
Optimization of Sporulation Conditions for Bacillus subtilis BSNK-5


Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain Culture
2.1.1. Microorganism and Culture Mediums
2.1.2. Culture Conditions
2.2. Measurement of Total Cell Density and Spore Density
2.3. Single Factor Test in Culture Medium
2.3.1. Carbon Source Screening
2.3.2. Nitrogen Source Screening
2.3.3. Inorganic Salts Screening
2.4. Optimize Culture Medium by Response Surface Methodology
2.5. Statistical Analysis
3. Results
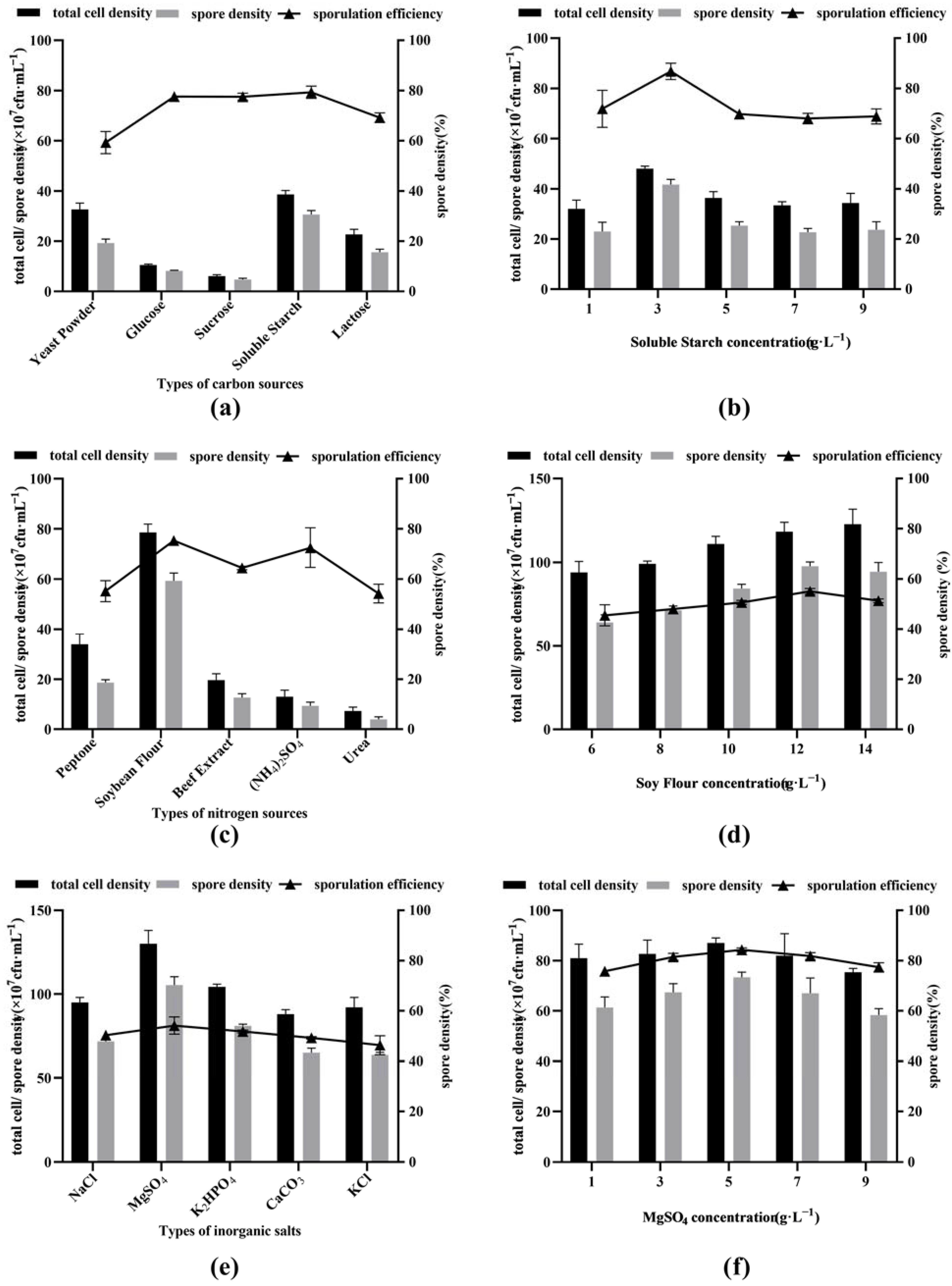
3.1. Screening the Best Carbon Source and Its Optimum Concentration
3.2. Screening the Best Nitrogen Source and Its Optimum Concentration
3.3. Screening the Best Inorganic Salts and Their Optimum Concentrations
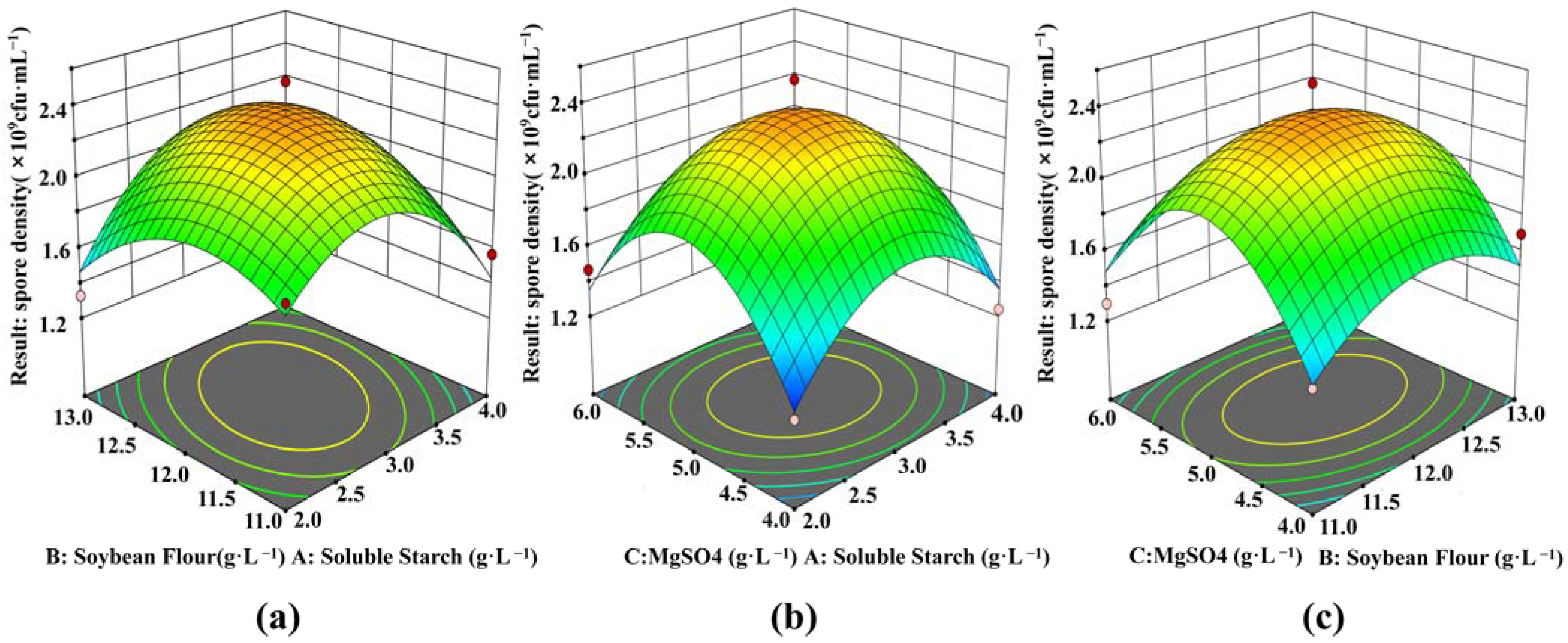
3.4. Optimizing Combination of Medium Composition by RSM
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, Y.M.; Lee, K.S.; Kim, W.M.; Kim, M.; Park, H.O.; Choi, C.W.; Han, J.S.; Park, S.Y.; Lee, K.S. Hydrochloric Acid-Treated Bacillus subtilis Ghosts Induce IL-1 beta, IL-6, and TNF-alpha in Murine Macrophage. Mol. Cell. Toxicol. 2022, 18, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Higgins, D.; Dworkin, J. Recent Progress in Bacillus subtilis Sporulation. FEMS Microbiol. Rev. 2012, 36, 131–148. [Google Scholar] [CrossRef] [Green Version]
- Idiyatov, I.I.; Eroshin, A.I.; Yusupov, S.A.; Zdoroveva, E.V.; Tremasova, A.M. Endophytic Isolates of Bacillus Subtilis: Prospects of Application for Improving the Quality of Food Raw Materials. IOP Conf. Ser. Earth Environ. Sci. 2022, 953, 012024. [Google Scholar] [CrossRef]
- Peng, Q.; Wu, J.; Chen, X.; Qiu, L.; Zhang, J.; Tian, H.; Song, F. Disruption of Two-component System LytSR Affects Forespore Engulfment in Bacillus thuringiensis. Front. Cell Infect. Microbiol. 2017, 7, 468. [Google Scholar] [CrossRef] [Green Version]
- Xing, J.H.; Li, Q.Y.; Zhao, W.; Yang, G.; Zhang, R.R.; Chen, H.L.; Li, Y.; Wan, D.; Zhao, D.D.; Huang, H.B.; et al. Bacillus Subtilis BSH has a Protective Effect on Salmonella Infection by Regulating the Intestinal Flora Structure in Chickens. Microb. Pathog. 2021, 155, 104898. [Google Scholar] [CrossRef]
