Propagation and Molecular Characterization of Fowl Adenovirus Serotype 8b Isolates in Chicken Embryo Liver Cells Adapted on Cytodex™ 1 Microcarrier Using Stirred Tank Bioreactor


 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Virus Isolates
2.3. Siliconization of Glassware
2.4. Setting up the BIOSTAT® B Bioreactor
2.5. Preparation of Cytodex™ 1 Microcarrier Beads
2.6. Preparation of Chicken Embryo Liver Cells
2.7. Adaptation of CEL Cells to Cytodex™ 1 Microcarrier Culture
2.8. Propagation of FAdV in Cytodex™ 1 Microcarrier Adapted CEL Cells in STB
2.9. Determination of the Infective Dose (TCID50) of the Passage Isolates
2.10. Indirect Immunofluorescence Assay
2.11. DNA Extraction and PCR Amplification of Hexon and Fiber Genes
2.12. Sequencing, Blast, Alignment, and Phylogentic Analysis
3. Results
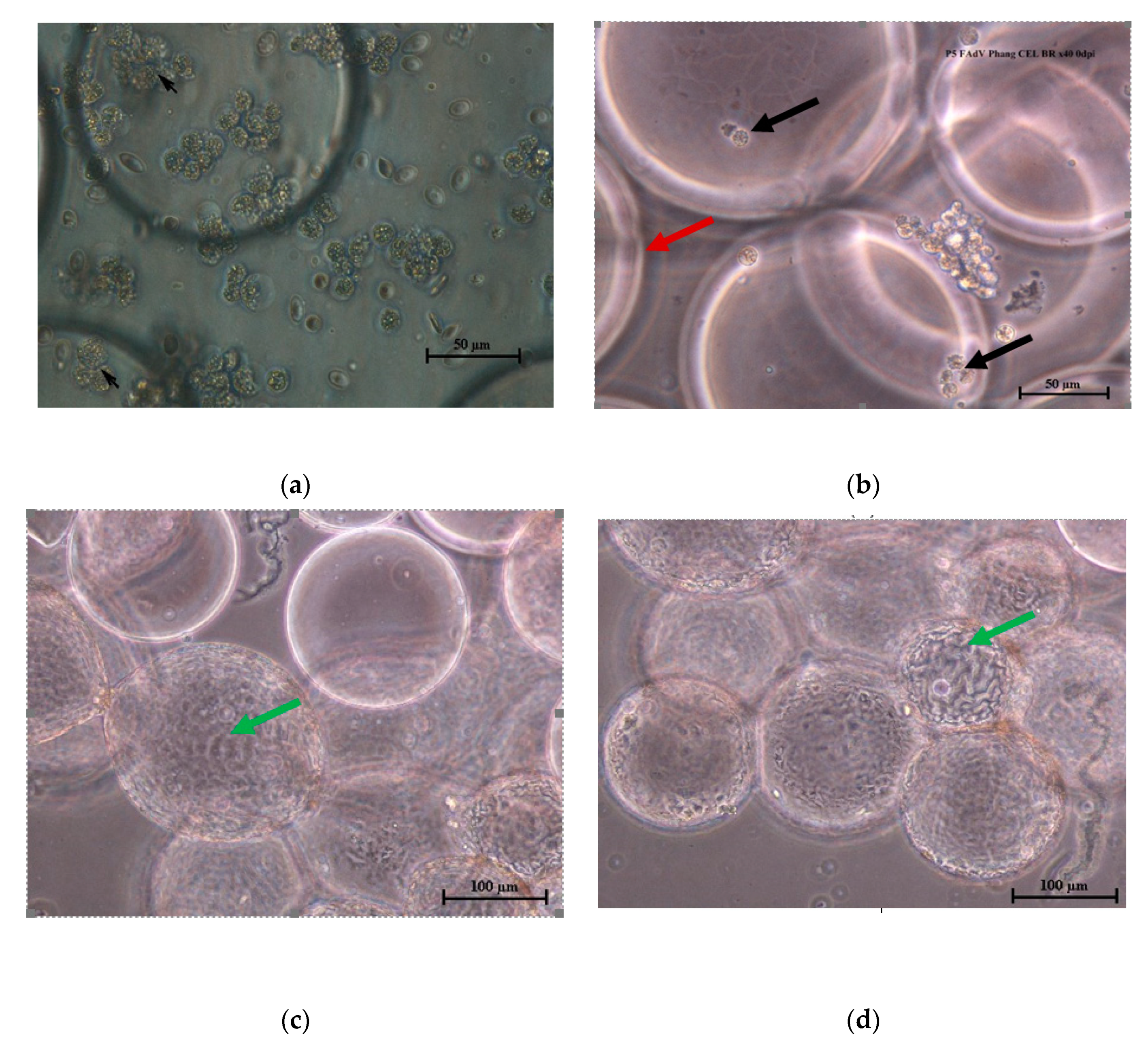
3.1. Adaptation of CEL Cells on Cytodex™ 1 Microcarriers
3.2. Propagation of FAdV in CEL Cells Adapted on Cytodex™ 1 Microcarrier
3.3. Infective Dose (TCID50) of UPM08136P5B1 and UPM08136P20B1 Isolates
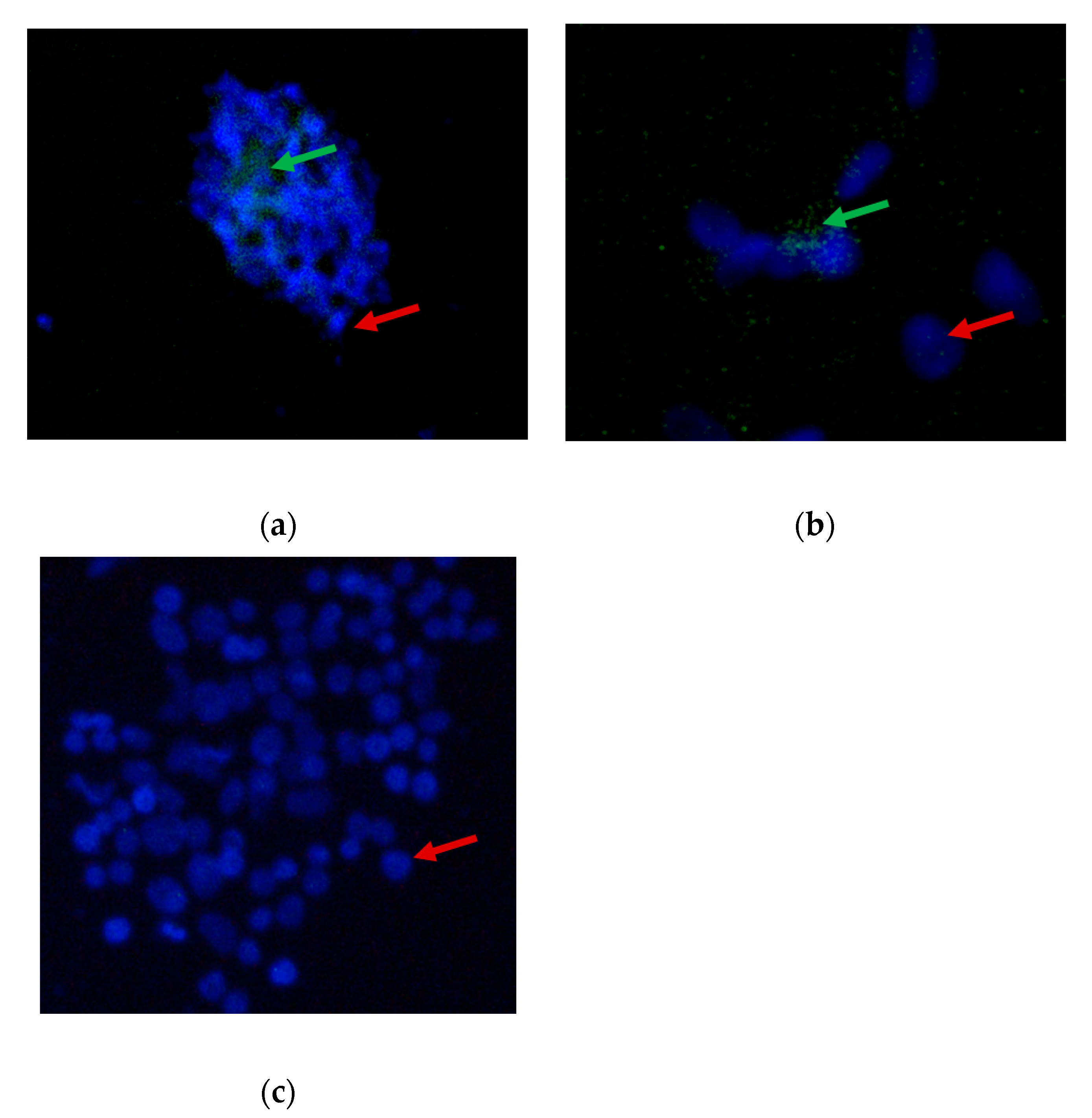
3.4. Indirect Immunofluorescence Assay of FAdV Isolates Propagated in CEL Cells Adapted Cytodex™ 1 Microcarrier in a Stirred Tank Bioreactor
3.5. Polymerase Chain Reaction Amplification of Hexon and Fiber Genes of the Propagated FAdV Isolates in CEL Cells Adapted on Cytodex™ 1 Microcarrier
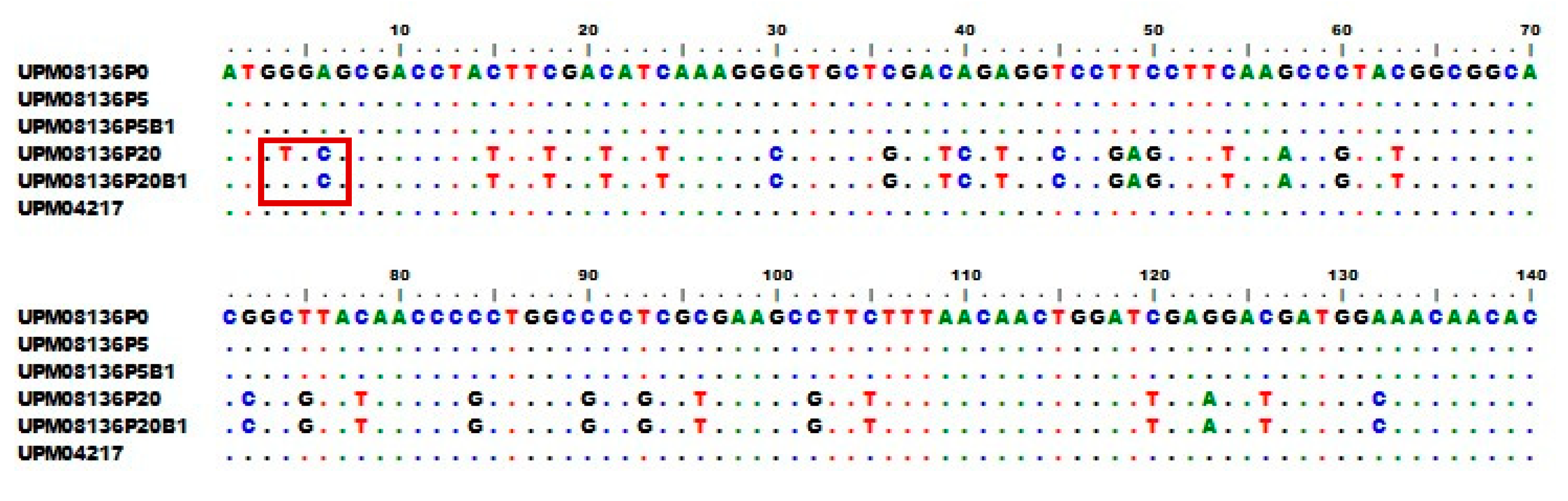
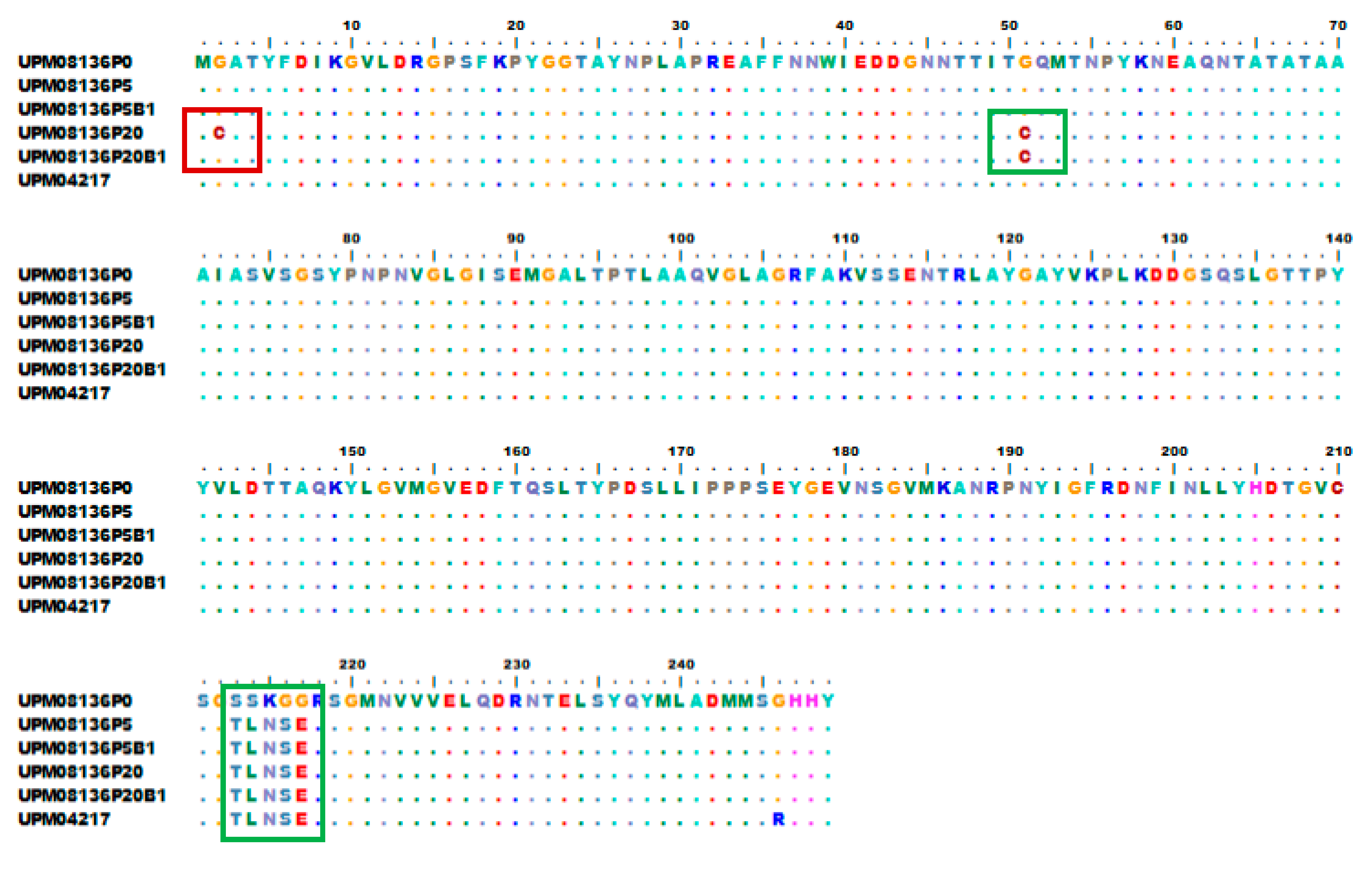
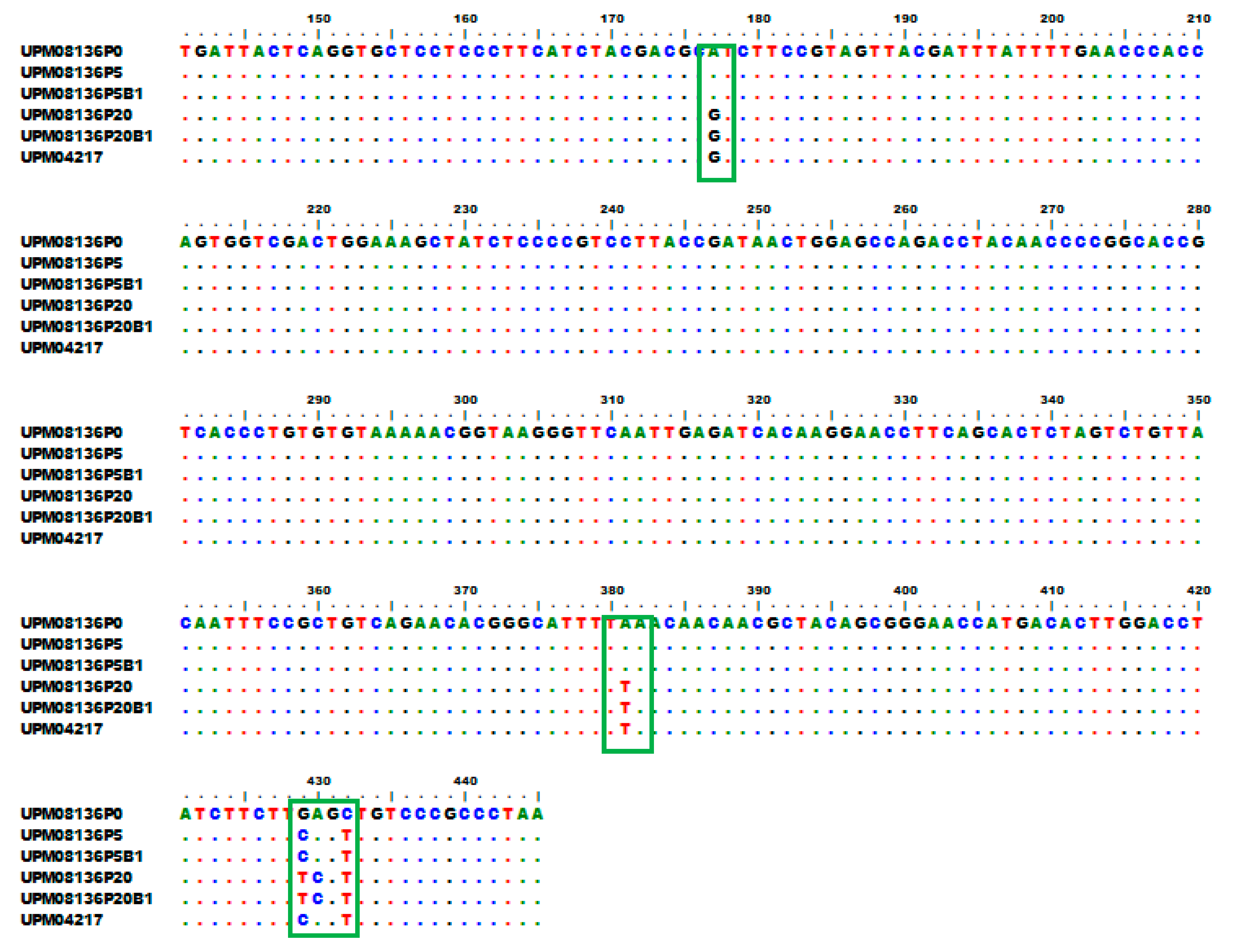
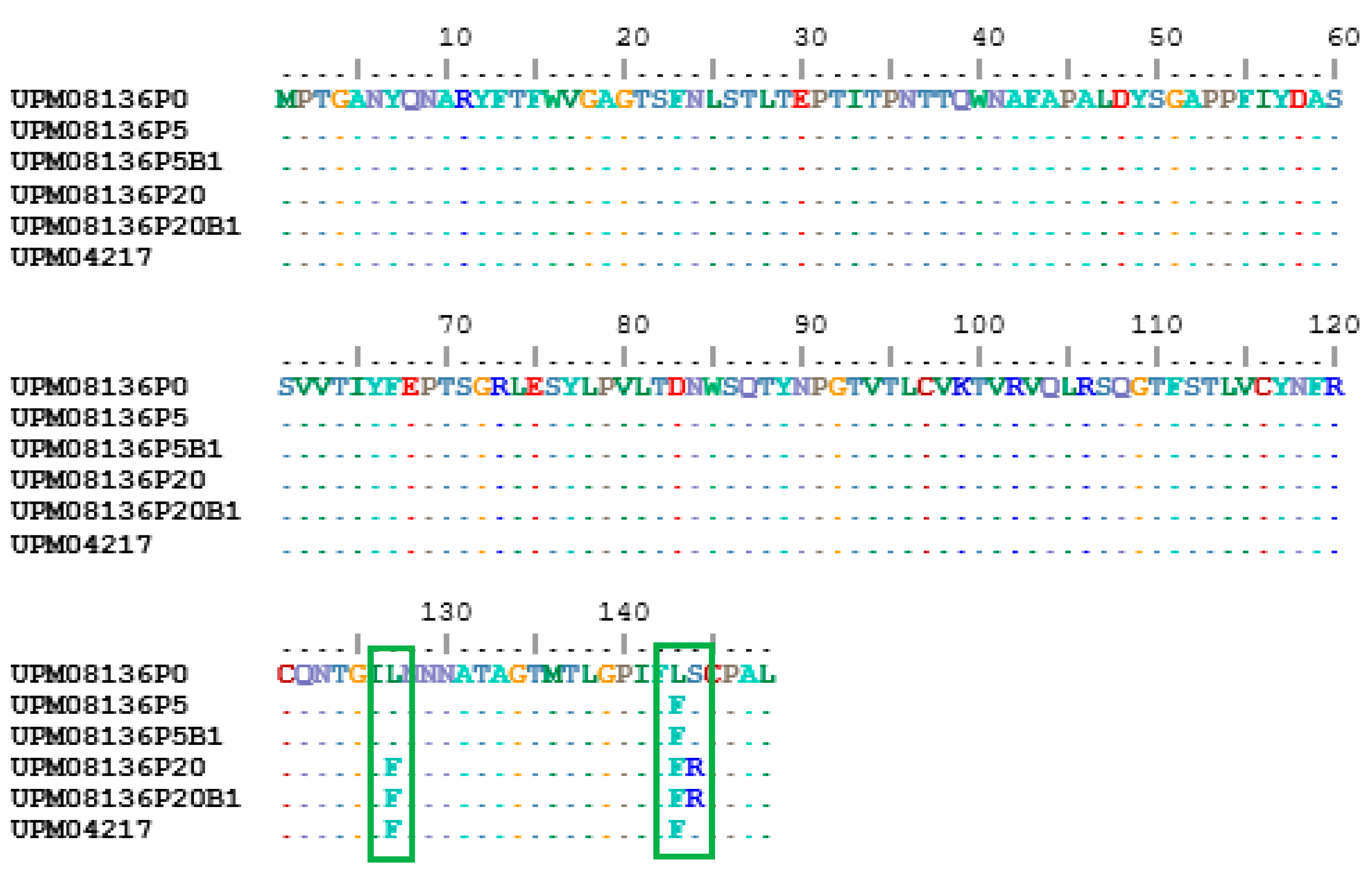
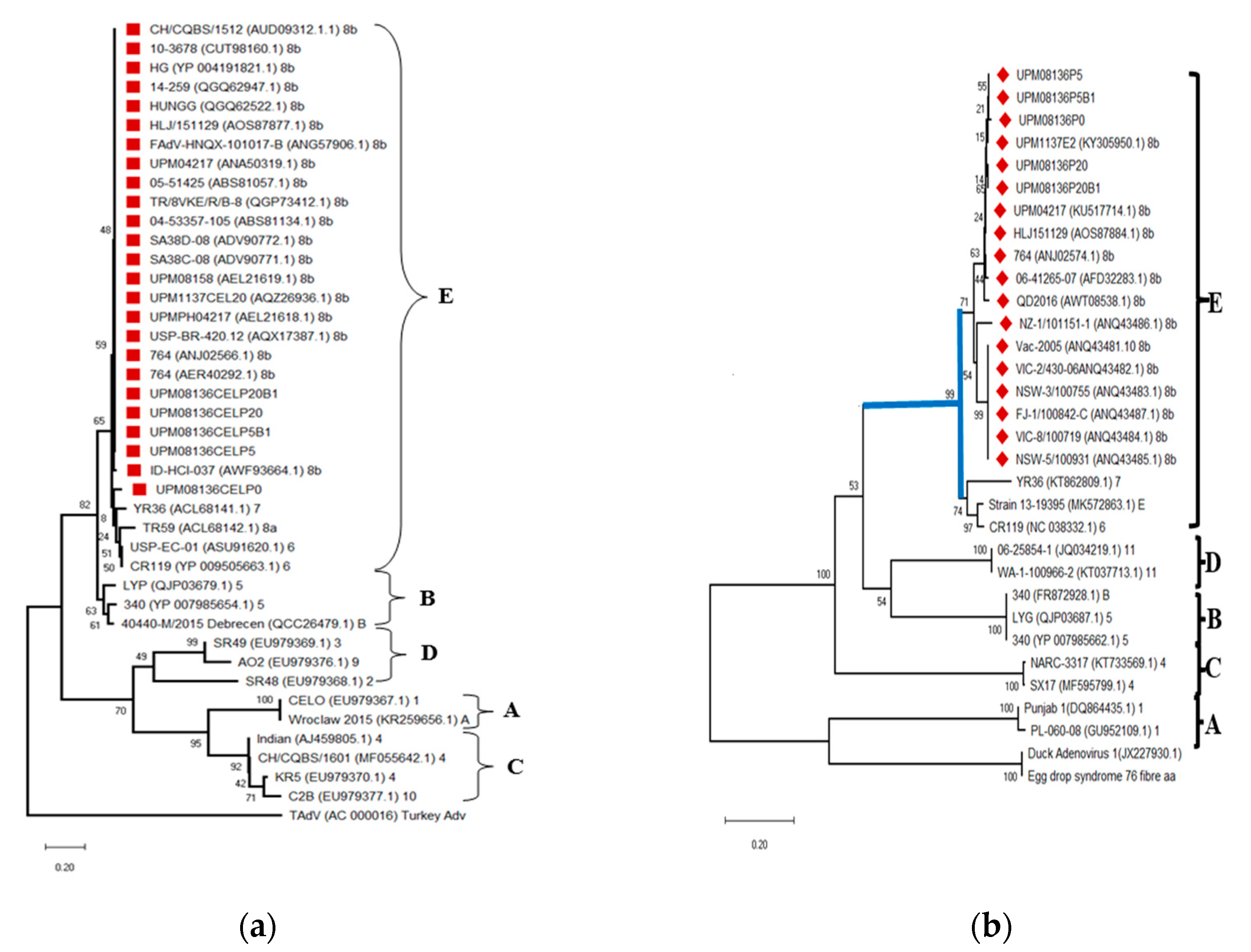
3.6. Multisequence Alignment and Phylogenetic Analysis of UPM08136CELP5B1 and UPM08136CELP20B1 with Reference Strains
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Dar, A.; Gomis, S.; Shirley, I.; Mutwiri, G.; Brownlie, R.; Potter, A.; Gerdts, V.; Tikoo, S.K. Pathotypic and Molecular Characterization of A Fowl Adenovirus Associated With Inclusion Body Hepatitis in Saskatchewan Chickens. Avian Dis. 2012, 56, 73–81. [Google Scholar] [CrossRef]
- Hair-Bejo, M. Inclusion Body Hepatitis in A Flock of Commercial Broiler Chickens. J. Vet. Malays. 2005, 17, 23–26. [Google Scholar]
- Norina, L.; Norsharina, A.; Nurnadiah, A.H.; Redzuan, I.; Ardy, A.; Nor-Ismaliza, I. Avian Adenovirus Isolated from Broiler Affected with Inclusion Body Hepatitis. Malays. J. Vet. Res. 2016, 7, 121–126. [Google Scholar]
