

Production and Recovery of Ectoine: A Review of Current State and Future Prospects



Abstract
:1. Introduction
1.1. Current State of Ectoine Production
1.2. Intracellular Ectoine Production
2. Extracellular Ectoine Production
Advances of Integrated Approaches for Ectoine Fermentation
3. Recovery and Downstream Processes of Ectoine
3.1. Coupled Cation-Exchange Resin and Crystallization
3.2. Ethanol Crystallization
3.3. Ultrafiltration with Hollow Fibers, Ethanol Precipitation, and Ion-Exchange Chromatography
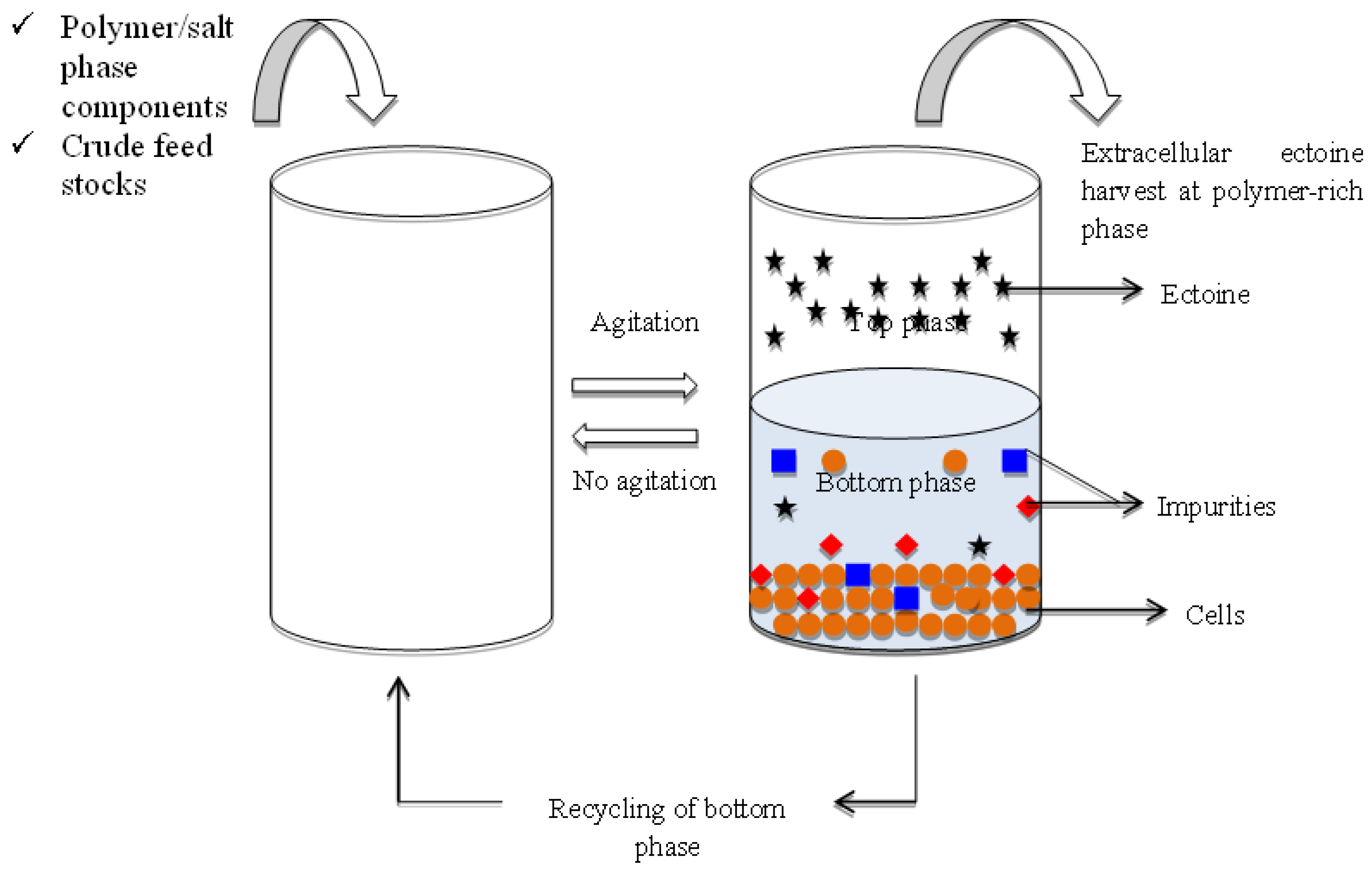
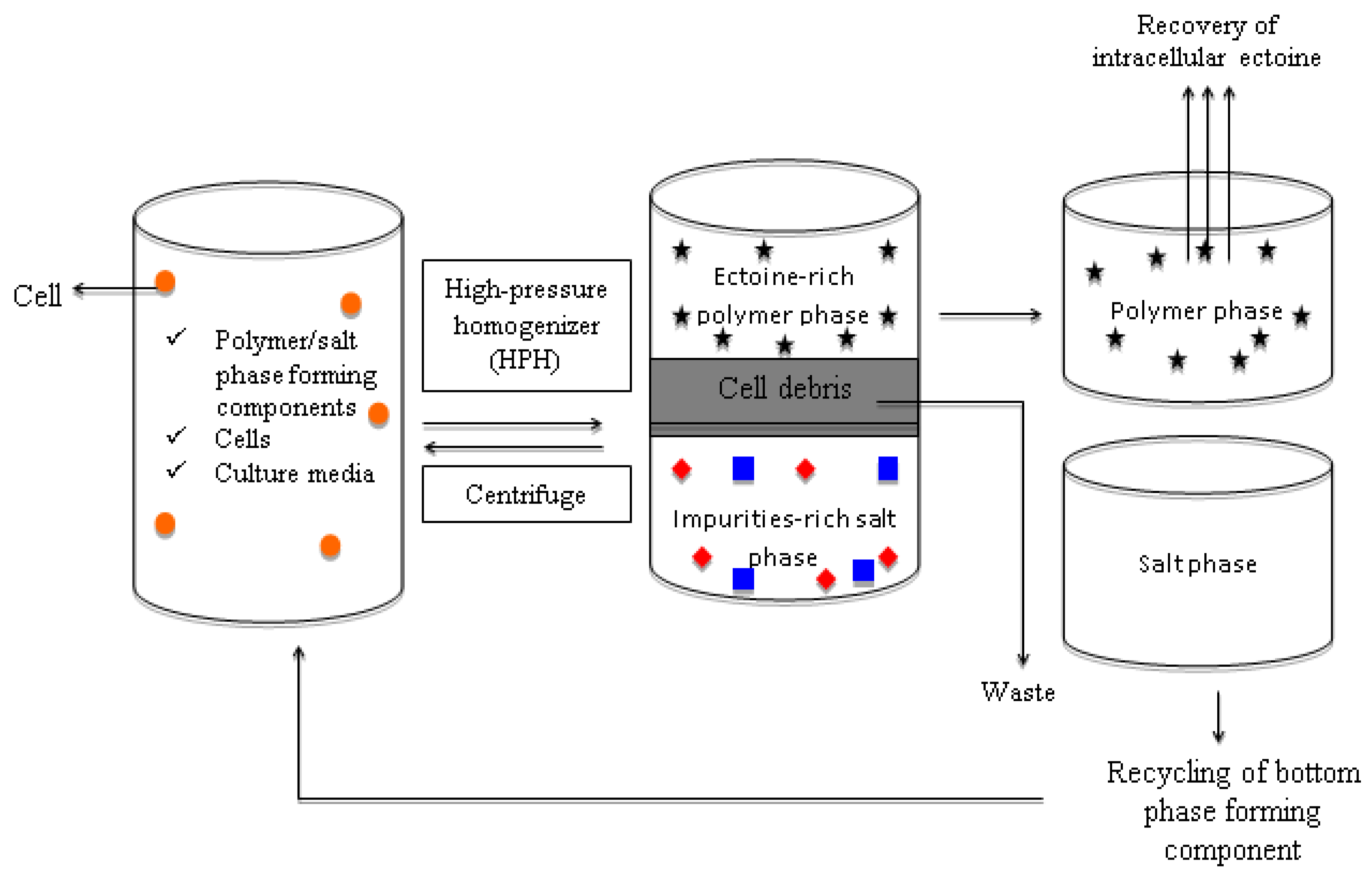
3.4. Applications of Aqueous Biphasic System in Recovery of Ectoine from Microbial Fermentation
4. Overall Perspective and Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shivanand, P.; Mugeraya, G. Halophilic bacteria and their compatible solutes–osmoregulation and potential applications. Curr. Sci. 2011, 100, 1516–1521. [Google Scholar]
- Roberts, M.F. Organic compatible solutes of halotolerant and halophilic microorganisms. Saline Syst. 2005, 1, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, C.; Jebbar, M.; Carrasco, R.; Blanco, C.; Calderón, M.I.; Iglesias-Guerra, F.; Nieto, J.J. Ectoines as compatible solutes and carbon and energy sources for the halophilic bacterium Chromohalobacter salexigens. J. Appl. Microbiol. 2006, 100, 98–107. [Google Scholar] [CrossRef]
- Galinski, E.A.; Pfeiffer, H.P.; Trüper, H.G. 1,4,5,6-Tetrahydro-2-methyl-4-pyrimidinecarboxylic acid: A novel cyclic amino acid from halophilic phototrophic bacteria of the genus Ectothiorhodospira. Eur. J. Biochem. 1985, 149, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Galinski, E.A. Osmoadaptation in bacteria. Adv. Microb. Physiol. 1995, 37, 273–328. [Google Scholar]
- Louis, P.; Galinski, E.A. Characterization of genes for the biosynthesis of the compatible solute ectoine from Marinococcus halophilus and osmoregulated expression in Escherichia coli. Microbiology 1997, 143, 1141–1149. [Google Scholar] [CrossRef] [Green Version]
- He, J.; Huang, X.; Gu, L.; Jiang, J.; Li, S. Cloning of the ectoine biosynthesis gene ectABC from Halomonas sp. BYS-1 and salt stressed expression in Escherichia coli. Wei Sheng Wu Xue Bao Acta Microbiol. Sin. 2006, 46, 28–32. [Google Scholar]
- Becker, J.; Schafer, R.; Kohlstedt, M.; Harder, B.J.; Borchert, N.S.; Stoveken, N.; Bremer, E.; Wittmann, C. Systems metabolic engineering of Corynebacterium glutamicum for production of the chemical chaperone ectoine. Microb. Cell Factories 2013, 12, 110. [Google Scholar] [CrossRef] [Green Version]
- Bestvater, T.; Louis, P.; Galinski, E.A. Heterologous ectoine production in Escherichia coli: By-passing the metabolic bottle-neck. Saline Syst. 2008, 4, 12. [Google Scholar] [CrossRef] [Green Version]
