
Analysis of the Fungi Community Variation during Rice Storage through High Throughput Sequencing


 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
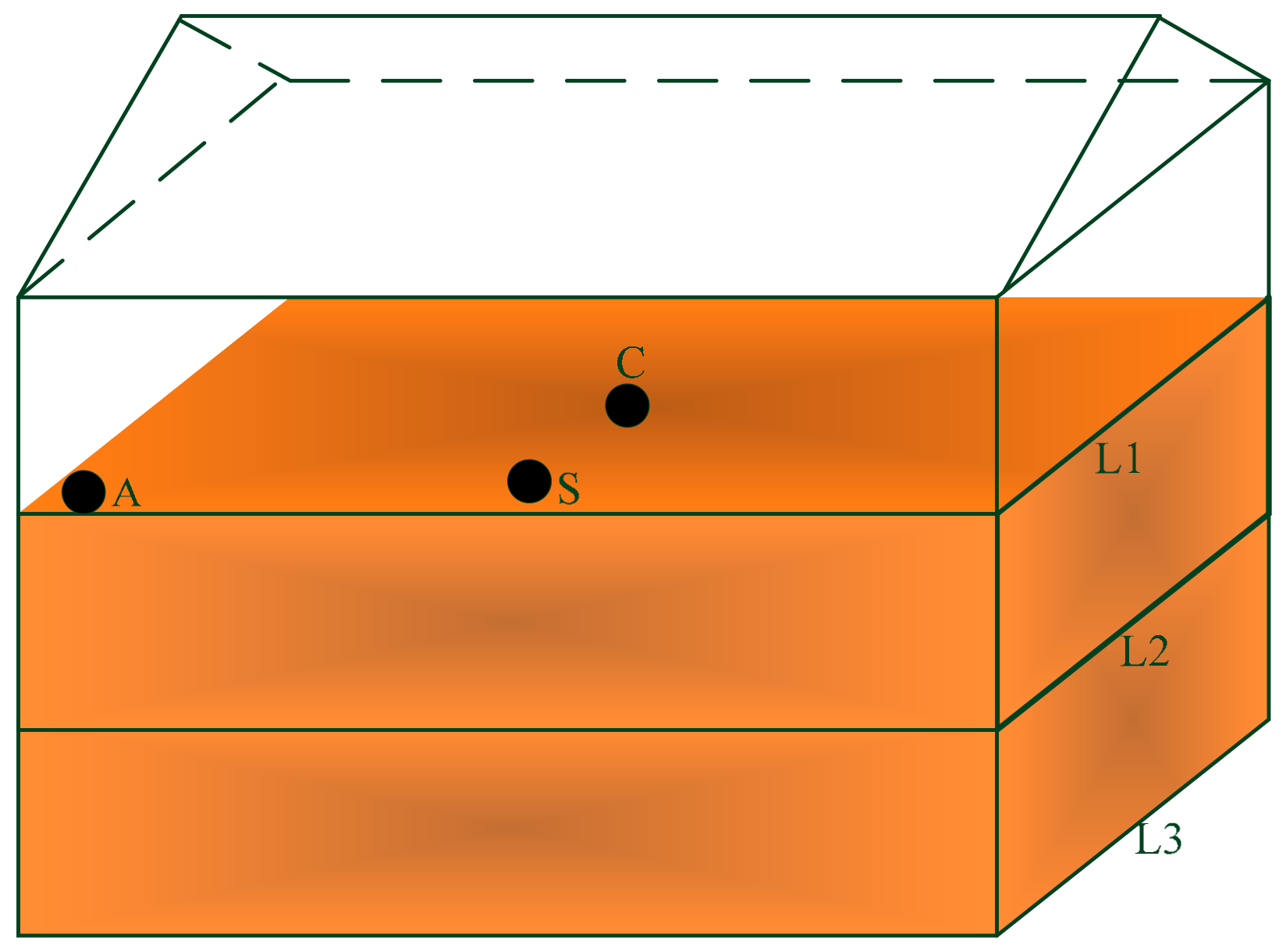
2.1. Rice Samples
2.2. Collection and Isolation of Fungi
2.3. Analysis of Fungi Diversity by High-Throughput Sequencing
2.3.1. DNA Extraction and Sequencing
2.3.2. Data Analysis
2.3.3. Statistical Analysis
3. Results
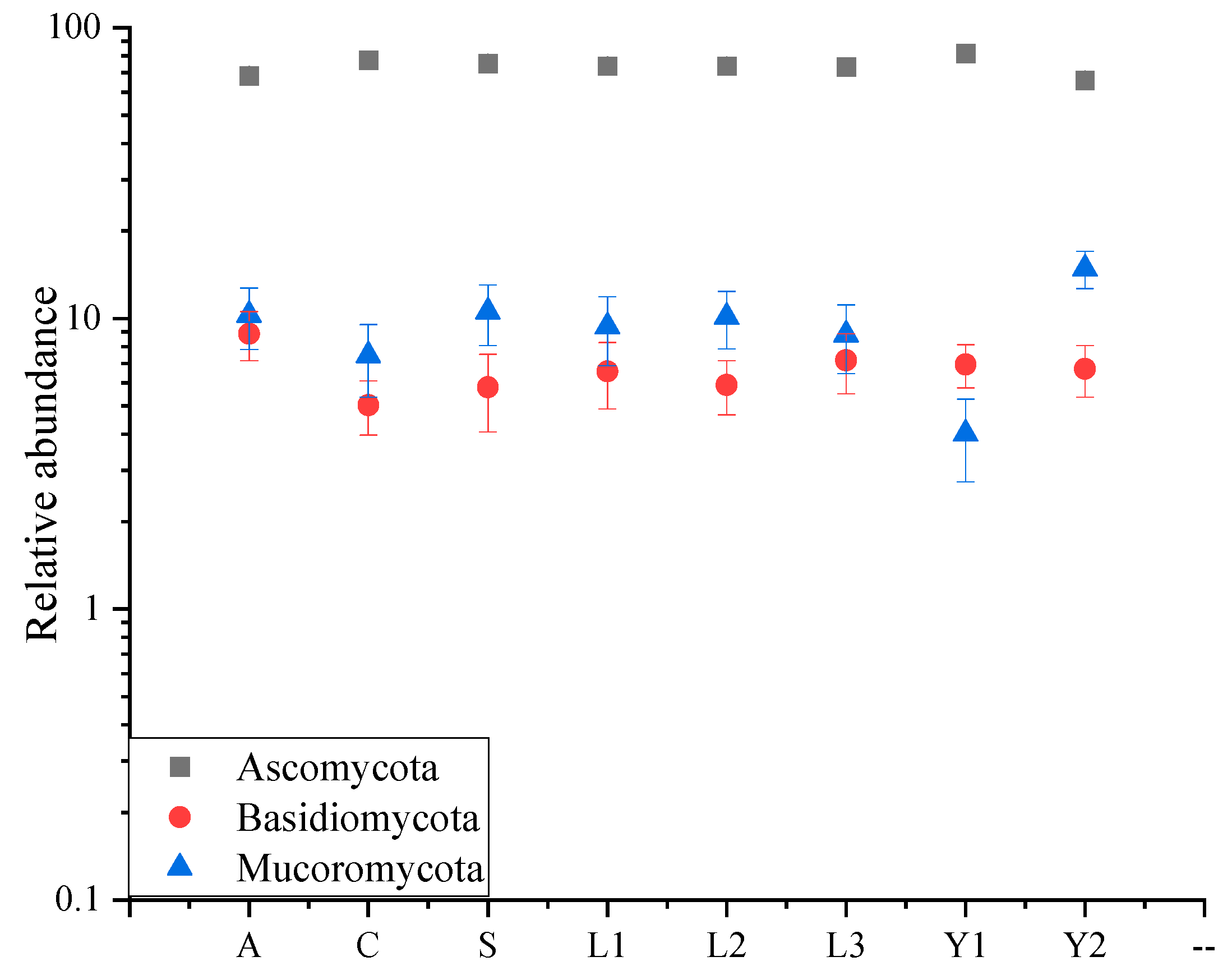
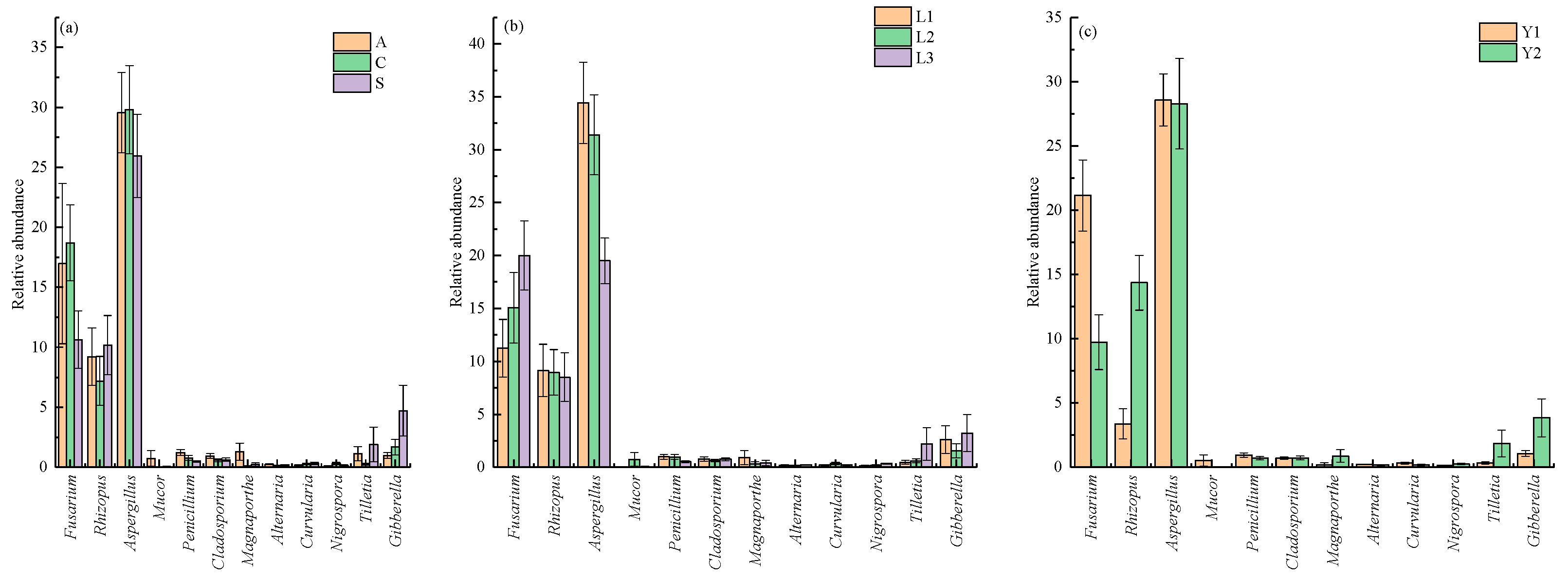
3.1. Fungal Diversity of Rice Samples
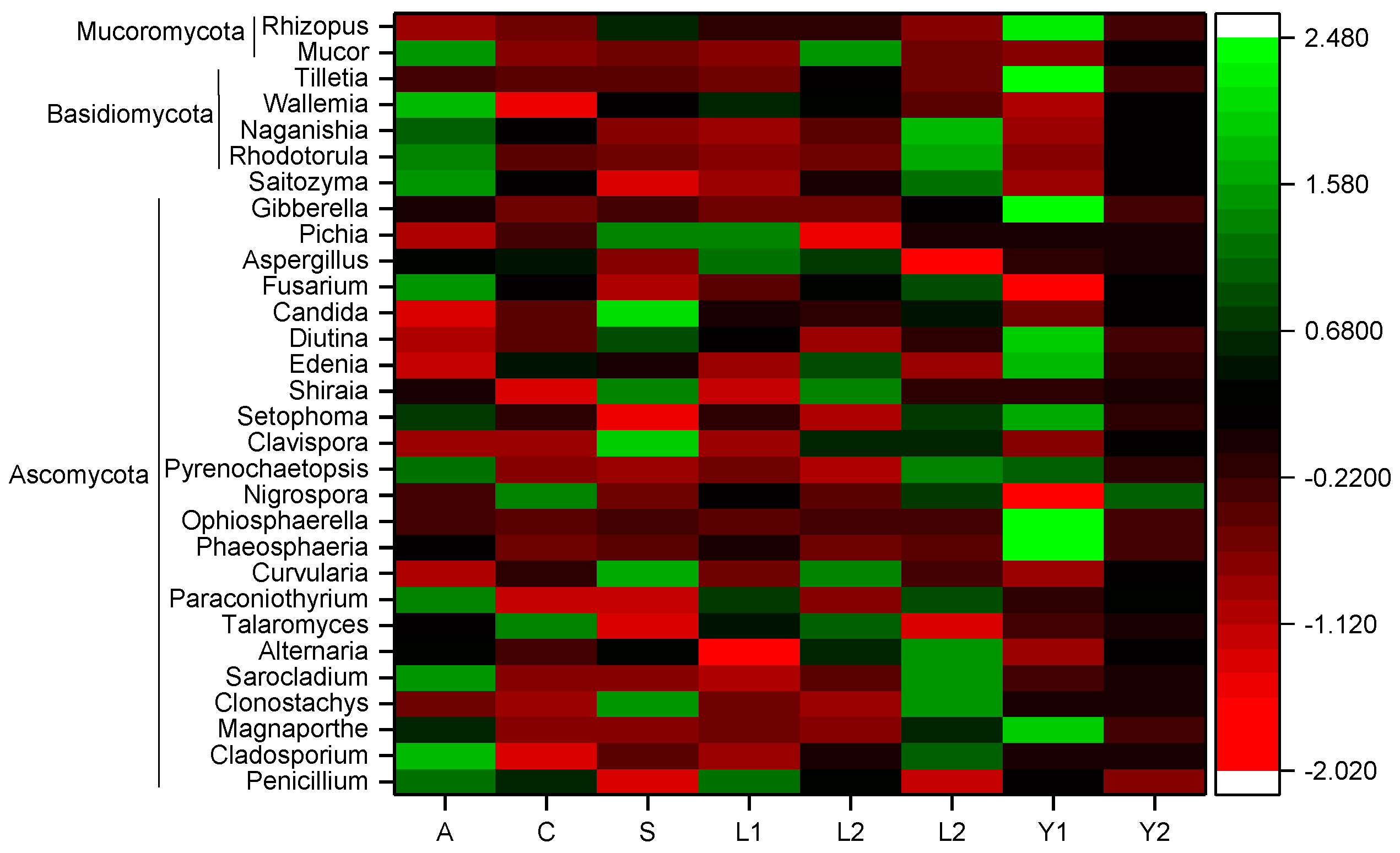
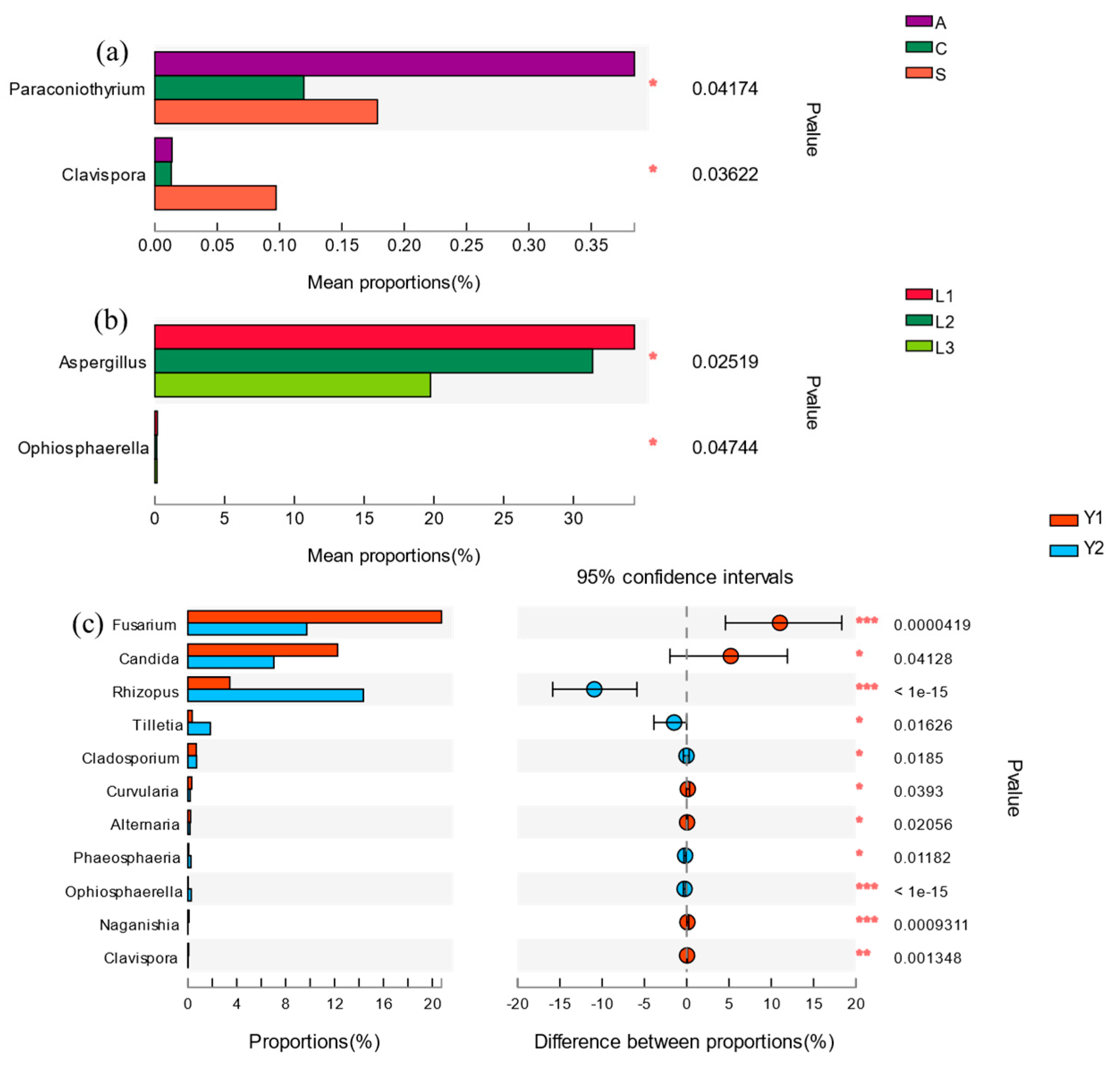
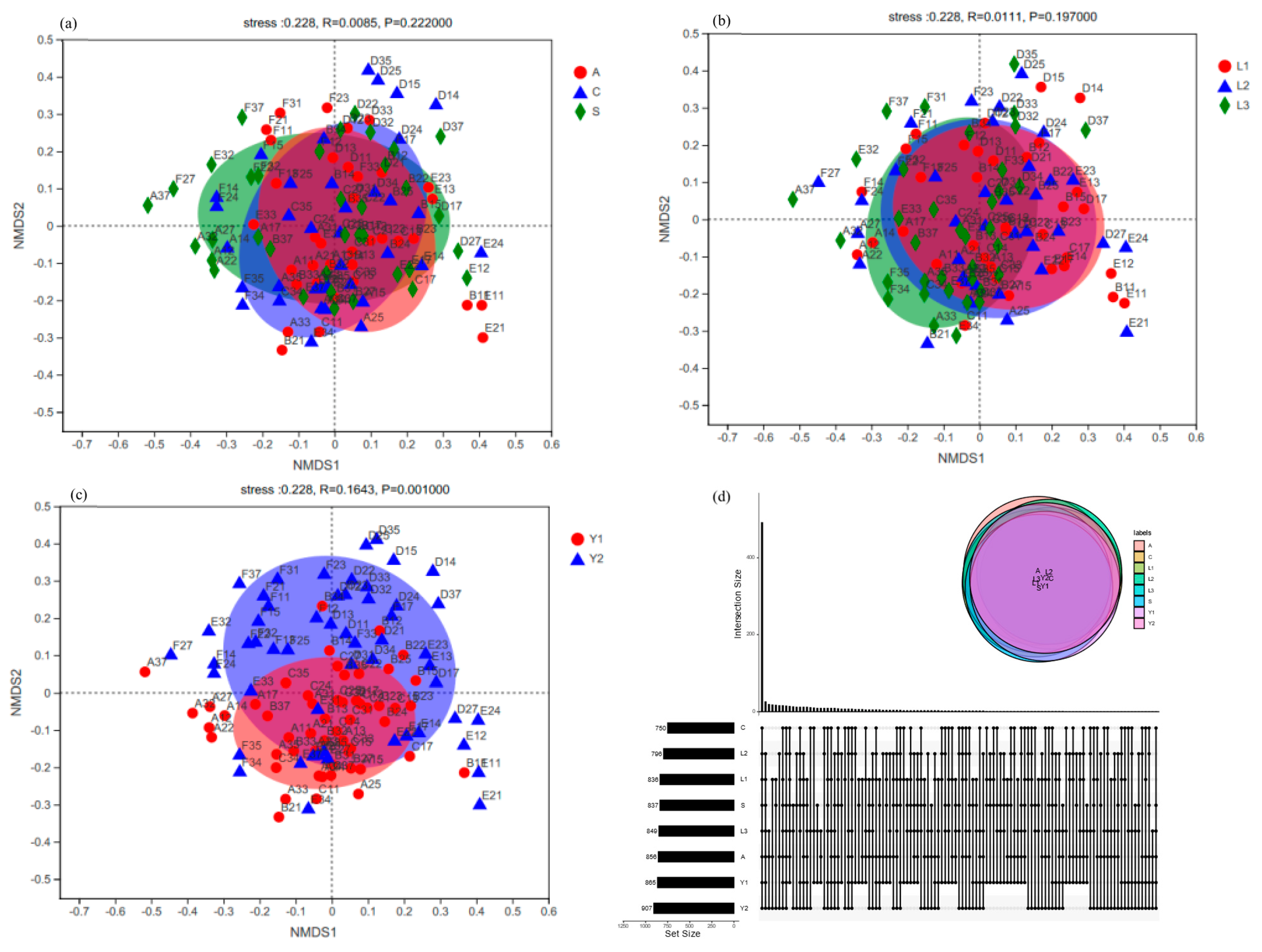
3.2. Fungi Communities Variation and Evolution in Storage
3.3. Core Fungi Community of Rice Samples
4. Discussion
4.1. Effect of Granary Orientation, Vertical Position and Storage Time on Rice Fungi Communities in Storage
4.2. Core Fungi of Rice Storage
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tong, C.; Gao, H.; Luo, S.; Liu, L.; Bao, J. Impact of Postharvest Operations on Rice Grain Quality: A Review. Compr. Rev. Food Sci. Food Saf. 2019, 18, 626–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Shad, Z.M.; Strappe, P.; Xu, L.; Zhang, F.; Chen, Y.; Li, D. A review on rice yellowing: Physicochemical properties, affecting factors, and mechanism. Food Chem. 2022, 370, 131265. [Google Scholar] [CrossRef]
- Prom-U-Thai, C.; Rerkasem, B. Rice quality improvement. A review. Agron. Sustain. Dev. 2020, 40, 28. [Google Scholar] [CrossRef]