- García, M. Current and Future Vaccines and Vaccination Strategies Against Infectious Laryngotracheitis (ILT) Respiratory Disease of Poultry. Vet. Microbiol. 2017, 206, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Minor, P.D. Live attenuated vaccines: Historical Successes and Current Challenges. Virology 2015, 479–480, 379–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinman, A.R.; Orenstein, W.A.; Santoli, J.M.; Rodewald, L.E.; Cochi, S.L. Vaccine Shortages: History, Impact, and Prospects for the Future. Annu. Rev. Public Health 2006, 27, 235–259. [Google Scholar] [CrossRef]
- Whitford, W.G.; Fairbank, A. Considerations in Scale-Up of Viral Vaccine Production Bioprocess International. 2011. Available online: https://bioprocessintl.com/manufacturing/monoclonal-antibodies/considerations-in-scale-up-of-viral-vaccine-production-320990/ (accessed on 25 May 2019).
- Haredy, A.M.; Takenaka, N.; Yamada, H.; Sakoda, Y.; Okamatsu, M.; Yamamoto, N.; Omasa, T.; Ohtake, H.; Mori, Y.; Kida, H.; et al. An MDCK Cell Culture-Derived Formalin-Inactivated Influenza Virus Whole-Virion Vaccine from an Influenza Virus Library Confers Cross-Protective Immunity by Intranasal Administration in Mice. Clin. Vaccine Immunol. 2013, 20, 998–1007. [Google Scholar] [CrossRef] [Green Version]
- Whitford, W. Using Disposables in Cell-Culture–Based Vaccine Production. BioProcess Int. 2010, 8, S20–S27. [Google Scholar]
- Swain, P.; Nanda, P.K.; Natak, S.K.; Mishra, S.S. Basic techniques and limitations in establishing cell culture: A mini review. Adv. Anim. Vet. Sci. 2014, 2, 1–10. [Google Scholar] [CrossRef]
- Moreira, A.R. The Evolution of Protein Expression and Cell Culture. BioPharm Int. 2007, 20. Available online: http://www.biopharminternational.com/evolution-protein-expression-and-cell-culture (accessed on 14 May 2019).
- Marquis, C.P. Mammalian Cell Culture. Biotechnology 2019, 1. Available online: https://www.eolss.net/Sample-Chapters/C17/E6-58-01-04.pdf (accessed on 14 May 2019).
- Ryan, J.A. Evolution of Cell Culture Surfaces. BioFiles 2008, 3.8, 21. [Google Scholar]
- Merten, O.W. Advances in Cell Culture: Anchorage Dependence. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2015, 370, 20140040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popovic, M.K.; Poertner, R. Bioreactors and Cultivation Systems for Cell and Tissue Culture. In Encyclopedia of Life Support Systems; Doelle, H.W., Rokem, S., Berovic, M., Eds.; Eolss Publications: Oxford, UK, 2011. [Google Scholar] [CrossRef]
- Blüml, G. Microcarrier Cell Culture Technology. In Animal Cell Biotechnology; Methods in Biotechnology; Pörtner, R., Ed.; Humana Press: Louisville, KY, USA, 2007; Volume 24. [Google Scholar] [CrossRef]
- Kluge, J.A.; Leisk, G.G.; Kaplan, D.L. Mechanical Determinants of Tissue Development. In Principles of Regenerative Medicine; Academic Press: Cambridge, MA, USA, 2008; pp. 480–497. [Google Scholar] [CrossRef]