- Bursy, J.; Pierik, A.J.; Pica, N.; Bremer, E. Osmotically induced synthesis of the compatible solute hydroxyectoine is mediated by an evolutionarily conserved ectoine hydroxylase. J. Biol. Chem. 2007, 282, 31147–31155. [Google Scholar] [CrossRef] [Green Version]
- Oren, A. Bioenergetic aspects of halophilism. Microbiol. Mol. Biol. Rev. 1999, 63, 334–348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastor, J.M.; Salvador, M.; Argandoña, M.; Bernal, V.; Reina-Bueno, M.; Csonka, L.N.; Iborra, J.L.; Vargas, C.; Nieto, J.J.; Cánovas, M.J. Ectoines in cell stress protection: Uses and biotechnological production. Biotechnol. Adv. 2010, 28, 782–801. [Google Scholar] [CrossRef] [PubMed]
- Lippert, K.; Galinski, E.A. Enzyme stabilization be ectoine-type compatible solutes: Protection against heating, freezing and drying. Appl. Microbiol. Biotechnol. 1992, 37, 61–65. [Google Scholar] [CrossRef]
- Kolp, S.; Pietsch, M.; Galinski, E.A.; Gütschow, M.J. Compatible solutes as protectants for zymogens against proteolysis. Biochim. Biophys. Acta BBA Proteins Proteom. 2006, 1764, 1234–1242. [Google Scholar] [CrossRef] [PubMed]
- Göller, K.; Galinski, E.A. Protection of a model enzyme (lactate dehydrogenase) against heat, urea and freeze-thaw treatment by compatible solute additives. J. Mol. Catal. B Enzym. 1999, 7, 37–45. [Google Scholar] [CrossRef]
- Khmelenina, V.N.; Mustakhimov, I.I.; Reshetnikov, A.S.; Kalyuzhnaya, M.G.; Trotsenko, Y.A. Genetic and biochemical aspects of ectoine biosynthesis in moderately halophilic and halotolerant methylotrophic bacteria. J. Agric. Biol. Sci. 2010, 5, 446–458. [Google Scholar] [CrossRef] [Green Version]
- Carmona-Martínez, A.A.; Marcos-Rodrigo, E.; Bordel, S.; Marín, D.; Herrero-Lobo, R.; García-Encina, P.A.; Muñoz, R. Elucidating the key environmental parameters during the production of ectoines from biogas by mixed methanotrophic consortia. J. Environ. Manag. 2021, 298, 113462. [Google Scholar] [CrossRef] [PubMed]
- Cantera, S.; Phandanouvong-Lozano, V.; Pascual, C.; García-Encina, P.A.; Lebrero, R.; Hay, A.; Muñoz, R. A systematic comparison of ectoine production from upgraded biogas using Methylomicrobium alcaliphilum and a mixed haloalkaliphilic consortium. Waste Manag. 2020, 102, 773–781. [Google Scholar] [CrossRef]
- Widderich, N.; Czech, L.; Elling, F.J.; Konneke, M.; Stoveken, N.; Pittelkow, M.; Riclea, R.; Dickschat, J.S.; Heider, J.; Bremer, E. Strangers in the archaeal world: Osmostress-responsive biosynthesis of ectoine and hydroxyectoine by the marine thaumarchaeon Nitrosopumilus maritimus. Environ. Microbiol. 2016, 18, 1227–1248. [Google Scholar] [CrossRef]
- Chen, Q.; Zhang, L.; Li, X.; Liu, S.; Li, D. Poly-β-hydroxybutyrate/ectoine co-production by ectoine-excreting strain Halomonas salina. Process Biochem. 2014, 49, 33–37. [Google Scholar] [CrossRef]
