
Investigation of the Relation between Temperature and M13 Phage Production via ATP Expenditure

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation for M13 Phage Production
2.2. M13 Phage Production
2.3. M13 Phage Purification via Polyethylene Glycol (PEG) Precipitation
2.4. E. coli Growth Level Analysis via Optical Density Measurements
2.5. Intracellular ATP Analysis via Luminescence Assay
2.6. M13 Phage Titer Calculation via Titration
3. Results and Discussion
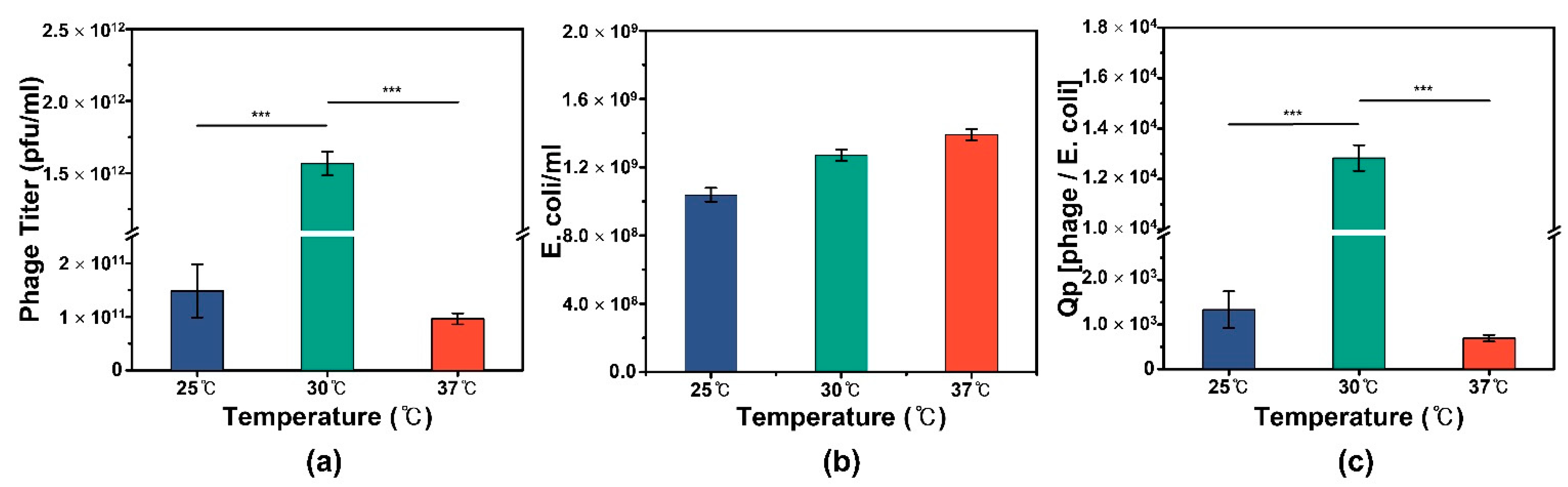
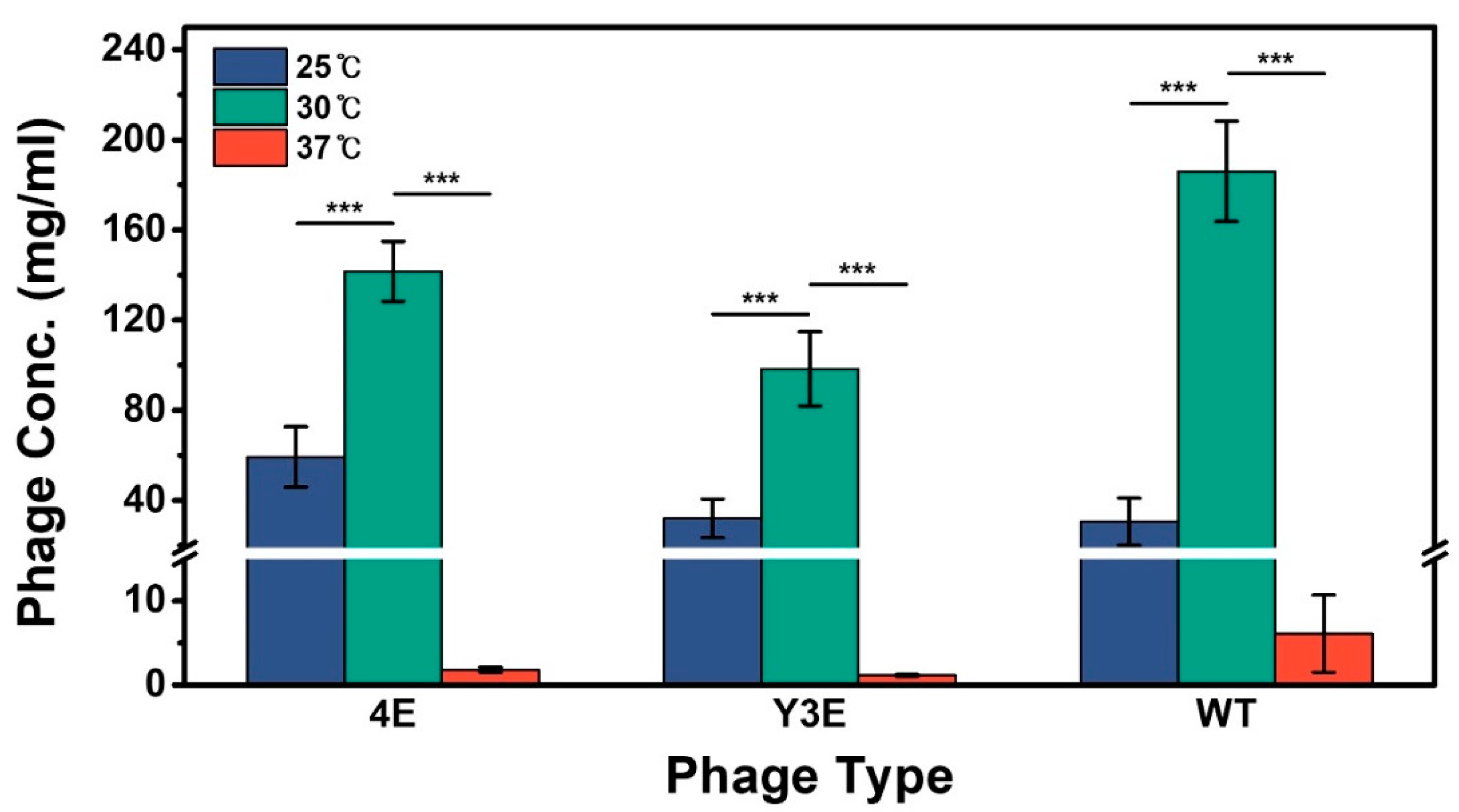
3.1. Comparison between Optimum and Control Temperatures for M13 Phage Production
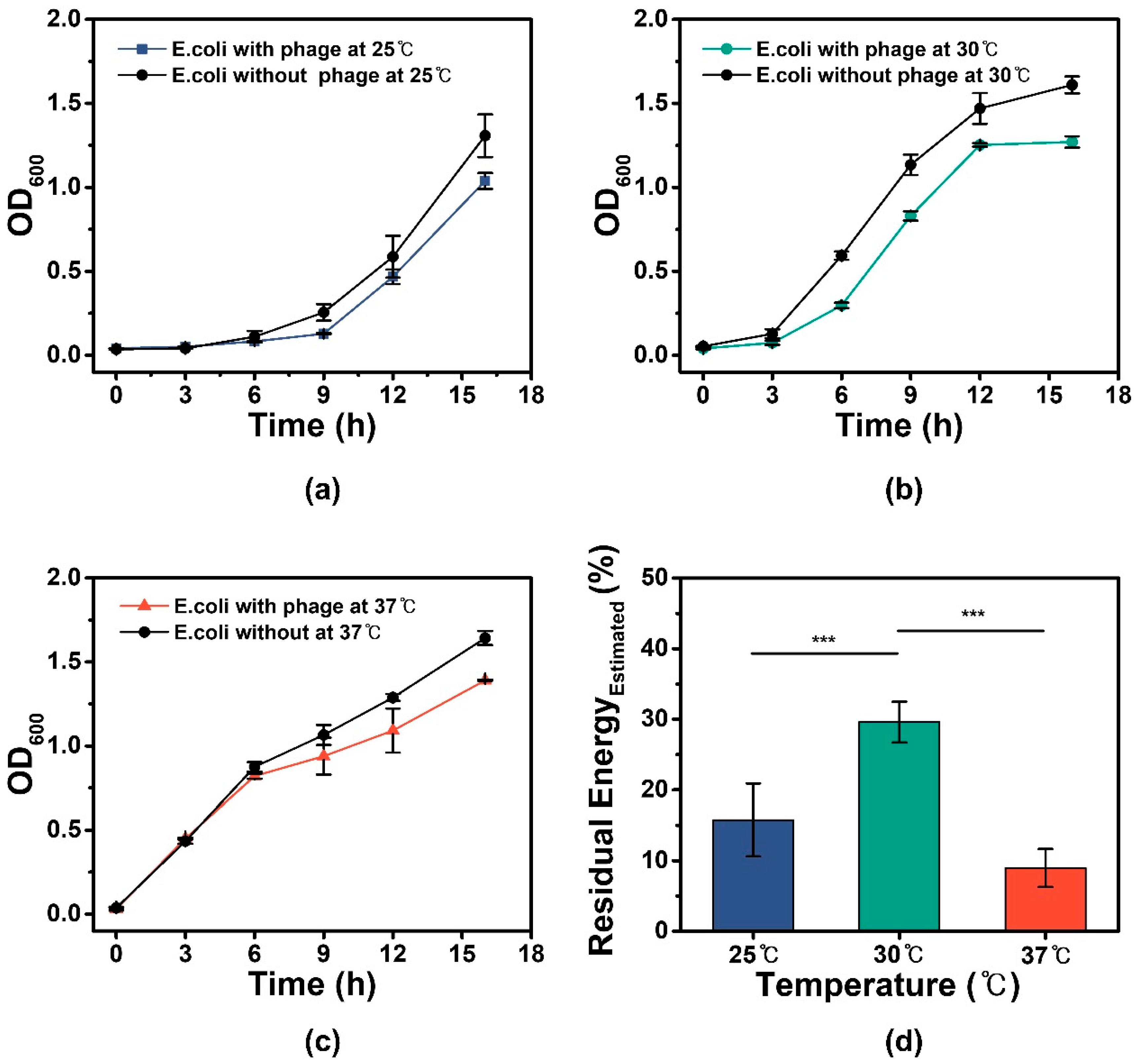
3.2. E. coli Specific Growth Rate Analysis and Estimation of Residual Energy for M13 Phage Production
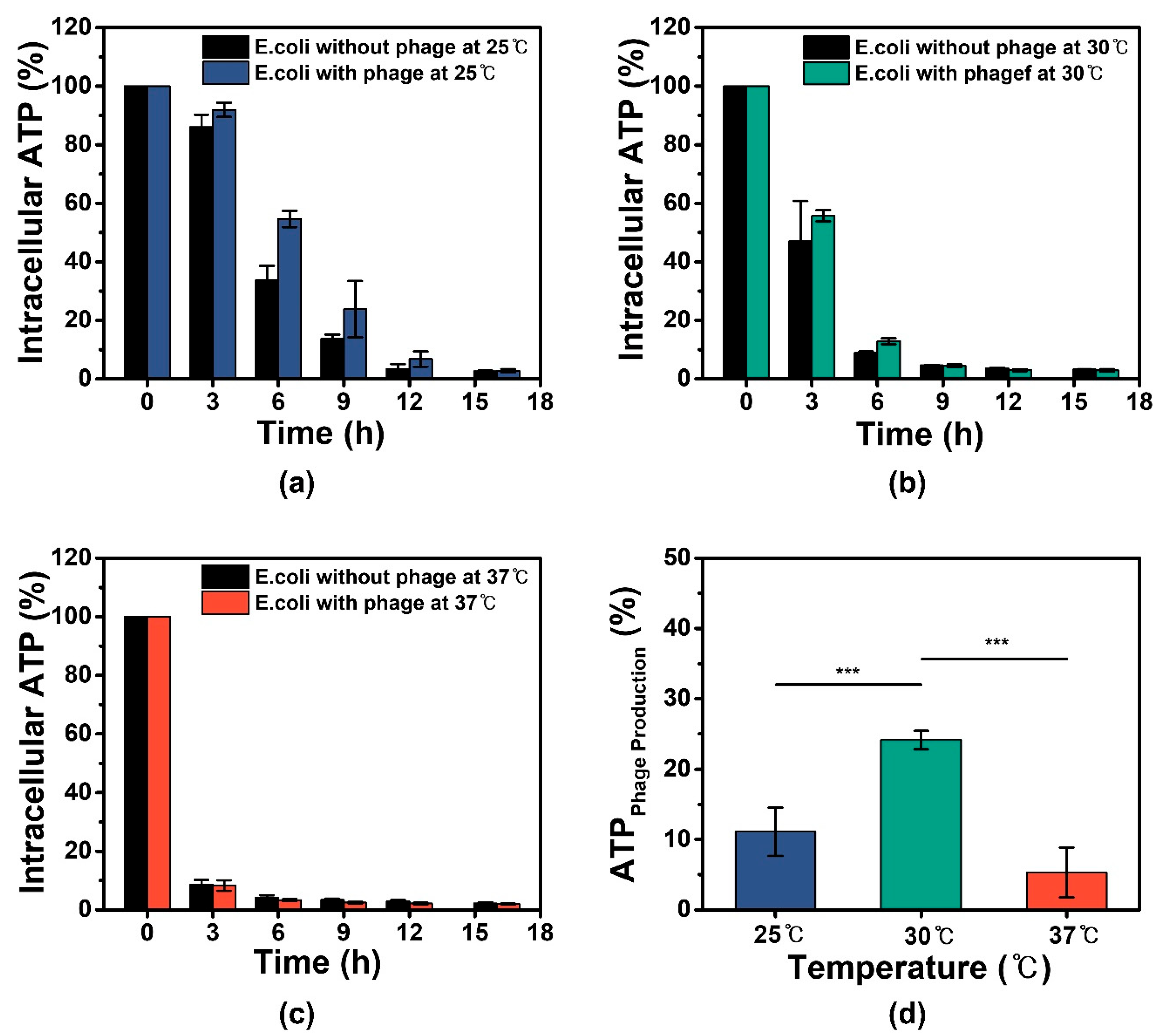
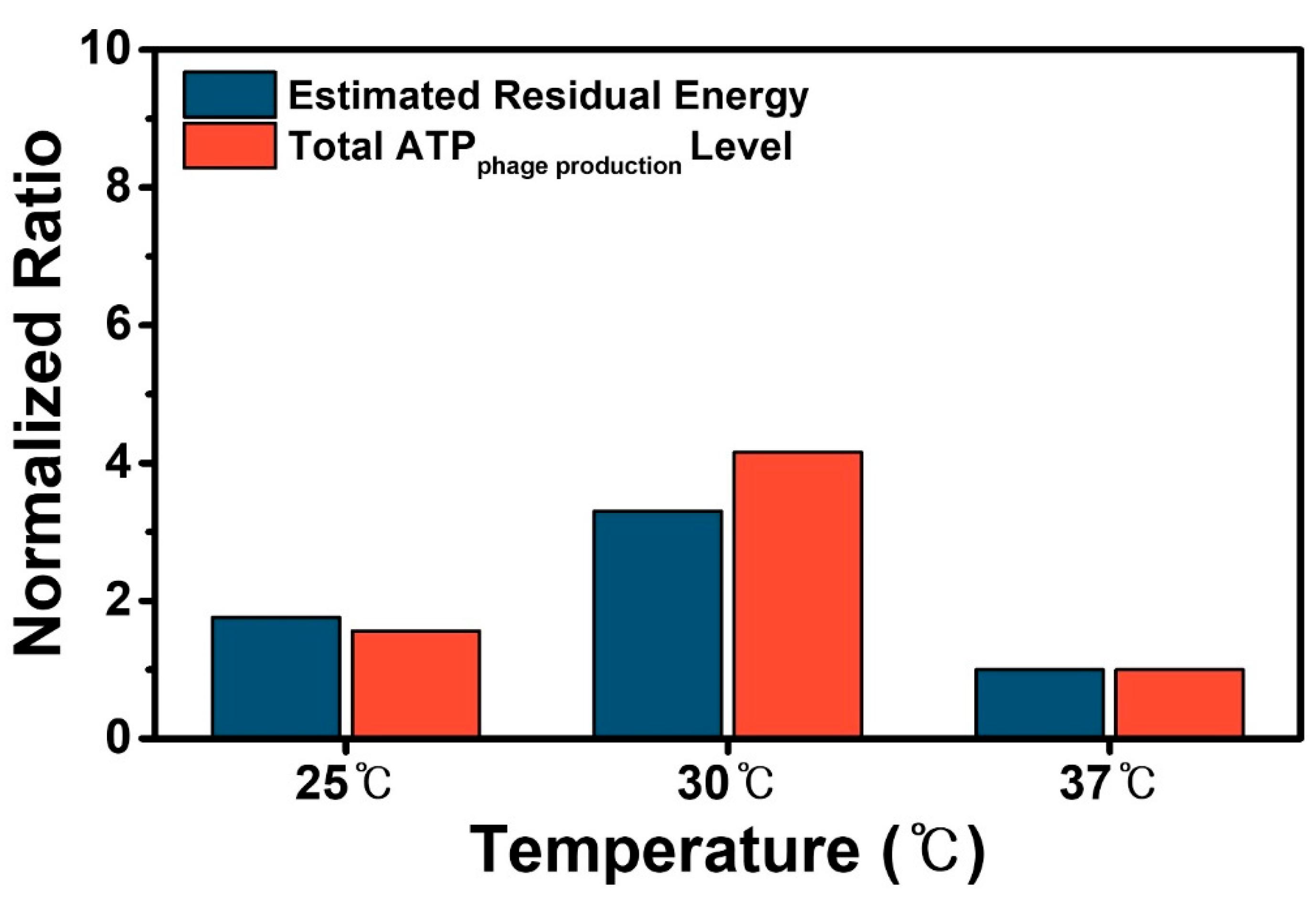
3.3. ATP Expenditure Analysis and Comparison with Residual Energy for M13 Phage Production
3.4. Verification of the Optimum Culture Temperature for Genetically Engineered M13 Phage Production
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smeal, S.W.; Schmitt, M.A.; Pereira, R.R.; Prasad, A.; Fisk, J.D. Simulation of the M13 life cycle I: Assembly of a genetically-structured deterministic chemical kinetic simulation. Virology 2017, 500, 259–274. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Warner, C.M.; Jin, H.-E.; Barnes, E.; Poda, A.R.; Perkins, E.J.; Lee, S.-W. Production of tunable nanomaterials using hierarchically assembled bacteriophages. Nat. Protoc. 2017, 12, 1999–2013. [Google Scholar] [CrossRef] [PubMed]
- Chung, W.J.; Oh, J.W.; Kwak, K.; Lee, B.Y.; Meyer, J.; Wang, E.; Hexemer, A.; Lee, S.W. Biomimetic self-templating supramolecular structures. Nature 2011, 478, 364–368. [Google Scholar] [CrossRef] [PubMed]
