1. Introduction
Tuberculosis (TB) has killed more people than any other infectious disease and continues to infect more people today than at any other time in history [
1]. In 2015, 10.4 million people were infected with Mycobacterium tuberculosis (Mtb), and 1.8 million died from TB disease [
2]. An individual inoculated with Mtb may experience a range of outcomes. The Mtb bacteria may be immediately destroyed by the host’s immune response, the immune response may isolate bacteria into granulomas where the infection persists in a latent state, or the bacteria may proliferate and manifest as active TB disease if the initial infection is not controlled by the immune response. A majority of people infected with Mtb have a clinically latent infection in which they do not show any symptoms of the infection. These individuals serve as a reservoir for the bacteria, and if their immune response system is compromised in such a way to trigger the penetration of the granulomas by active bacteria and formation of TB cavities, the infection may transition from latent to active TB disease. The major risk factors for activation of TB after an extended latent period include contact with an infectious TB patient, HIV co-infection, initiation of an anti-tumor necrosis factor (TNF) treatment, silicosis, and diabetes [
3]. About 5–10% of latent infections undergo granuloma activation and progress to active TB [
4]. However, the mechanism for the activation of latent TB is still unclear. Improved understanding of the triggers and dynamics of this transition could be useful for designing new therapies to prevent the activation of latent TB.
TB infection starts when infectious droplets containing Mtb reach the respiratory tract of an individual. After reaching the lung tissue, the Mtb is ingested by the resident alveolar macrophages. The host cellular immune response starts with the secretion of cytokines, such as interleukin 12 (IL-12) and tumor necrosis factor alpha (TNF-
), and chemokines that recruit the immune cells to the site of infection [
5] to form a compact cluster of immune cells, known as a granuloma. Latent infection is characterized by granuloma formation and steady state maintenance (
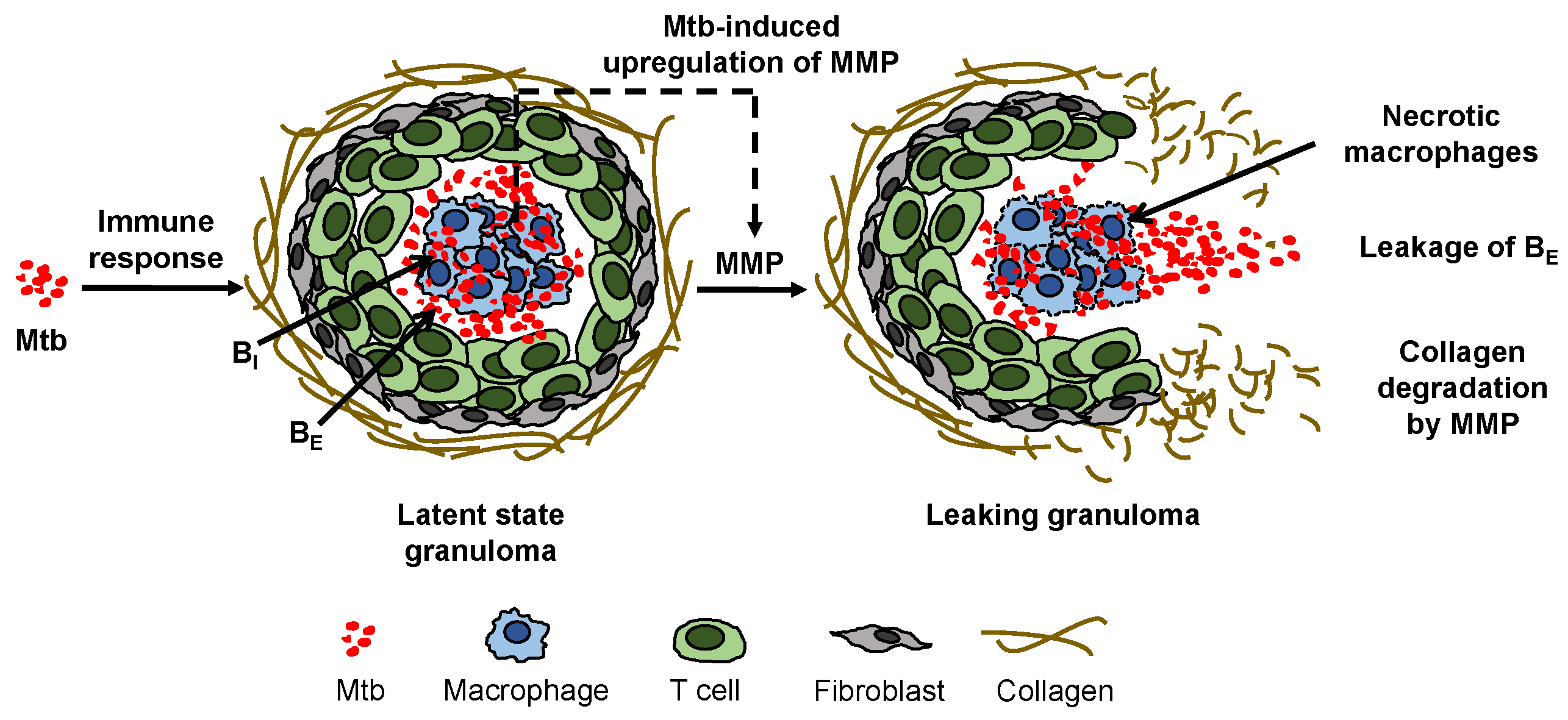
Figure 1). A granuloma is mostly comprised of an organized aggregate of blood-derived macrophages that ingest and contain the bacteria, differentiated macrophages, and T cells along with other cells such as neutrophils, multinucleated giant cells, dendritic cells, B cells, natural killer cells, fibroblasts, and cells that secrete extracellular matrix components. The exterior surface of the granuloma is composed largely of collagen fibers (
Figure 1). The granuloma acts as a microenvironment that walls off the bacteria from the rest of the body to control the infection [
1].
