii)
- Cellulase, pectinase, and xylanase were also immobilized in iron oxide magnetic nanoparticles functionalized with ATPES, followed by crosslinking with glutaraldehyde. Under optimized immobilization conditions, activity recovery of 85%, 81% and 76% were obtained for pectinase, cellulase and xylanase, respectively. Thermal stability was slightly enhanced within 55 to 75 °C, as the half-life of the immobilized form was marginally higher (under 2-fold) than that of the. Immobilization also marginally enhanced the stability upon incubation under pH 4.0 to 8.0 (up to 1.45-fold as compared to the respective free forms). As previously referred, the stabilization observed upon immobilization was ascribed to the cross-linking action of glutaraldehyde. The immobilized formulation depicted a residual activity slightly exceeding 85% after four consecutive batch runs. Afterwards, the residual activity decreased significantly to reach around 30% after eight consecutive batch runs. This pattern was tentatively associated with end-product inhibition and mechanical damage, which ultimately led to the deactivation of the immobilized enzymes. Immobilization also enhanced shelf-life, since after 36 days, the residual activity of xylanase, pectinase and cellulase increased 2.8-fold, 2.2-fold and 1.2-fold, respectively, as compared to the free form. The immobilized formulation was used in the extraction of piperine from black pepper, in the preparation of sugarcane cell protoplasts, and in the clarification of papaya juice. In all cases, the formulation was successfully used for at least three consecutive batch runs [259].
- (iii)
- Pectinase and cellulase for the clarification of fruit juices, were immobilized either by covalent binding to glutaraldehyde activated iron oxide magnetic nanoparticles functionalized with APTES (Enz-MNP) or as magnetic CLEAs (MN-CLEAs), through precipitation of both magnetic particles and enzymes followed by cross-linking with glutaraldehyde. The half-life at 60 ºC increased by an average of 2.4-fold and 1.4-fold for MN-CLEAs and Enz-MNP, respectively, as compared to the free enzymes. Again, stabilization was associated with chemical cross-linking involving glutaraldehyde, more so when MN-CLEAs are involved given the large number of multiples interactions between enzymes and enzymes and magnetic nanoparticles. Optimal temperature increased from 50 °C to 60 °C upon immobilization. Moreover, immobilization allowed for higher activities at high temperatures, e.g., at 80 °C MN-CLEAs and Enz-MNP displayed 84% and 67% of their maximal activities, respectively, as compared to 29% retained activity for the free form. The optimal pH was not changed by immobilization, yet at pH 3.0 MN-CLEA, displayed 80% of its maximal activity, compared with 30% of the free form. On the other hand, the free form depicted higher activities with pH 4.0 to 6.0. This behavior was associated with the ionization state of the microenvironment surrounding the active site due to the magnetic nanoparticles and/or glutaraldehyde. Recovered activities of 33.6% and 5.6% were observed for MN-CLEAs and Enz-MNP, respectively. Again, the vast multipoint interactions involving glutaraldehyde, enzymes and nanoparticles was suggested to have allowed a more assorted fraction of all enzymes than those allowed in Enz-MNP, hence increasing the activity of MN-CLEAs. Immobilization leading to Enz-MNP was suggested to have proceeded in such a manner that favorer pectinase binding over cellulase, therefore hampering overall catalytic activity. MN-CLEAs also outperformed Enz-MNP when operational stability was assessed. Hence, the former formulation displayed 10% residual activity after eight consecutive batch cycles, whereas the latter was fully exhausted after seven consecutive batch runs. This behavior was tentatively ascribed to several factors, e.g., enzyme leaching, thermal induced denaturation, mechanical or/and hydrodynamic stress. On the other hand, the shelf-life of the two immobilized formulation did not differ. Thus, the catalytic activity of both formulations remained unchanged for 30 days [256].
- (iv)
- Commercial preparations of pectinase (Pectinex®) and cellulase (Celluclast®) were immobilized in iron oxide magnetic nanoparticles functionalized with 3-aminopropyltriethoxysilane (APTES) followed by cross-linking with glutaraldehyde. Immobilization led to around 2.5-fold average increase in half-life within 50 to 70 °C [260]. Stabilization of the immobilized formulation was again related to glutaraldehyde cross-linking. Under optimized immobilization conditions, recovered activities of 92% and 87% were reported for pectinase and cellulase, respectively. The formulation displayed high operational stability, since 80% residual activity was reported after 10 consecutive batch runs. The immobilized formulation was for the extraction of antioxidant from real fruit waste peels, orange (Citrus sinensis), mango (Mangifera indica), and banana (Musa acuminate) [258].
- (v)
- Pectinase and cellulase in ferrite-based nanoparticles, functionalized with the agent, 3-aminopropyl triethoxysilane (APTES)/glutaraldehyde present a stability of the half-life of magnetic nanobiocatalyst increased by almost 3 folds as compared to the free form that confirmed enhanced thermal stability ranging from 30 to 60 °C, the enzyme loading of the xylanase (80 %) was observed, the reusability, as residual activity of immobilized biocatalyst was found to be 50 % after six consecutive cycles, the shelf life of the immobilized enzyme retained 90 % residual activity over 30 days, the application was in the extraction of lycopene from tomato peels [261,262,263]; α-amylase, pectinase and cellulase using covalent co-immobilization with use of glutaraldehyde as crosslinker, originated two fold increment in half-life, enhanced pH stability, and reusability for eight reaction cycles, the applicability is in fruit juice clarification [264,265].
6.2. Co-Immobilization of Glucosidases
7. Conclusions and Future Work
Author Contributions
Funding
Conflicts of Interest
References
- Sheldon, R.A. Metrics of Green Chemistry and Sustainability: Past, Present and Future. ACS Sustain. Chem. Eng. 2018, 6, 32–48. [Google Scholar] [CrossRef] [Green Version]
- Adams, J.P.; Brown, M.J.B.; Diaz-Rodriguez, A.; Lloyd, R.C.; Roiban, G.D. Biocatalysis: A Pharma Perspective. Adv. Synth. Catal. 2019, 361, 2421. [Google Scholar] [CrossRef] [Green Version]
- Heckmann, C.M.; Paradisi, F. Looking Back: A Short History of the Discovery of Enzymes and How They Became Powerful Chemical Tools. ChemCatChem 2020, 12, 6082–6102. [Google Scholar] [CrossRef]
- Choi, J.M.; Han, S.S.; Kim, H.S. Industrial applications of enzyme biocatalysis: Current status and future aspects. Biotechnol. Adv. 2015, 33, 1443–1454. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Kumar, M.; Mittal, A.; Mehta, P.K. Microbial enzymes: Industrial progress in 21st century. 3 Biotech 2016, 6, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chapman, J.; Ismail, A.E.; Dinu, C.Z. Industrial Applications of Enzymes: Recent Advances, Techniques, and Outlooks. Catalysts 2018, 8, 238. [Google Scholar] [CrossRef] [Green Version]
- Wackett, L.P. Microbial industrial enzymes: An annotated selection of World Wide Web sites relevant to the topics in microbial biotechnology. Microb. Biotechnol. 2019, 12, 405–406. [Google Scholar] [CrossRef] [Green Version]
- Abdelraheem, E.M.M.; Busch, H.; Hanefeld, U.; Tonin, F. Biocatalysis explained: From pharmaceutical to bulk chemical production. React. Chem. Eng. 2019, 4, 1878–1894. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Snajdrova, R.; Moore, J.C.; Baldenius, K.; Bornscheuer, U.T. Biocatalysis: Enzymatic Synthesis for Industrial Applications. Angew. Chem. Int. Ed. Engl. 2021, 60, 88–119. [Google Scholar] [CrossRef] [PubMed]
- Aleu, J.; Bustillo, A.J.; Hernandez-Galan, R.; Collado, I.G. Biocatalysis Applied to the Synthesis of Agrochemicals. Curr. Org. Chem. 2006, 10, 2037–2054. [Google Scholar] [CrossRef]
- Madhu, A.; Chakraborty, J.N. Developments in application of enzymes for textile processing. J. Clean. Prod. 2017, 145, 114–133. [Google Scholar] [CrossRef]
- Sá, A.G.A.; de Meneses, A.C.; de Araújo, P.H.H.; de Oliveira, D. A review on enzymatic synthesis of aromatic esters used as flavor ingredients for food, cosmetics and pharmaceuticals industries. Trends Food Sci. Technol. 2017, 69, 95–105. [Google Scholar] [CrossRef]
- Yarosh, D.B.; Rosenthal, A.; Moy, R. Six critical questions for DNA repair enzymes in skincare products: A review in dialog. Clin. Cosmet. Investig. Dermatol. 2019, 12, 617–624. [Google Scholar] [CrossRef] [Green Version]
- Woodley, J.M. Towards the sustainable production of bulk-chemicals using biotechnology. N. Biotechnol. 2020, 59, 59–64. [Google Scholar] [CrossRef]
- Gürkök, S. Microbial enzymes in detergents: A review. Int. J. Sci. Eng. Res. 2019, 10, 75–81. [Google Scholar]
- Al-Ghanayem, A.A.; Joseph, B. Current prospective in using cold-active enzymes as eco-friendly detergent additive. Appl. Microbiol. Biotechnol. 2020, 104, 2871–2882. [Google Scholar] [CrossRef] [PubMed]