- Onsager, L. The effects of shape on the interaction of colloidal particles. Ann. N. Y. Acad. Sci. 1949, 51, 627–659. [Google Scholar] [CrossRef]
- Lee, B.Y.; Zhang, J.; Zueger, C.; Chung, W.J.; Yoo, S.Y.; Wang, E.; Meyer, J.; Ramesh, R.; Lee, S.W. Virus-based piezoelectric energy generation. Nat. Nanotechnol. 2012, 7, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharjee, S.; Glucksman, M.J.; Makowski, L. Structural polymorphism correlated to surface charge in filamentous bacteriophages. Biophys. J. 1992, 61, 725–735. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Fan, B.; Samdin, T.D.; Monteiro, D.A.; Desai, M.S.; Scheideler, O.; Jin, H.E.; Kim, S.; Lee, S.W. Phage-Based Structural Color Sensors and Their Pattern Recognition Sensing System. ACS Nano 2017, 11, 3632–3641. [Google Scholar] [CrossRef]
- Gibaud, T.; Barry, E.; Zakhary, M.J.; Henglin, M.; Ward, A.; Yang, Y.; Berciu, C.; Oldenbourg, R.; Hagan, M.F.; Nicastro, D.; et al. Reconfigurable self-assembly through chiral control of interfacial tension. Nature 2012, 481, 348–351. [Google Scholar] [CrossRef]
- Ferrara, F.; Kim, C.-Y.; Naranjo, L.A.; Bradbury, A.R.M. Large scale production of phage antibody libraries using a bioreactor. mAbs 2015, 7, 26–31. [Google Scholar] [CrossRef] [Green Version]
- Grieco, S.H.; Lee, S.; Dunbar, W.S.; MacGillivray, R.T.; Curtis, S.B. Maximizing filamentous phage yield during computer-controlled fermentation. Bioprocess Biosyst. Eng. 2009, 32, 773–779. [Google Scholar] [CrossRef]
- Grieco, S.H.; Wong, A.Y.; Dunbar, W.S.; MacGillivray, R.T.; Curtis, S.B. Optimization of fermentation parameters in phage production using response surface methodology. J. Ind. Microbiol. Biotechnol. 2012, 39, 1515–1522. [Google Scholar] [CrossRef] [PubMed]