- Ali, S.; Rafique, A.; Ahmed, M.; Sakander, S. Different Types of Industrial Fermenters and Their Associated Operations for The Mass Production of Metabolites. Eur. J. Pharm. Med. Res. 2018, 5, 109–119. [Google Scholar]
- Surabattula, R.R.; Sambasiva, K.R.S.; Polavarapu, R. An Optimized Process for Expression, Scale-Up and Purification of Recombinant Erythropoietin Produced in Chinese Hamster Ovary Cell Culture. Res. Biotechnol. 2011, 2, 58–74. [Google Scholar]
- Lawal, N.; Hair-Bejo, M.; Arshad, S.S.; Omar, A.R.; Ideris, A. Propagation and Molecular Characterization of Bioreactor Adapted Very Virulent Infectious Bursal Disease Virus Isolates of Malaysia. J. Pathog. 2018. [Google Scholar] [CrossRef]
- Tapia, F.; Jordan, I.; Genzel, Y.; Reichl, U. Efficient and Stable Production of Modified Vaccinia Ankara Virus in Two-Stage Semi-Continuous and in Continuous Stirred Tank Cultivation Systems. PLoS ONE 2017, 12, e0182553. [Google Scholar] [CrossRef] [Green Version]
- Mancuso, F.; Shi, J.; Malik, D. High Throughput Manufacturing of Bacteriophages Using Continuous Stirred Tank Bioreactors Connected in Series to Ensure Optimum Host Bacteria Physiology for Phage Production. Viruses 2018, 10, 537. [Google Scholar] [CrossRef] [Green Version]
- Noteboom, W.D.; Will, P.C. Siliconizing Glassware to Be Used for Suspension Cell Culture. J. Tissue Cult. Methods 1982, 7, 9–11. [Google Scholar] [CrossRef]
- Soumyalekshmi, S.; Ajith, M.K.; Chandraprakash, M. Isolation of Fowl Adenovirus in Chicken Embryo Liver Cell Culture and Its Detection by Hexon Gene-Based PCR. Indian J. Sci. Technol. 2014, 2, 33–36. [Google Scholar]
- Sohaimi, N.M.; Bejo, M.H.; Omar, A.R.; Ideris, A.; Isa, N.M. Hexon and Fiber Gene Changes in an Attenuated Fowl Adenovirus Isolate from Malaysia in Embryonated Chicken Eggs and Its Infectivity in Chickens. J. Vet. Sci. 2018, 19, 759. [Google Scholar] [CrossRef] [PubMed]
- Zhihong, C.; Yinliang, C.; Yuan, S.; Xuepin, Y.; Guangzi, Y. Microcarrier Culture of Fish Cells and Viruses in Cell Culture Bioreactor. In Biochemical Engineering for 2001; Furusaki, S., Endo, I., Matsuno, R., Eds.; Springer: Tokyo, Japan, 1992; pp. 335–338. [Google Scholar]
- Möller, L.; Schünadel, L.; Nitsche, A.; Schwebke, I.; Hanisch, M.; Laue, M. Evaluation of Virus Inactivation by Formaldehyde to Enhance Biosafety of Diagnostic Electron Microscopy. Viruses 2015, 7, 666–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, L.J.; Muench, H.A. A Simple Method of Estimating Fifty Percent Endpoints. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Meulemans, G.; Boschmans, M.; van den Berg, T.P.; Decaesstecker, M. Polymerase Chain Reaction Combined with Restriction Enzyme Analysis for Detection and Differentiation of Fowl Adenoviruses. Avian Pathol. 