- Xu, Y.; Yu, Y.; Shen, Y.; Li, Q.; Lan, J.; Wu, Y.; Zhang, R.; Cao, G.; Yang, C. EFfects of Bacillus Subtilis and Bacillus Licheniformis on Growth Performance, Immunity, Short Chain Fatty Acid Production, Antioxidant Capacity, and Cecal Microflora in Broilers. Poult. Sci. 2021, 100, 101358. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wu, Q.; Nie, Y.; Wu, J.; Xu, Y. Construction of Synthetic Microbiota for Reproducible Flavor Compound Metabolism in Chinese Light-Aroma-Type Liquor Produced by Solid-State Fermentation. Appl. Environ. Microbiol. 2019, 85, e03090-18. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Mei, L.H.; Hu, S.; Sheng, Q.; Xu, J.; Wu, H.; Yao, S.J. Screening of Bacillus subtilis Natto from Traditional Japanese Food Natto and Separation of Nattokinase. J. Chem. Eng. Chin. Univ. 2005, 19, 518–522. [Google Scholar] [CrossRef]
- Gu, X.B.; Zheng, Z.M.; Yu, H.Q.; Wang, J.; Liang, F.L.; Liu, R.L. Optimization of Medium Constituents for a Novel Lipopeptide Production by Bacillus subtilis MO-01 by a Response Surface Method. Process Biochem. 2005, 40, 3196–3201. [Google Scholar] [CrossRef]
- Cho, J.H.; Kim, Y.B.; Kim, E.K. Optimization of Culture Media for Bacillus Species by Statistical Experimental Design Methods. Korean J. Chem. Eng. 2009, 26, 754–759. [Google Scholar] [CrossRef]
- Buhr, T.L.; McPherson, D.C.; Gutting, B.W. Analysis of Broth-Cultured Bacillus atrophaeus and Bacillus cereus Spores. J. Appl. Microbiol. 2008, 105, 1604–1613. [Google Scholar] [CrossRef] [PubMed]
- Firdous, S.; Iqbal, S.; Anwar, S. Optimization and Modeling of Glyphosate Biodegradation by a Novel Comamonas odontotermitis P2 Through Response Surface Methodology. Pedosphere 2017, 30, 618–627. [Google Scholar] [CrossRef]
- Korondi, P.; Marchi, M.; Poloni, C. Response Surface Methodology. In Optimization Under Uncertainty with Applications to Aerospace Engineering; Springer: Cham, Switzerland, 2021; pp. 387–409. [Google Scholar]
