Isolation, Identification and Antimicrobial Evaluation of Bactericides Secreting Bacillus subtilis Natto as a Biocontrol Agent

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials, Chemicals, and Reagents
2.2. Microbial Strains
2.3. Isolation and Screening of B. subtilis Natto
2.4. Molecular Identification: DNA Extraction and PCR Amplification
2.5. S rDNA Sequencing and Data Analysis
2.6. Microbial Cultural Conditions
2.7. Antimicrobial Activity of B. subtilis Natto Extract
2.8. Minimal Inhibitory Concentration of B. subtilis Natto against Pathogenic Bacteria
2.9. Optimization of Growth Medium to Induce the Antibacterial Activity of B. subtilis Natto
2.10. Extraction and Purification of Bactericides from B. subtilis Natto
2.11. Structural Identification of Bactericides from B. subtilis Natto
3. Results and Discussion
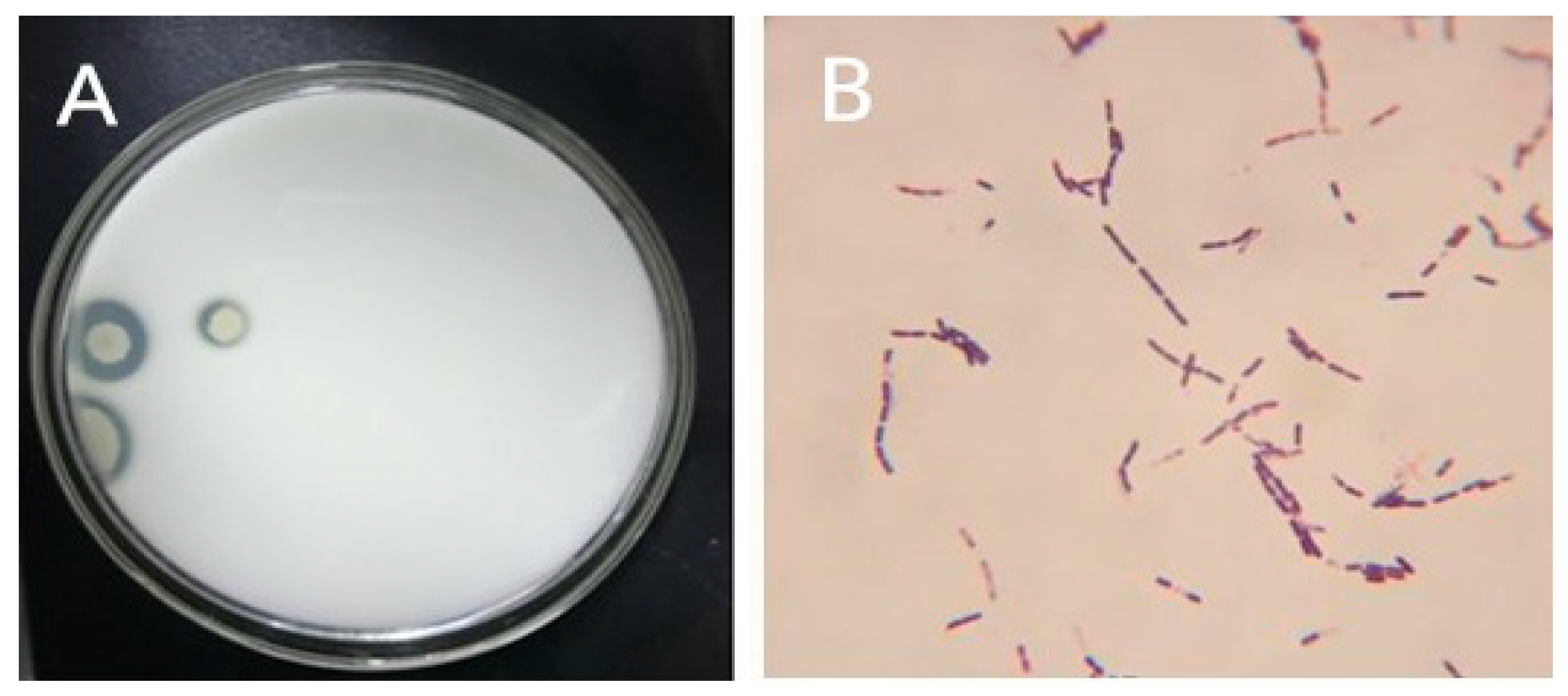
3.1. Isolation and Purification of B. subtilis Natto
3.2. Molecular Identification: 16S Ribosomal DNA Sequence Analysis
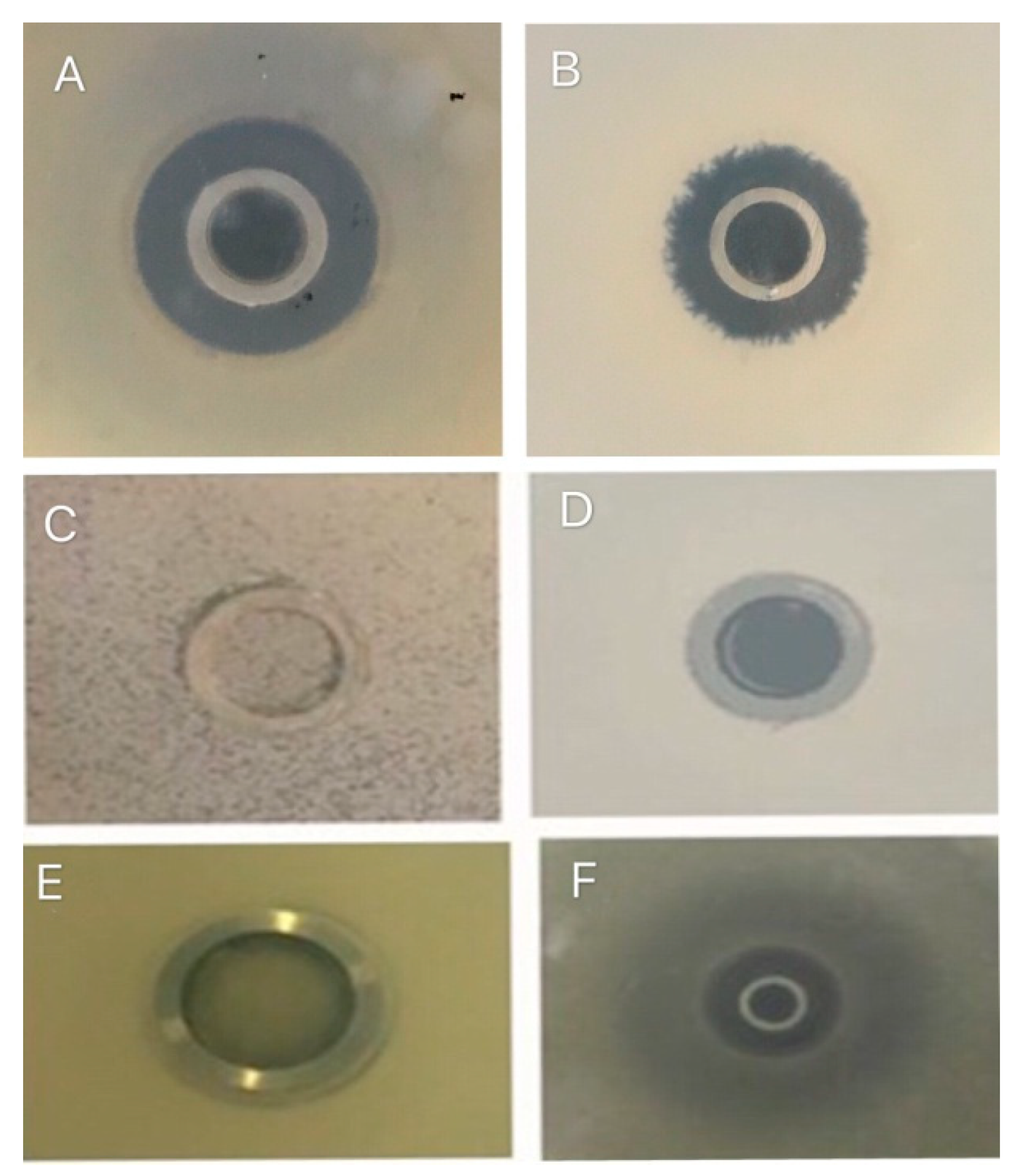
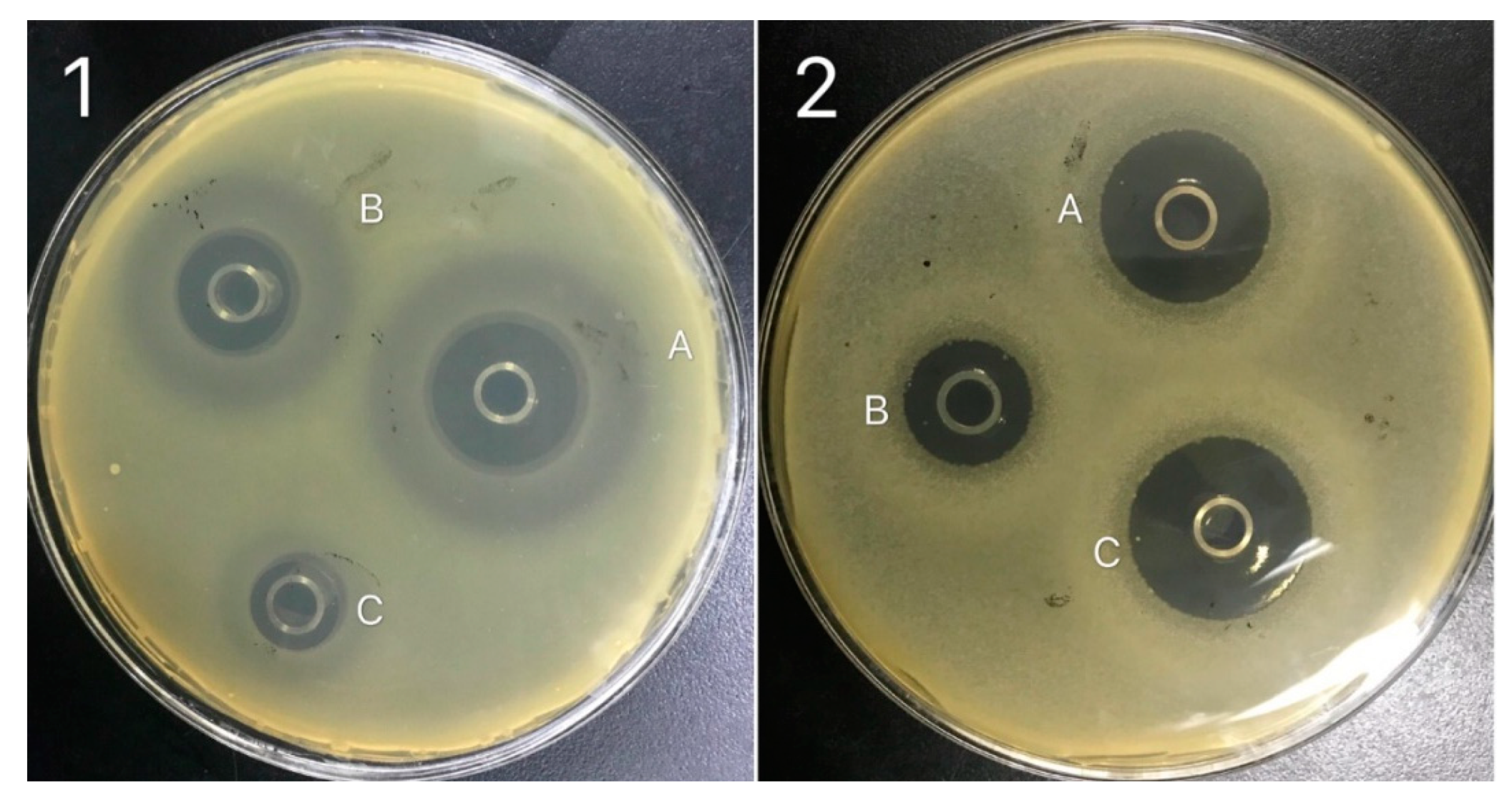
3.3. Antibacterial Activity of B. subtilis Natto
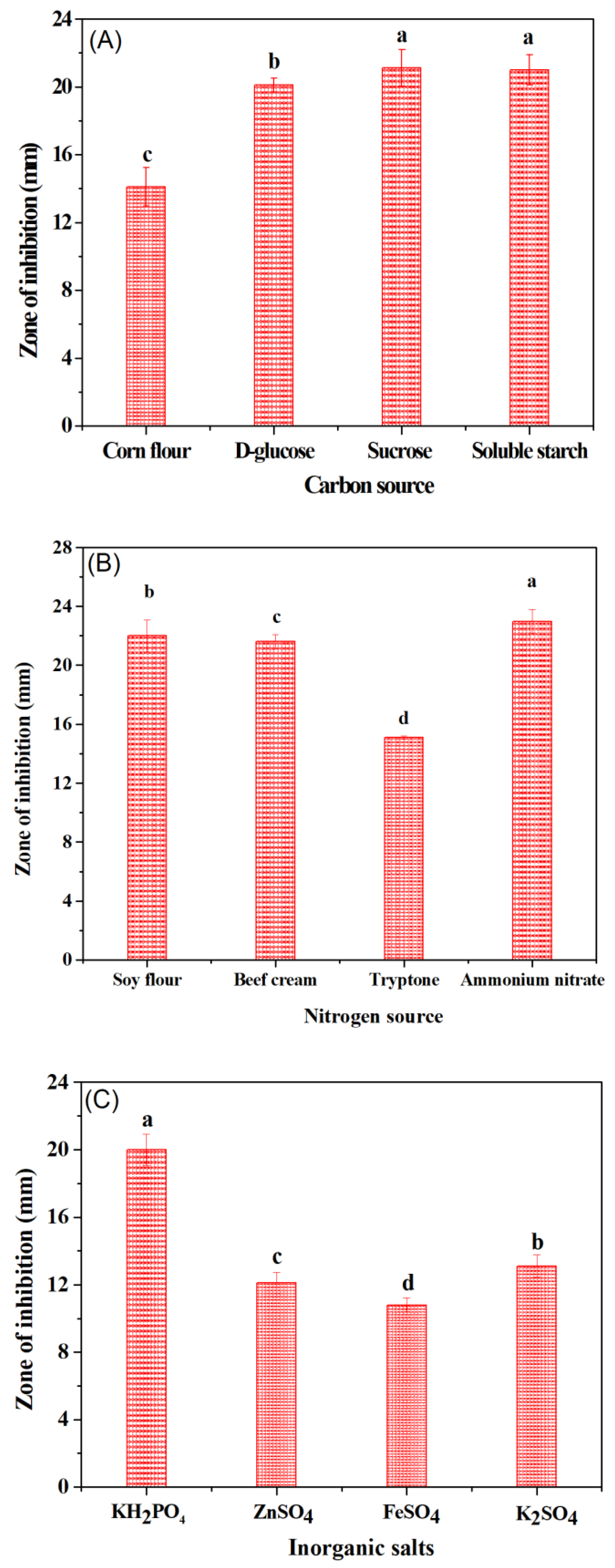
3.4. Optimization of Growth Medium to Induce the Antibacterial Activity of B. subtilis Natto
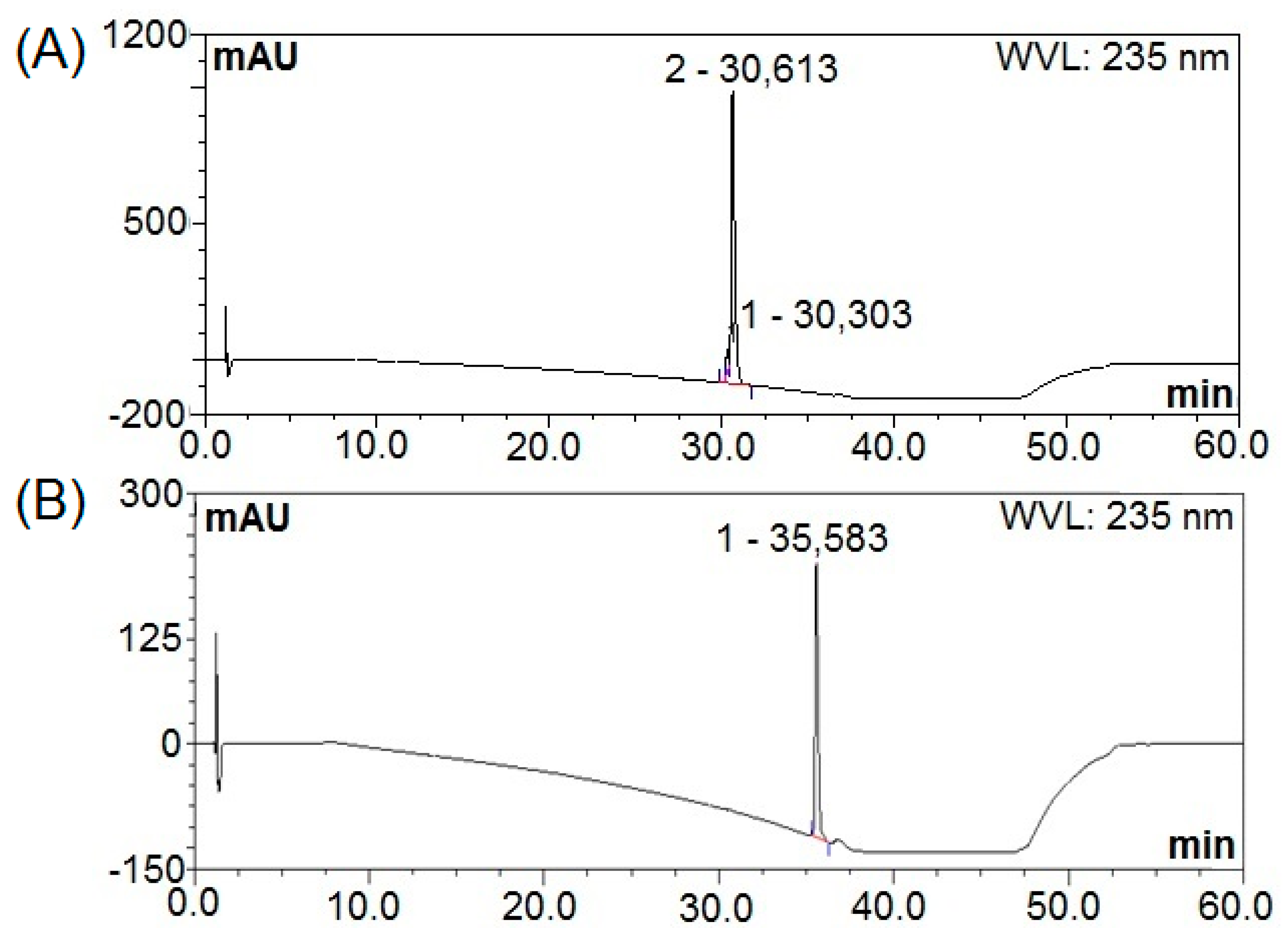
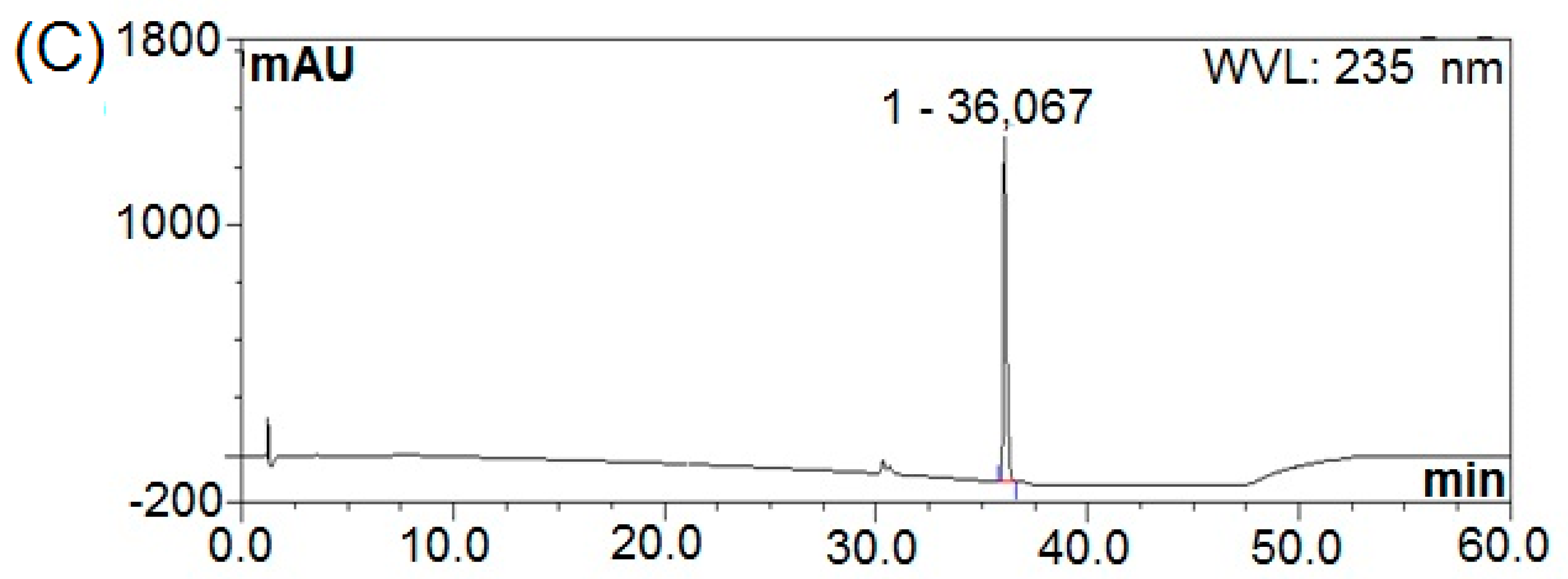
3.5. Purification of Bactericides from B. subtilis Natto
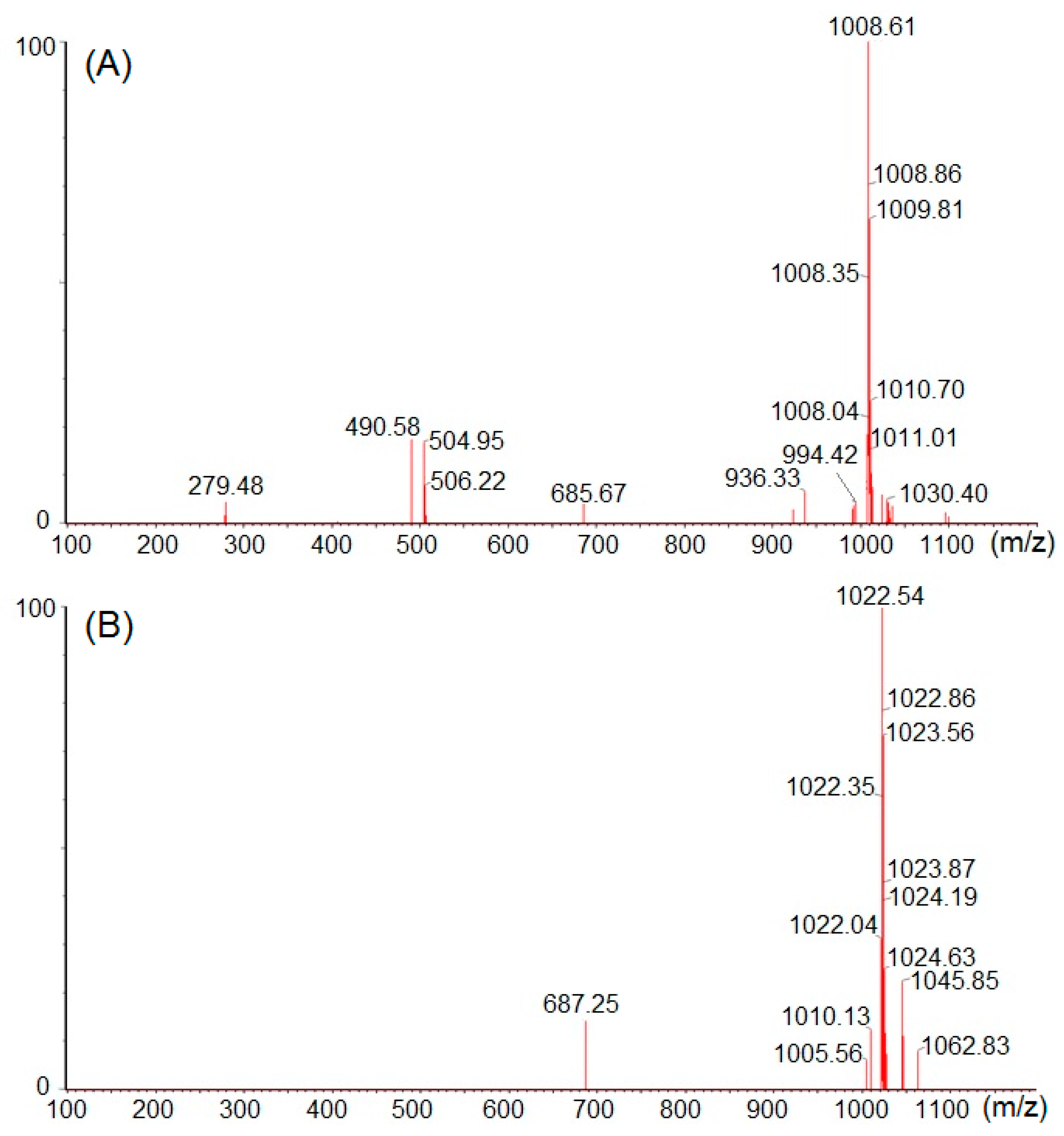
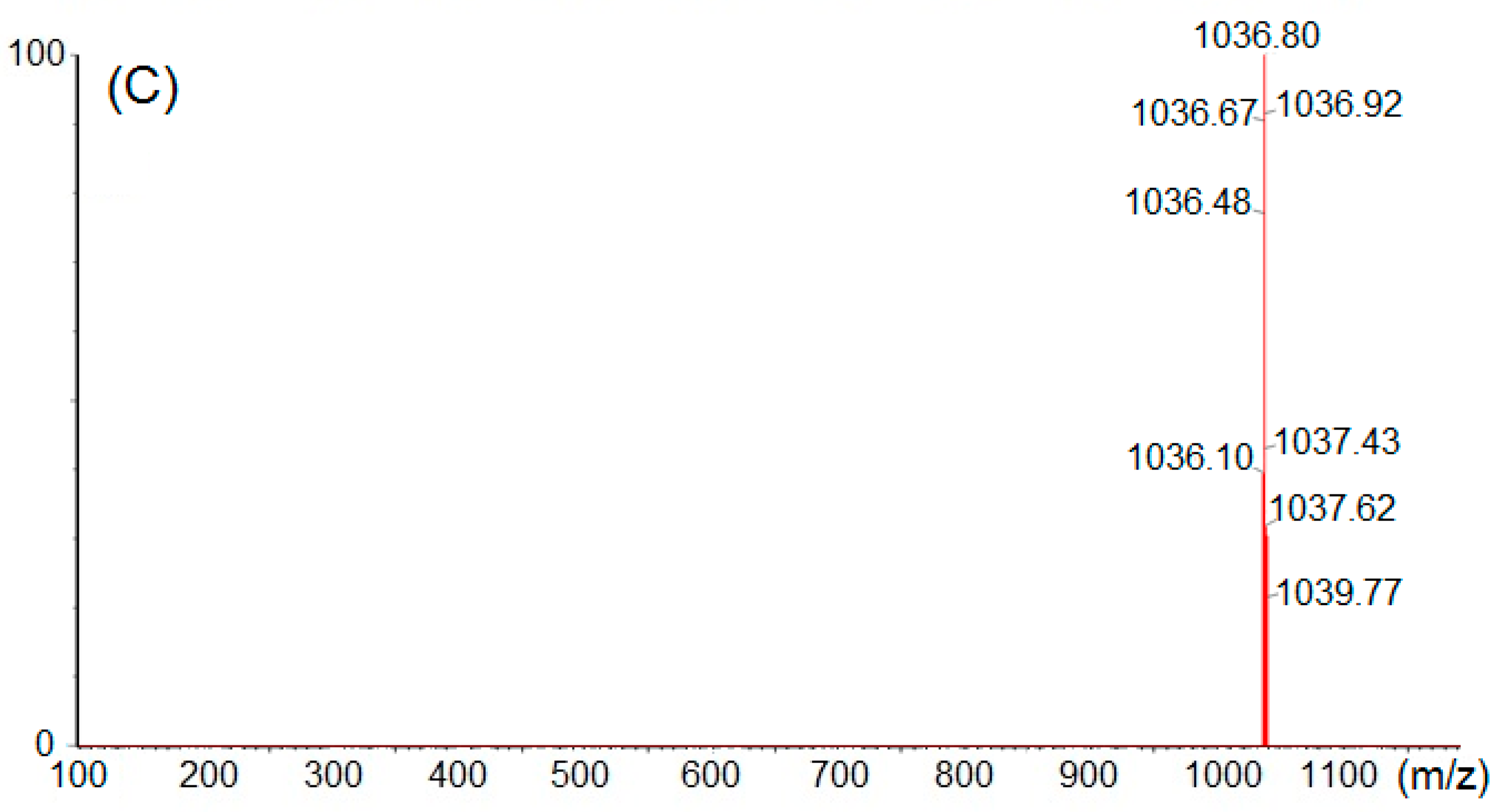
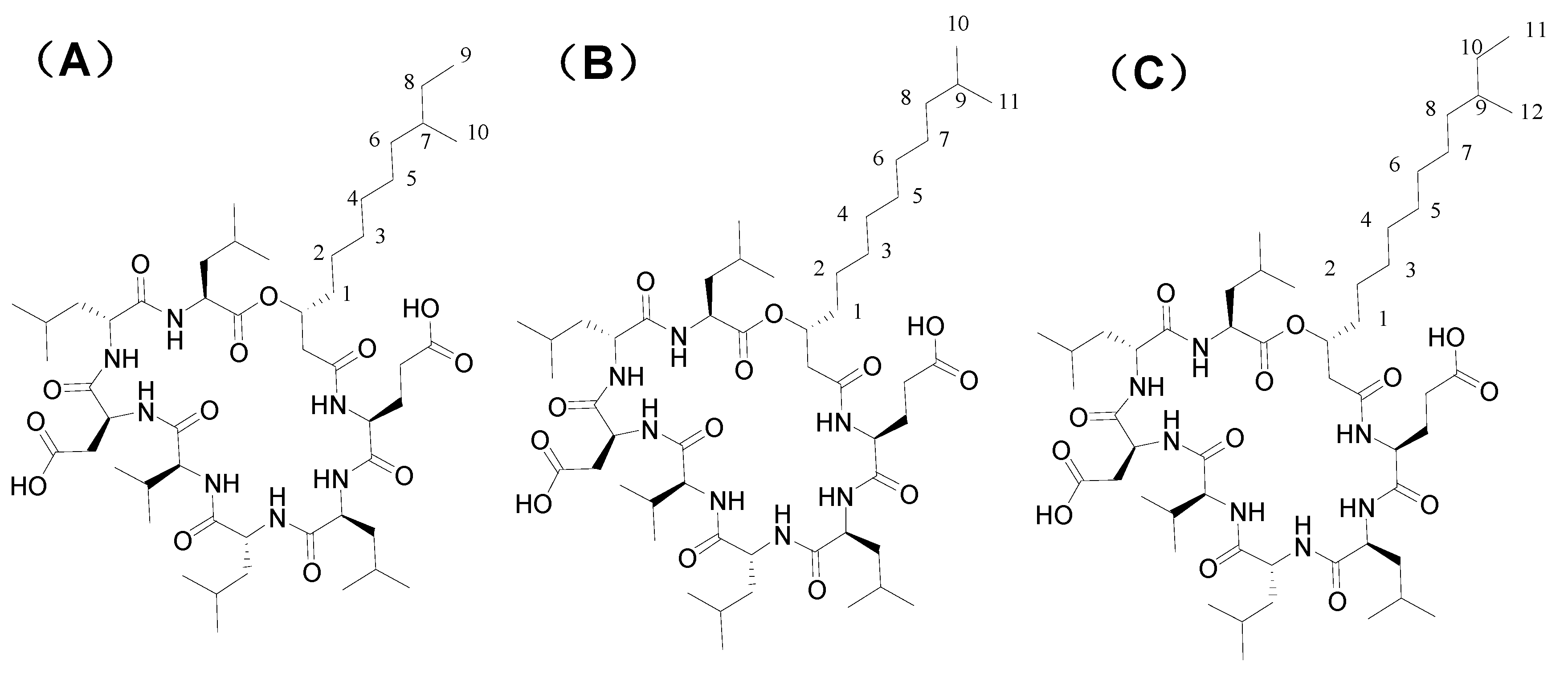
3.6. Structural Elucidation by 13C-NMR and Mass Spectral Analyses
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Tsukamoto, Y.; Ichise, H.; Kakuda, H.; Yamaguchi, M. Intake of fermented soybean (natto) increases circulating vitamin K 2 (menaquinone-7) and γ-carboxylated osteocalcin concentration in normal individuals. J. Bone Miner. Metab. 2000, 18, 216–222. [Google Scholar] [CrossRef]