2001, 30, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Maza, F.; Maldonado, J.; Vásquez-Dean, J.; Mandakovic, D.; Gaete, A.; Cambiazo, V.; González, M. Soil Bacterial Communities from the Chilean Andean Highlands: Taxonomic Composition and Culturability. Front. Bioeng. Biotechnol. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. Bioedit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, V.; Singh, G.; Sangwan, P.; Verma, A.K.; Agrawal, S. Cloning, Sequencing, and In Silico Analysis of-Propeller Phytase Bacillus licheniformis Strain PB-13. Biotechnol. Res. Int. 2014. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The Neighbor-Joining Method: A New Method for Reconstructing Phylogenetic Trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Nilsson, K. Microcarrier Cell Culture. Biotechnol. Genet. Eng. Rev. 1988, 6, 404–439. [Google Scholar] [CrossRef]
- van Wezel, A.L. Cultivation of Anchorage Dependent Cells and Their Applications. J. Chem. Technol. Biotechnol. 1982, 32, 318–323. [Google Scholar] [CrossRef]
- Frauenschuh, S.; Reichmann, E.; Ibold, Y.; Goetz, P.M.; Sittinger, M.; Ringe, J. A Microcarrier-Based Cultivation System for Expansion of Primary Mesenchymal Stem Cells. Biotechnol. Prog. 2007, 23, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Hirtenstein, M.; Clark, J.; Lindgren, G.; Vretblad, P. Microcarriers for Animal Cell Culture: A Brief Review of Theory and Practice. Dev. Biol. Stand. 1980, 46, 109–116. [Google Scholar] [PubMed]
- Mendonça, R.Z.; de Oliveira, M.I.; de Andrade Vaz-de-Lima, L.R.; Mendonça, R.M.Z.; Andrade, G.P.; Pereira, C.A.; Hoshino-Shimizu, S. Effect of Cell Culture System on the Production of Human Viral Antigens. J. Bras. Patol. Med. Lab. 2004, 40, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Asthana, M.; Singh, V.K.; Kumar, R.; Chandra, R. Isolation, Cloning and In Silico Study of Hexon Gene of Fowl Adenovirus 4 (FAV4) Isolates Associated with Hydro Pericardium Syndrome in Domestic Fowl. J Proteom. Bioinform 2011, 4, 190–195. [Google Scholar] [CrossRef] [Green Version]
- Heaton, N.S. Revisiting the Concept of a Cytopathic Viral Infection. PLoS Pathog 2017, 13, e1006409. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.; Poh, S.L.; Dietzsch, C.; Roethl, E.; Yan, M.L.; Ng, S.K. Serum-free Microcarrier Based Production of Replication Deficient Influenza Vaccine Candidate Virus Lacking NS1 Using Vero Cells. BMC Biotechnol. 2011, 11, 81. [Google Scholar] [CrossRef] [Green Version]
- Trabelsi, K.; Majoul, S.; Rourou, S.; Kallel, H. Development of a measles vaccine production process in MRC-5 cells grown on Cytodex1 microcarriers and in a stirred bioreactor. Appl. Microbiol. Biotechnol. 2014, 93, 1031–1040. [Google Scholar] [CrossRef]