- Van-Thuoc, D.; Guzmán, H.; Quillaguamán, J.; Hatti-Kaul, R. High productivity of ectoines by Halomonas boliviensis using a combined two-step fed-batch culture and milking process. J. Biotechnol. 2010, 147, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Schwibbert, K.; Marin-Sanguino, A.; Bagyan, I.; Heidrich, G.; Lentzen, G.; Seitz, H.; Rampp, M.; Schuster, S.C.; Klenk, H.P.; Pfeiffer, F. A blueprint of ectoine metabolism from the genome of the industrial producer Halomonas elongata DSM 2581T. Environ. Microbiol. 2011, 13, 1973–1994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widderich, N.; Hoppner, A.; Pittelkow, M.; Heider, J.; Smits, S.H.; Bremer, E. Biochemical properties of ectoine hydroxylases from extremophiles and their wider taxonomic distribution among microorganisms. PLoS ONE 2014, 9, e93809. [Google Scholar] [CrossRef]
- Fenizia, S.; Thume, K.; Wirgenings, M.; Pohnert, G. Ectoine from Bacterial and Algal Origin is a Compatible Solute in Microalgae. Mar. Drugs 2020, 18, 42. [Google Scholar] [CrossRef] [Green Version]
- Pérez, V.; Moltó, J.L.; Lebrero, R.; Muñoz, R. Ectoine production from biogas: A sensitivity analysis. Effect of local commodity prices, economy of scale, market trends and biotechnological limitations. J. Clean. Prod. 2022, 369, 133440. [Google Scholar] [CrossRef]
- Rodero, M.D.R.; Herrero-Lobo, R.; Pérez, V.; Muñoz, R. Influence of operational conditions on the performance of biogas bioconversion into ectoines in pilot bubble column bioreactors. Bioresour. Technol. 2022, 358, 127398. [Google Scholar] [CrossRef]
- Kang, J.Y.; Lee, B.; Kim, J.A.; Kim, M.-S.; Kim, C.H. Identification and characterization of an ectoine biosynthesis gene cluster from Aestuariispira ectoiniformans sp. nov., isolated from seawater. Microbiol. Res. 2022, 254, 126898. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, L. Ectoine improves yield of biodiesel catalyzed by immobilized lipase. J. Mol. Catal. B Enzym. 2010, 62, 90–95. [Google Scholar] [CrossRef]
- Bethlehem, L.; van Echten-Deckert, G. Ectoines as novel anti-inflammatory and tissue protective lead compounds with special focus on inflammatory bowel disease and lung inflammation. Pharmacol. Res. 2021, 164, 105389. [Google Scholar] [CrossRef]
- Buenger, J.; Driller, H. Ectoin: An effective natural substance to prevent UVA-induced premature photoaging. Ski. Pharmacol. Physiol. 2004, 17, 232–237. [Google Scholar] [CrossRef]