- Ju, H.K.; Jun, P.Y.; Lei, Z.; Yi, P.Q. Identification of two novel fibrinolytic enzymes from Bacillus subtilis QK02. Comparative. Biochem. Physio. Part C 2004, 137, 65–74. [Google Scholar]
- Cao, X.-H.; Liao, Z.-Y.; Wang, C.; Yang, W.-Y.; Lu, M.-F. Evaluation of a lipopeptide biosurfactant from Bacillus natto TK-1 as a potential source of anti-adhesive, antimicrobial and antitumor activities. Braz. J. Microbiol. 2009, 40, 373–379. [Google Scholar] [CrossRef] [Green Version]
- Sumi, H.; Hamada, H.; Nakanishi, K.; Hiratani, H. Enhancement of the Fibrinolytic Activity in Plasma by Oral Administration of Nattokinases. Acta Haematol. 1990, 84, 139–143. [Google Scholar] [CrossRef] [Green Version]
- Tam, N.K.M.; Uyen, N.Q.; Hong, H.A.; Duc, L.H.; Hoa, T.T.; Serra, C.; Henriques, A.; Cutting, S. The Intestinal Life Cycle of Bacillus subtilis and Close Relatives. J. Bacteriol. 2006, 188, 2692–2700. [Google Scholar] [CrossRef] [Green Version]
- Tomohiro, H.; Rieko, H.; Shizue, S.; Akio, A.; Kan, K.; Shuichi, K. Cytokine responses of human intestinal epithelial-like Caco- 2 cells to the nonpathogenic bacterium Bacillus subtilis (natto). Inter. J. Food. Microbiol. 2003, 82, 255–264. [Google Scholar]
- Itaya, M.; Nagasaku, M.; Shimada, T.; Ohtani, N.; Shiwa, Y.; Yoshikawa, H.; Kaneko, S.; Tomita, M.; Sato, M. Stable and efficient delivery of DNA toBacillus subtilis(natto) using pLS20 conjugational transfer plasmids. FEMS Microbiol. Lett. 2019, 366, 032. [Google Scholar] [CrossRef] [PubMed]
- Sumi, H.; Ohsugi, T.; Ikeda, S.; Kawahara, T.; Naito, S.; Yatagai, C. ID: 101 Anti-platelet aggregation and anti-Helicobacter pylori activity of dipicolinic acid contained in Bacillus subtilis natto. J. Thromb. Haemost. 2006, 4, 217. [Google Scholar] [CrossRef]