- Bertuzzi, T.; Romani, M.; Rastelli, S.; Giorni, P. Mycotoxins and Related Fungi in Italian Paddy Rice During the Growing Season and Storage. Toxins 2019, 11, 151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, S.; Jun, H.; Bang, J.; Chung, S.-H.; Kim, Y.; Kim, B.-S.; Kim, H.; Beuchat, L.R.; Ryu, J.-H. Behaviour of Aspergillus flavus and Fusarium graminearum on rice as affected by degree of milling, temperature, and relative humidity during storage. Food Microbiol. 2015, 46, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Zhang, B.; Chen, S.; Tao, T.; Liu, Q.; Liu, B.; Yuan, J.; Ding, C. Characterization of differences between microwave and traditional thermal sterilization to prevent fungal spoilage during storage of high-moisture paddy rice. Cereal Chem. 2021, 98, 154–163. [Google Scholar] [CrossRef]
- Chai, Q.; Li, Y.; Li, X.; Wu, W.; Peng, H.; Jia, R.; Sun, Q. Assessment of variation in paddy microbial communities under different storage temperatures and relative humidity by Illumina sequencing analysis. Food Res. Int. 2019, 126, 108581. [Google Scholar] [CrossRef]
- Shad, Z.M.; Atungulu, G.G. Post-harvest kernel discoloration and fungi activity in long-grain hybrid, pureline and medium-grain rice cultivars as influenced by storage environment and antifungal treatment. J. Stored Prod. Res. 2019, 81, 91–99. [Google Scholar] [CrossRef]
- Duan, S.; Yang, W.; Wang, X.; Mao, S.; Zhang, Y. Forecasting of Grain Pile Temperature From Meteorological Factors Using Machine Learning. IEEE Access 2019, 7, 130721–130733. [Google Scholar] [CrossRef]
