Tree Bark Phenols Regulate the Physiological and Biochemical Performance of Gladiolus Flowers

Abstract
:1. Introduction
2. Results
2.1. Morphological Responses and Bark Chemical Composition and Antioxidant Activities
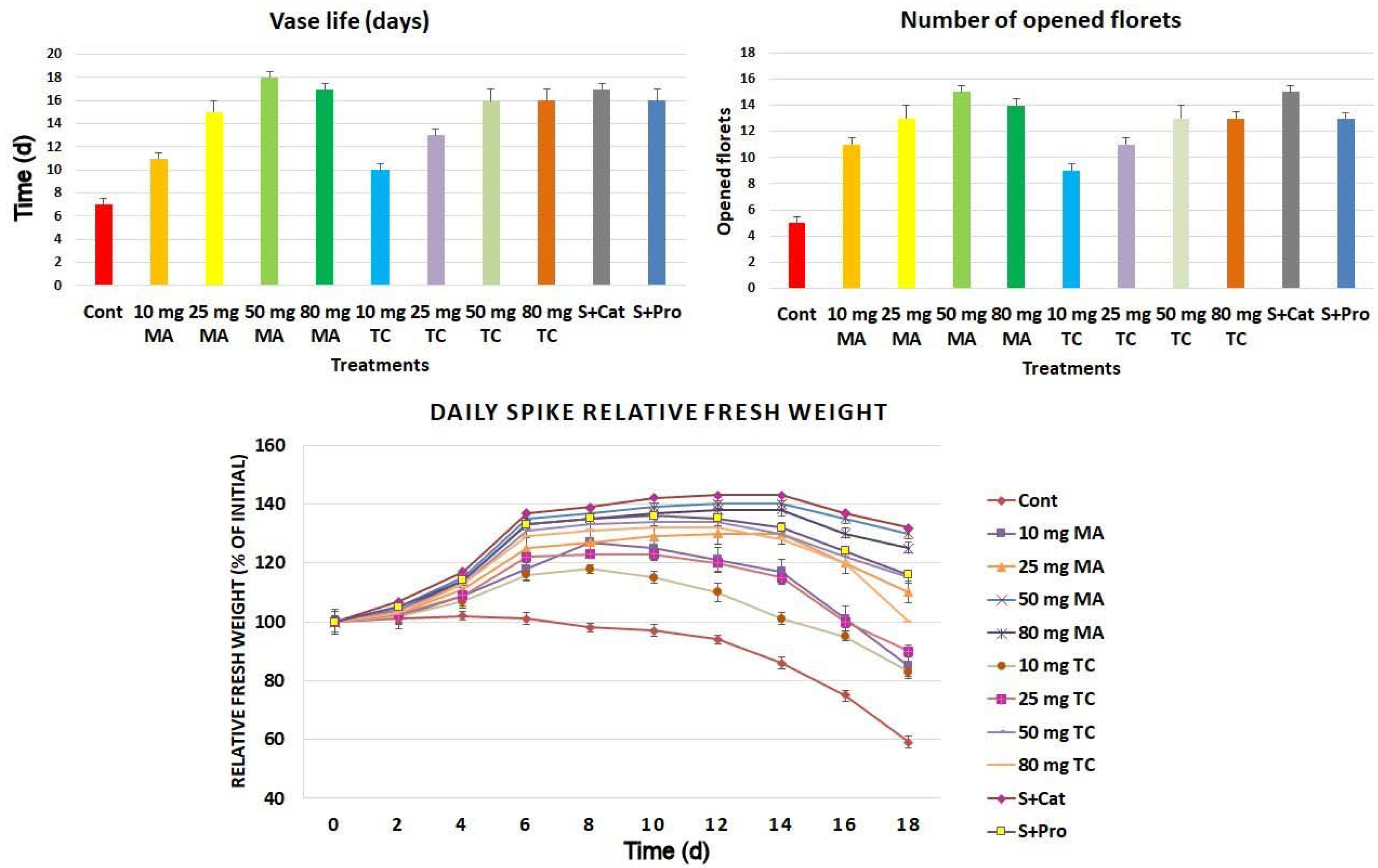
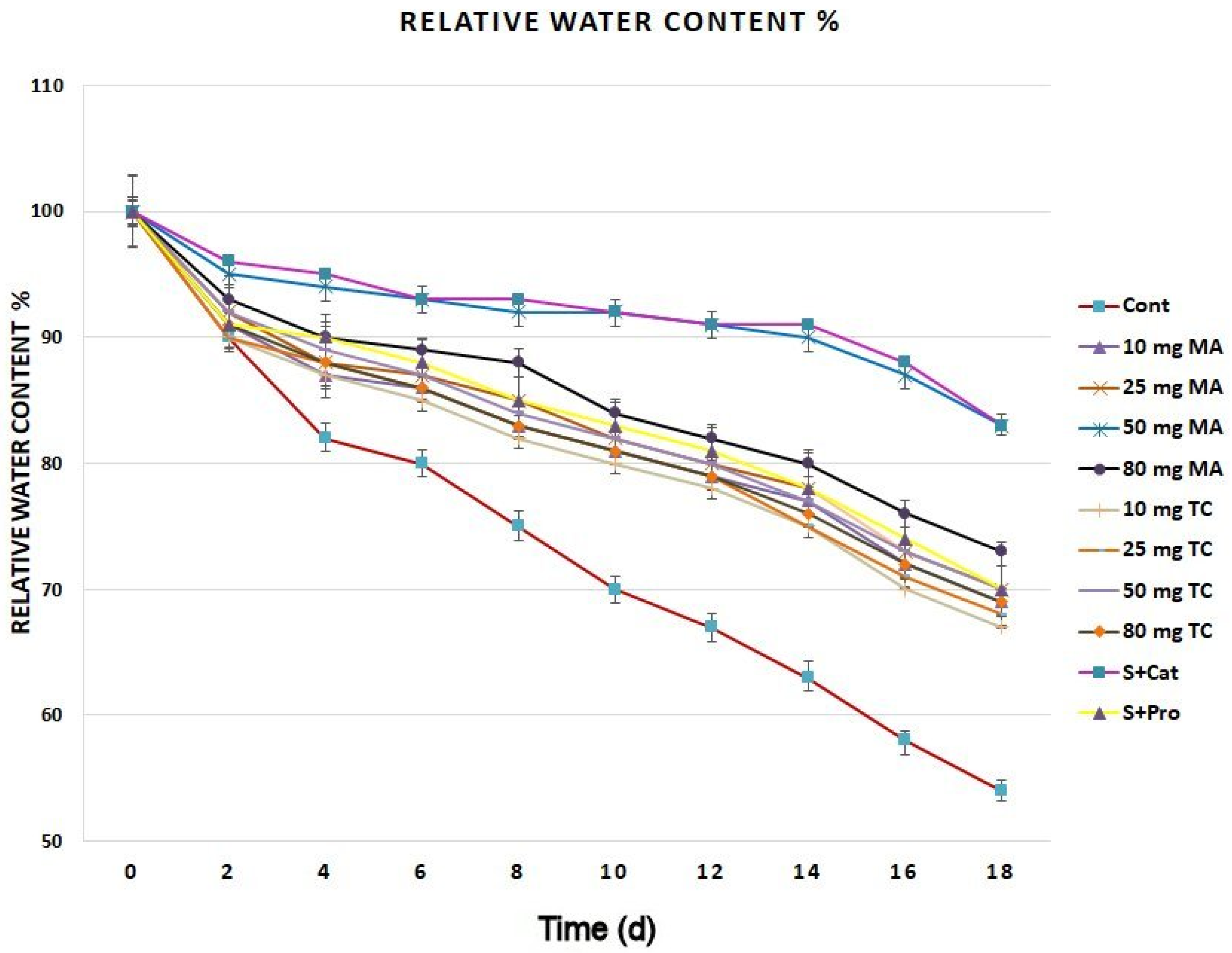
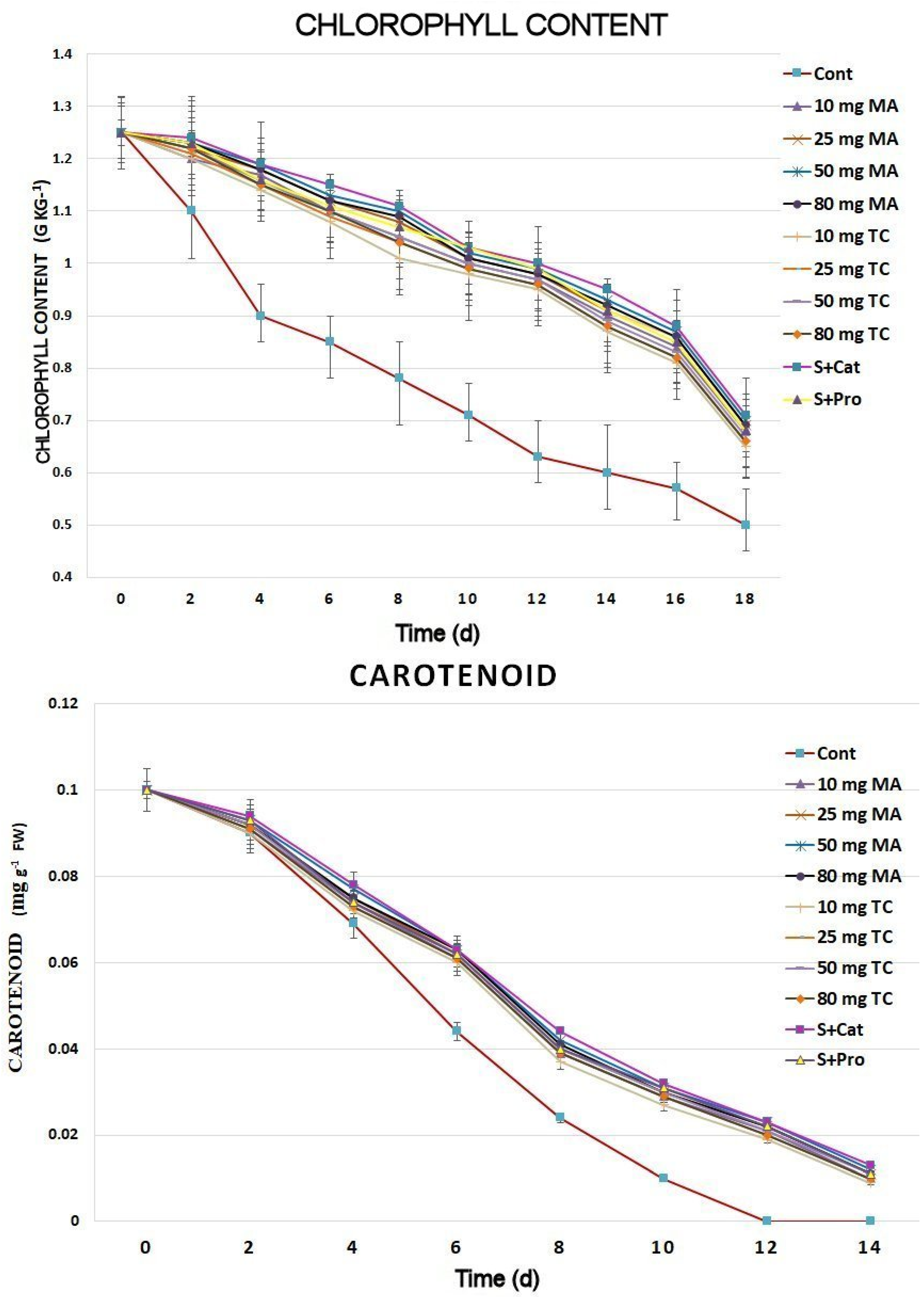
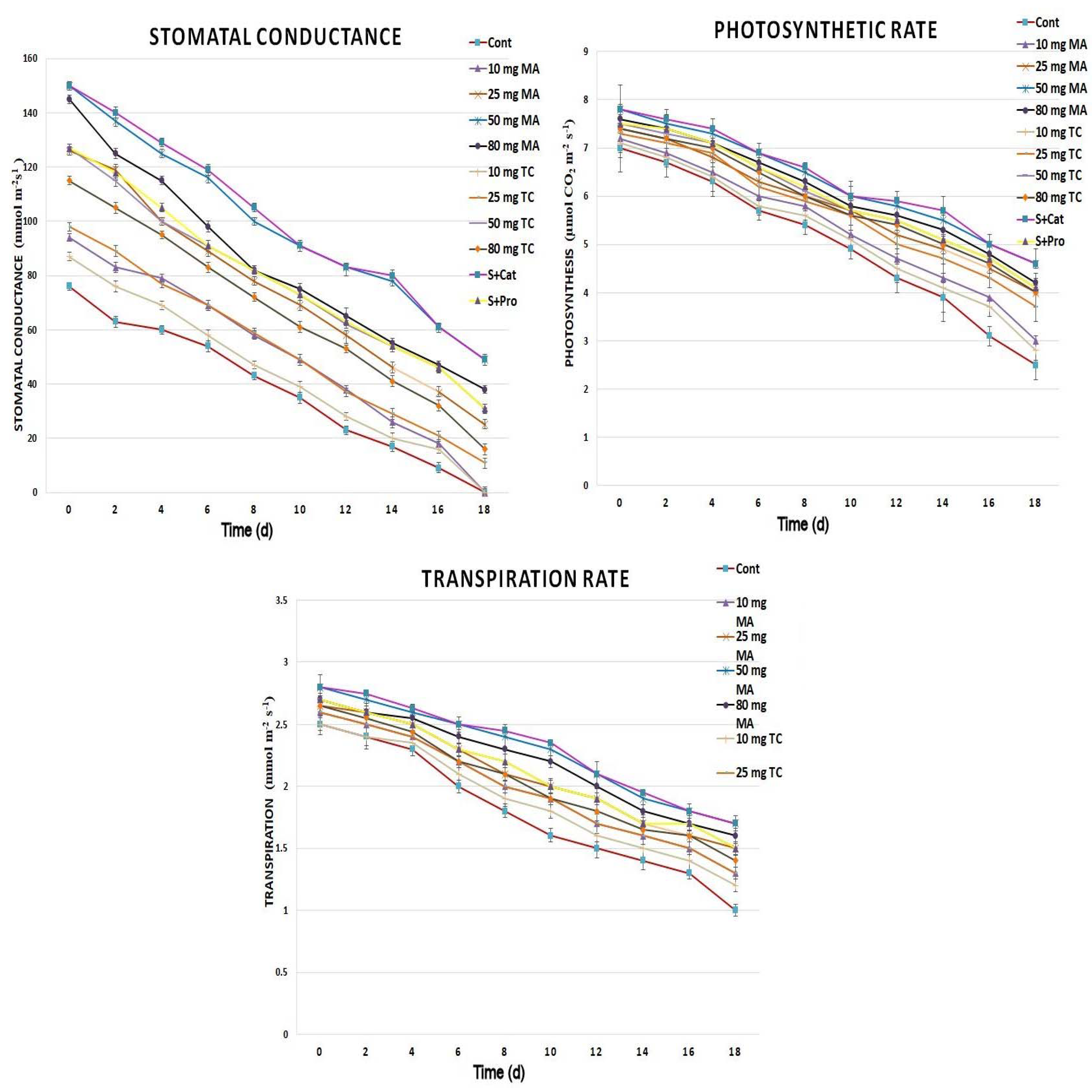
2.2. Physiological Responses
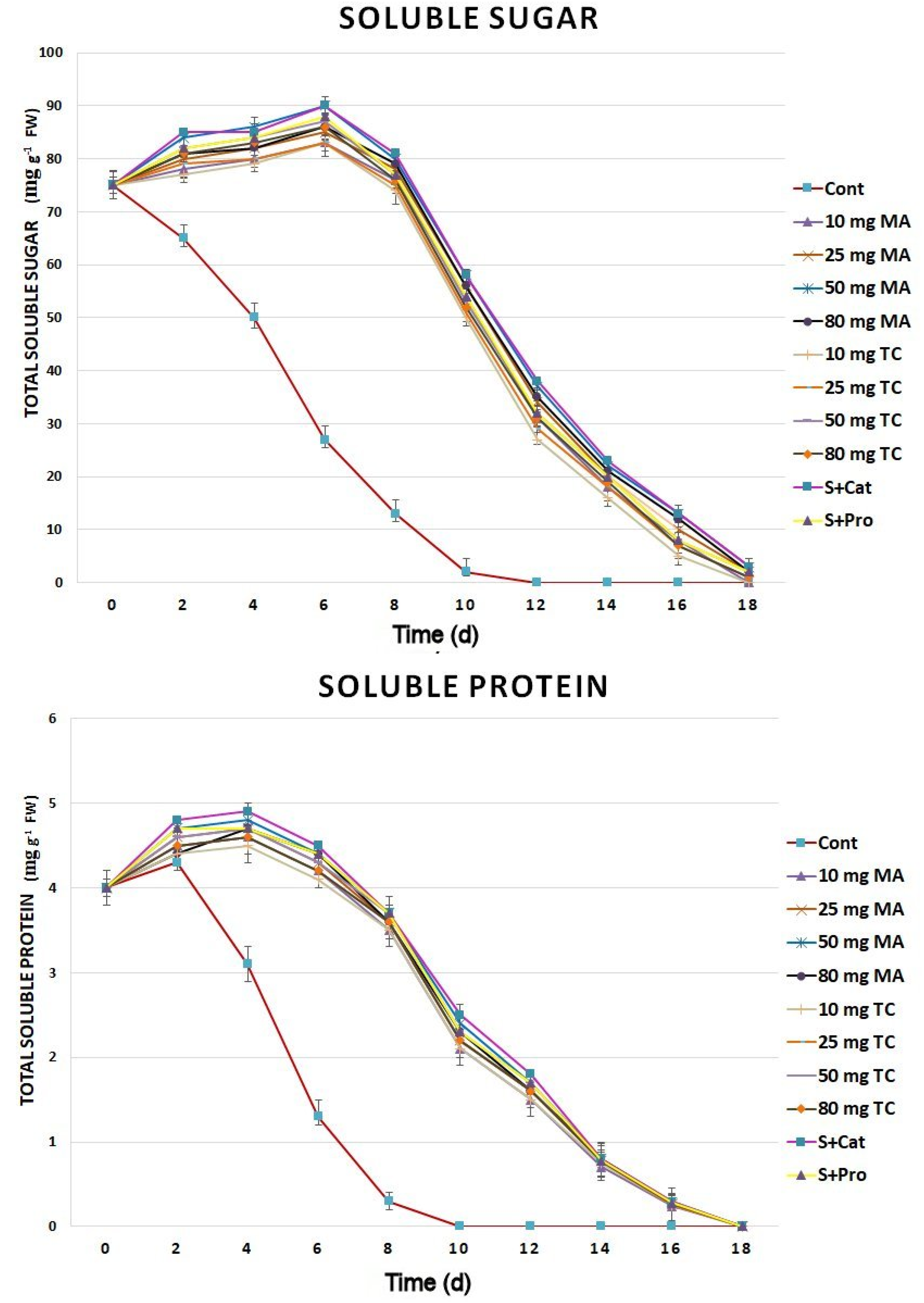
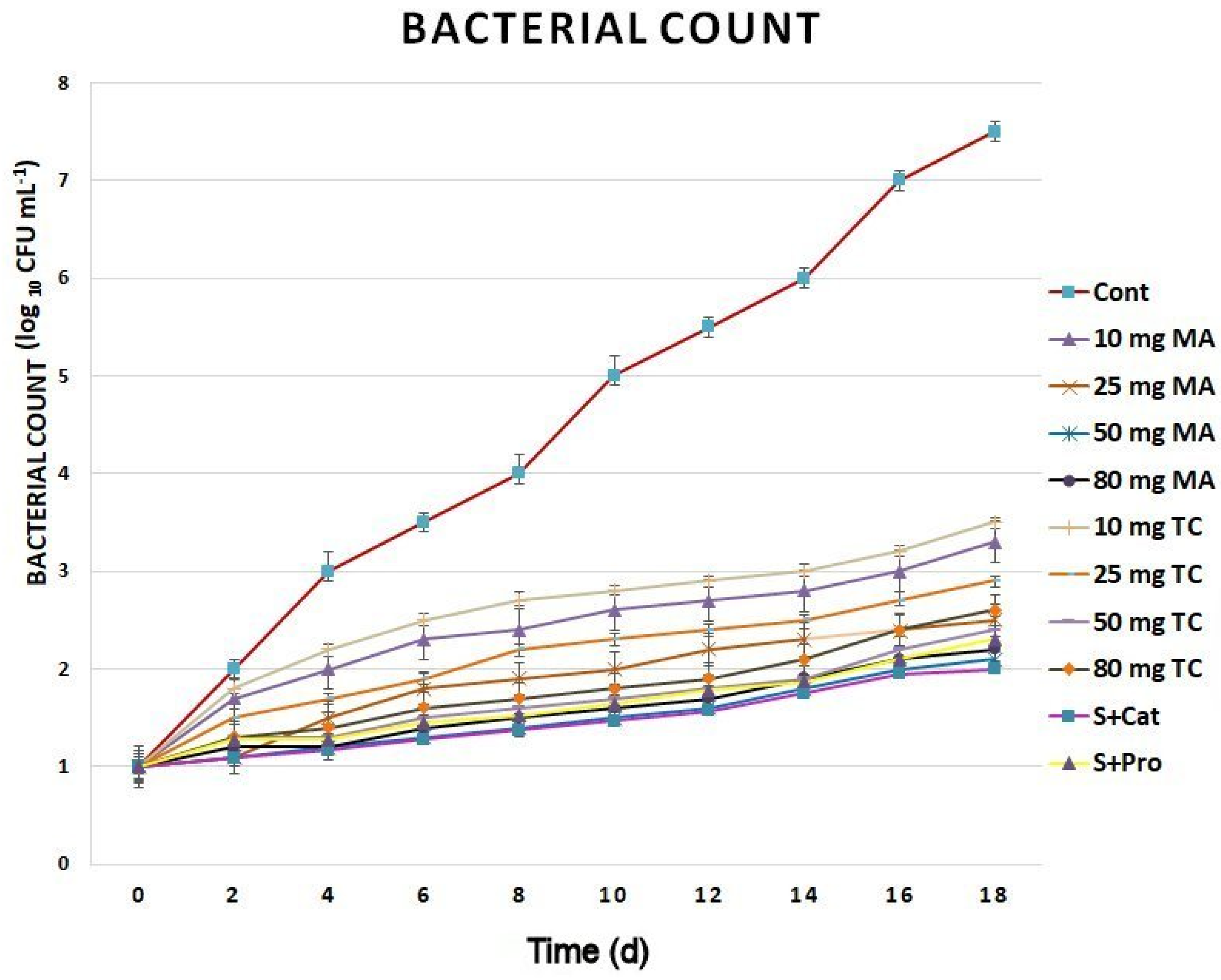
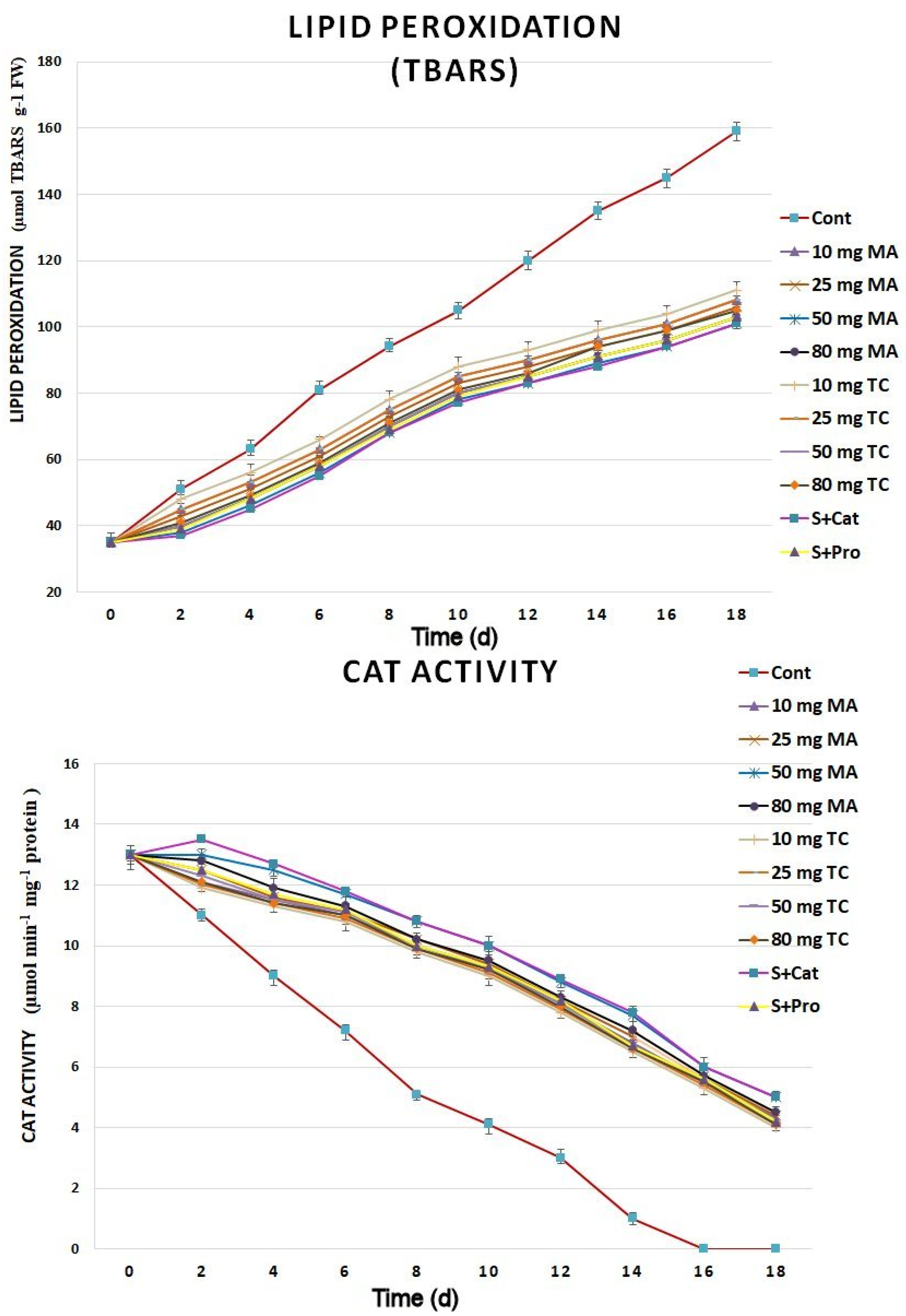
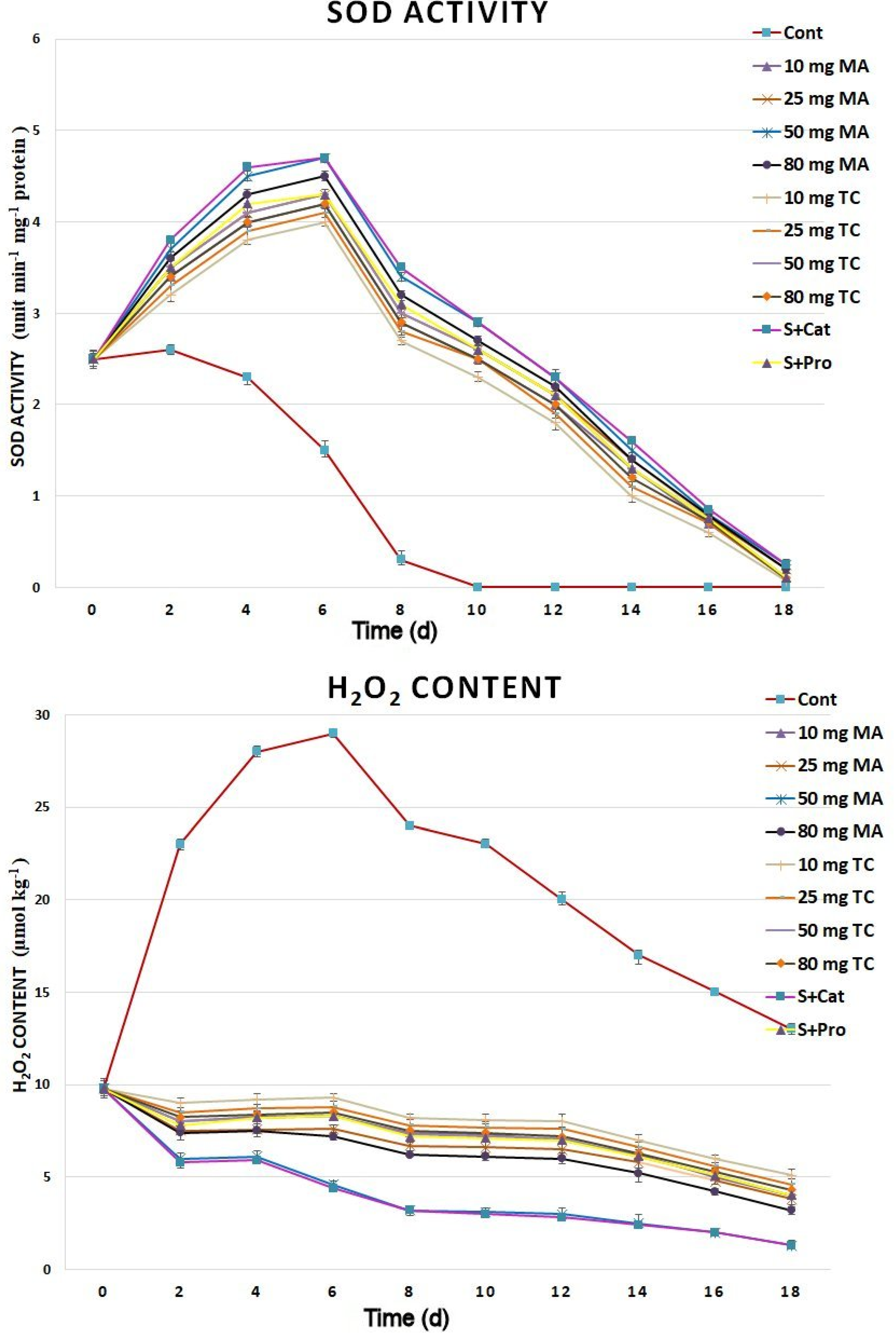
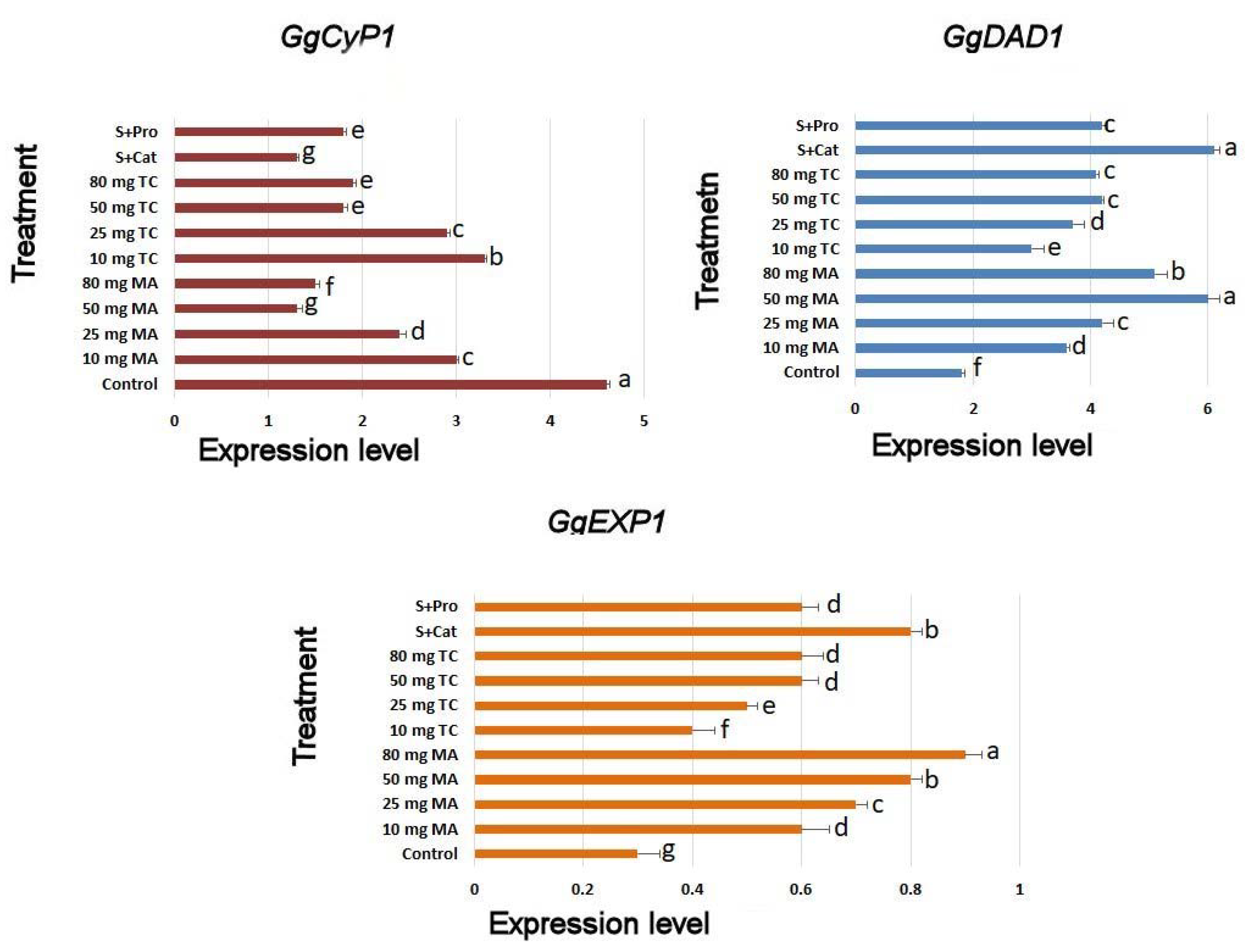
2.3. Biochemical Responses and Expression Analyses
3. Discussion
4. Material and Methods
4.1. Preparation and Analyses of Bark Extracts
4.2. Plant Material and Treatments
4.3. Morphological Parameters
4.4. Physiological and Biochemical Measurements
4.5. Expression Analyses
4.6. Statistical Analyses
5. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Ding, C.; Lei, L.; Yao, L.; Wang, L.; Hao, X.; Li, N.; Wang, Y.; Yin, P.; Guo, G.; Yang, Y.; et al. The involvements of calcium-dependent protein kinases and catechins in tea plant [Camellia sinensis (L.) O. Kuntze] cold responses. Plant Physiol. Biochem. 2019, 143, 190–202. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, K.; Watanabe, K.; Ono, M. CRISPR/Cas9-mediated mutagenesis of the EPHEMERAL1 locus that regulates petal senescence in Japanese morning glory. Plant Physiol. Biochem. 2018, 131, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Leoni, B.; Loconsole, D.; Cristiano, G.; De Lucia, B. Comparison between Chemical Fertilization and Integrated Nutrient Management: Yield, Quality, N, and P Contents in Dendranthema grandiflorum (Ramat.) Kitam. Cultivars 2019, 9, 202. [Google Scholar] [CrossRef] [Green Version]
- Tork, D.G.; Anderson, N.O.; Wyse, D.L.; Betts, K.J. Domestication of Perennial Flax Using an Ideotype Approach for Oilseed, Cut Flower, and Garden Performance. Agronomy 2019, 9, 707. [Google Scholar] [CrossRef] [Green Version]
