Nitrogen Removal from Agricultural Subsurface Drainage by Surface-Flow Wetlands: Variability


Abstract
:1. Introduction
1.1. Agriculture as Nitrogen Sources
1.2. Need to Recover Ecosystem Services at the Edge-of-Field
1.3. Aim of the Review
2. Variability in the Performance of Surface-Flow Constructed Wetlands
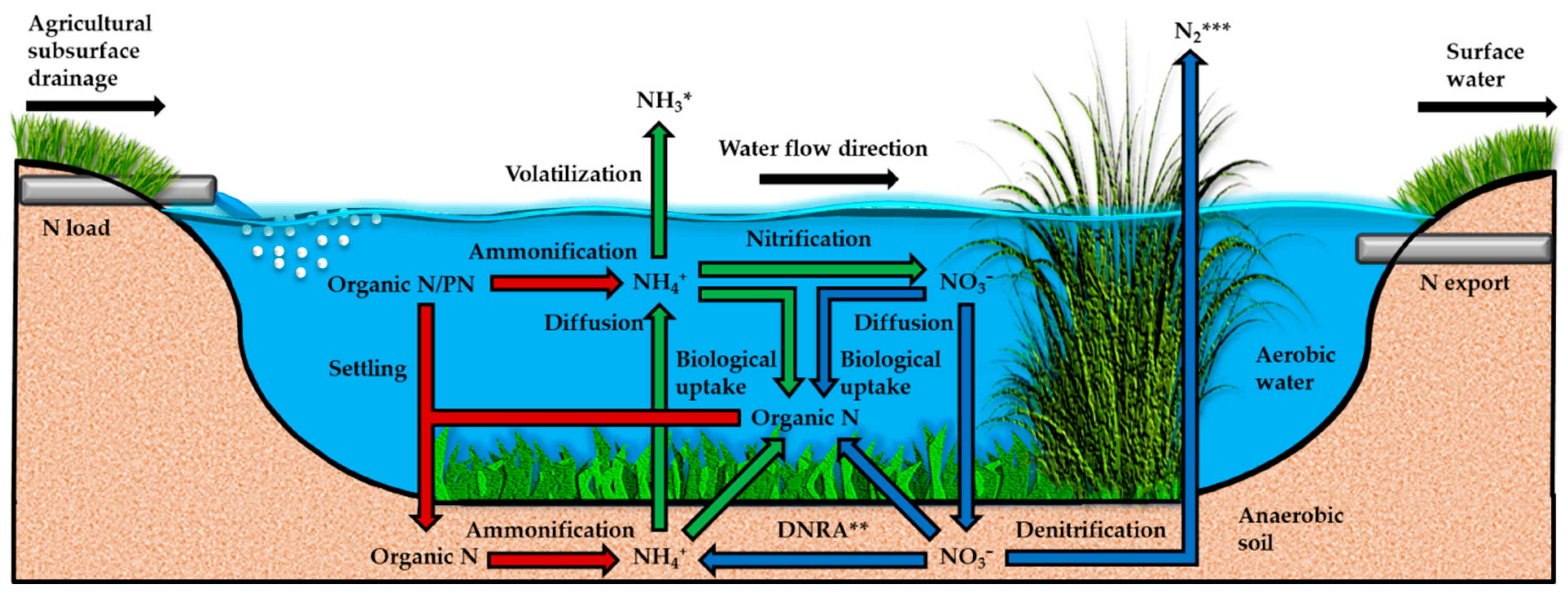
2.1. Nitrogen Load and Forms
2.1.1. Nitrogen Removal Rate

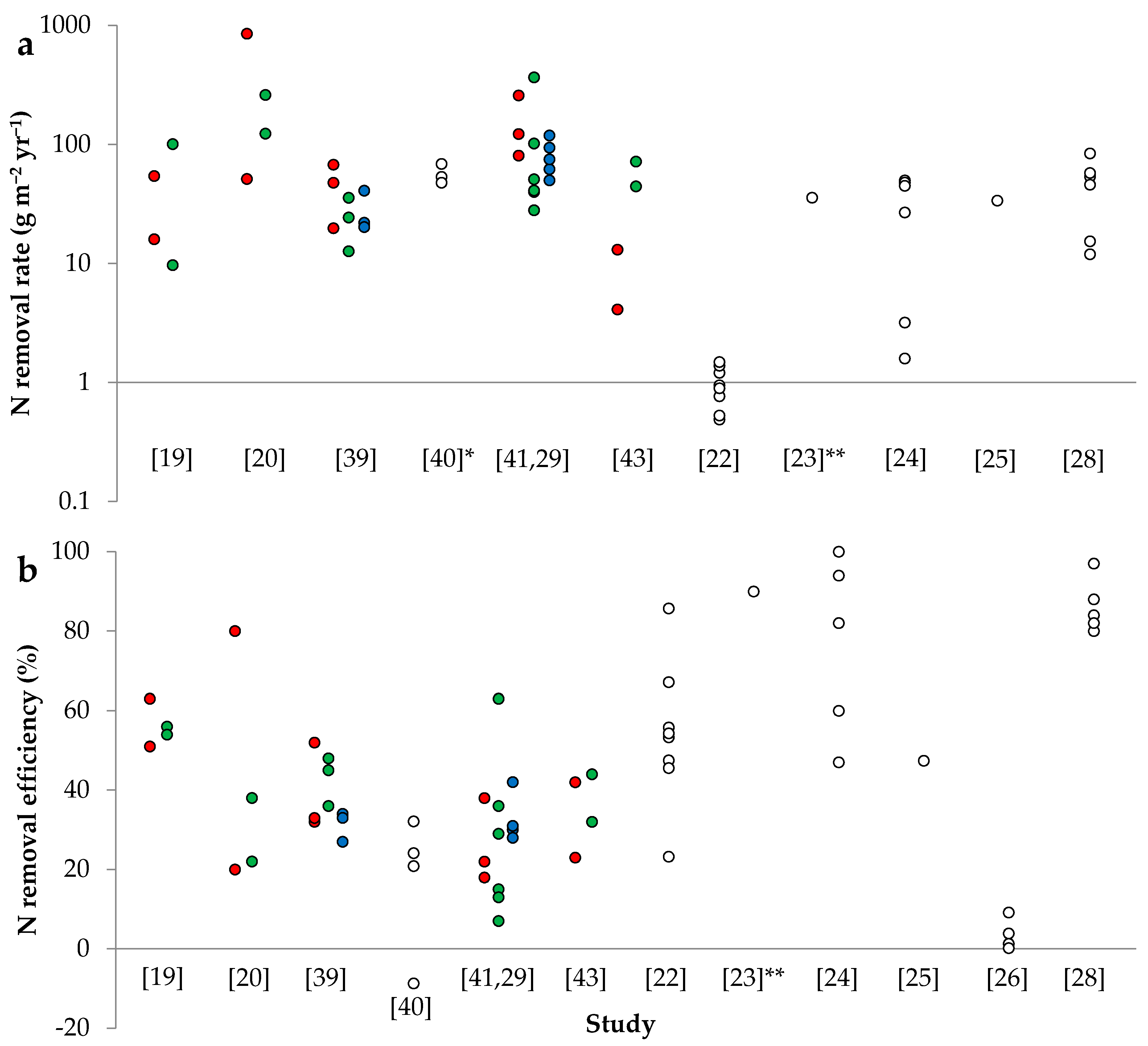{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Country | Name | SFW Area | Ratio SFW:AC Area | Monitoring Time | Hydraulic Load | N Concentration | N Load | NO3−/NH4+/Org. N Fraction | Hydraulic Residence Time | N Removal | Study | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| m2 | % | yr | m yr−1 | mg L−1 | g m−2 yr−1 | % | d | g m−2 yr−1 | % | |||
| USA | Wetland A | 6000 | 4 | 2 | 5.8 | 11.4 | 66 | >96/-/- | 56.8 | 35 | 53 | [19] |
| USA | Wetland B | 3000 | 3 | 2 | 6.4 | 15.7 | 101 | >96/-/- | 22.7 | 55 | 55 | [19] |
| New Zealand | Waikato | 260 | 1 | 2 | 25.1 | 12.7 a | 657 | 45/0/55 | 4.4 b | 452 | 69 | [20,42] |
| New Zealand | Northland | 898 | 1.6 | 2 | 47.4 | 11.1 a | 623 | 76/5/19 | - | 192 | 31 | [20] |