- Al-Hemaid, F.; Chandrasekaran, M. Enzymes in Processing of Nutraceuticals. In Enzymes in Food and Beverage Processing; Chandrasekaran, M., Ed.; CRC Press: Boca Raton, FL, USA, 2015; pp. 401–423. [Google Scholar]
- Raveendran, S.; Parameswaran, B.; Ummalyma, S.B.; Abraham, A.; Mathew, A.K.; Madhavan, A.; Rebello, S.; Pandey, A. Applications of Microbial Enzymes in Food Industry. Food Technol. Biotechnol. 2018, 56, 16–30. [Google Scholar] [CrossRef] [PubMed]
- Khambhaty, Y. Applications of enzymes in leather processing. Environ. Chem. Lett. 2020, 18, 747–769. [Google Scholar] [CrossRef]
- Singh, G.; Capalash, N.; Kaur, K.; Puri, S.; Sharma, P. Enzymes: Applications in pulp and paper industry. In Agro-Industrial Wastes as Feedstock for Enzyme Production: Apply and Exploit the Emerging and Valuable Use Options of Waste Biomass; Dhillon, G., Kaur, S., Eds.; Academic Press: San Diego, CA, USA, 2016; pp. 157–172. [Google Scholar]
- Meghwanshi, G.K.; Kaur, N.; Verma, S.; Dabi, N.K.; Vashishtha, A.; Charan, P.D.; Purohit, P.; Bhandari, H.; Bhojak, N.; Kumar, R. Enzymes for pharmaceutical and therapeutic applications. Biotechnol. Appl. Biochem. 2020, 67, 586–601. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Ortiz, C.; Berenguer-Murcia, A.; Torres, R.; Fernandez-Lafuente, R. Modifying enzyme activity and selectivity by immobilization. Chem. Soc. Rev. 2013, 42, 6290–6307. [Google Scholar] [CrossRef]
- Sheldon, R.A.; Woodley, J.M. Role of Biocatalysis in Sustainable Chemistry. Chem. Rev. 2018, 118, 801–838. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.; Martins, M.; Jing, S.; Fu, J.; Cavaco-Paulo, A. Practical insights on enzyme stabilization. Crit. Rev. Biotechnol. 2018, 38, 335–350. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A. CLEAs, Combi-CLEAs and ‘Smart’ Magnetic CLEAs: Biocatalysis in a Bio-Based Economy. Catalysts 2019, 9, 261. [Google Scholar] [CrossRef] [Green Version]
- Basso, A.; Serban, S. Industrial applications of immobilized enzymes—A review. Mol. Catal. 2019, 479, 110607. [Google Scholar] [CrossRef]
- Romero-Fernández, M.; Paradisi, F. General overview on immobilization techniques of enzymes for biocatalysis. In Catalysts Immobillization; Benaglia, M., Puglisi, A., Eds.; John Wiley & Sons, Ltd.: New York, NY, USA, 2019; pp. 409–435. [Google Scholar]
- Thompson, M.P.; Derrington, S.R.; Heath, R.S.; Porter, J.L.; Mangas-Sanchez, J.; Devine, P.N.; Truppo, M.D.; Turner, N.J. A generic platform for the immobilisation of engineered biocatalysts. Tetrahedron 2019, 75, 327–334. [Google Scholar] [CrossRef]
- Federsel, H.J.; Moody, T.S.; Taylor, S. Recent Trends in Enzyme Immobilization-Concepts for Expanding the Biocatalysis Toolbox. Molecules 2021, 26, 2822. [Google Scholar] [CrossRef]
- Sheldon, R.A.; Brady, D. Streamlining Design, Engineering, and Applications of Enzymes for Sustainable Biocatalysis. ACS Sustain. Chem. Eng. 2021, 9, 8032–8052. [Google Scholar] [CrossRef]
- Taheri-Kafrani, A.; Kharazmi, S.; Nasrollahzadeh, M.; Soozanipour, A.; Ejeian, F.; Etedali, P.; Mansouri-Tehrani, H.A.; Razmjou, A.; Yek, S.M.G.; Varma, R.S. Recent developments in enzyme immobilization technology for high-throughput processing in food industries. Crit. Rev. Food Sci. Nutr. 2021, 61, 3160–3196. [Google Scholar] [CrossRef] [PubMed]
- Nidetzky, B.; Bolivar, J. Continuous biocatalytic processing and synthesis: Variations on a theme of “flow chemistry”. Chim. Oggi-Chem. Today 2015, 33, 20–24. [Google Scholar]
- Bergquist, P.L.; Siddiqui, S.; Sunna, A. Cell-Free Biocatalysis for the Production of Platform Chemicals. Front. Energy Res. 2020, 8, 193. [Google Scholar] [CrossRef]
- Barbosa, O.; Ortiz, C.; Berenguer-Murcia, Á.; Torres, R.; Rodrigues, R.C.; Fernandez-Lafuente, R. Strategies for the one-step immobilization-purification of enzymes as industrial biocatalysts. Biotechnol. Adv. 2015, 33, 435–456. [Google Scholar] [CrossRef] [Green Version]
- Cai, L.; Chu, Y.; Liu, X.; Qiu, Y.; Ge, Z.; Zhang, G. A novel all-in-one strategy for purification and immobilization of β-1,3-xylanase directly from cell lysate as active and recyclable nanobiocatalyst. Microb. Cell Factories 2021, 20, 37. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, N.R.; Marzuki, N.H.C.; Buang, N.A.; Huyop, F.; Wahab, R.A. An overview of technologies for immobilization of enzymes and surface analysis techniques for immobilized enzymes. Biotechnol. Biotechnol. Equip. 2015, 29, 205–220. [Google Scholar] [CrossRef] [PubMed]
- Sheldon, R.A. Enzyme Immobilization: The Quest for Optimum Performance. Adv. Synth. Catal. 2007, 349, 1289–1307. [Google Scholar] [CrossRef]
- Romero-Fernández, M.; Paradisi, F. Protein immobilization technology for flow biocatalysis. Curr. Opin. Chem. Biol. 2020, 55, 1–8. [Google Scholar] [CrossRef]
- Hwang, E.T.; Gu, M.B. Enzyme stabilization by nano/microsized hybrid materials. Eng. Life Sci. 2013, 13, 49–61. [Google Scholar] [CrossRef]
- Bilal, M.; Iqbal, H.M.N. Chemical, physical, and biological coordination: An interplay between materials and enzymes as potential platforms for immobilization. Coord. Chem. Rev. 2019, 388, 1–23. [Google Scholar] [CrossRef]
- An, J.; Li, G.; Zhang, Y.; Zhang, T.; Liu, X.; Gao, F.; Peng, M.; He, Y.; Fan, H. Recent Advances in Enzyme-Nanostructure Biocatalysts with Enhanced Activity. Catalysts 2020, 10, 338. [Google Scholar] [CrossRef] [Green Version]
- Correa, S.; Ripoll, M.; Jackson, E.; Grazú, V.; Betancor, L. Stabilization of β-Glucuronidase by Immobilization in Magnetic-Silica Hybrid Supports. Catalysts 2020, 10, 669. [Google Scholar] [CrossRef]
- Singh, N.; Dhanya, B.S.; Verma, M.L. Nano-immobilized biocatalysts and their potential biotechnological applications in bioenergy production. Mater. Sci. Energy Technol. 2020, 3, 808–824. [Google Scholar] [CrossRef]
- Bolivar, J.M.; Nidetzky, B. Positively Charged Mini-Protein Zbasic2 as a Highly Efficient Silica Binding Module: Opportunities for Enzyme Immobilization on Unmodified Silica Supports. Langmuir 2012, 28, 10040–10049. [Google Scholar] [CrossRef]
- Rueda, N.; Albuquerque, T.L.; Bartolome-Cabrero, R.; Fernandez-Lopez, L.; Torres, R.; Ortiz, C.; Dos Santos, J.C.S.; Barbosa, O.; Fernandez-Lafuente, R. Reversible Immobilization of Lipases on Heterofunctional Octyl-Amino Agarose Beads Prevents Enzyme Desorption. Molecules 2016, 21, 646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateo, C.; Pessela, B.C.C.; Fuentes, M.; Torres, R.; Ortiz, C.; López-Gallego, F.; Betancor, L.; Alonso-Morales, N.; Guisan, J.M.; Fernandez-Lafuente, R. Very Strong but Reversible Immobilization of Enzymes on Supports Coated with Ionic Polymers. In Immobilization of Enzymes and Cells. Methods and Protocols; Guisan, J., Bolivar, J., López-Gallego, F., Rocha-Martín, J., Eds.; Humana: New York, NY, USA, 2020; Volume 2100, pp. 129–141. [Google Scholar]
- Del Monte-Martínez, A.; Cutiño-Avila, B.V. Rational design of immobilized lipases and phospholipases. Methods Mol. Biol. 2012, 861, 343–382. [Google Scholar] [PubMed]
- Sastre, D.E.; Reis, E.A.; Marques Netto, C.G.C. Strategies to rationalize enzyme immobilization procedures. Methods Enzymol. 2020, 630, 81–110. [Google Scholar] [PubMed]
- Roura Padrosa, D.; Marchini, V.; Paradisi, F. CapiPy: Python based GUI-application to assist in protein immobilization. Bioinformatics 2021, 18, btab030. [Google Scholar] [CrossRef] [PubMed]
- Range, J.; Halupczok, C.; Lohmann, J.; Swainston, N.; Kettner, C.; Bergmann, F.T.; Weidemann, A.; Wittig, U.; Schnell, S.; Pleiss, J. EnzymeML-a data exchange format for biocatalysis and enzymology. FEBS J. 2021. [Google Scholar] [CrossRef]
- Secundo, F. Conformational changes of enzymes upon immobilisation. Chem. Soc. Rev. 2013, 42, 6250–6261. [Google Scholar] [CrossRef] [PubMed]
- Liese, A.; Hilterhaus, L. Evaluation of immobilized enzymes for industrial applications. Chem. Soc. Rev. 2013, 42, 6236–6249. [Google Scholar] [CrossRef] [PubMed]
- Dias Gomes, M.; Woodley, J.M. Considerations when Measuring Biocatalyst Performance. Molecules 2019, 24, 3573. [Google Scholar] [CrossRef] [Green Version]
- Velasco-Lozano, S. Immobilization of Enzymes as Cross-Linked Enzyme Aggregates: General Strategy to Obtain Robust Biocatalysts. In Immobilization of Enzymes and Cells. Methods in Molecular Biology; Guisan, J., Bolivar, J., López-Gallego, F., Rocha-Martín, J., Eds.; Humana: New York, NY, USA, 2020; Volume 2100, pp. 345–361. [Google Scholar]
- Bommarius, A.S.; Paye, M.F. Stabilizing biocatalysts. Chem. Soc. Rev. 2013, 42, 6534–6565. [Google Scholar] [CrossRef] [PubMed]
- Bommarius, A.S. Biocatalysis: A Status Report. Annu. Rev. Chem. Biomol. Eng. 2015, 6, 319–345. [Google Scholar] [CrossRef]
- Bolivar, J.M.; López-Gallego, F. Characterization and evaluation of immobilized enzymes for applications in flow reactors. Curr. Opin. Green Sustain. Chem. 2020, 25, 100349. [Google Scholar] [CrossRef]
- Woodley, J.M. New frontiers in biocatalysis for sustainable synthesis. Curr. Opin. Green Sustain. Chem. 2020, 21, 22–26. [Google Scholar] [CrossRef]
- Sheldon, R.A.; van Pelt, S. Enzyme immobilisation in biocatalysis: Why, what and how. Chem. Soc. Rev. 2013, 42, 6223–6235. [Google Scholar] [CrossRef] [Green Version]
- Boudrant, J.; Woodley, J.M.; Fernandez-Lafuente, R. Parameters necessary to define an immobilized enzyme preparation. Process. Biochem. 2020, 90, 66–80. [Google Scholar] [CrossRef]