- Peng, D.; Nie, J.; Chen, W. Performance and energy consumption of liquid desiccant dehumidification grain in-bin drying system via layered model. Trans. Chin. Soc. Agric. Eng. 2021, 37, 274–282. [Google Scholar]
- Syamsiyah, J.; Sunarminto, B.H.; Hanudin, E.; Widada, J.; Herawati, A. Assessing soil carbon sequestration in upland rice systems using rice straw and mycorrhiza. IOP Conf. Ser. Earth Environ. Sci. 2018, 200, 012028. [Google Scholar] [CrossRef]
- He, X.; Liu, H.; Lv, C.; Wang, F.; Zhao, C.; Tao, R.; Li, J.; Liu, Z.; Du, L. Analysis of rice microbial communities under different storage conditions using culture-dependent and -independent techniques. Qual. Assur. Saf. Crop. Foods 2022, 14, 1–11. [Google Scholar] [CrossRef]
- Kumar, D.; Singh, K.N.; Shamim, M.; Kumar, M.; Siddiqui, M.W.; Srivastava, D.; Kumar, S.; Kumar, R.; Upadhyay, P.K. Storage of fungi with rice (Oryza sativa)-PRH 10 and their influence on seed quality. Indian J. Agric. Sci. 2020, 90, 1250–1253. [Google Scholar]
- Toju, H.; Tanabe, A.; Yamamoto, S.; Sato, H. High-Coverage ITS Primers for the DNA-Based Identification of Ascomycetes and Basidiomycetes in Environmental Samples. PLoS ONE 2012, 7, e40863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Z.; Zhou, X.; Tian, L.; Zhang, H.; Cai, L.; Tang, F. Distribution of mycotoxin-producing fungi across major rice production areas of China. Food Control 2022, 134, 108572. [Google Scholar] [CrossRef]
- He, P.; Hassan, M.; Tang, F.; Jiang, H.; Chen, M.; Liu, R.; Lin, H.; Chen, Q. Total Fungi Counts and Metabolic Dynamics of Volatile Organic Compounds in Paddy Contaminated by Aspergillus niger During Storage Employing Gas Chromatography-Ion Mobility Spectrometry. Food Anal. Methods 2022, 1–14. [Google Scholar] [CrossRef]