Collagen fibers provide tensile strength to the lungs and to granulomas and are highly resistant to enzymatic degradation. Only collagenolytic proteases such as matrix metalloproteinases (MMPs) are able to break the collagen fibers that encapsulate a granuloma. MMPs are a family of proteolytic enzymes that degrade the components of the extracellular matrix and are critical for matrix remodeling [
6]. MMPs are typically regulated by the complementary class of inhibitors called tissue inhibitors of metalloproteinases (TIMPs). MMPs have been implicated in the activation of latent tuberculosis infections [
7,
8,
9]. From the MMP family of proteases, MMP-1 specifically degrades type-1 collagen and drives the remodeling of pulmonary tissue in TB [
7]. Experimental data showed that TB activation involved a dysregulation in the balance of MMP-1 and its inhibitor TIMP-1 [
7,
10]. Direct infection of macrophages induced gene expression and secretion of MMP-1 along with a few other MMPs [
7]. Additionally, pro-inflammatory cytokines increased MMP secretion from stromal cells such as epithelial cells, fibroblasts, and astrocytes, while there was no compensatory increase in the production of TIMPs to regulate the MMP levels, thus causing an increase in collagen degradation [
7]. Measurements from both human plasma samples [
10] and sputum [
7] showed that the concentrations of MMP-1 were elevated in patients with active TB compared to latent TB patients or non-infected control subjects, and the levels of the associated inhibitor TIMP-1 were either decreased [
7] or changed insignificantly [
10] in the active TB cohorts. In [
7] MMP-1, degradation of lung collagen in TB was confirmed using a transgenic TB mouse model that overexpressed human MMP-1. As MMP-1 is the dominant MMP in granuloma degradation, we simply refer to MMP-1 as MMP in the rest of this article. A surplus concentration of MMP cleaves the collagen envelope of granulomas and eventually leads to the leakage of bacteria into the airways [
8] (
Figure 1); from the airways, the bacteria can spread into other regions of the lung and to the rest of the patient’s body in an active infection.
The animal models that are most commonly used to study TB do not develop lung pathology exactly the same as in humans [
11,
12]. Mouse models are useful for studying the infection stage of TB but not the long term latency or reactivation from the latent state because most mouse models do not form human-like granulomas [
7]. The rabbit model used in [
10] forms necrotic, leaking cavity structures characteristic of active TB after disruption of granulomas. These cavities in rabbits are consistent with the structure observed in humans, providing a valuable animal model for mechanistic insight into cavity formation due to MMP/TIMP imbalance [
10]. However, the one month time scale for the formation of active cavities from initial TB infection in the rabbit model is a much faster time scale than the typical cavity formation in human TB infection. Human studies to enhance mechanistic understanding of the untreated latent to active transition cannot be conducted ethically without applying the current standard of care (pharmaceutical interventions), disrupting the cavity formation and progress over time. Additionally, most animal models require that infected animals be sacrificed for invasive lung tissue sample collected to permit observation of the interaction between Mtb and host structures. Thus, conclusions have to be drawn at minimal discrete time points without providing much insight into the dynamic processes. Another surrogate model system is needed to overcome these challenges for understanding the dynamics of MMP dysregulation that can lead to TB cavity formation from latent granulomas and reactivation to TB disease from latent disease.
In lieu of biological experiments, computational models can be used to test possible mechanisms for triggering the switch from latent to active TB infection as well as to study the dynamic process. Mathematical models are useful tools for inexpensively conducting in silico experiments with multiple interacting factors and for testing hypotheses. We developed a mathematical model in this study to probe triggers for inducing the latent to active transition. Several computational and mathematical models have been developed to describe the granuloma formation stages of TB in response to an initial infection [
13,
14,
15,
16]. Another model was used to explore activation of TB due to a pharmaceutical intervention [
17]. No mathematical model has been published addressing the impact of MMP dysregulation or the dynamics of this process on the biological network of cells within a granuloma during TB. The model developed here builds on an existing model of the immune response to Mtb [
13] (referred to as the “immune response model” henceforth) by extending this model to explicitly consider dynamic regulation of MMP-1. We also share our open-source Python codes for ease of continued development by other computational researchers and by expanding the horizons of use of the models for further in silico experiments by collaborators and other scientists not necessarily trained in high performance computing.