- Xu, Q.; Shen, Y.; Wang, H.; Zhang, N.; Xu, S.; Zhang, L. Application of Response Surface Methodology to Optimise Extraction of Flavonoids from Fructus Sophorae. Food Chem. 2013, 138, 2122–2129. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.L.; Li, S.Y.; Nie, Y.; Li, Y.; Yuan, C.; Tang, X.M. Screening of Nattokinase Producing Strain and Characterazation of Nattokinase. Biotechnol. Bull. 2015, 31, 161–164. [Google Scholar]
- Chen, Z.M.; Li, Q.; Liu, H.M.; Yu, N.; Xie, T.J.; Yang, M.Y.; Shen, P.; Chen, X.D. Greater Enhancement of Bacillus Subtilis Spore Yields in Submerged Cultures by Optimization of Medium Composition through Statistical Experimental Designs. Appl. Microbiol. Biotechnol. 2010, 85, 1353–1360. [Google Scholar] [CrossRef]
- Gao, Y.X.; Xu, B.; Fan, H.R.; Zhang, M.R.; Zhang, L.J.; Lu, C.; Zhang, N.N.; Fan, B.; Wang, F.Z.; Li, S. 1H NMR-Based Chemometric Metabolomics Characterization of Soymilk Fermented by Bacillus subtilis BSNK-5. Food Res. Int. 2020, 138, 109686. [Google Scholar] [CrossRef] [PubMed]
- Adams, T.T.; Eiteman, M.A.; Adang, M.J. Bacillus thuringiensis Subsp. Kurstaki Spore Production in Batch Culture Using Broiler Litter Extracts as Complex Media. Bioresour. Technol. 1999, 67, 83–87. [Google Scholar] [CrossRef]
- Huang, C.T.; Xu, K.D.; McFeters, G.A.; Stewart, P.S. Spatial Patterns of Alkaline Phosphatase Expression within Bacterial Colonies and Biofilms in Response to Phosphate Starvation. Appl. Environ. Microbiol. 1998, 64, 1526–1531. [Google Scholar] [CrossRef] [Green Version]
- Lynd, L.R.; Weimer, P.J.; van Zyl, W.H.; Pretorius, I.S. Microbial Cellulose Utilization: Fundamentals and Biotechnology. Microbiol. Mol. Biol. Rev. 2002, 66, 739. [Google Scholar] [CrossRef] [Green Version]
- Sonenshein, A.L. Control of Sporulation Initiation in Bacillus subtilis. Curr. Opin. Microbiol. 2000, 3, 561–566. [Google Scholar] [CrossRef]
- Grimshaw, C.E.; Huang, S.; Hanstein, C.G.; Strauch, M.A.; Burbulys, D.; Wang, L.; Hoch, J.A.; Whiteley, J.M. Synergistic Kinetic Interactions between Components of the Phosphorelay Controlling Sporulation in Bacillus subtilis. Biochemistry 1998, 37, 1365–1375. [Google Scholar] [CrossRef] [PubMed]