- Jordan, B.A.; Blake, L.; Bisnath, J.; Ramgattie, C.; Carrington, C.V.; Oura, C.A.L. Identification of Four Serotypes of Fowl Adenovirus in Clinically Affected Commercial Poultry Co-Infected with Chicken Infectious Anaemia Virus in Trinidad and Tobago. Transbound. Emerg. Dis. 2019, 66, 1341–1348. [Google Scholar] [CrossRef]
- De la Torre, D.I.; Nuñez, L.F.; Astolfi-Ferreira, C.S.; Ferreira, A.J.P. Enteric Virus Diversity Examined by Molecular Methods in Brazilian Poultry Flocks. Vet. Sci. 2018, 5, 38. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xia, W.; Mao, S.; Lu, S.; Mo, K.; Liao, M.; Zhou, J.; Zheng, X. Isolation, Identification and Whole Genome Sequence Analysis of Serotype 4 Fowl Adenovirus Zhejiang Strain. Anim. Sci. Vet. Med. 2019, 45, 635–646. [Google Scholar] [CrossRef]
- Pauly, M.; Akoua-Koffi, C.; Buchwald, N.; Schubert, G.; Weiss, S.; Couacy-Hymann, E.; Anoh, A.E.; Mossoun, A.; Calvignac-Spencer, S.; Leendertz, S.A.; et al. Adenovirus in Rural Côte D‘Ivoire: High Diversity and Cross-Species Detection. EcoHealth 2015, 12, 441–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Counihan, K.L.; Skerratt, L.F.; Franson, J.C.; Hollmén, T.E. Phylogenetic and Pathogenic Characterization of Novel Adenoviruses Isolated from Long-Tailed Ducks (Clangula hyemalis). Virology 2015, 485, 393–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohizumi, T.; Nakamura, K.; Yamamoto, Y.; Mase, M.; Yamada, M. Detection of Fowl Adenovirus DNA from Formalin-Fixed and Paraffin-Embedded Sections by PCR and Classification of Serotypes by Sequencing of PCR Products. Avian Dis. 2012, 56, 741–743. [Google Scholar] [CrossRef] [PubMed]
- Marek, A.; Schulz, E.; Hess, C.; Hes, M. Comparison of the Fibers of Fowl Adenovirus a Serotype 1 Isolates from Chickens with Gizzard Erosions in Europe and Apathogenic Reference Strains. J. Vet. Diagn. Investig. 2010, 22, 937–941. [Google Scholar] [CrossRef] [Green Version]
- Pallister, J.; Wright, P.; Sheppard, M. A Single Gene Encoding the Fiber is Responsible for Variations in Virulence in the Fowl Adenoviruses. J. Virol. 1996, 70, 5115–5122. [Google Scholar] [CrossRef] [Green Version]
- Mase, M.; Nakamura, K.; Imada, T. Characterization of Fowl Adenovirus Serotype 4 Isolated from Chickens with Hydropericardium Syndrome Based on Analysis of the Short Fiber Protein Gene. J. Vet. Diagn. Investig. 2010, 22, 218–223. [Google Scholar] [CrossRef] [Green Version]
- Zhao, J.; Zhong, Q.; Zhao, Y.; Hu, Y.-X.; Zhang, G.-Z. Pathogenicity and Complete Genome Characterization of Fowl Adenoviruses Isolated from Chickens Associated with Inclusion Body Hepatitis and Hydropericardium Syndrome in China. PLoS ONE 2015, 10, e0133073. [Google Scholar] [CrossRef]