- Singhania, R.R.; Patel, A.K.; Pandey, A. The Industrial Production of Enzymes. In Industrial Biotechnology; Soetaert, W., Vandamme, E.J., Eds.; Wiley-VCH Verlag GmbH & Co.: Weinheim, Germany, 2021; pp. 207–225. [Google Scholar]
- Agyei, D.; Shanbhag, B.K.; He, L. Enzyme engineering (immobilization) for food applications. In Improving and Tailoring Enzymes for Food Quality and Functionality; Yada, R.Y., Ed.; Elsevier: Amsterdam, The Netherlands, 2015; pp. 213–236. [Google Scholar]
- Brena, B.; González-Pombo, P.; Batista-Viera, F. Immobilization of enzymes: A literature survey. Methods Mol. Biol. 2009, 1051, 15–31. [Google Scholar]
- Jesionowski, T.; Zdarta, J.; Krajewska, B. Enzyme immobilization by adsorption: A review. Adsorption 2014, 20, 801–821. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, M.; Amaral, J.C.; Lopes, L.A.; Fernandez-Lafuente, R.; Tardioli, P.W. Stabilization and operational selectivity alteration of Lipozyme 435 by its coating with polyethyleneimine: Comparison of the biocatalyst performance in the synthesis of xylose fatty esters. Int. J. Biol. Macromol. 2021, 192, 665–674. [Google Scholar] [CrossRef]
- Rueda, N.; Santos, C.S.D.; Rodriguez, M.D.; Albuquerque, T.L.; Barbosa, O.; Torres, R.; Ortiz, C.; Fernandez-Lafuente, R. Reversible immobilization of lipases on octyl-glutamic agarose beads: A mixed adsorption that reinforces enzyme immobilization. J. Mol. Catal. B Enzym. 2016, 128, 10–18. [Google Scholar] [CrossRef]
- Sheldon, R.A.; Basso, A.; Brady, D. New frontiers in enzyme immobilisation: Robust biocatalysts for a circular bio-based economy. Chem. Soc. Rev. 2021, 50, 5850–5862. [Google Scholar] [CrossRef]
- Zhou, L.J.; Li, R.F.; Li, X.Y.; Zhang, Y.W. One-step selective affinity purification and immobilization of His-tagged enzyme by recyclable magnetic nanoparticles. Eng. Life Sci. 2021, 21, 364–373. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, J.; Zhuang, N.; Zhang, A.; Chen, K.; Xu, N.; Xin, F.; Zhang, W.; Dong, W.; Jiang, M. Immobilization and Purification of Enzymes with the Novel Affinity Tag ChBD-AB from Chitinolyticbacter meiyuanensis SYBC-H1. Front. Bioeng. Biotechnol. 2020, 8, 579. [Google Scholar] [CrossRef]
- Wahab, R.A.; Elias, N.; Abdullah, F.; Ghoshal, S.K. On the taught new tricks of enzymes immobilization: An all-inclusive overview. React. Funct. Polym. 2020, 152, 104613. [Google Scholar] [CrossRef]
- Fernández-Lorente, G.; Lopez-Gallego, F.; Bolivar, J.M.; Rocha-Martin, J.; Moreno-Perez, S.; Guisán, J.M. Immobilization of proteins on highly activated glyoxyl supports: Dramatic increase of the enzyme stability via multipoint immobilization on pre-existing carriers. Curr. Org. Chem. 2015, 19, 1719–1731. [Google Scholar] [CrossRef] [Green Version]
- Weltz, J.S.; Kienle, D.F.; Schwart, D.K.; Kaar, J.L. Reduced Enzyme Dynamics upon Multipoint Covalent Immobilization Leads to Stability-Activity Trade-off. J. Am. Chem. Soc. 2020, 142, 3463–3471. [Google Scholar] [CrossRef] [PubMed]
- Novick, S.J.; Rozzell, J.D. Immobilization of Enzymes by Covalent Attachment. In Microbial Enzymes and Biotransformations. Methods in Biotechnology; Barredo, J.L., Ed.; Humana Press: Totowa, NJ, USA, 2005; Volume 17, pp. 247–271. [Google Scholar]
- Orrego, H.A.; Romero-Fernández, M.; Millán-Linares, M.C.; Pedroche, J.; Guisán, J.M.; Rocha-Martin, J. High Stabilization of Enzymes Immobilized on Rigid Hydrophobic Glyoxyl-Supports: Generation of Hydrophilic Environments on Support Surfaces. Catalysts 2020, 10, 676. [Google Scholar] [CrossRef]
- Bolivar, J.M.; Rocha-Martin, J.; Mateo, C.; Cava, F.; Berenguer, J.; Fernandez-Lafuente, R.; Guisan, J.M. Coating of soluble and immobilized enzymes with ionic polymers: Full stabilization of the quaternary structure of multimeric enzymes. Biomacromolecules 2009, 10, 742–747. [Google Scholar] [CrossRef] [PubMed]
- Roura Padrosa, D.; Benítez-Mateos, A.I.; Calvey, L.; Paradisi, F. Cell-free biocatalytic syntheses of l-pipecolic acid: A dual strategy approach and process intensification in flow. Green Chem. 2020, 22, 5310–5316. [Google Scholar] [CrossRef]
- Ovsejevi, K.; Manta, C.; Batista-Viera, F. Reversible covalent immobilization of enzymes via disulfide bonds. Methods Mol. Biol. 2013, 1051, 89–116. [Google Scholar] [PubMed]
- Guzik, U.; Hupert-Kocurek, K.; Marchlewicz, A.; Wojcieszyńska, D. Enhancement of biodegradation potential of catechol 1,2-dioxygenase through its immobilization in calcium alginate gel. Electron. J. Biotechnol. 2014, 17, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Schmieg, B.; Schimek, A.; Franzreb, M. Development and performance of a 3D-printable poly(ethylene glycol) diacrylate hydrogel suitable for enzyme entrapment and long-term biocatalytic applications. Eng. Life Sci. 2018, 18, 659–667. [Google Scholar] [CrossRef] [Green Version]
- Meyer, J.; Meyer, L.-E.; Kara, S. Enzyme immobilization in hydrogels: A perfect liaison for efficient and sustainable biocatalysis. Eng. Life. Sci. 2021, in press. [Google Scholar] [CrossRef]
- Cao, L.; van Langen, L.; Sheldon, R.A. Immobilised enzymes: Carrier-bound or carrier-free? Curr. Opin. Biotechnol. 2003, 14, 387–394. [Google Scholar] [CrossRef]
- Xu, D.Y.; Yang, Z. Cross-linked tyrosinase aggregates for elimination of phenolic compounds from wastewater. Chemosphere 2013, 92, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Minteer, S.D. Cell-free biotechnologies. In Biotechnology for Biofuel Production and Optimization; Eckert, C.A., Trinh, C.T., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 433–448. [Google Scholar]
- Husain, Q.; Ansari, S.A.; Alam, F.; Azam, A. Immobilization of Aspergillus oryzae β galactosidase on zinc oxide nanoparticles via simple adsorption mechanism. Int. J. Biol. Macromol. 2011, 49, 37–43. [Google Scholar] [CrossRef]
- Karakuş, E.; Pekyardımcı, S. Comparison of Covalent and Noncovalent Immobilization of Malatya Apricot Pectinesterase (Prunus armeniaca L.). Artif. Cells Blood Substit. Immobil. Biotechnol. 2012, 40, 132–141. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, N.M.; Garcia, K.C.; Bussamara, R.; Both, F.S.; Vainstein, M.H.; Soares, R.M.D. Poly (lactic acid)/chitosan fiber mats: Investigation of effects of the support on lipase immobilization. Int. J. Biol. Macromol. 2015, 72, 998–1004. [Google Scholar] [CrossRef]
- Lee, C.H.; Jin, E.S.; Lee, J.H.; Hwang, E.T. Immobilization and Stabilization of Enzyme in Biomineralized Calcium Carbonate Microspheres. Front. Bioeng. Biotechnol. 2020, 8, 553591. [Google Scholar] [CrossRef]
- Arıca, M.Y.; Bayramoğlu, G. Invertase reversibly immobilized onto polyethylenimine-grafted poly(GMA-MMA) beads for sucrose hydrolysis. J. Mol. Catal. B Enzym. 2006, 38, 131–138. [Google Scholar] [CrossRef]
- Cabral, B.V.; Santos, L.D.; Larissa, N.S.; Falleiros, S.; Taciana, S.C.; Freitas, F.F.; Cardoso, S.L. Sucrose hydrolysis by invertase immobilized on Duolite A-568 employing a packed-bed reactor. Chem. Eng. Commun. 2017, 204, 1007–1019. [Google Scholar] [CrossRef]
- Morshed, M.N.; Behary, N.; Guan, J.P.; Nierstrasz, V.A. Immobilizing redox enzyme on amino functional group-integrated tailor-made polyester textile: High loading, stability, and application in a bio-fenton system. ACS Sustain. Chem. Eng. 2021, 9, 8879–8894. [Google Scholar] [CrossRef]
- Dal Magro, L.; Kornecki, J.F.; Klein, M.P.; Rodrigues, R.C.; Fernandez-Lafuente, R. Optimized immobilization of polygalacturonase from Aspergillus niger following different protocols: Improved stability and activity under drastic conditions. Int. J. Biol. Macromol. 2019, 138, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.E.; Yang, Q.; Xiao, Z. Covalent immobilization of glucoamylase enzyme onto chemically activated surface of κ-carrageenan. Bull. Natl. Res. Cent. 2019, 43, 102. [Google Scholar] [CrossRef] [Green Version]
- Alagöz, D.; Toprak, A.; Yildirim, D.; Tükel, S.S.; Fernandez-Lafuente, R. Modified silicates and carbon nanotubes for immobilization of lipase from Rhizomucor miehei: Effect of support and immobilization technique on the catalytic performance of the immobilized biocatalysts. Enzym. Microb. Technol. 2021, 144, 109739. [Google Scholar] [CrossRef] [PubMed]
- Alagöz, D.; Varan, N.E.; Toprak, A.; Yildirim, D.; Tukel, S.S.; Fernandez-Lafuente, R. Immobilization of xylanase on differently functionalized silica gel supports for orange juice clarification. Process. Biochem. 2022, 113, 270–280. [Google Scholar] [CrossRef]
- Nawaz, M.A.; Rehman, H.U.; Bibi, Z.; Aman, A.; Ul Qader, S.A. Continuous degradation of maltose by enzyme entrapment technology using calcium alginate beads as a matrix. Biochem. Biophys. Rep. 2015, 4, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghu, S.; Pennathur, G. Enhancing the stability of a carboxylesterase by entrapment in chitosan coated alginate beads. Turk. J. Biol. 2018, 42, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Qi, D.; Gao, M.; Li, X.; Lin, J. Immobilization of Pectinase onto Porous Hydroxyapatite/Calcium Alginate Composite Beads for Improved Performance of Recycle. ACS Omega 2020, 5, 20062–20069. [Google Scholar] [CrossRef] [PubMed]
- Suvarli, N.; Wenger, L.; Serra, C.; Perner-Nochta, I.; Hubbuch, J.; Wörner, M. Immobilization of β-Galactosidase by Encapsulation of Enzyme-Conjugated Polymer Nanoparticles Inside Hydrogel Microparticles. Front. Bioeng. Biotechnol. 2022, 9, 818053. [Google Scholar] [CrossRef]