- Graf, R.; Anzali, S.; Buenger, J.; Pfluecker, F.; Driller, H. The multifunctional role of ectoine as a natural cell protectant. Clin. Dermatol. 2008, 26, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Marini, A.; Reinelt, K.; Krutmann, J.; Bilstein, A. Ectoine-containing cream in the treatment of mild to moderate atopic dermatitis: A randomised, comparator-controlled, intra-individual double-blind, multi-center trial. Ski. Pharmacol. Physiol. 2014, 27, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Bow, J.R.; Sonoki, Y.; Uchiyama, M.; Dauskardt, R.H. Ectoine disperses keratin and alters hydration kinetics in stratum corneum. Biochem. Biophys. Rep. 2021, 28, 101134. [Google Scholar] [CrossRef] [PubMed]
- Sydlik, U.; Gallitz, I.; Albrecht, C.; Abel, J.; Krutmann, J.; Unfried, K. The compatible solute ectoine protects against nanoparticle-induced neutrophilic lung inflammation. Am. J. Respir. Crit. Care Med. 2009, 180, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Wedeking, A.; Buttner, R.; Kalff, J.C.; Tolba, R.H.; van Echten-Deckert, G. A natural tetrahydropyrimidine protects small bowel from cold ischemia and subsequent warm in vitro reperfusion injury. Pathobiol. J. Immunopathol. Mol. Cell. Biol. 2009, 76, 212–220. [Google Scholar] [CrossRef]
- Kanapathipillai, M.; Lentzen, G.; Sierks, M.; Park, C.B. Ectoine and hydroxyectoine inhibit aggregation and neurotoxicity of Alzheimer’s beta-amyloid. FEBS Lett. 2005, 579, 4775–4780. [Google Scholar] [CrossRef] [Green Version]
- Harishchandra, R.K.; Sachan, A.K.; Kerth, A.; Lentzen, G.; Neuhaus, T.; Galla, H.J. Compatible solutes: Ectoine and hydroxyectoine improve functional nanostructures in artificial lung surfactants. Biochim. Biophys. Acta 2011, 1808, 2830–2840. [Google Scholar] [CrossRef] [Green Version]
- Bownik, A.; Stepniewska, Z. Protective effects of bacterial osmoprotectant ectoine on bovine erythrocytes subjected to staphylococcal alpha-haemolysin. Toxicon Off. J. Int. Soc. Toxinology 2015, 99, 130–135. [Google Scholar] [CrossRef]
- Unfried, K.; Kroker, M.; Autengruber, A.; Gotić, M.; Sydlik, U. The Compatible Solute Ectoine Reduces the Exacerbating Effect of Environmental Model Particles on the Immune Response of the Airways. J. Allergy 2014, 2014, 7. [Google Scholar] [CrossRef] [Green Version]
- Yao, C.-L.; Lin, Y.-M.; Mohamed, M.S.; Chen, J.-H. Inhibitory effect of ectoine on melanogenesis in B16-F0 and A2058 melanoma cell lines. Biochem. Eng. J. 2013, 78, 163–169. [Google Scholar] [CrossRef]
- Anzali, S.; Von Heydebreck, A.; Herget, T. Elucidation of the anti-aging effects of ectoine using cdna microarray analysis and signaling pathway evaluation. Int. J. Cosmet. Sci. 2010, 32, 319. [Google Scholar] [CrossRef]
- Werkhäuser, N.; Bilstein, A.; Sonnemann, U. Treatment of Allergic Rhinitis with Ectoine Containing Nasal Spray and Eye Drops in Comparison with Azelastine Containing Nasal Spray and Eye Drops or with Cromoglycic Acid Containing Nasal Spray. J. Allergy 2014, 2014, 13. [Google Scholar] [CrossRef] [Green Version]