The immune response model is able to simulate three physiologically-relevant regimes based on parameter values: (i) immediate clearance of Mtb; (ii) a mild initial infection followed by long-term latent TB; and (iii) an initial uncontrolled active infection [
13]. Here, a novel model for MMP dynamics, collagen degradation, and bacterial leakage is added to the immune response model.
Figure 1 illustrates the mechanisms we aim to capture in the model. The upregulation of MMP by Mtb drives the degradation of the granuloma envelope, which allows Mtb to leak out of the granuloma to the surroundings. Using the model that incorporates the MMP dynamics, we investigated conditions under which the biological system can be perturbed to switch to active infection after a steady latent infection has been established.
Section 2 details the equations used to define the mathematical model.
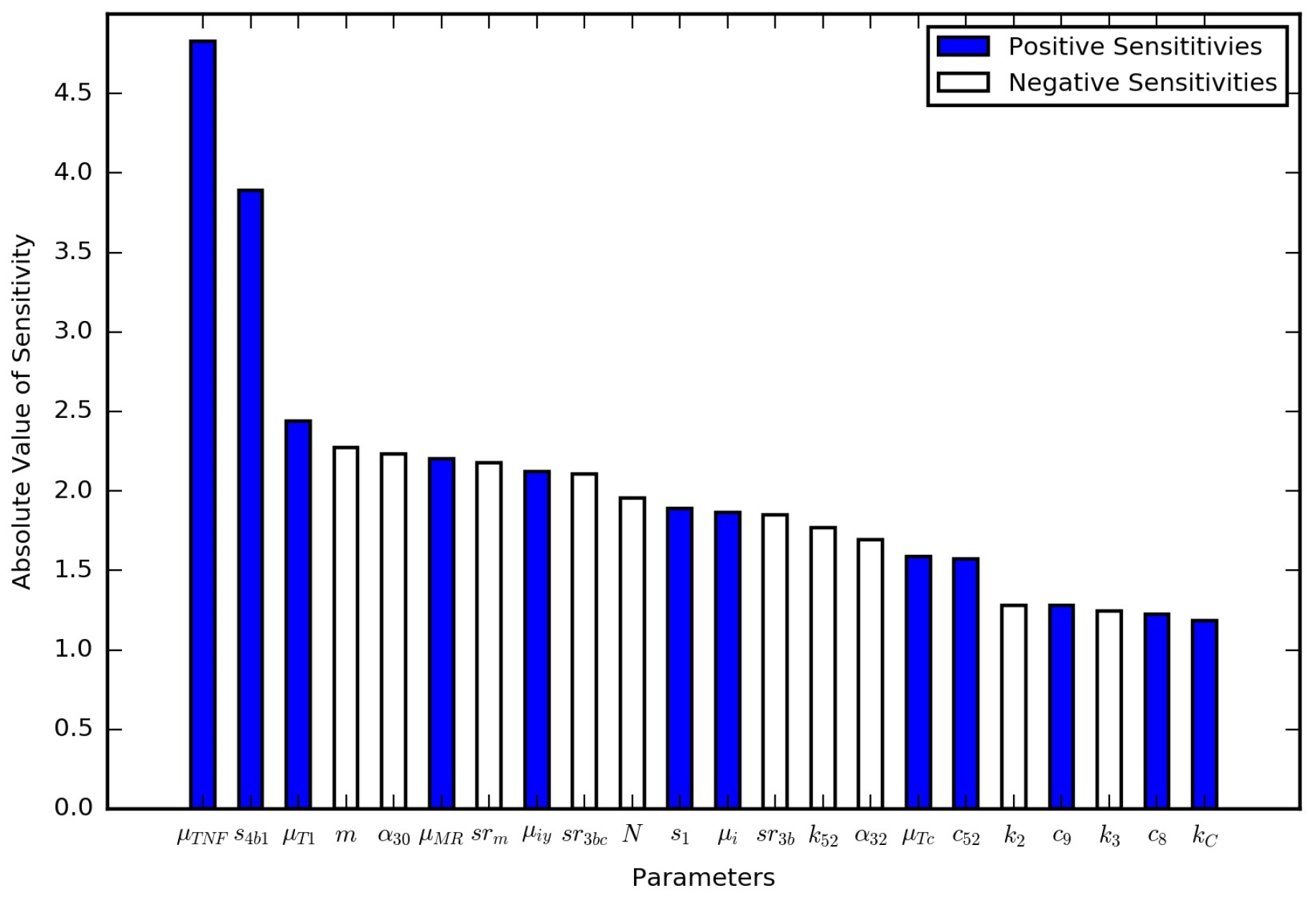
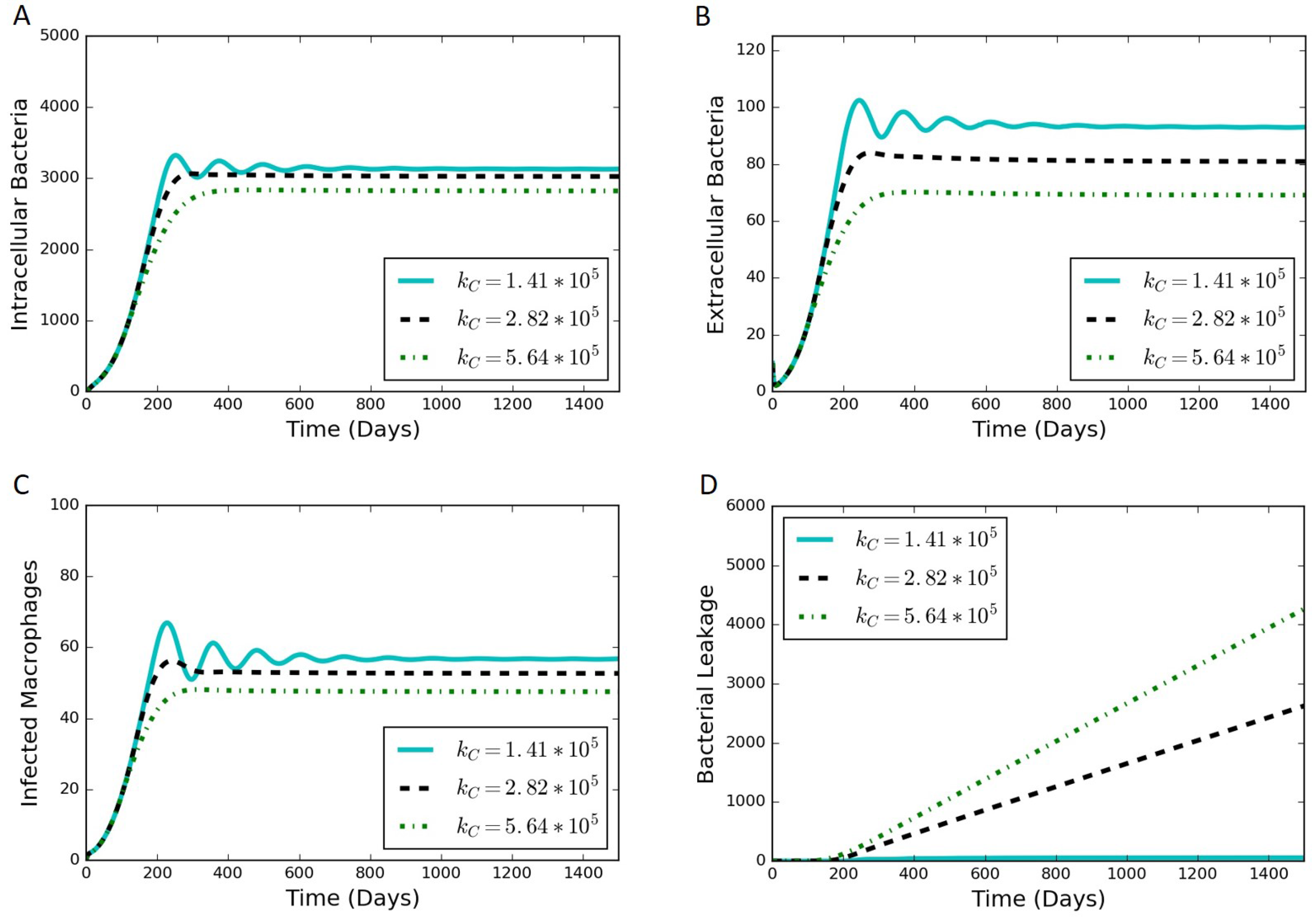
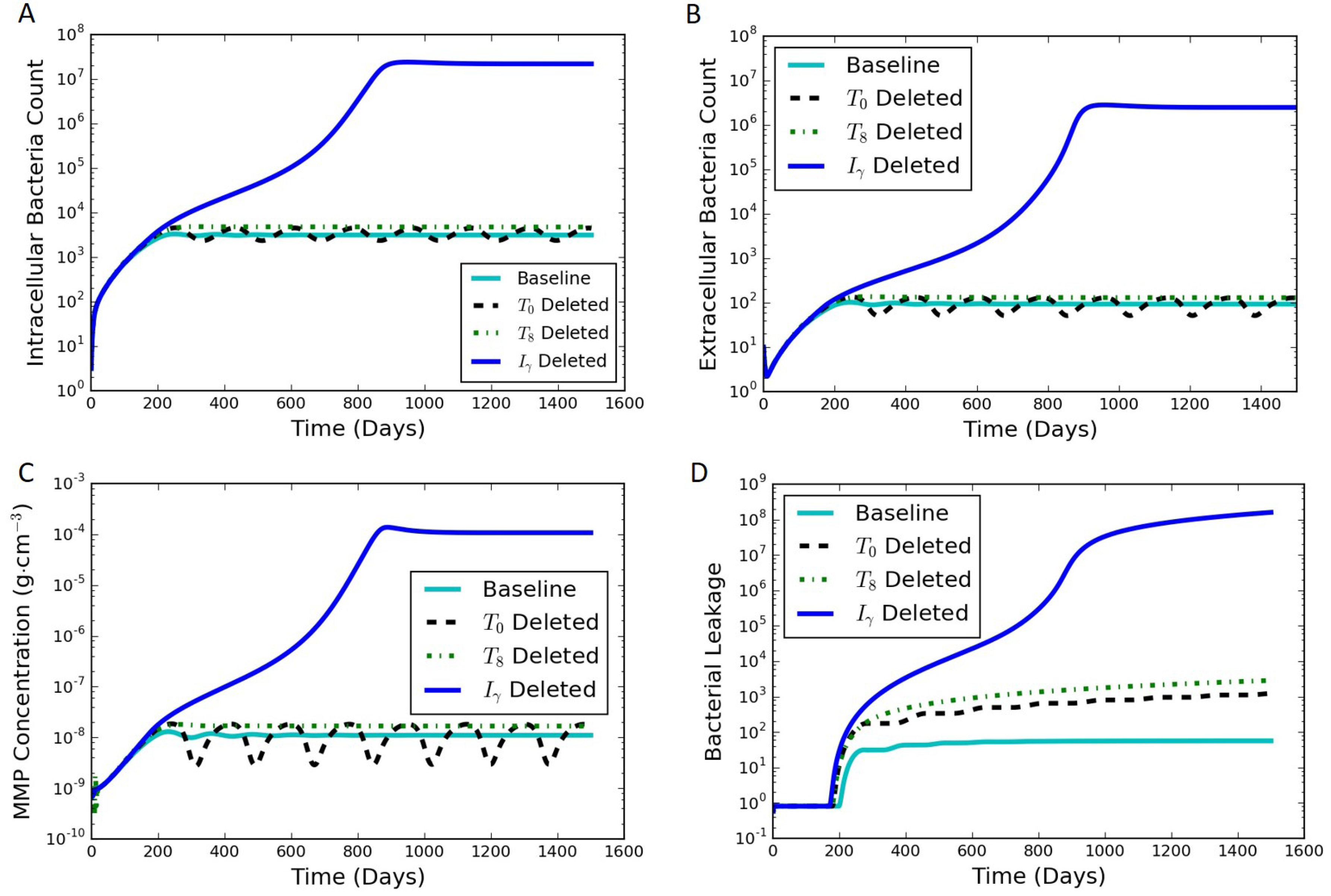
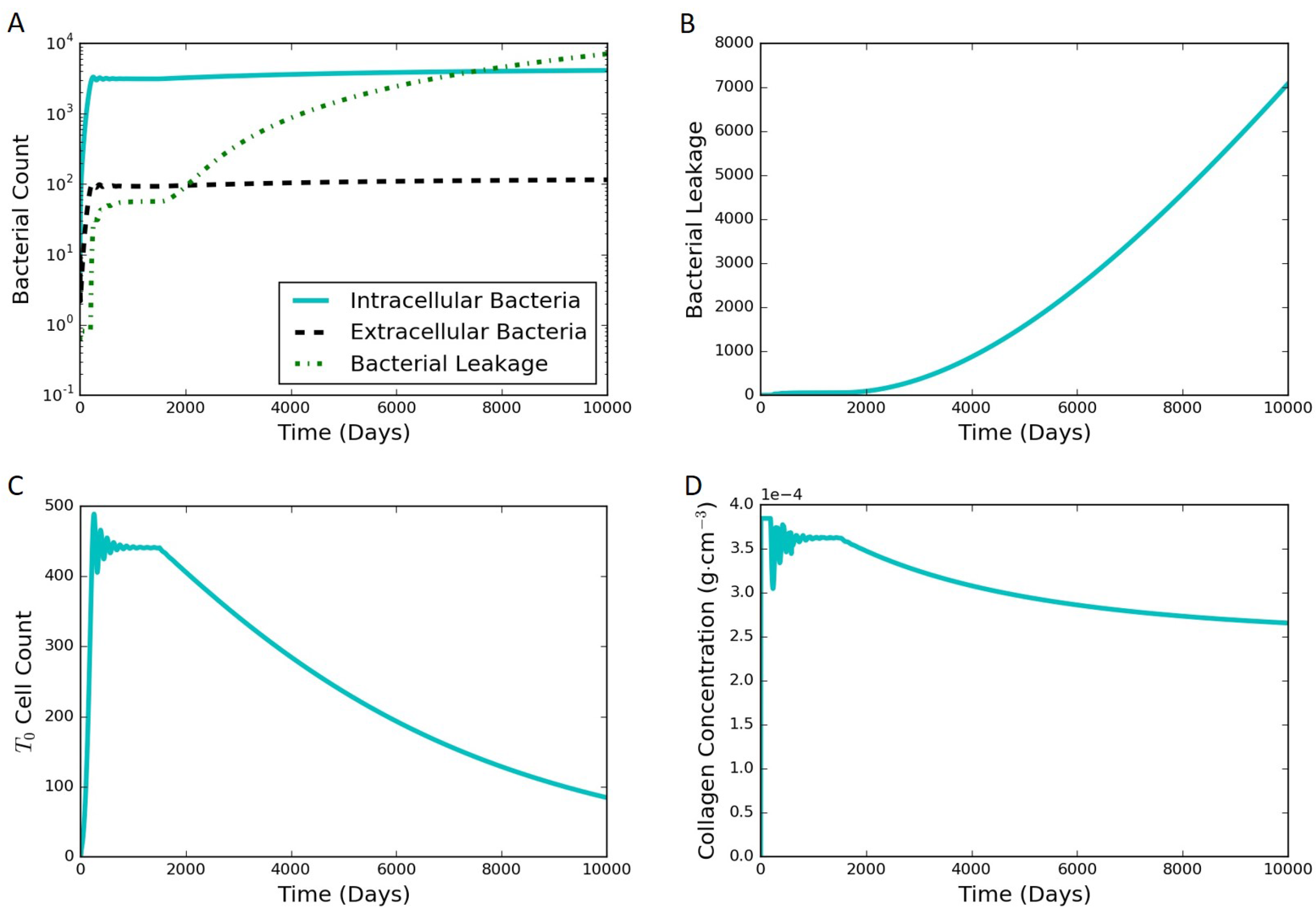
Section 3 includes model results under various scenarios as well as an analysis of the model sensitivity to parameter values.