| USA | - | 1012 | 1 | 3 | 8.4 | - | - | - | - | - | 68 c | [31] |
| USA | Wetland A | 6000 | 4 | 3 | 8.0 | 14.0 d | 336 | 93/7/0 | 41.1 | 136 | 40 | [39] |
| USA | Wetland B | 3000 | 6 | 3 | 5.3 | 10.4 d | 166 | 99/1/0 | 27.5 | 73 | 44 | [39] |
| USA | Wetland D | 8000 | 3.2 | 3 | 6.3 | 8.7 d | 266 | 98/2/0 | 37.9 | 83 | 31 | [39] |
| Canada | Walbridge | 1215 | 0.004 e | 4 | 58.3 | 3.4 c | 263 | 100/0/0 | - | 49 | 19 | [40] |
| New Zealand | Titoki | 898 | 1.6 | 3 | 53.1 | 10.7 | 564 | 76/19/5 | - | 154 | 27 | [41] |
| New Zealand | Toenepi | 293 | 1.1 | 5 | 25.3 | 13.4 | 342 | 83/1/16 | - | 117 | 34 | [41] |
| New Zealand | Bog Burn | 112.5 | 0.66 | 4 | 40.9 | 5.8 | 224 | 73/2/25 | - | 70 | 31 | [41] |
| USA | Wetland 1 | 1600 | 0.07 | 2 | 4.0 | 9.8 d | 39 | 90/0/10 | 38.1 | 12 | 30 | [43] |
| USA | Wetland 2 | 4000 | 0.03 | 2 | 6.5 | 13.2 d | 96 | 96/0/4 | 24.8 | 37 | 38 | [43] |
| Switzerland | Boden | 720 | 0.86 | 2.5 | 30.4 | 5.5 | 167 | 47.5/5/47.5 | 11.7 | 45 | 27 | [45] |
| France | - | 4165 | 1.2 | 8 | 0.1 | 14.1 c | 2 c | - | 7240.1 | 1 c | 50 c | [22] |
| Italy | - | 3200 | 5.3 | 5 | 5.7 | 7.0 | 40 | 87/-/- | - | 36 | 90 | [23] |