- Hageman, J.H.; Shankweiler, G.W.; Wall, P.R.; Franich, K.; McCowan, G.W.; Cauble, S.M.; Grajeda, J.; Quinones, C. Single, Chemically Defined Sporulation Medium for Bacillus subtilis: Growth, Sporulation, and Extracellular Protease Production. J. Bacteriol. 1984, 160, 438–441. [Google Scholar] [CrossRef] [Green Version]
- Tavares, M.B.; Souza, R.D.; Luiz, W.B.; Cavalcante, R.C.M.; Casaroli, C.; Martins, E.G.; Ferreira, R.C.C.; Ferreira, L.C.S. Bacillus subtilis Endospores at High Purity and Recovery Yields: Optimization of Growth Conditions and Purification Method. Curr. Microbiol. 2013, 66, 279–285. [Google Scholar] [CrossRef]
- Nierhaus, K. Mg2+, K+, and the Ribosome. J. Bacteriol. 2014, 196, 3817–3819. [Google Scholar] [CrossRef] [Green Version]
- Sreekumar, G.; Krishnan, S. Enhanced Biomass Production Study on Probiotic Bacillus subtilis SK09 by Medium Optimization Using Response Surface Methodology. Afr. J. Biotechnol. 2010, 9, 8078–8084. [Google Scholar]
- Saxena, R.; Singh, R. Contemporaneous Production of Amylase and Protease through CCD Response Surface Methodology by Newly Isolated Bacillus megaterium Strain B69. Enzyme Res. 2014, 2014, 601046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, H.; Cai, X. Optimization of the Extraction of Total Flavonoids from Scutellaria Baicalensis Georgi Using the Response Surface Methodology. J. Food Sci. Technol. 2015, 52, 2336–2343. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Cui, S.W.; Tang, J.; Gu, X. Optimization of Extraction Process of Crude Polysaccharides from Boat-Fruited Sterculia Seeds by Response Surface Methodology. Food Chem. 2007, 105, 1599–1605. [Google Scholar] [CrossRef]
- Li, S.; Zhang, X.; Li, Y.; Tao, L.; Li, T. Optimization of pH Conditions to Improve the Spore Production of Clostridium butyricum NN-2 during Fermentation Process. Arch. Microbiol. 2020, 202, 1251–1256. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Symbol | Variables | Levels (g·L−1) | ||
|---|---|---|---|---|
| −1 | 0 | 1 | ||
| A | Soluble Starch | 2 | 3 | 4 |
| B | Soybean Flour | 11 | 12 | 13 |
| C | MgSO4 | 4 | 5 | 6 |
| Std | Coded Value | Spore Density (×109 CFU·mL−1) | ||
|---|---|---|---|---|
| A | B | C | ||
| 1 | 1 | −1 | 0 | 1.57 |
| 2 | 0 | −1 | −1 | 1.4 |
| 3 | 1 | 0 | −1 | 1.24 |
| 4 | 0 | 1 | −1 | 1.7 |
| 5 | 0 | 0 | 0 | 2.53 |