- Kick, B.; Hensler, S.; Praetorius, F.; Dietz, H.; Weuster-Botz, D. Specific growth rate and multiplicity of infection affect high-cell-density fermentation with bacteriophage M13 for ssDNA production. Biotechnol. Bioeng. 2017, 114, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Warner, C.M.; Barker, N.; Lee, S.-W.; Perkins, E.J. M13 bacteriophage production for large-scale applications. Bioprocess Biosyst. Eng. 2014, 37, 2067–2072. [Google Scholar] [CrossRef] [PubMed]
- Bijlenga, R.K.L.; Aebi, U.; Kellenberger, E. Properties and structure of a gene 24-controlled T4 giant phage. J. Mol. Biol. 1976, 103, 469–498. [Google Scholar] [CrossRef]
- Groman, N.B.; Suzuki, G. Temperature and lambda phage reproduction. J. Bacteriol. 1962, 84, 431–437. [Google Scholar] [CrossRef] [Green Version]
- Krueger, A.P.; Fong, J. The relationship between bacterial growth and phage production. J. Gen. Physiol. 1937, 21, 137–150. [Google Scholar] [CrossRef] [Green Version]
- Lopez, J.; Webster, R.E. fipB and fipC: Two bacterial loci required for morphogenesis of the filamentous bacteriophage f1. J. Bacteriol. 1985, 163, 900–905. [Google Scholar] [CrossRef] [Green Version]
- Mitra, S.; Stallions, D.R. Role of dna genes of Escherichia coli in M13 phage replication. Virology 1973, 52, 417–424. [Google Scholar] [CrossRef]
- Olsen, W.L.; Staudenbauer, W.L.; Hofschneider, P.H. Replication of bacteriophage M13: Specificity of the Escherichia coli dnaB function for replication of double-stranded M13 DNA. Proc. Natl. Acad. Sci. USA 1972, 69, 2570–2573. [Google Scholar] [CrossRef] [Green Version]
- Speck, J.; Arndt, K.M.; Müller, K.M. Efficient phage display of intracellularly folded proteins mediated by the TAT pathway. Protein Eng. Des. Sel. 2011, 24, 473–484. [Google Scholar] [CrossRef] [Green Version]
- Mahmoudabadi, G.; Milo, R.; Phillips, R. Energetic cost of building a virus. Proc. Natl. Acad. Sci. USA 2017, 114, E4324–E4333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Serwer, P.; Wright, E.T. ATP-Driven Contraction of Phage T3 Capsids with DNA Incompletely Packaged In Vivo. Viruses 2017, 9, 119. [Google Scholar] [CrossRef] [Green Version]