- Sonnemann, U.; Scherner, O.; Werkhäuser, N. Treatment of Rhinitis Sicca Anterior with Ectoine Containing Nasal Spray. J. Allergy 2014, 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- Eichel, A.; Wittig, J.; Shah-Hosseini, K.; Mosges, R. A prospective, controlled study of SNS01 (ectoine nasal spray) compared to BNO-101 (phytotherapeutic dragees) in patients with acute rhinosinusitis. Curr. Med. Res. Opin. 2013, 29, 739–746. [Google Scholar] [CrossRef]
- Wan, P.K.; Lan, J.C.-W.; Chen, P.-W.; Tan, J.S.; Ng, H.S. Recovery of intracellular ectoine from Halomonas salina cells with poly(propylene) glycol/salt aqueous biphasic system. J. Taiwan Inst. Chem. Eng. 2018, 82, 28–32. [Google Scholar] [CrossRef]
- Zhang, L.H.; Lang, Y.J.; Nagata, S. Efficient production of ectoine using ectoine-excreting strain. Extrem. Life Under Extrem. Cond. 2009, 13, 717–724. [Google Scholar] [CrossRef]
- Chen, P.-W.; Cui, Z.-Y.; Ng, H.S.; Chi-Wei Lan, J. Exploring the additive bio-agent impacts upon ectoine production by Halomonas salina DSM5928T using corn steep liquor and soybean hydrolysate as nutrient supplement. J. Biosci. Bioeng. 2020, 130, 195–199. [Google Scholar] [CrossRef]
- Onraedt, A.E.; Walcarius, B.A.; Soetaert, W.K.; Vandamme, E.J. Optimization of ectoine synthesis through fed-batch fermentation of Brevibacterium epidermis. Biotechnol. Prog. 2005, 21, 1206–1212. [Google Scholar] [CrossRef]
- Fallet, C.; Rohe, P.; Franco-Lara, E. Process optimization of the integrated synthesis and secretion of ectoine and hydroxyectoine under hyper/hypo-osmotic stress. Biotechnol. Bioeng. 2010, 107, 124–133. [Google Scholar] [CrossRef]
- Sauer, T.; Galinski, E.A. Bacterial milking: A novel bioprocess for production of compatible solutes. Biotechnol. Bioeng. 1998, 57, 306–313. [Google Scholar] [CrossRef]
- Guzman, H.; Van-Thuoc, D.; Martin, J.; Hatti-Kaul, R.; Quillaguaman, J. A process for the production of ectoine and poly(3-hydroxybutyrate) by Halomonas boliviensis. Appl. Microbiol. Biotechnol. 2009, 84, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Lang, Y.J.; Bai, L.; Ren, Y.N.; Zhang, L.H.; Nagata, S. Production of ectoine through a combined process that uses both growing and resting cells of Halomonas salina DSM 5928T. Extrem. Life Under Extrem. Cond. 2011, 15, 303–310. [Google Scholar] [CrossRef] [PubMed]
- Jiang, A.; Song, Y.; You, J.; Zhang, X.; Xu, M.; Rao, Z. High-yield ectoine production in engineered Corynebacterium glutamicum by fine metabolic regulation via plug-in repressor library. Bioresour. Technol. 2022, 362, 127802. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Chen, J.; Wang, Y.; Du, G.; Kang, Z. Engineering Escherichia coli for high-yield production of ectoine. Green Chem. Eng. 2021, in press. [Google Scholar] [CrossRef]