- Drumonde-Neves, J.; Čadež, N.; Reyes-Domínguez, Y.; Gallmetzer, A.; Schuller, D.; Lima, T.; Pais, C.; Franco-Duarte, R. Clavispora santaluciae f.a., sp. nov., a novel ascomycetous yeast species isolated from grapes. Int. J. Syst. Evol. Microbiol. 2020, 70, 6307–6312. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Ren, F.; Che, Y.; Liu, G.; Liu, L. New Bergamotane Sesquiterpenoids from the Plant Endophytic Fungus Paraconiothyrium brasiliense. Molecules 2015, 20, 14611–14620. [Google Scholar] [CrossRef] [Green Version]
- Saleh, A.S.M.; Wang, P.; Wang, N.; Yang, L.; Xiao, Z. Brown Rice Versus White Rice: Nutritional Quality, Potential Health Benefits, Development of Food Products, and Preservation Technologies. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1070–1096. [Google Scholar] [CrossRef] [Green Version]
- Golroodbari, M.S.; Nosrati, A.C. Evaluation of Aspergillus spp. reflecting to subgenus circumdati section flavi contamination in the gilan province, is stored and fresh harvested rice crops. IIOAB J. 2016, 7, 58–61. [Google Scholar]
- Xu, J.; Liu, K.; Zhang, C. Electronic nose for volatile organic compounds analysis in rice aging. Trends Food Sci. Technol. 2021, 109, 83–93. [Google Scholar] [CrossRef]