- Hay, I.D.; Lithgow, T. Filamentous phages: Masters of a microbial sharing economy. EMBO Rep. 2019, 20, e47427. [Google Scholar] [CrossRef] [PubMed]
- Loh, B.; Kuhn, A.; Leptihn, S. The fascinating biology behind phage display: Filamentous phage assembly. Mol. Microbiol. 2019, 111, 1132–1138. [Google Scholar] [CrossRef] [Green Version]
- Loh, B.; Haase, M.; Mueller, L.; Kuhn, A.; Leptihn, S. The Transmembrane Morphogenesis Protein gp1 of Filamentous Phages Contains Walker A and Walker B Motifs Essential for Phage Assembly. Viruses 2017, 9, 73. [Google Scholar] [CrossRef] [Green Version]
- Feng, D.-F.; Cho, G.; Doolittle, R.F. Determining divergence times with a protein clock: Update and reevaluation. Proc. Natl. Acad. Sci. USA 1997, 94, 13028–13033. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.N.; Model, P.; Russel, M. A trans-envelope protein complex needed for filamentous phage assembly and export. Mol. Microbiol. 1999, 34, 745–755. [Google Scholar] [CrossRef]
- Smeal, S.W.; Schmitt, M.A.; Pereira, R.R.; Prasad, A.; Fisk, J.D. Simulation of the M13 life cycle II: Investigation of the control mechanisms of M13 infection and establishment of the carrier state. Virology 2017, 500, 275–284. [Google Scholar] [CrossRef]
- Maitra, A.; Dill, K.A. Bacterial growth laws reflect the evolutionary importance of energy efficiency. Proc. Natl. Acad. Sci. USA 2015, 112, 406–411. [Google Scholar] [CrossRef] [Green Version]
- Bremer, H.; Dennis, P.P. Modulation of Chemical Composition and Other Parameters of the Cell at Different Exponential Growth Rates. EcoSal Plus 2008, 3, 1–48. [Google Scholar] [CrossRef]