- Zhao, X.; Zhou, Z.-J.; Han, Y.; Wang, Z.-Z.; Fan, J.; Xiao, H.-Z. Isolation and identification of antifungal peptides from Bacillus BH072, a novel bacterium isolated from honey. Microbiol. Res. 2013, 168, 598–606. [Google Scholar] [CrossRef]
- Xu, D.; Wang, Y.; Sun, L.; Liu, H.; Li, J. Inhibitory activity of a novel antibacterial peptide AMPNT-6 from Bacillus subtilis against Vibrio parahaemolyticus in shrimp. Food Control. 2013, 30, 58–61. [Google Scholar] [CrossRef]
- Mo, H.; Zhu, Y.; Chen, Z. Microbial fermented tea—A potential source of natural food preservatives. Trends Food Sci. Technol. 2008, 19, 124–130. [Google Scholar] [CrossRef]
- Schmidt, S.E.; Taylor, T.M.; Davidson, P.M. Chemical Preservatives and Natural Antimicrobial Compounds. Food Microbiol. 2013, 765–801. [Google Scholar]
- Goñi, P.; López, P.; Sanchez, C.; Gómez-Lus, R.; Becerril, R.; Nerín, C. Antimicrobial activity in the vapour phase of a combination of cinnamon and clove essential oils. Food Chem. 2009, 116, 982–989. [Google Scholar] [CrossRef]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48, 5–16. [Google Scholar] [CrossRef] [Green Version]
- Fazeli, M.R.; Amin, G.R.; Ahmadian-Attari, M.M.; Ashtiani, H.; Jamalifar, H.; Samadi, N. Antimicrobial activities of Iranian sumac and avishan-e shirazi (Zataria multiflora) against some food-borne bacteria. Food Control. 2007, 18, 646–649. [Google Scholar] [CrossRef]
- Vaquero, M.R.; Alberto, M.; De Nadra, M.C.M. Antibacterial effect of phenolic compounds from different wines. Food Control. 2007, 18, 93–101. [Google Scholar] [CrossRef]
- Pang, X.; Zhao, J.; Fang, X.; Liu, H.; Zhang, Y.; Cen, S.; Yu, L. Surfactin derivatives from Micromonospora sp. CPCC 202787 and their anti-HIV activities. J. Antibiot. 2016, 70, 105–108. [Google Scholar] [CrossRef]
- Raaijmakers, J.M.; Vlami, M.; De Souza, J.T. Antibiotic production by bacterial biocontrol agents. Antonie Leeuwenhoek 2002, 81, 537–547. [Google Scholar] [CrossRef]