- Slaine, P.D.; Ackford, J.G.; Kropinski, A.M.; Kozak, R.A.; Krell, P.J.; Nagy, E. Molecular Characterization of Pathogenic and Nonpathogenic Fowl Aviadenovirus Serotype 11 Isolates. Can. J. Microbiol. 2016, 62, 993–1002. [Google Scholar] [CrossRef]
- Kang, S.; Kim, Y.-C. Identification of Viral Taxon-Specific Genes (VTSG): Application toCaliciviridae. Genom. Inf. 2018, 16, e23. [Google Scholar] [CrossRef] [Green Version]
- Stano, M.; Beke, G.; Klucar, L. viruSITE—Integrated Database for Viral Genomics. Database 2016. [Google Scholar] [CrossRef]
- Goodacre, N.; Aljanahi, A.; Nandakumar, S.; Mikailov, M.; Khan, A.S. A Reference Viral Database (RVDB) To Enhance Bioinformatics Analysis of High-Throughput Sequencing for Novel Virus Detection. mSphere 2018, 3, e00069-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cochrane, G.; Karsch-Mizrachi, I.; Takagi, T. International Nucleotide Sequence Database Collaboration. The International Nucleotide Sequence Database Collaboration. Nucleic Acids Res. 2016, 44, D48–D50. [Google Scholar] [CrossRef] [PubMed]
- Consortium, U. UniProt: A hub for protein information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Guertin, P.; Jia, G.; Lv, Z.; Yang, H.; Ju, D. Large-Scale Microcarrier Culture of HEK293T Cells and Vero Cells in Single-Use Bioreactors. AMB Express 2019, 9, 70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grein, T.A.; Weidner, T.; Czermak, P. Concepts for the Production of Viruses and Viral Vectors in Cell Culture. In New Insights into Cell Culture Technology; Gowder, S.J.T., Ed.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Isolate | Microcarrier Concentration (/mL) | Surface Area | Virus Innoculum Titre (/mL) | Inoculation Volume (mL) | CEL Cells Inoculated (/mL) |
|---|---|---|---|---|---|---|
| 1 | UPM08136 P5 | 8.6 × 106 | 17.6 m2 | 107.67 | 10 | 3.1 × 107 |
| 2 | UPM08136 P20 | 8.6 × 106 | 17.6 m2 | 105.5 | 10 | 3.1 × 107 |
| No | Isolate Name | Serotype | Origin | Accession Number | Uploaded By |
|---|---|---|---|---|---|
| 1 | UPM04217 | 8b | Malaysia | ANA50319.1 | Juliana et al., 2016 |
| 2 | UPM08158 | 8b | Malaysia | AEL21619.1 | Juliana et al., 2011 |
| 3 | UPM1137CEL20 | 8b | Malaysia | AQZ26936.1 | Norfitriah et al., 2017 |
| 4 | UPM08158 | 8b | Malaysia | AEL21619.1 | Juliana et al., 2011 |
| 5 | Celo | 1 | Australia | EU979367.1 | Steer et al., 2009 |
| 6 | SR48 | 2 | Australia | EU979368.1 | Steer et al., 2009 |
| 7 | SR49 | 3 | Australia | EU070369.1 | Steer et al., 2009 |
| 8 | KR5 | 4 | Australia | EU979370.1 | Steer et al., 2009 |
| 9 | 340 | 5 | Australia | YP 007985654.1 | Marek et al., 2013 |