- Gourieroux, A.M.; McCully, M.E.; Holzapfel, B.P.; Scollary, G.R.; Rogiers, S.Y. Flowers regulate the growth and vascular development of the inflorescence rachis in Vitis vinifera L. Plant Physiol. Biochem. 2016, 108, 519–529. [Google Scholar] [CrossRef]
- Van Meeteren, U.; Arévalo-Galarza, L.; Van Doorn, W. Inhibition of water uptake after dry storage of cut flowers: Role of aspired and wound-induced processes in Crysanthemum. Postharvest Biol. Technol. 2006, 41, 70–77. [Google Scholar] [CrossRef]
- Sankat, C.K.; Mujaffar, S. Water Balance in Cut Anthurium Flowers in Storage and Its Effect on Quality; International Society for Horticultural Science (ISHS): Leuven, Belgium, 1994; pp. 723–732. [Google Scholar]
- Singh, A.; Kumar, J.; Kumar, P. Effects of plant growth regulators and sucrose on post harvest physiology, membrane stability and vase life of cut spikes of gladiolus. Plant Growth Regul. 2008, 55, 221–229. [Google Scholar] [CrossRef]
- Elansary, H.O.; Zin El-Abedin, T.K. Omeprazole alleviates water stress in peppermint and modulates the expression of menthol biosynthesis genes. Plant Physiol. Biochem. 2019, 139, 578–586. [Google Scholar] [CrossRef]
- Zhang, J.; Lv, J.; Dawuda, M.M.; Xie, J.; Yu, J.; Li, J.; Zhang, X.; Tang, C.; Wang, C.; Gan, Y. Appropriate Ammonium-Nitrate Ratio Improves Nutrient Accumulation and Fruit Quality in Pepper (Capsicum annuum L.). Agronomy 2019, 9, 683. [Google Scholar] [CrossRef] [Green Version]
- Sotek, Z.; Białecka, B.; Pilarczyk, B.; Drozd, R.; Pilarczyk, R.; Tomza-Marciniak, A.; Kruzhel, B.; Lysak, H.; Bąkowska, M.; Vovk, S. Antioxidant Activity and Selenium and Polyphenols Content from Selected Medicinal Plants Natives from Various Areas Abundant in Selenium (Poland, Lithuania, and Western Ukraine). Processes 2019, 7, 878. [Google Scholar] [CrossRef] [Green Version]
- Al-Fatimi, M. Antifungal Activity of Euclea divinorum Root and Study of its Ethnobotany and Phytopharmacology. Processes 2019, 7, 680. [Google Scholar] [CrossRef] [Green Version]