- Rasbold, L.M.; Delai, V.M.; da Cruz Kerber, C.M.; Simões, M.R.; Heinen, P.R.; da Conceição Silva, J.L.; Simão, R.C.G.; Kadowaki, M.K.; Maller, A. Production, immobilization and application of invertase from new wild strain Cunninghamella echinulata PA3S12MM. J. Appl. Microbiol. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- van Pelt, S.; Quignard, S.; Kubac, D.; Dimitry, Y.S.B.; van Rantwijk, F.; Sheldon, R.A. Nitrile hydratase CLEAs: The immobilization and stabilization of an industrially important enzyme. Green Chem. 2008, 10, 395–400. [Google Scholar] [CrossRef]
- Nadar, S.S.; Muley, A.B.; Ladole, M.R.; Joshi, P.U. Macromolecular cross-linked enzyme aggregates (M-CLEAs) of α-amylase. Int. J. Biol. Macromol. 2016, 84, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Rehman, S.; Bhatti, H.N.; Bilal, M.; Asgher, M. Cross-linked enzyme aggregates (CLEAs) of Pencilluim notatum lipase enzyme with improved activity, stability and reusability characteristics. Int. J. Biol. Macromol. 2016, 91, 1161–1169. [Google Scholar] [CrossRef] [PubMed]
- Zang, Y.Y.; Yang, S.; Xu, Y.Q.; Chen, Z.G.; Wu, T. Carrier-Free Immobilization of Rutin Degrading Enzyme Extracted From Fusarium spp. Front. Bioeng. Biotechnol. 2020, 8, 470. [Google Scholar] [CrossRef] [PubMed]
- Yan, H.; Shi, B.; Ding, J.; Zhu, H.; Li, X.; Qian, J. Preparation and characterization of a nontoxic cross-linked lipase aggregate by using sodium tripolyphosphate and chitosan. J. Appl. Polym. Sci. 2022, 139, e51871. [Google Scholar] [CrossRef]
- Doraiswamy, N.; Sarathi, M.; Pennathur, G. Cross-linked esterase aggregates (CLEAs) using nanoparticles as immobilization matrix. Prep. Biochem. Biotechnol. 2019, 49, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Paitaid, P.; H-Kittikun, A. Magnetic Cross-Linked Enzyme Aggregates of Aspergillus oryzae ST11 Lipase Using Polyacrylonitrile Coated Magnetic Nanoparticles for Biodiesel Production. Appl. Biochem. Biotechnol. 2020, 190, 1319–1332. [Google Scholar] [CrossRef] [PubMed]
- Lucena, G.N.; dos Santos, C.C.; Pinto, G.C.; Piazza, R.D.; Guedes, W.N.; Jafelicci Junior, M.; de Paula, A.V.; Marques, R.F.C. Synthesis and characterization of magnetic cross-linked enzyme aggregate and its evaluation of the alternating magnetic field (AMF) effects in the catalytic activity. J. Magn. Magn. Mater. 2020, 516, 167326. [Google Scholar] [CrossRef]
- Wei, B.; Liu, F.; Liu, X.; Cheng, L.; Yuan, O.; Gao, H.; Liang, H. Enhancing stability and by-product tolerance of β-glucuronidase based on magnetic cross-linked enzyme aggregates. COLSUB 2022, 210, 112241. [Google Scholar] [CrossRef]
- Faccio, G. From Protein Features to Sensing Surfaces. Sensors 2018, 18, 1204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benítez-Mateos, A.I.; Contente, M.L. Agarose vs. Methacrylate as Material Supports for Enzyme Immobilization and Continuous Processing. Catalysts 2021, 11, 814. [Google Scholar] [CrossRef]
- Engelmann, C.; Ekambaram, N.; Johannsen, J.; Fellechner, O.; Waluga, T.; Fieg, G.; Liese, A.; Bubenheim, P. Enzyme Immobilization on Synthesized Nanoporous Silica Particles and their Application in a Bi-enzymatic Reaction. ChemCatChem 2020, 12, 2245–2252. [Google Scholar] [CrossRef]
- Kahar, U.M.; Sani, M.H.; Chan, K.-G.; Goh, K.M. Immobilization of α-Amylase from Anoxybacillus sp. SK3-4 on ReliZyme and Immobead Supports. Molecules 2016, 21, 1196. [Google Scholar] [CrossRef] [Green Version]
- Dinu, C.Z.; Zhu, G.; Bale, S.S.; Anand, G.; Reeder, P.J.; Sanford, K.; Whited, G.; Kane, R.S.; Dordick, J.S. Enzyme-based nanoscale composites for use as active decontamination surfaces. Adv. Funct. Mater. 2010, 20, 392–398. [Google Scholar] [CrossRef]
- Moskovitz, Y.; Srebnik, S. Conformational changes of globular proteins upon adsorption on a hydrophobic surface. Phys. Chem. Chem. Phys. 2014, 16, 11698–11707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, G.; Sharma, S.; Dutta, A.K.; Kumar, S.K.; Belfort, G. Conformational Transitions of Adsorbed Proteins on Surfaces of Varying Polarity. Langmuir 2010, 26, 10803–10811. [Google Scholar] [CrossRef] [PubMed]
- Sethuraman, A.; Vedantham, G.; Imoto, T.; Przybycien, T.; Belfort, G. Protein Unfolding at Interfaces: Slow Dynamics of α-Helix to β-Sheet Transition. Proteins 2004, 56, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Jin, Q.; Jia, G.; Zhang, Y.; Yang, Q.; Li, C. Hydrophobic Surface Induced Activation of Pseudomonas Cepacia Lipase Immobilized into Mesoporous Silica. Langmuir 2011, 27, 12016–12024. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Lopez, L.; Rueda, N.; Bartolome-Cabrero, R.; Rodriguez, M.D.; Albuquerque, T.L.; Dos Santos, J.C.S.; Barbosa, O.; Fernandez-Lafuente, R. Improved immobilization and stabilization of lipase from Rhizomucor miehei on octyl-glyoxyl agarose beads by using CaCl2. Process. Biochem. 2016, 51, 48–52. [Google Scholar] [CrossRef]
- Rodrigues, R.C.; Virgen-Ortíz, J.J.; dos Santos, J.C.S.; Berenguer-Murcia, Á.; Alcantara, A.R.; Barbosa, O.; Ortiz, C.; Fernandez-Lafuente, R. Immobilization of lipases on hydrophobic supports: Immobilization mechanism, advantages, problems, and solutions. Biotechnol. Adv. 2019, 37, 746–770. [Google Scholar] [CrossRef] [Green Version]
- Benítez-Mateos, A.I.; Huber, C.; Nidetzky, B.; Bolivar, J.M.; López-Gallego, F. Design of the Enzyme–Carrier Interface to Overcome the O2 and NADH Mass Transfer Limitations of an Immobilized Flavin Oxidase. ACS Appl. Mater. Interfaces 2020, 12, 56027–56038. [Google Scholar] [CrossRef]
- Grimaldi, J.; Radhakrishna, M.; Kumar, S.K.; Belfort, G. Stability of proteins on hydrophilic surfaces. Langmuir 2015, 31, 1005–1010. [Google Scholar] [CrossRef] [PubMed]
- Bayne, L.; Ulijn, R.V.; Halling, P.J. Effect of pore size on the performance of immobilised enzymes. Chem. Soc. Rev. 2013, 42, 9000–9010. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.; Qiu, J.; Mo, H.; Yu, Y.; Ito, K.; Sakai, E.; Feng, H. Synthesis of mesoporous silica with different pore sizes for cellulase immobilization: Pure physical adsorption. New J. Chem. 2017, 41, 9338–9345. [Google Scholar] [CrossRef]
- Takimoto, A.; Shiomi, T.; Ino, K.; Tsunoda, T.; Kawai, A.; Mizukami, F.; Sakaguchi, K. Encapsulation of Cellulase with Mesoporous Silica (SBA-15). Microporous Mesoporous Mater. 2008, 116, 601–606. [Google Scholar] [CrossRef]
- Kuo, P.-C.; Lin, Z.-X.; Wu, T.-Y.; Hsu, C.-H.; Lin, H.-P.; Wu, T.-S. Effects of morphology and pore size of mesoporous silicas on the efficiency of an immobilized enzyme. RSC Adv. 2021, 11, 10010–10017. [Google Scholar] [CrossRef]
- Caldas, E.M.; Novatzky, D.; Deon, M.; de Menezes, E.W.; Hertz, P.F.; Costa, T.M.H.; Arenas, L.T.; Benvenutti, E.V. Pore size effect in the amount of immobilized enzyme for manufacturing carbon ceramic biosensor. Microporous Mesoporous Mater. 2017, 247, 95–102. [Google Scholar] [CrossRef]
- Califano, V.; Costantini, A.; Silvestri, B.; Venezia, V.; Cimino, S.; Sannino, F. The effect of pore morphology on the catalytic performance of β-glucosidase immobilized into mesoporous silica. Pure Appl. Chem. 2019, 91, 1583–1592. [Google Scholar] [CrossRef]
- Tadepalli, S.; Wang, Z.; Slocik, J.; Naik, R.R.; Singamaneni, S. Effect of size and curvature on the enzyme activity of bionanoconjugates. Nanoscale 2017, 9, 15666–15672. [Google Scholar] [CrossRef]
- Peng, G.; Hou, X.; Liu, B.; Chen, H.; Luo, R. Stabilized enzyme immobilization on micron-size PSt-GMA microspheres: Different methods to improve the carriers’ surface biocompatibility. RSC Adv. 2016, 6, 91431–91439. [Google Scholar] [CrossRef]
- Shinde, P.; Musameh, M.; Gao, Y.; Robinson, A.J.; Kyratzis, I. Immobilization and stabilization of alcohol dehydrogenase on polyvinyl alcohol fibre. Biotechnol. Rep. 2018, 19, e00260. [Google Scholar] [CrossRef] [PubMed]
- Abaházi, E.; Lestál, D.; Boros, Z.; Poppe, L. Tailoring the Spacer Arm for Covalent Immobilization of Candida antarctica Lipase B—Thermal Stabilization by Bisepoxide-Activated Aminoalkyl Resins in Continuous-Flow Reactors. Molecules 2016, 21, 767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Contente, M.L.; Farris, S.; Tamborini, L.; Molinari, F.; Paradisi, F. Flow-Based Enzymatic Synthesis of Melatonin and Other High Value Tryptamine Derivatives: A Five-Minute Intensified Process. Green Chem. 2019, 21, 3263–3266. [Google Scholar] [CrossRef] [Green Version]
- Romero, O.; Guisán, J.M.; Illanes, A.; Wilson, L. Reactivation of penicillin acylase biocatalysts: Effect of the intensity of enzyme–support attachment and enzyme load. J. Mol. Catal. B Enzym. 2012, 74, 224–229. [Google Scholar] [CrossRef]
- Arana-Peña, S.; Rios, N.S.; Carballares, D.; Mendez-Sanchez, C.; Lokha, Y.; Gonçalves, L.R.B.; Fernandez-Lafuente, R. Effects of Enzyme Loading and Immobilization Conditions on the Catalytic Features of Lipase From Pseudomonas fluorescens Immobilized on Octyl-Agarose Beads. Front. Bioeng. Biotechnol. 2020, 8, 36. [Google Scholar] [CrossRef] [Green Version]
- .García-García, P.; Fernandez-Lorente, G.; Guisan, J.M. Capture of enzyme aggregates by covalent immobilization on solid supports. Relevant stabilization of enzymes by aggregation. J. Biotechnol. 2021, 325, 138–144. [Google Scholar] [CrossRef]