- Anbu Rajan, L.; Joseph, T.C.; Thampuran, N.; James, R.; Ashok Kumar, K.; Viswanathan, C.; Bansal, K.C. Cloning and heterologous expression of ectoine biosynthesis genes from Bacillus halodurans in Escherichia coli. Biotechnol. Lett. 2008, 30, 1403–1407. [Google Scholar] [CrossRef]
- Schubert, T.; Maskow, T.; Benndorf, D.; Harms, H.; Breuer, U. Continuous synthesis and excretion of the compatible solute ectoine by a transgenic, nonhalophilic bacterium. Appl. Environ. Microbiol. 2007, 73, 3343–3347. [Google Scholar] [CrossRef] [Green Version]
- He, Y.-Z.; Gong, J.; Yu, H.-Y.; Tao, Y.; Zhang, S.; Dong, Z.-Y. High production of ectoine from aspartate and glycerol by use of whole-cell biocatalysis in recombinant Escherichia coli. Microb. Cell Factories 2015, 14, 55. [Google Scholar] [CrossRef] [Green Version]
- Tanimura, K.; Nakayama, H.; Tanaka, T.; Kondo, A. Ectoine production from lignocellulosic biomass-derived sugars by engineered Halomonas elongata. Bioresour. Technol. 2013, 142, 523–529. [Google Scholar] [CrossRef]
- Tanimura, K.; Matsumoto, T.; Nakayama, H.; Tanaka, T.; Kondo, A. Improvement of ectoine productivity by using sugar transporter-overexpressing Halomonas elongata. Enzym. Microb. Technol. 2016, 89, 63–68. [Google Scholar] [CrossRef]
- Ning, Y.; Wu, X.; Zhang, C.; Xu, Q.; Chen, N.; Xie, X. Pathway construction and metabolic engineering for fermentative production of ectoine in Escherichia coli. Metab. Eng. 2016, 36, 10–18. [Google Scholar] [CrossRef]
- Wei, Y.H.; Yuan, F.W.; Chen, W.C.; Chen, S.Y. Production and characterization of ectoine by Marinococcus sp. ECT1 isolated from a high-salinity environment. J. Biosci. Bioeng. 2011, 111, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Simental-Martínez, J.; Montalvo-Hernández, B.; Rito-Palomares, M.; Benavides, J. Application of Aqueous Two-Phase Systems for the Recovery of Bioactive Low-Molecular Weight Compounds. Sep. Sci. Technol. 2014, 49, 1872–1882. [Google Scholar] [CrossRef]
- Ng, H.S.; Wang, C.-C.; Tan, J.S.; Lan, J.C.-W. Primary recovery of recombinant human serum albumin from transgenic Oryza sativa with a single-step aqueous biphasic system. J. Taiwan Inst. Chem. Eng. 2018, 84, 60–66. [Google Scholar] [CrossRef]
- Gu, B.H.; Zheng, P.; Yan, Q.; Liu, W. Aqueous two-phase system: An alternative process for recovery of succinic acid from fermentation broth. Sep. Purif. Technol. 2014, 138, 47–54. [Google Scholar] [CrossRef]
- Campos-Pinto, I.; Espitia-Saloma, E.; Rosa, S.A.S.L.; Rito-Palomares, M.; Aguilar, O.; Arévalo-Rodríguez, M.; Aires-Barros, M.R.; Azevedo, A.M. Integration of cell harvest with affinity-enhanced purification of monoclonal antibodies using aqueous two-phase systems with a dual tag ligand. Sep. Purif. Technol. 2017, 173, 129–134. [Google Scholar] [CrossRef]