| 6 | 0 | 1 | 1 | 1.5 |
| 7 | −1 | 0 | 1 | 1.47 |
| 8 | 0 | 0 | 0 | 2.21 |
| 9 | 0 | 0 | 0 | 2.28 |
| 10 | −1 | −1 | 0 | 1.82 |
| 11 | 1 | 1 | 0 | 1.76 |
| 12 | 1 | 0 | 1 | 1.33 |
| 13 | 0 | −1 | 1 | 1.30 |
| 14 | −1 | 1 | 0 | 1.33 |
| 15 | −1 | 0 | −1 | 1.21 |
| Source | Adj SS | df | Adj MS | F-Value | p-Value |
|---|---|---|---|---|---|
| Model | 2.14 | 9 | 0.2378 | 5.95 | 0.0319 * |
| A-Soluble Starch | 0.0006 | 1 | 0.0006 | 0.0153 | 0.9063 |
| B-Soybean Flour C-MgSO4 | 0.0050 | 1 | 0.0050 | 0.1250 | 0.7380 |
| 0.0003 | 1 | 0.0003 | 0.0078 | 0.9330 | |
| AB | 0.1156 | 1 | 0.1156 | 2.89 | 0.1498 |
| AC | 0.0072 | 1 | 0.0072 | 0.1807 | 0.6885 * |
| BC | 0.0025 | 1 | 0.0025 | 0.0625 | 0.8125 * |
| A2 | 0.7189 | 1 | 0.7189 | 17.98 | 0.0082 * |
| B2 | 0.2869 | 1 | 0.2869 | 7.18 | 0.0439 * |
| C2 | 1.27 | 1 | 1.27 | 31.74 | 0.0024 * |
| Residual | 0.1999 | 5 | 0.0400 | ||
| Lack of Fit | 0.1433 | 3 | 0.0478 | 1.69 | 0.3930 |
| Pure Error | 0.0566 | 2 | 0.0283 | ||
| Cor Total | 2.34 | 14 |
| Parameters | Coded Factor Levels | Actual Factor Levels (g·L−1) |
|---|---|---|
| Soluble Starch | 0 | 3 |
| Soybean Flour | 0 | 12 |
| MgSO4 | 0 | 5 |
| Response | Predicted | Actual |
| Spore cell density (×109 CFU·ml−1) | 2.34 | 2.43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, Z.; Hou, L.; Hu, M.; Gao, Y.; Li, D.; Fan, B.; Wang, F.; Li, S. Optimization of Sporulation Conditions for Bacillus subtilis BSNK-5. Processes 2022, 10, 1133. https://doi.org/10.3390/pr10061133
Tian Z, Hou L, Hu M, Gao Y, Li D, Fan B, Wang F, Li S. Optimization of Sporulation Conditions for Bacillus subtilis BSNK-5. Processes. 2022; 10(6):1133. https://doi.org/10.3390/pr10061133
Chicago/Turabian StyleTian, Zhiliang, Lizhen Hou, Miao Hu, Yaxin Gao, Danfeng Li, Bei Fan, Fengzhong Wang, and Shuying Li. 2022. "Optimization of Sporulation Conditions for Bacillus subtilis BSNK-5" Processes 10, no. 6: 1133. https://doi.org/10.3390/pr10061133
APA StyleTian, Z., Hou, L., Hu, M., Gao, Y., Li, D., Fan, B., Wang, F., & Li, S. (2022). Optimization of Sporulation Conditions for Bacillus subtilis BSNK-5. Processes, 10(6), 1133. https://doi.org/10.3390/pr10061133