- Li, Y.-M.; Yuan, J.; Ren, H.; Ji, C.-Y.; Tao, Y.; Wu, Y.; Chou, L.-Y.; Zhang, Y.-B.; Cheng, L. Fine-Tuning the Micro-Environment to Optimize the Catalytic Activity of Enzymes Immobilized in Multivariate Metal–Organic Frameworks. J. Am. Chem. Soc. 2021, 143, 15378–15390. [Google Scholar] [CrossRef]
- Ruiz, M.; Plata, E.; Castillo, J.J.; Ortiz, C.C.; López, G.; Baena, S.; Torre, R.; Fernandez-Lafuente, R. Modulation of the Biocatalytic Properties of a Novel Lipase from Psychrophilic Serratia sp. (USBA-GBX-513) by Different Immobilization Strategies. Molecules 2021, 26, 1574. [Google Scholar] [CrossRef]
- Fernandez-Lafuente, R. Stabilization of multimeric enzymes: Strategies to prevent subunit dissociation. Enzym. Microb. Technol. 2009, 45, 405–418. [Google Scholar] [CrossRef]
- García-García, P.; Guisan, J.M.; Fernandez-Lorente, G.A. Mild intensity of the enzyme-support multi-point attachment promotes the optimal stabilization of mesophilic multimeric enzymes: Amine oxidase from Pisum sativum. J. Biotechnol. 2020, 318, 39–44. [Google Scholar] [CrossRef]
- Zhu, L.; Wang, S.; Tian, W.; Zhang, Y.; Song, Y.; Zhang, J.; Mu, B.; Peng, C.; Deng, Z.; Ma, H.; et al. Stabilization of multimeric proteins via intersubunit cyclization. Appl. Environ. Microbiol. 2017, 83, e01239-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Q.; Li, A.; Zhang, Z.; Ma, R.; Shi, L. Stabilization of multimeric enzymes against heat inactivation by chitosan-graft-poly(N-isopropylacrylamide) in confined spaces. ACS Biomater. Sci. Eng. 2017, 3, 3141–3145. [Google Scholar] [CrossRef] [PubMed]
- Trobo-Maseda, L.; Orrego, A.H.; Moreno-Pérez, S.; Fernández-Lorente, G.; Guisan, J.M.; Rocha-Martin, J. Stabilization of multimeric sucrose synthase from Acidithiobacillus caldus via immobilization and post-immobilization techniques for synthesis of UDP-glucose. Appl. Microbiol. Biotechnol. 2018, 102, 773–787. [Google Scholar] [CrossRef] [PubMed]
- Mateo, C.; Pessela, B.; Fuentes, M.; Torres, R.; Betancor, L.; Hidalgo, A.; Fernandez-Lorente, G.; Fernandez-Lafuente, R.; Guisan, J.M. Stabilization of Multimeric Enzymes via Immobilization and Further Cross-Linking with Aldehyde-Dextran. In Immobilization of Enzymes and Cells. Methods in Molecular Biology; Guisan, J., Bolivar, J., López-Gallego, F., Rocha-Martín, J., Eds.; Humana: New York, NY, USA, 2020; Volume 2100, pp. 175–187. [Google Scholar]
- Rodrigues, R.C.; Berenguer-Murcia, Á.; Carballares, D.; Morellon-Sterling, R.; Fernandez-Lafuente, R. Stabilization of enzymes via immobilization: Multipoint covalent attachment and other stabilization strategies. Biotechnol. Adv. 2021, 52, 107821. [Google Scholar] [CrossRef] [PubMed]
- Cavalcante, F.T.T.; Cavalcante, A.L.G.; de Sousa, I.G.; Neto, F.S.; dos Santos, J.C.S. Current Status and Future Perspectives of Supports and Protocols for Enzyme Immobilization. Catalysts 2021, 11, 1222. [Google Scholar] [CrossRef]
- Liu, X.; Formanek, P.; Voit, B.; Appelhans, D. Functional Cellular Mimics for the Spatiotemporal Control of Multiple Enzymatic Cascade Reactions. Angew. Chem. Int. Ed. Engl. 2017, 56, 16233–16238. [Google Scholar] [CrossRef]
- Bernal, C.; Rodríguez, K.; Martínez, R. Integrating enzyme immobilization and protein engineering: An alternative path for the development of novel and improved industrial biocatalysts. Biotechnol Adv. 2018, 36, 1470–1480. [Google Scholar] [CrossRef]
- Zdarta, J.; Meyer, A.S.; Jesionowski, T.; Pinelo, M.A. General Overview of Support Materials for Enzyme Immobilization: Characteristics, Properties, Practical Utility. Catalysts 2018, 8, 92. [Google Scholar] [CrossRef] [Green Version]
- Cen, Y.-K.; Liu, Y.-X.; Xue, Y.-P.; Zheng, Y.-G. Immobilization of Enzymes in/on Membranes and their Applications. Adv. Synth. Catal. 2019, 361, 5500–5515. [Google Scholar] [CrossRef]
- Bilal, M.; Asgher, M.; Cheng, H.; Yan, Y.; Iqbal, H.M.N. Multi-point enzyme immobilization, surface chemistry, and novel platforms: A paradigm shift in biocatalyst design. Crit Rev. Biotechnol. 2019, 39, 202–219. [Google Scholar] [CrossRef]
- Ye, N.; Kou, X.; Shen, J.; Huang, S.; Chen, G.; Ouyang, G. Metal-Organic Frameworks: A New Platform for Enzyme Immobilization. ChemBioChem 2020, 21, 2585–2590. [Google Scholar] [CrossRef] [PubMed]
- Aggarwal, S.; Chakravarty, A.; Ikram, S. A comprehensive review on incredible renewable carriers as promising platforms for enzyme immobilization & thereof strategies. Int. J. Biol. Macromol. 2021, 167, 962–986. [Google Scholar] [PubMed]
- Ding, S.S.; Zhu, J.P.; Wang, Y.; Yu, Y.; Zhao, Z. Recent progress in magnetic nanoparticles and mesoporous materials for enzyme immobilization: An update. Braz. J. Biol. 2021, 82, e244496. [Google Scholar] [CrossRef]
- Liu, D.M.; Chen, J.; Shi, Y.P. Advances on methods and easy separated support materials for enzymes immobilization. Trends Analyt. Chem. 2018, 102, 332–342. [Google Scholar] [CrossRef]
- Rodríguez-Núñez, K.; Berna, C.; Martínez, R. Immobilized Biocatalyst Engineering: High throughput enzyme immobilization for the integration of biocatalyst improvement strategies. Biotechnol. Adv. 2021, 36, 1470–1480. [Google Scholar] [CrossRef] [PubMed]
- Gkantzou, E.; Chatzikonstantinou, A.V.; Fotiadou, R.; Giannakopoulou, A.; Patila, M.; Stamatis, H. Trends in the development of innovative nanobiocatalysts and their application in biocatalytic transformations. Biotechnol. Adv. 2021, 51, 107738. [Google Scholar] [CrossRef] [PubMed]
- Betancor, L.; Luckarift, H. Co-immobilized coupled enzyme systems in biotechnology. Biotechnol. Genetic Eng. Rev. 2010, 27, 95–114. [Google Scholar] [CrossRef] [Green Version]
- Gruber, P.; Marques, M.P.C.; O’Sullivan, B.; Baganz, F.; Wohlgemuth, R.; Szita, N. Conscious coupling: The challenges and opportunities of cascading enzymatic microreactors. Biotechnol. J. 2017, 12, 1700030. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; Li, Z.; Zheng, S.; Xu, H.; Zhou, Y.J.; Gao, Z.; Meng, C.; Li, S. Establishing an enzyme cascade for one-pot production of α-olefins from low-cost triglycerides and oils without exogenous H2O2 addition. Biotechnol. Biofuels 2020, 13, 52. [Google Scholar] [CrossRef]
- Metzner, R.; Hummel, W.; Wetterich, F.; König, B.; Gröger, H. Integrated biocatalysis in multistep drug synthesis without intermediate isolation: A de novo approach toward a rosuvastatin key building block. Org. Process. Res. Dev. 2015, 19, 635–638. [Google Scholar] [CrossRef]
- Rudroff, F.; Mihovilovic, M.D.; Gröger, H.; Snajdrova, R.; Iding, H.; Bornscheuer, U.T. Opportunities and challenges for combining chemo- and biocatalysis. Nat. Catal. 2018, 1, 12–22. [Google Scholar] [CrossRef]
- Mestrom, L.; Przypis, M.; Kowalczykiewicz, D.; Pollender, A.; Kumpf, A.; Marsden, S.R.; Bento, I.; Jarzębski, A.B.; Szymańska, K.; Chruściel, A.; et al. Leloir Glycosyltransferases in Applied Biocatalysis: A Multidisciplinary Approach. Int. J. Mol. Sci. 2019, 20, 5263. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Wen, L.; Tan, T.; Lv, Y. Sequential Co-immobilization of Enzymes in Metal-Organic Frameworks for Efficient Biocatalytic Conversion of Adsorbed CO2 to Formate. Front. Bioeng. Biotechnol. 2019, 7, 394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arana-Peña, S.; Carballares, D.; Morellon-Sterlling, R.; Berenguer-Murcia, Á.; Alcántara, A.R.; Rodrigues, R.C.; Fernandez-Lafuente, R. Enzyme co-immobilization: Always the biocatalyst designers’ choice…or not? Biotechnol. Adv. 2020, 51, 107584. [Google Scholar] [CrossRef] [PubMed]
- Ricca, E.; Brucher, B.; Schrittwieser, J.H. Multi-enzymatic cascade reactions: Overview and perspectives. Adv. Synth. Catal. 2011, 353, 2239–2262. [Google Scholar] [CrossRef]
- Muschiol, J.; Peters, C.; Oberleitner, N.; Mihovilovic, M.D.; Bornscheuer, U.T.; Rudroff, F. Cascade catalysis—Strategies and challenges en route to preparative synthetic biology. Chem Commun. 2015, 51, 5798–5811. [Google Scholar] [CrossRef]
- Robescu, M.S.; Serra, I.; Terreni, M.; Ubiali, D.; Bavaro, T. A Multi-Enzymatic Cascade Reaction for the Synthesis of Vidarabine 5′-Monophosphate. Catalysts 2020, 10, 60. [Google Scholar] [CrossRef] [Green Version]
- Velasco-Lozano, S.; Santiago-Arcos, J.; Mayoral, J.A.; López-Gallego, F. Co-immobilization and colocalization of multi-enzyme systems for the cell-free biosynthesis of aminoalcohols. ChemCatChem 2020, 12, 3030–3041. [Google Scholar] [CrossRef]
- Kazenwadel, F.; Franzreb, M.; Rapp, B.E. Synthetic enzyme supercomplexes: Co-immobilization of enzyme cascades. Anal. Methods 2015, 7, 4030–4037. [Google Scholar] [CrossRef] [Green Version]
- Quin, M.B.; Wallin, K.K.; Zhang, G.; Schmidt-Dannert, C. Spatial organization of multi-enzyme biocatalytic cascades. Org. Biomol. Chem. 2017, 15, 4260–4271. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.T.; Lee, S. Multienzymatic cascade reactions via enzyme complex by immobilization. ACS Catal. 2019, 9, 4402–4425. [Google Scholar] [CrossRef]
- Giannakopoulou, A.; Gkantzou, E.; Polydera, A.; Stamatis, H. Multienzymatic Nanoassemblies: Recent Progress and Applications. Trends Biotechnol. 2020, 38, 202–216. [Google Scholar] [CrossRef]
- Xu, K.; Chen, X.; Zheng, R.; Zheng, Y. Immobilization of Multi-Enzymes on Support Materials for Efficient Biocatalysis. Front. Bioeng. Biotechnol. 2020, 8, 660. [Google Scholar] [CrossRef] [PubMed]