- Ding, C.; Khir, R.; Pan, Z.; Wood, D.F.; Venkitasamy, C.; Tu, K.; El Mashad, H.; Berrios, J. Influence of infrared drying on storage characteristics of brown rice. Food Chem. 2018, 264, 149–156. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Khir, R.; Pan, Z.; El Mashad, H.; Atungulu, G.G.; Ma, H.; McHugh, T.H.; Qu, W.; Wu, B. Effective Disinfection of Rough Rice Using Infrared Radiation Heating. J. Food Prot. 2014, 77, 1538–1545. [Google Scholar] [CrossRef] [PubMed]
- Müller, A.; Nunes, M.T.; Maldaner, V.; Coradi, P.C.; de Moraes, R.S.; Martens, S.; Leal, A.F.; Pereira, V.F.; Marin, C.K. Rice Drying, Storage and Processing: Effects of Post-Harvest Operations on Grain Quality. Rice Sci. 2022, 29, 16–30. [Google Scholar] [CrossRef]
- Lim, D.; Mo, C.; Lee, D.-K.; Long, N.P.; Lim, J.; Kwon, S.W. Non-destructive profiling of volatile organic compounds using HS-SPME/GC–MS and its application for the geographical discrimination of white rice. J. Food Drug Anal. 2018, 26, 260–267. [Google Scholar] [CrossRef]
- Gu, S.; Wang, Z.; Wang, J. Untargeted rapid differentiation and targeted growth tracking of fungal contamination in rice grains based on headspace-gas chromatography-ion mobility spectrometry. J. Sci. Food Agric. 2021. [Google Scholar] [CrossRef]
- Zhao, C.; Su, W.; Mu, Y.; Jiang, L.; Mu, Y. Correlations between microbiota with physicochemical properties and volatile flavor compo-nents in black glutinous rice wine fermentation. Food Res. Int. 2020, 138, 109800. [Google Scholar] [CrossRef]
- Dossou, B.; Silue, D. Rice pathogens intercepted on seeds originating from 11 African countries and from the USA. Seed Sci. Technol. 2018, 46, 31–40. [Google Scholar] [CrossRef]