- Knee, M. Selection of biocides for use in floral preservatives. Postharvest Biol. Technol. 2000, 18, 227–234. [Google Scholar] [CrossRef]
- Saeed, T.; Hassan, I.; Abbasi, N.A.; Jilani, G. Effect of gibberellic acid on the vase life and oxidative activities in senescing cut gladiolus flowers. Plant Growth Regul. 2014, 72, 89–95. [Google Scholar] [CrossRef]
- Saeed, T.; Hassan, I.; Jilani, G.; Abbasi, N.A. Zinc augments the growth and floral attributes of gladiolus, and alleviates oxidative stress in cut flowers. Sci. Hortic Amst. 2013, 164, 124–129. [Google Scholar] [CrossRef]
- Hassan, F.A.S.; Fetouh, M.I. Does moringa leaf extract have preservative effect improving the longevity and postharvest quality of gladiolus cut spikes? Sci. Hortic Amst. 2019, 250, 287–293. [Google Scholar] [CrossRef]
- Maity, T.R.; Samanta, A.; Datta, S. Evaluation of Piper betle mediated silver nanoparticle in post-harvest physiology in relation to vase life of cut spike of Gladiolus. Bull. Natl. Res. Cent. 2019, 43, 9. [Google Scholar] [CrossRef] [Green Version]
- Chantuma, P.; Lacointe, A.; Kasemsap, P.; Thanisawanyangkura, S.; Gohet, E.; Clement, A.; Guilliot, A.; Ameglio, T.; Thaler, P. Carbohydrate storage in wood and bark of rubber trees submitted to different level of C demand induced by latex tapping. Tree Physiol. 2009, 29, 1021–1031. [Google Scholar] [CrossRef] [Green Version]
- Mayans, B.; Pérez-Esteban, J.; Escolástico, C.; Eymar, E.; Masaguer, A. Evaluation of Commercial Humic Substances and Other Organic Amendments for the Immobilization of Copper Through 13C CPMAS NMR, FT-IR, and DSC Analyses. Agronomy 2019, 9, 762. [Google Scholar] [CrossRef] [Green Version]
- Sakaue, Y.; Domon, H.; Oda, M.; Takenaka, S.; Kubo, M.; Fukuyama, Y.; Okiji, T.; Terao, Y. Anti-biofilm and bactericidal effects of magnolia bark-derived magnolol and honokiol on Streptococcus mutans. Microbiol. Immunol. 2016, 60, 10–16. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Szopa, A.; Kubica, P.; Al-Mana, F.A.; Mahmoud, E.A.; El-Abedin, T.K.A.Z.; Mattar, M.A.; Ekiert, H. Phenolic Compounds of Catalpa speciosa, Taxus cuspidata, and Magnolia acuminata have Antioxidant and Anticancer Activity. Molecules 2019, 24, 412. [Google Scholar] [CrossRef] [Green Version]