- Júnior, A.A.D.T.; Ladeira, Y.F.X.; França, A.D.S.; Souza, R.O.M.A.D.; Moraes, A.H.; Wojcieszak, R.; Itabaiana, I., Jr.; Miranda, A.S.D. Multicatalytic Hybrid Materials for Biocatalytic and Chemoenzymatic Cascades—Strategies for Multicatalyst (Enzyme) Co-Immobilization. Catalysts 2021, 11, 936. [Google Scholar] [CrossRef]
- Cutlan, R.; De Rose, S.; Isupov, M.N.; Littlechild, J.A.; Harmer, N.J. Using enzyme cascades in biocatalysis: Highlight on transaminases and carboxylic acid reductases. Biochim. Biophys. Acta Proteins Proteom. 2020, 1868, 140322. [Google Scholar] [CrossRef]
- McIntosh, J.A.; Owens, A. Enzyme engineering for biosynthetic cascades. Curr. Opin. Green Sustain. Chem. 2021, 29, 100448. [Google Scholar] [CrossRef]
- Schwizer, F.; Okamoto, Y.; Heinisch, T.; Gu, Y.; Pellizzoni, M.M.; Lebrun, V.; Reuter, R.; Köhler, V.; Lewis, J.C.; Ward, T.R. Artificial Metalloenzymes: Reaction Scope and Optimization Strategies. Chem. Rev. 2018, 118, 142–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomo, J.M. Artificial enzymes with multiple active sites. Curr. Opin. Green Sustain. Chem. 2021, 29, 100452. [Google Scholar] [CrossRef]
- Küchler, A.; Yoshimoto, M.; Luginbühl, S.; Mavelli, F.; Walde, P. Enzymatic reactions in confined environments. Nat. Nanotech. 2016, 11, 409–420. [Google Scholar] [CrossRef]
- Shi, J.; Wu, Y.; Zhang, S.; Tian, Y.; Yang, D.; Jiang, Z. Bioinspired construction of multi-enzyme catalytic systems. Chem. Soc. Rev. 2018, 47, 4295–4313. [Google Scholar] [CrossRef]
- Lechner, H.; Ferruz, N.; Höcker, B. Strategies for designing non-natural enzymes and binders. Curr. Opin. Chem. Biol. 2018, 47, 67–76. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Cao, Y.; Luo, K.; Sun, Y.Z.; Xiong, J.R.; Wang, L.C.; Liu, Z.; Li, J.; Ma, J.Y.; Ge, J.; et al. Highly active enzyme–metal nanohybrids synthesized in protein–polymer conjugates. Nat. Catal. 2019, 2, 718–725. [Google Scholar] [CrossRef]
- Ren, S.; Wang, Z.; Bilal, M.; Feng, Y.; Jiang, Y.; Jia, S.; Cui, J. Co-immobilization multienzyme nanoreactor with co-factor regeneration for conversion of CO2. Int. J. Biol. Macromol. 2020, 155, 110–118. [Google Scholar] [CrossRef] [PubMed]
- Orrego, A.H.; López-Gallego, F.; Fernandez-Lorente, G.; Guisan, J.M.; Rocha-Martín, J. Co-Immobilization and Co-Localization of Multi-Enzyme Systems on Porous Materials. In Immobilization of Enzymes and Cells. Methods in Molecular Biology; Guisan, J., Bolivar, J., López-Gallego, F., Rocha-Martín, J., Eds.; Humana: New York, NY, USA, 2020; Volume 2100, pp. 297–308. [Google Scholar]
- Schoffelen, S.; van Hest, J.C. Chemical approaches for the construction of multi-enzyme reaction systems. Curr. Opin. Struct. Biol. 2013, 23, 613–621. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, D.; Wu, C.; Yao, K.; Li, Z.; Shi, N.; Wen, F.; Gates, I.D. Co-immobilization of cellulase and lysozyme on amino-functionalized magnetic nanoparticles: An activity-tunable biocatalyst for extraction of lipids from microalgae. Bioresour. Technol. 2018, 263, 317–324. [Google Scholar] [CrossRef]
- Vong, T.; Schoffelen, S.; van Dongen, S.F.M.; van Beek, T.A.; Zuilhof, H.; van Hest, J.C.M. A DNA-Based Strategy for Dynamic Positional Enzyme Immobilization Inside Fused Silica Microchannels. Chem. Sci. 2011, 2, 1278–1285. [Google Scholar] [CrossRef]
- Boehm, C.R.; Freemont, P.S.; Ces, O. Design of a prototype flow microreactor for synthetic biology in vitro. Lab. Chip 2013, 13, 3426–3432. [Google Scholar] [CrossRef]
- Chen, Y.; Ke, G.; Ma, Y.; Zhu, Z.; Liu, M.; Liu, Y.; Yan, H.; Yang, C.J. A Synthetic Light-Driven Substrate Channeling System for Precise Regulation of Enzyme Cascade Activity Based on DNA Origami. J. Am. Chem. Soc. 2018, 140, 8990–8996. [Google Scholar] [CrossRef] [PubMed]
- Klein, W.P.; Thomsen, R.P.; Turner, K.B.; Walper, S.A.; Vranish, J.; Kjems, J.; Ancona, M.G.; Medintz, I.L. Enhanced catalysis from multienzyme cascades assembled on a DNA origami triangle. ACS Nano 2019, 13, 13677–13689. [Google Scholar] [CrossRef]
- Zhang, L.; Shi, J.F.; Jiang, Z.Y.; Jiang, Y.J.; Qiao, S.Z.; Li, J.A.; Wang, R.; Meng, R.; Zhu, Y.; Zheng, Y. Bioinspired preparation of polydopamine microcapsule for multienzyme system construction. Green Chem. 2011, 13, 300–306. [Google Scholar] [CrossRef]
- Begum, G.; Goodwin, W.B.; deGlee, B.M.; Sandhage, K.H.; Kröger, N. Compartmentalisation of enzymes for cascade reactions through biomimetic layer-by-layer mineralization. J. Mat. Chem. B 2015, 3, 5232–5240. [Google Scholar] [CrossRef] [Green Version]
- Bilal, M.; Hussain, N.; Américo-Pinheiro, J.H.P.; Almulaiky, Y.Q.; Iqbal, H.M. Multi-enzyme co-immobilized nano-assemblies: Bringing enzymes together for expanding bio-catalysis scope to meet biotechnological challenges. Int. J. Biol. Macromol. 2021, 186, 735–749. [Google Scholar] [CrossRef] [PubMed]
- Omidvar, M.; Zdarta, J.; Sigurdardóttir, S.B.; Pinelo, M. Mimicking natural strategies to create multi-environment enzymatic reactors: From natural cell compartments to artificial polyelectrolyte reactors. Biotechnol. Adv. 2022, 54, 107798. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Chen, Q.-S.; Zong, M.-H.; Lou, W.-Y. Sequential co-immobilization of multienzyme nanodevices based on SpyCatcher and SpyTag for robust biocatalysis. Mol. Catal. 2021, 510, 111673. [Google Scholar] [CrossRef]
- Ren, S.; Li, C.; Jiao, X.; Jia, S.; Jiang, Y.; Bilal, M.; Cui, J. Recent progress in multienzymes co-immobilization and multienzyme system applications. Chem. Eng. J. 2019, 373, 1254–1278. [Google Scholar] [CrossRef]
- Zhang, H.; Bai, Y.; Zhu, N.; Xu, J. Microfluidic reactor with immobilized enzyme-from construction to applications: A review. Chin. J. Chem. Eng. 2021, 30, 136–145. [Google Scholar] [CrossRef]
- Jia, F.; Narasimhan, B.; Mallapragada, S. Materials-based strategies for multi-enzyme immobilization and co-localization: A review. Biotechnol. Bioeng. 2014, 111, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Remonatto, D.; Ferrari, B.R.; Bassan, J.C.; Mussagy, C.U.; de Carvalho Santos-Ebinuma, V.; Veloso de Paula, A. Utilization of Clay Materials as Support for Aspergillus japonicus Lipase: An Eco-Friendly Approach. Catalysts 2021, 11, 1173. [Google Scholar] [CrossRef]
- Lyubyakina, P.; Tambasova, D.; Antonov, D.; Kovaleva, E. Complex enzymatic preparations immobilized on aluminum oxide in chitosan breakdown. AIP Conf. Proc. 2020, 2280, 050031. [Google Scholar] [CrossRef]
- Nishida, H.; Kajisa, T.; Miyazawa, Y.; Tabuse, Y.; Yoda, T.; Takeyama, H.; Kambara, H.; Sakata, T. Self-oriented immobilization of DNA polymerase tagged by titanium-binding peptide motif. Langmuir 2015, 31, 732–740. [Google Scholar] [CrossRef] [PubMed]
- Masuda, Y.; Kugimiya, S.-I.; Kato, K. Improvement of thermal-stability of enzyme immobilized onto mesoporous zirconia. J. Asian Ceram. Soc. 2014, 2, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Bôa Morte, E.F.; Marum, D.S.; Saitovitch, E.B.; Alzamora, M.; Monteiro, S.N.; Sanchez Rodriguez, R.J. Modified magnetite nanoparticle as biocatalytic support for magnetically stabilized fluidized bed reactors. J. Mater. Res. Technol. 2021, 14, 1112–1125. [Google Scholar] [CrossRef]
- Moisa, M.E.; Spelmezan, C.G.; Paul, C.; Bartha-Vari, J.H.; Bencze, L.C.; Irimie, F.D.; Paizs, C.; Peter, F.; Tosa, M.I. Tailored sol–gel immobilized lipase preparates for the enzymatic kinetic resolution of heteroaromatic alcohols in batch and continuous flow systems. RSC Adv. 2017, 7, 52977–52987. [Google Scholar] [CrossRef] [Green Version]
- Anastasescu, C.; Anastasescu, M.; Balint, I.; Zaharescu, M. SiO2 Based Materials for Immobilization of Enzymes (2019). In Nanomaterials—Toxicity, Human Health and Environment; Clichici, S., Filip, A., Nascimento, G.M., Eds.; IntechOpen: London, UK, 2019; pp. 1–21. [Google Scholar] [CrossRef] [Green Version]
- Hartmann, M.; Kostrov, X. Immobilization of enzymes on porous silicas—Benefits and challenges. Chem. Soc. Rev. 2013, 42, 6277–6289. [Google Scholar] [CrossRef] [PubMed]
- Pazarlioglu, N.K.; Sariisçik, M.; Telefoncu, A. Treating denim fabrics with immobilized commercial cellulases. Process. Biochem. 2005, 40, 767–771. [Google Scholar] [CrossRef]
- Sattar, H.; Aman, A.; Qader, S. Agar-agar immobilization: An alternative approach for the entrapment of protease to improve the catalytic efficiency, thermal stability and recycling efficiency. Int. J. Biol. Macromol. 2018, 111, 917–922. [Google Scholar] [CrossRef] [PubMed]
- Zucca, P.; Fernandez-Lafuente, R.; Sanjust, E. Agarose and Its Derivatives as Supports for Enzyme Immobilization. Molecules 2016, 21, 1577. [Google Scholar] [CrossRef]
- Kurtinaĭtene, B.S.; Kulis, I. Stabilization of native and immobilized urease. Priklad. Biokhim. Mikrobiol. 1987, 23, 299–302. [Google Scholar]
- Yoon, J.; Lee, S.H.; Tieves, F.; Rauch, M.; Hollmann, F.; Park, C.B. Light-harvesting dye–alginate hydrogel for solar-driven, sustainable biocatalysis of asymmetric hydrogenation. ACS Sustain. Chem. Eng. 2019, 7, 5632–5637. [Google Scholar] [CrossRef] [Green Version]