2. Methods
The immune response model considers the local immune response to Mtb in the lungs. The immune response model includes population balances for macrophages, two families of T cells (CD4+ and CD8+), intracellular bacteria (inside infected macrophages), and extracellular bacteria (inside the granuloma but outside the macrophages). The model also includes the signaling network that connects the various cell populations via the cytokines TNF-
, interferon gamma (IFN-
), IL-4, IL-10, and IL-12 [
13]. The equations and parameters of the immune response model are summarized in the
Supplementary Materials. The immune response model can generate three regimes representing the infection outcomes of clearance, latency, and active TB. However, the immune response model does not consider the dynamic effects of MMP on the collagen on the surface of the granuloma, which, if breached, can lead to bacterial leakage. In addition, the immune response model cannot predict the transition from latent to activate TB after a period of latency. The present work extends the immune response model by considering the effects of intracellular bacteria on reprogramming infected macrophages to increase production of MMP and the subsequent degradation of the collagen envelope of granuloma by MMP. Here, we focus on additions to the original immune response model that represent the local changes in MMP concentration, collagen concentration, and the leakage of extracellular bacteria from the granuloma. Each of these additions is described in turn in the following subsections. Here, the granulomas are considered as well-mixed zones without transport limitations to facilitate adaptation of the ordinary differential equation based immune response model. An alternate partial differential equation model for the immune response with spatial effects including diffusion has been formulated [
16]. However, the simulation results for that model were only shown for spatially averaged populations of cells and cytokines. We seek to improve understanding of the process dynamics in the present work and thus follow these previously published models in neglecting spatial effects inside of granulomas.
2.1. MMP Dynamics
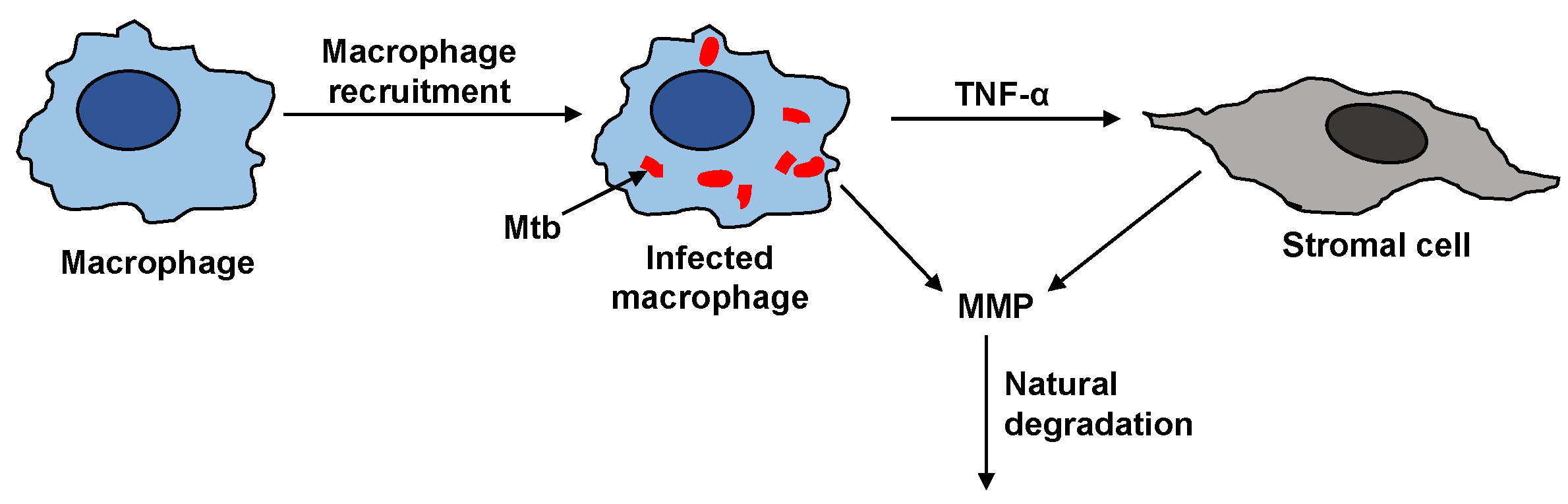
The steps that affect the MMP dynamics are illustrated in
Figure 2. The activation of resting macrophages, the recruitment of additional macrophages, and the infection of macrophages by Mtb are well-characterized by the immune response model. Macrophages infected with Mtb have been observed to induce gene expression and secretion of MMP; however, a compensatory increase in secretion of TIMP was not observed [
7]. The infected macrophages are not the only source of MMP in a granuloma. The infected macrophages interact with the stromal cells like epithelial cells, fibroblasts, and astrocytes, which further secrete MMP and together amplify the MMP upregulation. The pro-inflammatory cytokines especially TNF-
have been found to play a key role in triggering the upregulation of MMPs by stromal cells [
18]. It has been found that interaction between macrophages and stromal cell requires TNF-
to increase the MMP secretion by stromal cell networks [
18,
19,
20,
21].