- Bajpai, V.K.; Sharma, A.; Baek, K.-H. Antibacterial Mechanism of Action of Taxus cuspidata Stem Essential Oil against Selected Foodborne Pathogens. Food Saf. 2013, 33, 348–359. [Google Scholar] [CrossRef]
- Grobelna, A.; Kalisz, S.; Kieliszek, M. The Effect of the Addition of Blue Honeysuckle Berry Juice to Apple Juice on the Selected Quality Characteristics, Anthocyanin Stability, and Antioxidant Properties. Biomolecules 2019, 9, 744. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Szopa, A.; Kubica, P.; Ekiert, H.; Mattar, M.; Al-Yafrasi, M.A.; El-Ansary, D.O.; Zin El-Abedin, T.K.; Yessoufou, K. Polyphenol Profile and Pharmaceutical Potential of Quercus spp. Bark Extracts. Plants 2019, 8, 486. [Google Scholar] [CrossRef] [Green Version]
- Andrade, C.; Ferreres, F.; Gomes, G.M.N.; Duangsrisai, S.; Srisombat, N.; Vajrodaya, S.; Pereira, M.D.; Gil-Izquierdo, A.; Andrade, B.P.; Valentão, P. Phenolic Profiling and Biological Potential of Ficus curtipes Corner Leaves and Stem Bark: 5-Lipoxygenase Inhibition and Interference with NO Levels in LPS-Stimulated RAW 264.7 Macrophages. Biomolecules 2019, 9, 400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maity, T.R.; Samanta, A.; Jana, D.; Saha, B.; Datta, S. Effect of Piper betle leaf extract on post-harvest physiology and vascular blockage in relation to vase life and keeping quality of cut spike of tuberose (Polianthes tuberosa L. cv. Single). Indian J. Plant Physiol. 2014, 19, 250–256. [Google Scholar] [CrossRef]
- Cavaiuolo, M.; Cocetta, G.; Ferrante, A. The Antioxidants Changes in Ornamental Flowers during Development and Senescence. Antioxidants 2013, 2, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, A.J.; Cramer, M.D.; Watt, D.A. Regulation of photosynthesis by sugars in sugarcane leaves. J. Plant Physiol. 2008, 165, 1817–1829. [Google Scholar] [CrossRef]