- Lee, S.-B.; Kim, N.; Hur, Y.-J.; Cho, S.-M.; Kim, T.-H.; Lee, J.-Y.; Cho, J.-H.; Lee, J.-H.; Song, Y.-C.; Seo, Y.-S.; et al. Fine mapping of qBK1, a major QTL for bakanae disease resistance in rice. Rice 2019, 12, 136. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Nysetvold, E.; Zhou, X.-G. Culture media promoting sporulation of rice kernel smut fungus Tilletia barclayana. Eur. J. Plant Pathol. 2021, 161, 629–635. [Google Scholar] [CrossRef]
- Makun, H.A.; Gbodi, T.A.; Akanya, O.H.; Salako, E.A.; Ogbadu, G.H. Fungi and some mycotoxins contaminating rice (Oryza sativa) in Niger State, Nigeria. Afr. J. Biotechnol. 2007, 6, 99–108. [Google Scholar]
- Khosravi, A.R.; Shokri, H.; Zaboli, F. Grain-Borne Mycoflora and Fumonisin B1 From Fresh-Harvested and Stored Rice in Northern Iran. Jundishapur J. Microbiol. 2013, 6, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Khaleghian, H.; Karimi, K.; Behzad, T. Ethanol production from rice straw by sodium carbonate pretreatment and Mucor hiemalis fermentation. Ind. Crop. Prod. 2015, 76, 1079–1085. [Google Scholar] [CrossRef]
- Liu, N.; Pan, J.; Miao, S.; Qin, L. Microbial community in Chinese traditional fermented acid rice soup (rice-acid) and its correlations with key organic acids and volatile compounds. Food Res. Int. 2020, 137, 109672. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Chen, J.; Wu, W.; Du, Z.; Guan, Y. Immobilization of Laccase on Magnetic Nanoparticles and Application in the Detoxification of Rice Straw Hydrolysate for the Lipid Production of Rhodotorula glutinis. Appl. Biochem. Biotechnol. 2021, 193, 998–1010. [Google Scholar] [CrossRef]