- Stouthamer, A.H.; Bettenhaussen, C. Utilization of energy for growth and maintenance in continuous and batch cultures of microorganisms. A reevaluation of the method for the determination of ATP production by measuring molar growth yields. Biochim. Biophys. Acta 1973, 301, 53–70. [Google Scholar] [CrossRef]
- Tempest, D.W.; Neijssel, O.M. The status of YATP and maintenance energy as biologically interpretable phenomena. Annu. Rev. Microbiol. 1984, 38, 459–486. [Google Scholar] [CrossRef] [PubMed]
- Scott, M.; Gunderson, C.W.; Mateescu, E.M.; Zhang, Z.; Hwa, T. Interdependence of cell growth and gene expression: Origins and consequences. Science 2010, 330, 1099–1102. [Google Scholar] [CrossRef] [PubMed]
- Kepes, A.; Beguin, S. Peptide chain initiation and growth in the induced synthesis of β-galactosidase. Biochim. Biophys. Acta Nucleic Acids Protein Synth. 1966, 123, 546–560. [Google Scholar] [CrossRef]
- Young, R.; Bremer, H. Polypeptide-chain-elongation rate in Escherichia coli B/r as a function of growth rate. Biochem. J. 1976, 160, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Barbas, C.F.; Burton, D.R.; Silverman, G.J. Phage Display: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Spring harbor, New York, USA, 2004; ISBN 978-087969740-2. [Google Scholar]
- Crookes-Goodson, W.J.; Slocik, J.M.; Naik, R.R. Bio-directed synthesis and assembly of nanomaterials. Chem. Soc. Rev. 2008, 37, 2403–2412. [Google Scholar] [CrossRef]
- Lee, S.K.; Lee, J.H.; Park, C. Rapid and concise quantification of mycelial growth by microscopic image intensity model and application to mass cultivation of fungi. Sci. Rep. 2021, 11, 24157. [Google Scholar]
- Zhang, W.; Chen, X.; Sun, W.; Nie, T.; Quanquin, N.; Sun, Y. Escherichia coli Increases its ATP Concentration in Weakly Acidic Environments Principally through the Glycolytic Pathway. Genes 2020, 11, 991. [Google Scholar] [CrossRef]