| 10 | CR119 | 6 | Australia | YP 009505663.1 | Steer et al., 2009 |
| 11 | YR36 | 7 | Australia | ACL68141.1 | Steer et al., 2009 |
| 12 | AO2 | 9 | Australia | EU979376.1 | Steer et al., 2009 |
| 13 | C2B | 10 | Australia | EU979377.1 | Steer et al., 2009 |
| 14 | Wroclaw 2015 | 1 | Poland | KR259656.1 | Wieliczko et al., 2015 |
| 15 | USP-EC-01 | 6 | Ecuador | ASU91620.1 | De la Torre et al., 2017 |
| 16 | TR59 | 8b | Australia | ACL68142.1 | Steer et al., 2008 |
| 17 | 40440-M/2015 | 5 | Hungary | QCC26479.1 | Kajan et al., 2018 |
| 18 | LYP | 5 | China | QJP03679.1 | Chen, 2019 |
| 19 | CH/CQBS/1601 | 4 | China | MF055642 | Xia et al., 2017 |
| 20 | CH/CBQS/1512 | 8b | China | AUD09312.1 | Xia et al., 2017 |
| 21 | 05-51425 | 8b | Canada | ABS81057.1 | Ojkic et al., 2007 |
| 22 | Indian | 4 | India | AJ459805 | Barua et al., 2002 |
| 23 | 10-3678 | 8b | Poland | CUT98160 | Schachner et al., 2016 |
| 24 | HG | 8b | Canada | YP 004191821.1 | Grgic et al., 2011 |
| 25 | 14-259 | 8b | France | QGQ62947.1 | Schachner et al., 2019 |
| 26 | HUNGG | 8b | Hungary | QGQ62522.1 | Schachner et al., 2019 |
| 27 | HLJ/151129 | 8b | China | AOS87877.1 | Wang et al., 2016 |
| 28 | FAdV-HNQX-101017-B | 8b | China | ANG57906.1 | Li et al., 2016 |
| 29 | TR/8VKE/R/B-8 | 8b | Turkey | QGP73412.1 | Sahindokuyucu et al., 2019 |
| 30 | 04-53357-105 | 8B | Canada | ABS81134.1 | Ojkic et al., 2007 |
| 31 | SA38D-08 | 8b | South Africa | ADV90772.1 | Joubert et al., 2010 |
| 32 | SA38C-08 | 8b | South Africa | ADV90771.1 | Joubert et al., 2010 |
| 33 | USP-BR-420.12 | 8b | Brazil | AQX17387.1 | De la Torre et al., 2016 |
| 34 | 764 | 8b | United Kingdom | ANJ02566.1 | Marek et al., 2016 |
| 35 | 764 | 8b | Canada | AER40292.1 | Dar et al., 2012 |
| 36 | ID-HCI-037 | 8b | Peru | AWF93664.1 | Caballero-Garcia et al., 2018 |
| 37 | Turkey Adv | 3 | United Kingdom | AC_000016 | Davison et al., 2003 |
| No | Isolate Name | Serotype | Origin | Accession Number | Uploaded By |
|---|---|---|---|---|---|
| 1 | UPM04217 | 8b | Malaysia | KU517714.1 | Juliana et al., 2015 |
| 2 | UPM1137E2 | 8b | Malaysia | KY305950.1 | Norfitriah et al., 2017 |
| 3 | PL-060-08 | 1 | Poland | GU952109.1 | Domanska-Blicharz et al., 2011 |
| 4 | YR36 | 7 | Austria | KT862809.1 | Marek et al., 2015 |
| 5 | CR119 | 6 | USA | NC_038332.1 | Schachner et al., 2018 |
| 6 | NARC-3317 | 4 | Pakistan | KT733569.1 | Jabeen et al., 2015 |
| 7 | SX17 | 4 | China | MF595799.1 | Feng et al., 2017 |
| 8 | Punjab1 | 1 | India | DQ864435.1 | Bhan et al., 2006 |
| 9 | 06-25854-1 | 11 | Canada | JQ034219.1 | Grgic et al., 2014 |
| 10 | WA-1/100966-2 | 11 | Australia | KT037713.1 | Steer et al., 2015 |