- Tan, Z.; Li, F.; Xu, X. Isolation and purification of aloe anthraquinones based on an ionic liquid/salt aqueous two-phase system. Sep. Purif. Technol. 2012, 98, 150–157. [Google Scholar] [CrossRef]
- Asenjo, J.A.; Andrews, B.A. Aqueous two-phase systems for protein separation: A perspective. J. Chromatogr. A 2011, 1218, 8826–8835. [Google Scholar] [CrossRef]
- Aziz, N.F.H.A.; Abbasiliasi, S.; Ng, H.S.; Phapugrangkul, P.; Bakar, M.H.A.; Tam, Y.J.; Tan, J.S. Purification of β-mannanase derived from Bacillus subtilis ATCC 11774 using ionic liquid as adjuvant in aqueous two-phase system. J. Chromatogr. B 2017, 1055, 104–112. [Google Scholar] [CrossRef]
- Ruiz, C.S.; van den Berg, C.; Wijffels, R.H.; Eppink, M.H.M. Rubisco separation using biocompatible aqueous two-phase systems. Sep. Purif. Technol. 2018, 196, 254–261. [Google Scholar] [CrossRef]
- Ng, H.S.; Ooi, C.W.; Mokhtar, M.N.; Show, P.L.; Ariff, A.; Tan, J.S.; Ng, E.-P.; Ling, T.C. Extractive bioconversion of cyclodextrins by Bacillus cereus cyclodextrin glycosyltransferase in aqueous two-phase system. Bioresour. Technol. 2013, 142, 723–726. [Google Scholar] [CrossRef] [Green Version]
- Ng, H.S.; Wan, P.K.; Ng, T.-C.; Lan, J.C.-W. Primary purification of intracellular Halomonas salina ectoine using ionic liquids-based aqueous biphasic system. J. Biosci. Bioeng. 2020, 130, 200–204. [Google Scholar] [CrossRef] [PubMed]
- Ooi, C.W.; Hii, S.L.; Kamal, S.M.M.; Ariff, A.; Ling, T.C. Extractive fermentation using aqueous two-phase systems for integrated production and purification of extracellular lipase derived from Burkholderia pseudomallei. Process Biochem. 2011, 46, 68–73. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Phylum | Microorganisms | ectD is Outside of ect Gene Cluster | ectD is within the ect Gene Cluster |
|---|---|---|---|
| Actinobacteria | Brachybacterium faecium DSM 4810 Brevibacterium album DSM 18261 | √ √ | |
| Gordonia terrae NBRC 100016 | √ | ||
| Kytococcus sedentarius DSM 20547 | √ | ||
| Streptomyces coelicolor A3 (2) | √ | ||
| Firmicutes | Bacillus halodurans DSM 497T Bacillus pseudofirmus OF4 | √ √ | |
| Halobacillus halophilus DSMZ 2266T | √ | ||
| Marinococcus halotolerans DSM 16375 Marinococcus sp. M52 Virgibacillus salexigens DSM 11438 | √ √ | √ | |
| Proteobacteria | Achromobacter xylosidans A8 | √ | |
| Acidiphilum crytum JF-5 | √ | ||
| Cellvibrio japonicus Ueda107 Chromohalobacter salexigens DSM 3043 | √ | √ | |
| Halomonas boliviensis Halomonas elongata DSM 2581 | √ √ | ||
| Nitrosococcus oceani ATCC 19707 Pseudomonas stutzeri A1501 Roseobacter sp. MED193 Vibrio cholerae O395 | √ √ | √ √ | |
| Archaea | Nitrosopumilus sp. AR2 Candidatus | √ |