- Asthana, R.K.; Deepali; Tripathi, M.K.; Srivastava, A.; Singh, A.P.; Singh, S.P.; Nath, G.; Srivastava, R.; Srivastava, B.S. Isolation and identification of a new antibacterial entity from the Antarctic cyanobacterium Nostoc CCC 537. Environ. Boil. Fishes 2008, 21, 81–88. [Google Scholar]
- Yang, L.; Quan, X.; Xue, B.; Goodwin, P.H.; Lu, S.; Wang, J.; Du, W.; Wu, C. Isolation and identification of Bacillus subtilis strain YB-05 and its antifungal substances showing antagonism against Gaeumannomyces graminis var. tritici. Boil. Control. 2015, 85, 52–58. [Google Scholar] [CrossRef]
- Bilal, M.; Rasheed, T.; Iqbal, H.M.; Hu, H.; Zhang, X. Silver Nanoparticles: Biosynthesis and Antimicrobial Potentialities. Int. J. Pharmacol. 2017, 13, 832–845. [Google Scholar] [CrossRef]
- Bilal, M.; Rasheed, T.; Iqbal, H.M.; Hu, H.; Wang, W.; Zhang, X. Macromolecular agents with antimicrobial potentialities: A drive to combat antimicrobial resistance. Int. J. Boil. Macromol. 2017, 103, 554–574. [Google Scholar] [CrossRef]
- Rasheed, T.; Bilal, M.; Iqbal, H.M.; Li, C. Green biosynthesis of silver nanoparticles using leaves extract of Artemisia vulgaris and their potential biomedical applications. Colloids Surfaces B Biointerfaces 2017, 158, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, T.; Bilal, M.; Li, C.; Iqbal, H.M. Biomedical Potentialities of Taraxacum officinale-based Nanoparticles Biosynthesized Using Methanolic Leaf Extract. Curr. Pharm. Biotechnol. 2017, 18, 1116–1123. [Google Scholar] [CrossRef] [PubMed]
- Tabbene, O.; Benslimen, I.; Djebali, K.; Mangoni, M.-L.; Urdaci, M.C.; Limam, F. Optimization of medium composition for the production of antimicrobial activity by Bacillus subtilis B38. Biotechnol. Prog. 2009, 25, 1267–1274. [Google Scholar] [CrossRef]