- Nikinmaa, E.; Sievänen, R.; Hölttä, T. Dynamics of leaf gas exchange, xylem and phloem transport, water potential and carbohydrate concentration in a realistic 3-D model tree crown. Ann. Bot. 2014, 114, 653–666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nazemi Rafi, Z.; Ramezanian, A. Vase life of cut rose cultivars ‘Avalanche’ and ‘Fiesta’ as affected by Nano-Silver and S-carvone treatments. S. Afr. J. Bot. 2013, 86, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Semaming, Y.; Pannengpetch, P.; Chattipakorn, S.C.; Chattipakorn, N. Pharmacological Properties of Protocatechuic Acid and Its Potential Roles as Complementary Medicine. Evid. Based Complement. Altern. Med. 2015, 2015, 593902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taylor, P.W.; Hamilton-Miller, J.M.T.; Stapleton, P.D. Antimicrobial properties of green tea catechins. Food Sci. Technol. Bull. 2005, 2, 71–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borges, J.d.C.; Perim, M.C.; de Castro, R.O.; Araújo, T.A.d.S.; Peixoto Sobrinho, T.J.d.S.; da Silva, A.C.O.; Mariano, S.M.B.; Carreiro, S.C.; Pranchevicius, M.C.d.S. Evaluation of antibacterial activity of the bark and leaf extracts of Brosimum gaudichaudii Trécul against multidrug resistant strains. Nat. Prod. Res. 2017, 31, 2931–2935. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.S.; Tahir, I. Regulatory role of phenols in flower development and senescence in the genus Iris. Indian J. Plant Physiol. 2017, 22, 135–140. [Google Scholar] [CrossRef]
- Naing, A.H.; Win, N.M.; Han, J.-S.; Lim, K.B.; Kim, C.K. Role of Nano-silver and the Bacterial Strain Enterobacter cloacae in Increasing Vase Life of Cut Carnation ‘Omea’. Front. Plant Sci. 2017, 8, 1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Put, H.M.C. Micro-organisms from freshly harvested cut flower stems and developing during the vase life of chrysanthemum, gerbera and rose cultivars. Sci. Hortic Amst. 1990, 43, 129–144. [Google Scholar] [CrossRef]