- Elnashar, M.M.; Awad, G.E.; Hassan, M.E.; Eldin, M.S.M.; Haroun, B.M.; El-Diwany, A.L. Optimal Immobilization of β-Galactosidase onto κ-Carrageenan Gel Beads Using Response Surface Methodology and Its Applications. Sci. World J. 2014, 2014, 571682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, M.L.; Kumar, S.; Das, A.; Randhawa, J.S.; Chamundeeswari, M. Enzyme Immobilization on Chitin and Chitosan-Based Supports for Biotechnological Applications. In Sustainable Agriculture Reviews 35; Crini, G., Lichtfouse, E., Eds.; Springer: Cah, Switzerland, 2019; pp. 147–173. [Google Scholar] [CrossRef]
- He, S.; Song, D.; Chen, M.; Cheng, H. Immobilization of Lipases on Magnetic Collagen Fibers and Its Applications for Short-Chain Ester Synthesis. Catalysts 2017, 7, 178. [Google Scholar] [CrossRef] [Green Version]
- Labus, K.; Wolanin, K.; Radosiński, Ł. Comparative Study on Enzyme Immobilization Using Natural Hydrogel Matrices—Experimental Studies Supported by Molecular Models Analysis. Catalysts 2020, 10, 489. [Google Scholar] [CrossRef]
- Jawaheer, S.; White, S.F.; Rughooputh, S.D.D.V.; Cullen, D.C. Enzyme stabilization using pectin as a novel entrapment matrix in biosensors. Anal. Lett. 2002, 35, 2077–2091. [Google Scholar] [CrossRef]
- Lee, J.-Y.; Jeon, S.J. Characterization and immobilization on nickel-chelated Sepharose of a glutamate decarboxylase A from Lactobacillus brevis BH2 and its application for production of GABA. Biosci. Biotechnol. Biochem. 2014, 78, 1656–1661. [Google Scholar] [CrossRef] [PubMed]
- Bedzo, O.; Trollope, K.; Gottumukkala, L.D.; Coetzee, G.; Görgens, J.F. Amberlite IRA 900 versus calcium alginate in immobilization of a novel, engineered β-fructofuranosidase for short-chain fructooligosaccharide synthesis from sucrose. Biotechnol. Prog. 2019, 35, e2797. [Google Scholar] [CrossRef] [PubMed]
- Rehman, H.U.; Nawaz, M.A.; Pervez, S.; Jamal, M.; Attaullah, M.; Aman, A.; Ul Qader, S.A. Encapsulation of pectinase within polyacrylamide gel: Characterization of its catalytic properties for continuous industrial uses. Heliyon 2020, 6, e04578. [Google Scholar] [CrossRef] [PubMed]
- Dencheva, N.; Braz, J.; Scheibel, D.; Malfois, M.; Denchev, Z.; Gitsov, I. Polymer-Assisted Biocatalysis: Polyamide 4 Microparticles as Promising Carriers of Enzymatic Function. Catalysts 2020, 10, 767. [Google Scholar] [CrossRef]
- Ghosh, S.; Chaganti, S.R.; Prakasham, R.S. Polyaniline nanofiber as a novel immobilization matrix for the anti-leukemia enzyme L-asparaginase. J. Mol. Catal. B Enzym. 2012, 74, 132–137. [Google Scholar] [CrossRef]
- Kumar, N.; Upadhyay, L.S.B. Enzyme Immobilization over Polystyrene Surface Using Cysteine Functionalized Copper Nanoparticle as a Linker Molecule. Appl. Biochem. Biotechnol. 2020, 191, 1247–1257. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Li, L.; Li, M. Improvement of lipase activity by synergistic immobilization on polyurethane and its application for large-scale synthesizing vitamin A palmitate. Prep. Biochem. Biotechnol. 2019, 49, 485–492. [Google Scholar] [CrossRef]
- Alonso-González, M.; Corral-González, A.; Felix, M.; Romero, A.; Martin-Alfonso, J.E. Developing active poly(vinyl alcohol)-based membranes with encapsulated antimicrobial enzymes via electrospinning for food packaging. Int. J. Biol. Macromol. 2020, 162, 913–921. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Fang, Z.; Duan, H.; Liang, L. Polydopamine-mediated synthesis of core–shell gold@calcium phosphate nanoparticles for enzyme immobilization. Biomater. Sci. 2019, 7, 2841–2849. [Google Scholar] [CrossRef] [PubMed]
- Shao, P.; He, Z.; Hu, Y.; Shen, Y.; Zhang, S.; Yu, Y. Zeolitic imidazolate frameworks with different organic ligands as carriers for Carbonic anhydrase immobilization to promote the absorption of CO2 into tertiary amine solution. Chem. Eng. J. 2022, 435, 134957. [Google Scholar] [CrossRef]
- Du, K.; Sun, J.; Zhou, X.; Feng, W.; Jiang, X.; Ji, P. A two-enzyme immobilization approach using carbon nanotubes/silica as support. Biotechnol. Prog. 2015, 31, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Laranjo, M.T.; Morawski, F.M.; Dias, S.L.P.; Benvenutti, E.V.; Arenas, L.T.; Costa, T.M.H. Silica/Titania Graphite Composite Modified with Chitosan and Tyrosinase Employed as a Sensitive Biosensor for Phenolic Compounds. J. Braz. Chem. Soc. 2019, 30, 2660–2672. [Google Scholar] [CrossRef]
- Convery, N.; Gadegaard, N. 30 Years of Microfluidics. Micro Nano Eng. 2019, 2, 76–91. [Google Scholar] [CrossRef]
- Benítez-Mateos, A.I.; Contente, M.L.; Roura Padrosa, D.; Paradisi, F. Flow biocatalysis: Design, development and applications. React. Chem. Eng. 2021, 6, 599–611. [Google Scholar] [CrossRef]
- Ward, K.; Fan, Z.H. Mixing in microfluidic devices and enhancement methods. J. Micromech. Microeng. 2015, 25, 094001. [Google Scholar] [CrossRef]
- Elvira, K.; Solvas, X.; Wootton, R.; deMello, A.J. The past, present and potential for microfluidic reactor technology in chemical synthesis. Nat. Chem. 2013, 5, 905–915. [Google Scholar] [CrossRef]
- Šalić, A.; Zelić, B. Synergy of Microtechnology and Biotechnology: Microreactors as an Effective Tool for Biotransformation Processes. Food Technol. Biotechnol. 2018, 56, 464–479. [Google Scholar] [CrossRef] [PubMed]
- Prado, R.C.; Borges, E.R. Microbioreactors as Engineering Tools for Bioprocess Development. Braz. J. Chem. Eng. 2018, 35, 1163–1182. [Google Scholar] [CrossRef] [Green Version]
- Calleri, E.; Temporini, C.; Colombo, R.; Tengattini, S.; Rinaldi, F.; Brusotti, G.; Furlanetto, S.; Massolini, G. Analytical settings for in-flow biocatalytic reaction monitoring, TrAC. Trends Anal. Chem. 2021, 143, 116348. [Google Scholar] [CrossRef]
- Gkantzou, E.; Patila, M.; Stamatis, H. Magnetic Microreactors with Immobilized Enzymes—From Assemblage to Contemporary Applications. Catalysts 2018, 8, 282. [Google Scholar] [CrossRef] [Green Version]
- Žnidaršič-Plazl, P. The Promises and the Challenges of Biotransformations in Microflow. Biotechnol. J. 2019, 14, 1800580. [Google Scholar] [CrossRef] [PubMed]
- De Santis, P.; Meyer, L.-E.; Kara, S. The rise of continuous flow biocatalysis–Fundamentals, very recent developments and future perspectives. React. Chem. Eng. 2020, 5, 2155–2184. [Google Scholar] [CrossRef]
- Zhu, Y.; Chen, Q.; Shao, L.; Jia, Y.; Zhang, X. Microfluidic immobilized enzyme reactors for continuous biocatalysis. React. Chem. Eng. 2020, 5, 9–32. [Google Scholar] [CrossRef]
- Santi, M.; Sancineto, L.; Nascimento, V.; Braun Azeredo, J.; Orozco, E.V.M.; Andrade, L.H.; Gröger, H.; Santi, C. Flow Biocatalysis: A Challenging Alternative for the Synthesis of APIs and Natural Compounds. Int. J. Mol. Sci. 2021, 22, 990. [Google Scholar] [CrossRef] [PubMed]
- Amin, F.; Haq, N.B.; Muhammad, B. Recent advances in the production strategies of microbial pectinases—A review. Biol. Macromol. 2019, 122, 1017–1026. [Google Scholar] [CrossRef]
- Huang, D.; Yuyang, S.; Yanlin, L.; Yi, Q. A new strain of Aspergillus tubingensis for high-activity pectinase production. Braz. J. Microbiol. 2019, 50, 53–65. [Google Scholar] [CrossRef]
- Satapathy, S.; Rout, J.R.; Kerry, R.G.; Thatoi, H.; Sahoo, S.L. Biochemical Prospects of Various Microbial Pectinase and Pectin: An Approachable Concept in Pharmaceutical Bioprocessing. Front. Nutr. 2020, 7, 117. [Google Scholar] [CrossRef]
- Khan, M.; Ekambaram, N.; Umesh-Kumar, S. Potential Application of Pectinase in Developing Functional Foods. Annu. Rev. Food Sci. Technol. 2013, 4, 21–34. [Google Scholar] [CrossRef] [PubMed]
- Alimardani-Theuil, P.; Gainvors-Claisse, A.; Duchiron, F. Yeasts: An attractive source of pectinases—From gene expression to potential applications: A review. Process. Biochem. 2011, 46, 1525–1537. [Google Scholar] [CrossRef]
- de Oliveira, R.L.; Osmar, S.S.; Attilio, C.; Tatiana, S.P. Thermodynamic and kinetic studies on pectinase extracted from Aspergillus aculeatus: Free and immobilized enzyme entrapped in alginate beads. Int. J. Biol. Macromol. 2018, 115, 1088–1093. [Google Scholar] [CrossRef] [PubMed]
- Takc, H.A.; Mercimek, U.T. Extracellular pectinase production and purification from a newly isolated Bacillus subtilis strain. Int. J. Food Prop. 2016, 19, 2443–2450. [Google Scholar]
- Voragen, A.G.J.; Gerd-Jan, C.; Rene, P.V.; Henk, A.S.P. Pectin a versatile polysaccharide present in plant cell walls. Struct. Chem. 2009, 20, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Favela, T.; Volke-Sepúlveda, T.; Viniegra-González, G. Production of Hydrolytic Depolymerising Pectinases. Food Technol. Biotechnol. 2006, 44, 221–227. [Google Scholar]
- Gonçalves, D.B.; Teixeira, J.A.; Bazzolli, D.M.S.; Queiroz, M.V.D.; Araújo, E.F. Use of response surface methodology to optimize production of pectinases by recombinant Penicillium griseoroseum T20. Biocat. Agric. Biotechnol. 2012, 1, 140–146. [Google Scholar] [CrossRef]
- Govindaraji, P.K.; Vuppu, S. Characterization of pectin and optimization of pectinase enzyme from novel Streptomyces fumigatiscleroticus VIT-SP4 for drug delivery and concrete crack-healing applications: An eco-friendly approach. Saudi J. Biol. Sci. 2020, 20, 3529–3540. [Google Scholar] [CrossRef]
- Ortiz, G.E.; María, C.; Ponce, M.; Diego, G.N.; Gabriela, C.; Celina, S.; María, C.L.; Guillermo, P.G.; Martín, B.; Edgardo, O.A. Pectinase production by Aspergillus giganteus in solid state fermentation: Optimization, scale-up, biochemical characterization and its application in olive oil extraction. J. Ind. Microbiol. Technol. 2017, 44, 197–211. [Google Scholar] [CrossRef] [PubMed]