| 11 | 340 | 5 | Austria | FR872928.1 | Marek et al., 2015 |
| 12 | HLJ151129 | 8b | China | AOS87884.1 | Wang et al., 2016 |
| 13 | 764 | 8b | United Kingdom | ANJ02574.1 | Marek et al., 2015 |
| 14 | 06-41265-07 | 8b | Canada | AFD32283.1 | Grgic et al., 2014 |
| 15 | QD2016 | 8b | China | AWT08538.1 | Hu et al.,2017 |
| 16 | NZ-1/101151-1 | 8b | New Zealand | ANQ43486.1 | Steer et al., 2015 |
| 17 | Vac-2005 | 8b | Australia | ANQ43481.1 | Steer et al., 2015 |
| 18 | VIC-2/430-6 | 8b | Australia | ANQ43482.1 | Steer et al., 2015 |
| 19 | NSW-3/100931 | 8b | Australia | ANQ43483.1 | Steer et al., 2015 |
| 20 | FJ-1/100842-C | 8b | Fiji | ANQ43487.1 | Steer et al., 2015 |
| 21 | VIC-8/100719 | 8b | Australia | ANQ43484.1 | Steer et al., 2015 |
| 22 | NSW-5/100931 | 8b | Australia | ANQ43485.1 | Steer et al., 2015 |
| 23 | Strain 13-19395 | - | Germany | MK572863.1 | Schachner et al., 2019 |
| 24 | LYG | 5 | China | QJP03687.1 | Chen, 2019 |
| 25 | 340 | 5 | Northern Ireland | YP 007985662.1 | Marek et al.,2013 |
| 26 | 127 | DAdV 1 | Russia | Z86065.1 | Akopian et al., 1997 |
| 27 | D11-JW-010 | DAdV 1 | South Korea | JX227930 | Cha et al., 2012 |
| No | FAdV Isolate | Initial Titre (/mL) | Final Titre (/mL) | Final Volume (/mL) |
|---|---|---|---|---|
| 1 | UPM08136CELP5B1 | 105.5 | 107.5 | 500 |
| 2 | UPM08136CELP20B1 | 107.67 | 106.5 | 600 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ugwu, C.C.; Hair-Bejo, M.; Nurulfiza, M.I.; Omar, A.R.; Ideris, A. Propagation and Molecular Characterization of Fowl Adenovirus Serotype 8b Isolates in Chicken Embryo Liver Cells Adapted on Cytodex™ 1 Microcarrier Using Stirred Tank Bioreactor. Processes 2020, 8, 1065. https://doi.org/10.3390/pr8091065
Ugwu CC, Hair-Bejo M, Nurulfiza MI, Omar AR, Ideris A. Propagation and Molecular Characterization of Fowl Adenovirus Serotype 8b Isolates in Chicken Embryo Liver Cells Adapted on Cytodex™ 1 Microcarrier Using Stirred Tank Bioreactor. Processes. 2020; 8(9):1065. https://doi.org/10.3390/pr8091065
Chicago/Turabian StyleUgwu, Chidozie C., Mohd Hair-Bejo, Mat I. Nurulfiza, Abdul R. Omar, and Aini Ideris. 2020. "Propagation and Molecular Characterization of Fowl Adenovirus Serotype 8b Isolates in Chicken Embryo Liver Cells Adapted on Cytodex™ 1 Microcarrier Using Stirred Tank Bioreactor" Processes 8, no. 9: 1065. https://doi.org/10.3390/pr8091065
APA StyleUgwu, C. C., Hair-Bejo, M., Nurulfiza, M. I., Omar, A. R., & Ideris, A. (2020). Propagation and Molecular Characterization of Fowl Adenovirus Serotype 8b Isolates in Chicken Embryo Liver Cells Adapted on Cytodex™ 1 Microcarrier Using Stirred Tank Bioreactor. Processes, 8(9), 1065. https://doi.org/10.3390/pr8091065