- Mitani, T.; Ota, K.; Inaba, N.; Kishida, K.; Koyama, H.A. Antimicrobial Activity of the Phenolic Compounds of Prunus mume against Enterobacteria. Biol. Pharm. Bull. 2018, 41, 208–212. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Szopa, A.; Klimek-Szczykutowicz, M.; Jafernik, K.; Ekiert, H.; Mahmoud, E.A.; Barakat, A.A.; El-Ansary, D.O. Mammillaria Species—Polyphenols Studies and Anti-Cancer, Anti-Oxidant, and Anti-Bacterial Activities. Molecules 2019, 25, 131. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Salem, M.Z.M.; Ashmawy, N.A.; Yessoufou, K.; El-Settawy, A.A.A. In vitro antibacterial, antifungal and antioxidant activities of Eucalyptus spp. leaf extracts related to phenolic composition. Nat. Prod. Res. 2017, 31, 2927–2930. [Google Scholar] [CrossRef]
- Elansary, H.O.; Mahmoud, A.E.; El-Ansary, D.O.; Mattar, M.A. Effects of Water Stress and Modern Biostimulants on Growth and Quality Characteristics of Mint. Agronomy 2019, 10, 6. [Google Scholar] [CrossRef] [Green Version]
- Khenizy, S.A.; Abd El-Moneim, A.M.; Abdel-Fattah, G.H. Effect of natural extracts on vase life of Gypsophila cut flowers. Sci. J. Flowers Ornam. Plants 2014, 1, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48. [Google Scholar] [CrossRef] [PubMed]
- Saeed, T.; Hassana, I.; Abbasia, N.A.; Jilani, G.J.S.H. Antioxidative activities and qualitative changes in gladiolus cutflowers in response to salicylic acid application. Sci. Hortic Amst. 2016, 210. [Google Scholar] [CrossRef]
- Dwivedi, S.K.; Arora, A.; Singh, V.P.; Sairam, R.; Bhattacharya, R.C. Effect of sodium nitroprusside on differential activity of antioxidants and expression of SAGs in relation to vase life of gladiolus cut flowers. Sci. Hortic Amst. 2016, 210, 158–165. [Google Scholar] [CrossRef]
- Yamada, T.; Takatsu, Y.; Kasumi, M.; Marubashi, W.; Ichimura, K. A homolog of the defender against apoptotic death gene (DAD1) in senescing gladiolus petals is down-regulated prior to the onset of programmed cell death. Plant Physiol. 2004, 161, 1281–1283. [Google Scholar] [CrossRef] [PubMed]