- Bais, H.P.; Fall, R.; Vivanco, J.M. Biocontrol of Bacillus subtilis against Infection of Arabidopsis Roots by Pseudomonas syringae Is Facilitated by Biofilm Formation and Surfactin Production1. Plant Physiol. 2004, 134, 307–319. [Google Scholar] [CrossRef] [Green Version]
- Caldeira, A.T.; Feio, S.; Arteiro, J.; Coelho, A.V.; Roseiro, J. Environmental dynamics of Bacillus amyloliquefaciens CCMI 1051 antifungal activity under different nitrogen patterns. J. Appl. Microbiol. 2008, 104, 808–816. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.-Y.; Lin, Q.; Peng, K.; Cao, J.-Z.; Yang, C.; Xu, N. Surfactin Variants from Bacillus subtilis natto CSUF5 and Their Antifungal Properities Against Aspergillus niger. J. Biobased Mater. Bioenergy 2017, 11, 210–215. [Google Scholar] [CrossRef]
- Wei, Y.; Wang, L.; Chang, J.-S. Optimizing Iron Supplement Strategies for Enhanced Surfactin Production with Bacillus subtilis. Biotechnol. Prog. 2004, 20, 979–983. [Google Scholar] [CrossRef]
- Yeh, M.-S.; Wei, Y.-H.; Chang, J.-S. Enhanced Production of Surfactin from Bacillussubtilis by Addition of Solid Carriers. Biotechnol. Prog. 2008, 21, 1329–1334. [Google Scholar] [CrossRef]
- Cooper, D.G.; Macdonald, C.R.; Duff, S.J.B.; Kosaric, N. Enhanced Production of Surfactin from Bacillus subtilis by Continuous Product Removal and Metal Cation Additions. Appl. Environ. Microbiol. 1981, 42, 408–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Das, P.; Mukherjee, S.; Sen, R. Genetic Regulations of the Biosynthesis of Microbial Surfactants: An Overview. Biotechnol. Genet. Eng. Rev. 2008, 25, 165–186. [Google Scholar] [CrossRef] [PubMed]
- Yakimov, M.M.; Timmis, K.N.; Wray, V.; Fredrickson, H.L. Characterization of a new lipopeptide surfactant produced by thermotolerant and halotolerant subsurface Bacillus licheniformis BAS50. Appl. Environ. Microbiol. 1995, 61, 1706–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiradate, S.; Yoshida, S.; Sugie, H.; Yada, H.; Fujii, Y. Mulberry anthracnose antagonists (iturins) produced by Bacillus amyloliquefaciens RC-2. Phytochemistry 2002, 61, 693–698. [Google Scholar] [CrossRef]