- Li, N.; Zhang, B.; Zhao, S.; Niu, M.; Jia, C.; Huang, Q.; Liu, Y.; Lin, Q. Influence of Lactobacillus/Candida fermentation on the starch structure of rice and the related noodle features. Int. J. Biol. Macromol. 2019, 121, 882–888. [Google Scholar] [CrossRef]
- Zhu, A.; Tan, H.; Cao, L. Isolation of phytase-producing yeasts from rice seedlings for prospective probiotic applications. 3 Biotech 2019, 9, 216. [Google Scholar] [CrossRef]
- Samsudin, N.I.P.; Abdullah, N. A preliminary survey on the occurrence of mycotoxigenic fungi and mycotoxins contaminating red rice at consumer level in Selangor, Malaysia. Mycotoxin Res. 2013, 29, 89–96. [Google Scholar] [CrossRef]
- Bernaola, L.; Cange, G.; Way, M.O.; Gore, J.; Hardke, J.; Stout, M. Natural Colonization of Rice by Arbuscular Mycorrhizal Fungi in Different Production Areas. Rice Sci. 2018, 25, 169–174. [Google Scholar] [CrossRef]
- Qiu, J.H.; Meng, S.; Deng, Y.Z.; Huang, S.W.; Kou, Y.J. Ustilaginoidea virens: A Fungus Infects Rice Flower and Threats World Rice Production. Rice Sci. 2019, 26, 199–206. [Google Scholar] [CrossRef]
- Pitt, J.I.; Miller, J.D. A Concise History of Mycotoxin Research. J. Agric. Food Chem. 2017, 65, 7021–7033. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Fungus | ||
|---|---|---|---|
| Shannon | Chao1 | ||
| Orientation | A | 2.68 ± 0.11 a | 317.02 ± 16.90 a |
| C | 2.58 ± 0.09 a | 264.65 ± 15.80 b | |
| S | 2.41 ± 0.10 a | 291.46 ± 13.08 ab | |
| Vertical position | L1 | 2.58 ± 0.11 a | 290.07 ± 17.68 a |
| L2 | 2.57 ± 0.10 a | 283.26 ± 14.36 a | |