- Sun, Y.; Fukamachi, T.; Saito, H.; Kobayashi, H. ATP Requirement for Acidic Resistance in Escherichia coli. J. Bacteriol. 2011, 193, 3072–3077. [Google Scholar] [CrossRef] [Green Version]
- Mempin, R.; Tran, H.; Chen, C.; Gong, H.; Kim Ho, K.; Lu, S. Release of extracellular ATP by bacteria during growth. BMC Microbiol. 2013, 13, 301. [Google Scholar] [CrossRef] [Green Version]
- Kaufmann, S.H.; Reimann, J. Immunology of infection. In Vaccines; Springer: Berlin/Heidelberg, Germany, 1999; pp. 21–42. ISBN 3540647406 9783540647409. [Google Scholar]
- Au-Baer, A.; Au-Kehn-Hall, K. Viral Concentration Determination Through Plaque Assays: Using Traditional and Novel Overlay Systems. JoVE 2014, 4, e52065. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Spring harbor, New York, USA, 2014; ISBN 978-1-936113-41-5. [Google Scholar]
- Farewell, A.; Neidhardt, F.C. Effect of temperature on in vivo protein synthetic capacity in Escherichia coli. J. Bacteriol. 1998, 180, 4704–4710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gant Kanegusuku, A.; Stankovic, I.N.; Cote-Hammarlof, P.A.; Yong, P.H.; White-Ziegler, C.A. A Shift to Human Body Temperature (37 °C) Rapidly Reprograms Multiple Adaptive Responses in Escherichia coli That Would Facilitate Niche Survival and Colonization. J. Bacteriol. 2021, 203, e0036321. [Google Scholar] [CrossRef] [PubMed]
- Hall, B.G.; Acar, H.; Nandipati, A.; Barlow, M. Growth rates made easy. Mol. Biol. Evol. 2014, 31, 232–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Son, J.; Lee, K.H.; Park, C. Enhanced Production of Bacterial Cellulose from Miscanthus as Sustainable Feedstock through Statistical Optimization of Culture Conditions. Int. J. Environ. Res. Public Health 2022, 19, 866. [Google Scholar] [CrossRef]
- Kim, H.; Son, J.; Park, C. Improved production of bacterial cellulose through investigation of effects of inhibitory compounds from lignocellulosic hydrolysates. Glob. Chang. Biol. Bioenergy 2021, 13, 436–444. [Google Scholar] [CrossRef]