- Sandrin, C.; Peypoux, F.; Michel, G. Coproduction of surfactin and iturin A, lipopeptides with surfactant and antifungal properties, by Bacillus subtilis. Biotechnol. Appl. Biochem. 1990, 12, 370–375. [Google Scholar]
- Vollenbroich, D.; Pauli, G.; Ozel, M.; Vater, J. Antimycoplasma properties and application on cell cultures of surfactin, a lipopeptide antibiotic from Bacillus subtilis. Appl. Environ. Microbiol. 1997, 63, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Sinclair, J.; Hartman, G.; Bertagnolli, B. Production of iturin A by Bacillus amyloliquefaciens suppressing Rhizoctonia solani. Soil Boil. Biochem. 2002, 34, 955–963. [Google Scholar] [CrossRef]
- Cho, S.-J.; Lee, S.K.; Cha, B.J.; Kim, Y.H.; Shin, K.-S. Detection and characterization of the Gloeosporium gloeosporioides growth inhibitory compound iturin A from Bacillus subtilis strain KS03. FEMS Microbiol. Lett. 2003, 223, 47–51. [Google Scholar] [CrossRef] [Green Version]
- Lim, J.-H.; Park, B.-K.; Kim, M.-S.; Hwang, M.-H.; Rhee, M.H.; Park, S.-C.; Yun, H.-I. The anti-thrombotic activity of surfactins. J. Veter Sci. 2005, 6, 353–355. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Reaction Response | Parameter | Reaction Response |
|---|---|---|---|
| Sportiness | + | Peroxidase | + |
| Anaerobic culture | - | Hydrogen sulfide | |
| Salt tolerance 1% | + | Lecithinase | + |
| 3% | + | Using citrate | + |
| 5% | + | Use of malonate | + |
| 7% | + | Nitrate reduction | + |
| 9% | - | Indigo matrix production | - |
| V-P experiment | + | Sticky brushed | + |
| Starch hydrolysis | + | Catalase | + |
| Gelatin liquefaction | + |
| Level | Factor | ||
|---|---|---|---|