- Angelim, A.L.; Samantha, P.C.; Cibelle, B.F.L.; Freitas, A.; Vânia, M.M. An innovative bioremediation strategy using a bacterial consortium entrapped in chitosan beads. J. Environ. Manag. 2013, 127, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Di Carlo, D. Inertial Microfluidics. Lab. Chip 2009, 9, 3038–3046. [Google Scholar] [CrossRef]
- Dai, X.Y.; Kong, L.M.; Wang, X.L.; Zhu, Q.; Chen, K.; Zhou, T. Preparation, characterization and catalytic behavior of pectinase covalently immobilized onto sodium alginate/graphene oxide composite beads. Food Chem. 2018, 253, 185–193. [Google Scholar] [CrossRef]
- Dal Magro, L.; Silveira, V.C.C.; de Menezes, E.W.; Benvenutti, E.V.; Nicolodi, S.; Hertz, P.F.; Klein, M.P.; Rodrigues, R.C. Magnetic biocatalysts of pectinase and cellulase: Synthesis and characterization of two preparations for application in grape juice clarification. Int. J. Biol. Macromol. 2018, 115, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Djordjevic, V.; Willaert, R.; Gibson, B.; Nedovic, V. Immobilized yeast cells and secondary metabolites. In Fungal Metabolites; Reference Series in Phytochemistry; Mérillon, J.M., Ramawat, K., Eds.; Springer: Cham, Switzerland, 2016; pp. 599–638. [Google Scholar]
- Sojitra, U.V.; Nadar, S.S.; Rathod, V.K. A magnetic tri-enzyme nanobiocatalyst for fruit juice clarification. Food Chem. 2016, 213, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Muley, A.B.; Thorat, A.S.; Singhal, R.S.; Harinath Babu, K. A tri-enzyme coimmobilized magnetic complex: Process details, kinetics, thermodynamics and applications. Int. J. Biol. Macromol. 2018, 118, 1781–1795. [Google Scholar] [CrossRef]
- Nadar, S.S.; Rathod, V.K. A co-immobilization of pectinase and cellulase onto magnetic nanoparticles for antioxidant extraction from waste fruit peels. Biocatal. Agric. Biotechnol. 2019, 17, 470–479. [Google Scholar] [CrossRef]
- Voberkov, S.; Solcany, V.; Vrsansk, M.; Adam, V. Immobilization of ligninolytic enzymes from white rot fungi in cross-linked aggregates. Chemosphere 2020, 18, 694–707. [Google Scholar] [CrossRef]
- Trappey, A.F.; Johnson, C.E.; Wilson, P.W. Use of a Commercial Pectolytic Enzyme to Extract Juice from Frozen Mayhaw (Crataegus opaca Hook.) Fruit. Int. J. Fruit Sci. 2007, 7, 77–86. [Google Scholar] [CrossRef]
- Cho, E.; Jung, S.; Kim, H.; Lee, H.Y.; Nam, K.; Lee, H.; Bae, H. Co-immobilization of three cellulases on Au-doped magnetic silica nanoparticles for the degradation of cellulose. Chem. Commun. 2012, 48, 886–888. [Google Scholar] [CrossRef] [PubMed]
- Nadar, S.S.; Rathod, V.K. Magnetic nanobiocatalyst for extraction of bioactive ingredients: A novel approach. Trends Food Sci. Technol. 2020, 103, 225–238. [Google Scholar] [CrossRef]
- Demir, N.; Jale, A.; Kemal, S.; Mehmet, M. The use of commercial pectinase in fruit juice industry, Part 3: Immobilized pectinase for mash treatment. J. Food Eng. 2001, 47, 275–280. [Google Scholar] [CrossRef]
- Spagna, G.; Romagnoli, D.; Martino, A.; Bianchi, G.; Pier, B.; Pifferi, G. A simple method for purifying glycosidases: α-l-rhamnopyranosidase from Aspergillus niger to increase the aroma of Moscato wine Giovanni Spagna. Enzym. Microb. Technol. 2000, 27, 522–530. [Google Scholar] [CrossRef]
- Puri, M. Updates on naringinase: Structural and biotechnological aspects. Appl. Microbiol. Biotechnol. 2012, 93, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, M.H.L. Naringinases: Occurrence, characteristics, and applications. Appl. Microbiol. Biotechnol. 2011, 90, 1883–1895. [Google Scholar] [CrossRef]
- Ferreira, L.; Afonso, C.; Vila-Real, H.; Alfaia, A.; Ribeiro, M.H.L. Evaluation of the Effect of High Pressure on Naringin Hydrolysis in Grapefruit Juice with Naringinase Immobilised in Calcium Alginate Beads. Food Technol. Biotechnol. 2008, 46, 146–150. [Google Scholar]
- Ribeiro, M.H.L.; Rabaça, M. Cross-linked enzyme aggregates of naringinase: Novel biocatalysts for naringin hydrolysis. Enzyme Res. 2011, 8, 851272. [Google Scholar] [CrossRef] [Green Version]
- Martins, S.L.; Albuquerque, B.F.; Nunes, M.A.P.; Ribeiro, M.H.L. Exploring magnetic and imprinted cross-linked enzyme aggregates of rhamnopyranosidase in microbioreactors. Bioresour. Technol. 2017, 247, 704–712. [Google Scholar] [CrossRef]
- Nunes, M.A.P.; Fernandes, P.C.B.; Ribeiro, M.H.L. High affinity water-soluble system for efficient naringinase immobilization in polyvinyl alcohol-dimethyl sulfoxide lens-shaped particles. J. Mol. Recognit. 2012, 25, 580–594. [Google Scholar] [CrossRef]
- Onbas, R.; Ozlem, Y.C. Synthesis of alginate-silica hybrid hydrogel for biocatalytic conversion by β-glucosidase in microreactor. Eng. Life Sci. 2018, 19, 37–46. [Google Scholar] [CrossRef]
- Berbegal, C.; Spano, G.; Tristezza, M.; Griego, F.; Capozzi, V. Microbial resources and innovation in the wine production sector. S. Afr. J. Enol. Vitic. 2017, 38, 156–166. [Google Scholar] [CrossRef]
- Park, M.; Sun, Q.; Liu, F.; DeLisa, M.P.; Chen, W. Positional assembly of enzymes on bacterial outer membrane vesicles for cascade reactions. PLoS ONE 2014, 9, e97103. [Google Scholar] [CrossRef] [Green Version]


| Immobilization Method | Advantages | Limitations | References |
|---|---|---|---|
| Adsorption: involves weak interactions between enzyme and carrier, e.g., van der Waals intermolecular forces, hydrophobic interactions | Simple, low-cost preparation No modification of the enzyme required, active site typically not affected, maximizes activity retention. Carrier regeneration most feasible | Weak binding force, most prone to enzyme leakage (most sensitive to shifts in temperature and pH, and to strong hydrodynamic forces and concomitant shear stress). Random, non-specific immobilization. Operational stability relatively poor due to enzyme leakage | [66,84,85,86,87] |
| Ionic interaction: involves electrostatic interactions between oppositely charged residues in carrier and enzyme | Simple, low-cost preparation Active site typically hardly affected, high activity retention. Intermediate binding force. Carrier regeneration possible | Less prone to enzyme leakage than adsorption Intermediate operational stability | [46,88,89,90] |
| Affinity binding: involves specific interactions between complementary molecules, e.g., poly-histidine tagged enzyme-metal ion activated carrier | Oriented, well-controlled immobilization Highly specific Carrier regeneration possible | Cost and complexity Specific groups are required in the enzyme surface, complementary binding agent needed | [44,68,69] |
| Covalent binding: involves strong chemical bonds established between carrier and enzyme residues, e.g., through Schiff bases | Strong binding High stability Most feasible for long term use | Carrier regeneration largely unfeasible. Complexity and cost. Risk of activity loss during immobilization | [36,91,92,93,94] |
| Entrapment/encapsulation: involves physical retainment of the enzyme within a polymeric network (entrapment in e.g., hydrogel) or enzyme containment inside a semipermeable barrier (encapsulation in e.g., a membrane) | No chemical interaction between enzyme and carrier, enzyme activity site unhindered Moderate cost and complexity Multienzyme immobilization easy | Diffusion limitations Prone to enzyme leakage, particularly low molecular wight enzymes | [79,95,96,97,98,99] |
| CLEAs: carrier-free method, involves chemical crosslinking of enzyme molecules and co-aggregates such as bovine serum albumin, using bifunctional compounds, e.g., glutaraldehyde | Strong binding Relatively simple preparation Biocatalyst devoided of non-catalytic material Stabilization of multimers | Activity loss during immobilization Small pore size may result in poor diffusion of high molecular weight substrates. Compression and cluster/clump formation during recovery by filtration/centrifugation, further hindering diffusion | [25,100,101,102,103,104] |
| Magnetic-CLEAs: the same as CLEAs but magnetic particles are also crosslinked | The same as for CLEAs plus eased recovery of the biocatalyst using a magnetic field | Same as for CLEAs but cluster/clump formation avoided since recovery circumvents filtration/centrifugation | [25,105,106,107,108] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bié, J.; Sepodes, B.; Fernandes, P.C.B.; Ribeiro, M.H.L. Enzyme Immobilization and Co-Immobilization: Main Framework, Advances and Some Applications. Processes 2022, 10, 494. https://doi.org/10.3390/pr10030494
Bié J, Sepodes B, Fernandes PCB, Ribeiro MHL. Enzyme Immobilization and Co-Immobilization: Main Framework, Advances and Some Applications. Processes. 2022; 10(3):494. https://doi.org/10.3390/pr10030494
Chicago/Turabian StyleBié, Joaquim, Bruno Sepodes, Pedro C. B. Fernandes, and Maria H. L. Ribeiro. 2022. "Enzyme Immobilization and Co-Immobilization: Main Framework, Advances and Some Applications" Processes 10, no. 3: 494. https://doi.org/10.3390/pr10030494
APA StyleBié, J., Sepodes, B., Fernandes, P. C. B., & Ribeiro, M. H. L. (2022). Enzyme Immobilization and Co-Immobilization: Main Framework, Advances and Some Applications. Processes, 10(3), 494. https://doi.org/10.3390/pr10030494