The mass balance for MMP in terms of concentration for a constant volume system,
, is
where the first term represents secretion of MMPs indirectly by the stromal cells that requires both TNF-
,
, and infected macrophages,
; the second term represents production of MMPs by reprogrammed infected macrophages; the last two terms represent the natural first-order degradation and constant production of MMP;
is the rate constant for indirect production of MMP;
is the constant where the effect of TNF-
on the indirect MMP production has reached half of its saturation level;
is the rate constant for direct production of MMP by
;
is the half life of MMP; and
is the basal constant recruitment rate of MMP. The last two terms maintain the constant concentration of MMP at equilibrium in the latent state. The functional forms for the two terms representing the upregulation of MMP in the presence of Mtb infection were based on the general mathematical forms defined in [
13]: i.e., all terms that require the presence of infected macrophages to upregulate a process are given a linear dependence on
, while all terms that are upregulated by a cytokine such as TNF-
are given a Michaelis-Menten type saturation equation dependent on
.
2.2. Collagen Dynamics
The collagen dynamics during granuloma formation at the onset of TB infection is beyond the scope of the current work focused on the latent to active transition. Therefore, for simplicity, we consider a constant source term for collagen representative of the source after latency is achieved. The effects of MMP on degrading the collagen are incorporated to study how the granulomas can be compromised after latent TB is established (
Figure 3). The cleavage of collagen by MMPs was found to display Michaelis–Menten kinetics [
22,
23]. We recognize that the granulomas should have the collagen fibers concentrated on the exterior surfaces. In the model proposed here, there is no spatial variation. This could be a realistic approximation if the MMP is uniformly secreted within and adjacent to the granulomas or if the transport occurs faster than the degradation time scale. Slow collagen degradation is considered here, making this well-mixed model reasonable. The mass balance for the change in concentration of collagen,
C, in the well-mixed granuloma is
where
is a constant recruitment term representing the external build up of the collagen envelope around the granuloma,
is the rate constant of collagen degradation, and
is the Michaelis constant for collagen degradation catalyzed by MMP.
2.3. Bacterial Leakage
The change in the population of the extracellular bacteria inside the granuloma,
, has two terms:
where
is the non-leaking extracullular bacteria and
is the leaking extracellular bacteria (
Figure 1). The first term
is the contribution from the immune response model and is given by
(S16) in the Supplementary Materials, which considers all the different mechanisms for the gain and loss in extracellular bacterial count corresponding to the release and uptake of intracellular bacteria by the macrophages, respectively, and the constant turnover number. However, this term does not capture the loss of extracellular bacteria population when the granuloma starts leaking extracellular bacteria into the lung. To account for this case, the second term is added to (
3) to track the leakage of bacteria through deteriorated collagen and is represented by
where
is the rate constant for the maximum rate of bacteria exiting the granuloma,
is the half saturation constant for the inhibitory effect of collagen on this process, and
is the expected collagen concentration at latency. When the concentration of collagen is equal to the concentration of collagen in the latent case (
), the granuloma is intact with no leakage of bacteria, thus making the leakage term (
4) zero. Zero is the maximum value for (
4), i.e., the bacterial leakage never has a positive value. This is ensured by the maximum value of
C in the model formulation, which is
. The maximum collagen concentration with respect to changes in MMP concentration is determined by
The solution to the maximization problem is
, which is how
was defined using the values
and
. At the other extreme, when the concentration of collagen goes to zero, there is no longer a barrier around the bacteria, and the rate of leakage is directly dependent on the extracellular bacterial count giving the fastest bacteria leakage rate from the granuloma.