| Sucrose | Soy Flour | Potassium Sulfate | |
| 1 | 1% | 0.20% | 0.20% |
| 2 | 2% | 0.50% | 0.50% |
| 3 | 3% | 0.80% | 0.80% |
| Factor | Sucrose (A) | Ammonium Nitrate (B) | KH2PO4 (C) | Inhibition Zone (mm) |
|---|---|---|---|---|
| Experiment 1 | 1.0 | 1.0 | 1.0 | 19 |
| Experiment 2 | 1.0 | 2.0 | 2.0 | 23 |
| Experiment 3 | 1.0 | 3.0 | 3.0 | 22 |
| Experiment 4 | 2.0 | 1.0 | 2.0 | 16 |
| Experiment 5 | 2.0 | 2.0 | 3.0 | 20 |
| Experiment 6 | 2.0 | 3.0 | 1.0 | 27 |
| Experiment 7 | 3.0 | 1.0 | 3.0 | 21 |
| Experiment 8 | 3.0 | 2.0 | 1.0 | 22 |
| Experiment 9 | 3.0 | 3.0 | 2.0 | 20 |
| K1 | 6.0 | 5.5 | 6.3 | - |
| K2 | 6.0 | 6.1 | 5.7 | - |
| K3 | 6.0 | 6.4 | 6.0 | - |
| k1 | 2.0 | 1.83 | 2.1 | - |
| k2 | 2.0 | 2.03 | 1.9 | - |
| k3 | 2.0 | 2.13 | 2.0 | - |
| R | 0.0 | 0.90 | 0.6 | - |
| Position | NT6 | NT-7 | NT-5 |
|---|---|---|---|
| 1 Glu C = O | 171.0 | 171.0 | 171.0 |
| α-C | 52.4 | 52.4 | 52.4 |
| β-C | 27.1 | 27.1 | 27.1 |
| γ-C | 29.7 | 29.7 | 29.7 |
| δ-C = O | 173.9 | 173.9 | 173.9 |
| -OCH3 | - | - | - |
| 2 Leu C = O | 172.6 | 172.6 | 172.6 |
| α-C | 51.9 | 51.9 | 51.9 |
| β-C | 40.0 | 40.0 | 40.0 |
| γ-C | 24.2 | 24.2 | 24.2 |
| δ-C | 23.0/22.9 | 23.0/22.9 | 23.0/22.9 |
| 3 Leu C = O | 172.9 | 172.9 | 172.9 |
| α-C | 51.8 | 51.8 | 51.8 |
| β-C | 38.9 | 38.9 | 38.9 |
| γ-C | 24.2 | 24.2 | 24.2 |
| δ-C | 22.9/22.6 | 22.9/22.6 | 22.9/22.6 |
| 4 Val C = O | 170.7 | 170.7 | 170.7 |
| α-C | 58.6 | 58.6 | 58.6 |
| β-C | 30.1 | 30.1 | 30.1 |
| γ-C | 19.1/17.9 | 19.1/17.9 | 19.1/17.9 |
| 5 Asp C = O | 169.9 | 169.9 | 169.9 |
| α-C | 49.6 | 49.6 | 49.6 |
| β-C | 35.7 | 35.7 | 35.7 |
| γ-C = O | 171.7 | 171.7 | 171.7 |
| -OCH3 | - | - | - |
| 6 Leu C = O | 171.6 | 171.6 | 171.6 |
| α-C | 50.8 | 50.8 | 50.8 |
| β-C | 41.6 | 41.6 | 41.6 |
| γ-C | 24.2 | 24.2 | 24.2 |
| δ-C | 21.7/21.7 | 21.7/21.7 | 21.7/21.7 |
| 7 Leu/Val C = O | 171.9 | 171.9 | 171.9 |