| Protective Effects against/on: | Treatment Target | Ref. |
|---|---|---|
| Nanoparticle | Neutrophilic lung inflammation | [34] |
| Small bowel | Cold ischemia, reperfusion injury | [35] |
| Aggregation and neurotoxicity of Alzheimer’s β-amyloid | Alzheimer’s disease | [36] |
| Improvement on functional nanostructures | Artificial lung surfactants | [37] |
| Staphylococcal α-hemolysin | Bovine erythrocytes | [38] |
| Exacerbating effect of environmental model particles | Immune response of the airways | [39] |
| Melanogens | Melanogenesis | [40] |
| Ultraviolet A, anti-aging | Skin protection | [23,41] |
| Mild to moderate atopic dermatitis | Allergy | [25] |
| Transepidermal water loss | Skin dehydration | [24] |
| In vivo protection | Allergic rhinitis, Acute rhinosinusitis, rhinitis sicca anterior | [42,43] |
| Microorganisms | Fermentation Setting | Intracellular /Extracellular Ectoine | Carbon Source | Concentration of NaCl (M) | Maximum Production Yield (mg/gcdw) | Maximum Productivity (g/L/day) | Ref. |
|---|---|---|---|---|---|---|---|
| Brevibacterium epidermis DSM 20659 | Fed-batch | Intracellular | Sodium glutamate | 1.0 | 160 | 2 | [48] |
| Chromohalobacter salexigens | Continuous cell retention | Intracellular | Glucose | 1.8 | 540 | 32.5 | [49] |
| Halomonas elongata DSM 142T | Batch | Intracellular | Glucose, ammonium chloride | 2.6 | 155 | 5.3 | [50] |
| Halomonas boliviensis DSM 15516 | Two-step fed-batch | Intracellular | Glucose and sodium glutamate (c) | 0.17 (a) 2.13 (b) | 170 | 3.4 | [51] |
| Halomonas salina DSM 5928 | Batch | Both | Sodium glutamate | 0.5 | 245.9 | 7.9 | [46] |
| Halomonas salina DSM 5928T | Combined Batch | Both | Sodium glutamate | 0.5 | 140 | 7.75 | [52] |
| Halomonas boliviensis LC1T | Two-step fed-batch, batch | Intracellular | Glucose and sodium glutamate (c) | 0.77 (a) 2.55 (b) | 154 | 9.1 | [21] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ng, H.S.; Wan, P.-K.; Kondo, A.; Chang, J.-S.; Lan, J.C.-W. Production and Recovery of Ectoine: A Review of Current State and Future Prospects. Processes 2023, 11, 339. https://doi.org/10.3390/pr11020339
Ng HS, Wan P-K, Kondo A, Chang J-S, Lan JC-W. Production and Recovery of Ectoine: A Review of Current State and Future Prospects. Processes. 2023; 11(2):339. https://doi.org/10.3390/pr11020339
Chicago/Turabian StyleNg, Hui Suan, Pik-Ki Wan, Akihiko Kondo, Jo-Shu Chang, and John Chi-Wei Lan. 2023. "Production and Recovery of Ectoine: A Review of Current State and Future Prospects" Processes 11, no. 2: 339. https://doi.org/10.3390/pr11020339
APA StyleNg, H. S., Wan, P. -K., Kondo, A., Chang, J. -S., & Lan, J. C. -W. (2023). Production and Recovery of Ectoine: A Review of Current State and Future Prospects. Processes, 11(2), 339. https://doi.org/10.3390/pr11020339