| L3 | 2.52 ± 0.11 a | 299.30 ± 14.75 a | |
| Storage time | Y1 | 2.53 ± 0.09 a | 321.08 ± 10.09 a |
| Y2 | 2.58 ± 0.08 a | 261.08 ± 13.87 b | |
| Shannon | Chao1 | |
|---|---|---|
| Horizontal | 2.92 | 5.98 |
| Vertical | 0.09 | 0.11 |
| Storage time | 0.20 | 17.18 |
| Sums of Squares | Mean Squares | F.Model | R2 | p-Value | |
|---|---|---|---|---|---|
| Horizontal | 0.67926 | 0.33963 | 1.31205 | 0.02438 | 0.13 |
| Vertical | 0.62467 | 0.31234 | 1.2042 | 0.02242 | 0.266 |
| Storage time | 1.76889 | 1.76889 | 7.18676 | 0.06349 | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Cui, J.; Li, J.; Guo, J.; Huang, T.; Zhang, J.; Hu, H.; Liu, X. Analysis of the Fungi Community Variation during Rice Storage through High Throughput Sequencing. Processes 2022, 10, 754. https://doi.org/10.3390/pr10040754
Li W, Cui J, Li J, Guo J, Huang T, Zhang J, Hu H, Liu X. Analysis of the Fungi Community Variation during Rice Storage through High Throughput Sequencing. Processes. 2022; 10(4):754. https://doi.org/10.3390/pr10040754
Chicago/Turabian StyleLi, Wanting, Jie Cui, Jiafeng Li, Jian Guo, Tao Huang, Jiaojiao Zhang, Hao Hu, and Xingquan Liu. 2022. "Analysis of the Fungi Community Variation during Rice Storage through High Throughput Sequencing" Processes 10, no. 4: 754. https://doi.org/10.3390/pr10040754
APA StyleLi, W., Cui, J., Li, J., Guo, J., Huang, T., Zhang, J., Hu, H., & Liu, X. (2022). Analysis of the Fungi Community Variation during Rice Storage through High Throughput Sequencing. Processes, 10(4), 754. https://doi.org/10.3390/pr10040754