- Azeez, A.; Sane, A.P.; Tripathi, S.K.; Bhatnagar, D.; Nath, P. The gladiolus GgEXPA1 is a GA-responsive alpha-expansin gene expressed ubiquitously during expansion of all floral tissues and leaves but repressed during organ senescence. Postharvest Biol. Technol. 2010, 58, 48–56. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Elansary, H.O.; El-Shanhorey, N.A.; Abdel-Hamid, A.M.E.; Ali, H.M.; Elshikh, M.S. Salicylic Acid-Regulated Antioxidant Mechanisms and Gene Expression Enhance Rosemary Performance under Saline Conditions. Front. Physiol. 2017, 8. [Google Scholar] [CrossRef]
- Irigoyen, J.J.; Einerich, D.W.; Sánchez-Díaz, M. Water stress induced changes in concentrations of proline and total soluble sugars in nodulated alfalfa (Medicago sativd) plants. Physiol. Plant. 1992, 84, 55–60. [Google Scholar] [CrossRef]
- Olajuyigbe, O.O.; Afolayan, A.J. Phenolic content and antioxidant property of the bark extracts of Ziziphus mucronata Willd. subsp. mucronata Willd. BMC Complement. Altern. Med. 2011, 11, 130. [Google Scholar] [CrossRef] [Green Version]
- Magel, E.; Einig, W.; Hampp, R. Carbohydrates in trees. In Developments in Crop Science; Gupta, A.K., Kaur, N., Eds.; Elsevier: Amsterdam, The Netherlands, 2000; Volume 26, pp. 317–336. [Google Scholar]
- He, S.; Joyce, D.C.; Irving, D.E.; Faragher, J.D. Stem end blockage in cut Grevillea ‘Crimson Yul-lo’ inflorescences. Postharvest Biol. Technol. 2006, 41, 78–84. [Google Scholar] [CrossRef] [Green Version]
- Weatherley, P.E. Studies in water relations of cotton plants I. The field measurement of water deficit in leaves. New Phytol. 1950, 49, 81–97. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elansary, H.O.; Yessoufou, K.; Abdel-Hamid, A.M.E.; El-Esawi, M.A.; Ali, H.M.; Elshikh, M.S. Seaweed Extracts Enhance Salam Turfgrass Performance during Prolonged Irrigation Intervals and Saline Shock. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamo, K.; Kim, A.-Y.; Park, S.H.; Joung, Y.H. The 5’UTR-intron of the Gladiolus polyubiquitin promoter GUBQ1 enhances translation efficiency in Gladiolus and Arabidopsis. BMC Plant Biol. 2012, 12, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Chemical Compound | Amount |