- Maaløe, O.; Hanawalt, P.C. Thymine deficiency and the normal DNA replication cycle. I. J. Mol. Biol. 1961, 3, 144–155. [Google Scholar] [CrossRef]
- Karlsson, F.; Malmborg-Hager, A.C. Genome-wide comparison of phage M13-infected vs. uninfected Escherichia coli. Can. J. Microbiol. 2005, 51, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Zhan, X.; Gao, J. RraA: A Protein Inhibitor of RNase E Activity that Globally Modulates RNA Abundance in E. coli. Cell 2003, 114, 623–634. [Google Scholar] [CrossRef]
- Lee, S.Y. Systems Biology and Biotechnology of Escherichia Coli; Springer: Berlin/Heidelberg, Germany, 2009; ISBN 978-1-4020-9393-7. [Google Scholar]
- Schoenhofen, I.C.; Li, G. Purification and Characterization of the N-Terminal Domain of ExeA: A Novel ATPase Involved in the Type II Secretion Pathway of Aeromonas hydrophila. J. Bacteriol. 2005, 187, 6370–6378. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.W.; Liu, H.P. Dual ATPase and GTPase activity of the replication-associated protein (Rep) of beak and feather disease virus. Virus Res. 2016, 213, 149–161. [Google Scholar] [CrossRef] [PubMed]
- Leigh, R.A.; Walker, R.R. ATPase and Acid Phosphatase Activities Associated with Vacuoles Isolated from Storage Roots of Red Beet (Beta vulgaris L.). Planta 1980, 150, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Rocak, S.; Emery, B. Characterization of the ATPase and unwinding activities of the yeast DEAD-box protein Has1p and the analysis of the roles of the conserved motifs. Nucleic Acids Res. 2005, 33, 999–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. a | Amplification Ratio b | Temperature (°C) | pH | Medium | Additives | E. coli Strain | Ref. |
|---|---|---|---|---|---|---|---|
| 1 | 2.50 × 103 | 30 | 7.4 | LB | N/A c | XL1-Blue | Our Study |