| α-C | 50.8 | 50.8 | 50.8 |
| β-C | 38.8 | 38.8 | 38.8 |
| γ-C | 24.0 | 24.0 | 24.0 |
| δ-C | 21.2/21.1 | 21.2/21.1 | 21.2/21.1 |
| Fatty acid-1 | 169.9 | 169.9 | 169.9 |
| 2 | 41.1 | 41.1 | 36.0 |
| 3 | 71.4 | 71.4 | 71.4 |
| 4 | 33.6 | 33.6 | 33.6 |
| 5 | 24.3 | 24.3 | 24.3 |
| 6 | 28.6-29.3 | 28.6-29.4 | 28.6-29.4 |
| 7 | 28.6-29.3 | 28.6-29.4 | 28.6-29.4 |
| 8 | 28.6-29.3 | 28.6-29.4 | 28.6-29.4 |
| 9 | 28.6-29.3 | 28.6-29.4 | 28.6-29.4 |
| 10 | 26.8 | 28.6-29.4 | 33.7 |
| 11 | 38.5 | 36.0 | 26.4 |
| 12 | 27.4 | 33.7 | 11.2 |
| 13 | 22.5 | 26.4 | 19.1 |
| 14 | 22.5 | 11.2 | - |
| 15 | - | 19.1 | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Bilal, M.; Liu, S.; Zhang, J.; Lu, H.; Luo, H.; Luo, C.; Shi, H.; Iqbal, H.M.N.; Zhao, Y. Isolation, Identification and Antimicrobial Evaluation of Bactericides Secreting Bacillus subtilis Natto as a Biocontrol Agent. Processes 2020, 8, 259. https://doi.org/10.3390/pr8030259
Zhang J, Bilal M, Liu S, Zhang J, Lu H, Luo H, Luo C, Shi H, Iqbal HMN, Zhao Y. Isolation, Identification and Antimicrobial Evaluation of Bactericides Secreting Bacillus subtilis Natto as a Biocontrol Agent. Processes. 2020; 8(3):259. https://doi.org/10.3390/pr8030259
Chicago/Turabian StyleZhang, Jing, Muhammad Bilal, Shuai Liu, Jiaheng Zhang, Hedong Lu, Hongzhen Luo, Chuping Luo, Hao Shi, Hafiz M. N. Iqbal, and Yuping Zhao. 2020. "Isolation, Identification and Antimicrobial Evaluation of Bactericides Secreting Bacillus subtilis Natto as a Biocontrol Agent" Processes 8, no. 3: 259. https://doi.org/10.3390/pr8030259
APA StyleZhang, J., Bilal, M., Liu, S., Zhang, J., Lu, H., Luo, H., Luo, C., Shi, H., Iqbal, H. M. N., & Zhao, Y. (2020). Isolation, Identification and Antimicrobial Evaluation of Bactericides Secreting Bacillus subtilis Natto as a Biocontrol Agent. Processes, 8(3), 259. https://doi.org/10.3390/pr8030259