|---|---|---|
| M. acuminata | Total phenolics | 32 ± 0.53 mg g−1 |
| soluble sugars | 35 ± 0.71 mg g−1 | |
| Total antioxidants (DPPH) | 2.97 ± 0.13 (IC50, µg mL−1) | |
| Total antioxidants (β-carotene-linoleic acid) | 3.5 ± 0.12 (IC50, µg mL−1) | |
| Total antioxidants (ABTS) | 1.6 ± 0.1 (IC50, µg mL−1) | |
| Identified phenolics | ||
| Ellagic acid | 0.42 ± 0.03 (mg 100 g−1) | |
| Protocatechuic acid | 15.01 ± 0.9 (mg 100 g−1) | |
| Catechin | 83.12 ± 1.35 (mg 100 g−1) | |
| Epicatechin | 21.54 ± 1.01 (mg 100 g−1) | |
| Epigallocatechin gallate | 13.13 ± 0.17 (mg 100 g−1) | |
| T. cuspidata | Total phenolics | 26 mg g−1 |
| soluble sugars | 28 mg g−1 | |
| DPPH radical scavenging activity | 4.0 ± 0.2 (IC50, µg mL−1) | |
| β-carotene bleaching inhibition activity | 4.7 ± 0.2 (IC50, µg mL−1) | |
| ABTS radical scavenging activity | 2.0 ± 0.1 (IC50, µg mL−1) | |
| Identified phenolics | ||
| Caffeic acid | 3.12 ± 0.03 (mg 100 g−1) | |
| Chlorogenic acid | 8.20 ± 0.56 (mg 100 g−1) | |
| Gallic acid | 2.1 ± 0.09 (mg 100 g−1) | |
| p-Hydroxybenzoic acid | 2.13 ± 0.09 (mg 100 g−1) | |
| Hydroxycaffeic acid | 22.76 ± 0.8 (mg 100 g−1) | |
| Protocatechuic acid | 21.08 ± 1.23 (mg 100 g−1) |
© 2020 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elansary, H.O. Tree Bark Phenols Regulate the Physiological and Biochemical Performance of Gladiolus Flowers. Processes 2020, 8, 71. https://doi.org/10.3390/pr8010071
Elansary HO. Tree Bark Phenols Regulate the Physiological and Biochemical Performance of Gladiolus Flowers. Processes. 2020; 8(1):71. https://doi.org/10.3390/pr8010071
Chicago/Turabian StyleElansary, Hosam O. 2020. "Tree Bark Phenols Regulate the Physiological and Biochemical Performance of Gladiolus Flowers" Processes 8, no. 1: 71. https://doi.org/10.3390/pr8010071
APA StyleElansary, H. O. (2020). Tree Bark Phenols Regulate the Physiological and Biochemical Performance of Gladiolus Flowers. Processes, 8(1), 71. https://doi.org/10.3390/pr8010071