2.4. Biological Feedback
Although not shown explicitly, a feedback loop is formed between the equations introduced here (
1)–(
4) via the species included in the immune response model (see
Supplementary Materials (S1)–(S16)). It is apparent that Equation (
2) depends on the value of (MMP), and Equations (
3)–(
4) depend on
C. The contribution to the extracellular bacterial count from the immune response
depends on multiple species in (S16) including
. The extracellular bacteria count
in-turn leads directly to changes in species
,
,
,
,
,
, and
through dependence on
or
in equations (S1), (S2), (S3), (S10), (S11), (S14), and (S15), respectively. Furthermore, changes in
,
,
,
, and some T cells affect the production of
. Changes in
and
directly lead to changes in the production of MMP given by (
1). Other pathways for indirect feedback exist between the cytokines and the bacterial-population-sensitive macrophages.
2.5. Parameter Values
Parameters need to be specified to define the system before performing any simulations. A value of
is taken from a mathematical model for MMP in fibrosis [
24] and is used as the basis for calculating the rest of the parameters for (
1). The value of
is calculated by evaluating (
1) with no infection and data from [
25]. The value of
at the end of a typical latent simulation is used to calculate
. Values of
and
are then calculated using data from [
7,
25]. Both
and
are kinetic parameters taken from published experimental data on characterizing the kinetics of MMP. However, the parameters for the breakdown of collagen in literature have two sets of parameters: one for each of the two proteins,
-1 and
-2, that compose collagen I [
23]. For this model, a weighted average of the parameters based on the number of proteins in each stand is used and converted into the appropriate units. The source term for collagen I,
, is then calculated from a typical collagen concentration [
24]. The value of collagen expected in the latent case,
, is set based on data from the same fibrosis model used for
[
24]. The rest of the parameters in (
4),
,
, and
, are calculated to give the steady state and reasonable limiting behavior. The parameters are defined in
Table 1. The parameters listed in the
Supplementary Materials Table S1 were validated for the immune response model [
13].
2.6. Numerical Methods and Code Repository
The system of ordinary differential equations in the immune response model (see
Supplementary Materials (S1)–(S16)) and the TB granuloma active model defined by (
1)–(
4) was solved with
odeint solver from the SciPy Integrate Python module, which uses the classic
lsoda routine from the FORTAN library
odepack. The default options were used in the solver. The parameter values in
Table 1 were used to generate the results in
Section 3, unless otherwise indicated. The initial conditions are given in
Table 2. To enable code reuse, we wrote the model in Python and shared the code, parameter files, and documentation in an open-source software repository at
http://github.com/ashleefv/tbActivationDynamics [
26].













{kind=link}
{kind=link}
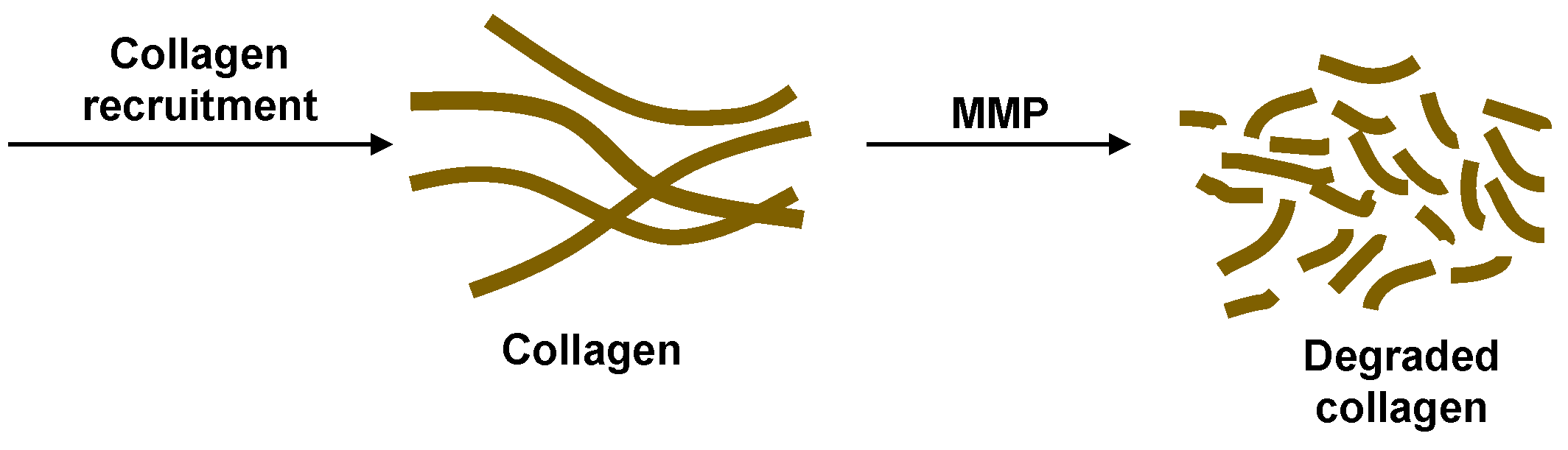{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}