| 2 | 2.00 × 103 | 34 | N/A c | Hanawalt | Vitamin B1 Thymidine | HfrH 165/70 | [19] |
| 3 | 1.76 × 103 | 29 | N/A c | LB d | Tetracycline | XL1-Blue | [13] |
| 4 | 1.00 × 103 | 32→44→33 | N/A c | LB d | N/A c | BT100 | [18] |
| 5 | 1.40 × 102 | 31 | N/A c | 2xYT d | Chloramphenicol Tetracycline Glucose | XL1-Blue | [20] |
| 6 | 1.30 × 102 | 37 | 7.4 | NZY d | Tetracycline | K91 | [10] |
| No. b | Temperature (°C) | pH | Ionic Concentration | ATPase Type | Function | Ref. |
|---|---|---|---|---|---|---|
| 1 | 37~42 | 7~8 | 5 mM Mg2+ | ExeA | Secretion of exotoxins | [54] |
| 2 | 56 | 7.5 | 2.5 mM Mg2+ | Rep c | Initiation of replication | [55] |
| 3 | N/A a | 7.5~8.5 | 3 mM Mg2+ 50 mM KCl | ATPase of Beta vulgaris L. | Transport of solutes | [56] |
| 4 | N/A | 6.5 | 1.5~2.0 mM MgCl2 | Has1p | Remodel macromolecular interaction | [57] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Choi, Y.K.; Han, S.M.; Lee, S.M.; Soh, J.O.; Lee, S.K.; Lee, J.H. Investigation of the Relation between Temperature and M13 Phage Production via ATP Expenditure. Processes 2022, 10, 962. https://doi.org/10.3390/pr10050962
Choi YK, Han SM, Lee SM, Soh JO, Lee SK, Lee JH. Investigation of the Relation between Temperature and M13 Phage Production via ATP Expenditure. Processes. 2022; 10(5):962. https://doi.org/10.3390/pr10050962
Chicago/Turabian StyleChoi, Young Kyun, Sang Min Han, Sang Min Lee, Jeong Ook Soh, Seung Kyu Lee, and Ju Hun Lee. 2022. "Investigation of the Relation between Temperature and M13 Phage Production via ATP Expenditure" Processes 10, no. 5: 962. https://doi.org/10.3390/pr10050962
APA StyleChoi, Y. K., Han, S. M., Lee, S. M., Soh, J. O., Lee, S. K., & Lee, J. H. (2022). Investigation of the Relation between Temperature and M13 Phage Production via ATP Expenditure. Processes, 10(5), 962. https://doi.org/10.3390/pr10050962