| Italy | - | 3750 | 3 | 6 | 2.5 | 13.5 | 42 | - | 58.8 | 29 | 69 | [24] |
| USA | Wetland B | 3000 | 6 | 1 | 8.8 | 8.2 | 72 | 100/0/0 | 19.4 | 34 | 47 | [25] |
| Germany | - | 4632 | 0.4 | 4 | 20.1 | 9.0 a | 181 | 89/0/11 | - | - | 3 | [26] |
| Netherlands | - | 64 | 0.26 | 2 | - | 30.0 | - | 96/-/- | - | 166 | 58 | [27] |
| Italy | - | 3200 | 7.1 | 6 | 7.8 | 5.9 a | 54 | 74/-/- | - | 45 | 84 | [28] |
| New Zealand | Toenepi | 293 | 1.1 | 1 | 30.4 | 10.4 | 316 | 96/1/3 | - | 41 | 13 | [29] |
| New Zealand | Bog Burn | 112.5 | 0.66 | 1 | 38.2 | 7.4 | 284 | 78/4/18 | - | 119 | 42 | [29] |
| Sweden | Bölarp | 2800 | 0.14 | 2 | - | - | 2338 | - | - | 100 | 4 | [30] |
| Sweden | Edenberga | 2200 | 0.37 | 2 | - | - | 625 | - | - | 58 | 9 | [30] |
| Sweden | Södra Stene | 21,000 | 2.1 | 2 | - | - | 14 | - | - | 2 | 12 | [30] |
| Name | NO3− | NH4+ | Organic N | Hydraulic Residence Time | Study | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Concentration | Load | Removal | Concentration | Load | Removal | Concentration | Load | Removal | ||||||
| mg L−1 | g m−2 yr−1 | % | mg L−1 | g m−2 yr−1 | % | mg L−1 | g m−2 yr−1 | % | d | |||||
| Wetland A | 11.0 | 64 | 36 | 56 | - | - | - | - | - | - | - | - | 56.8 | [19] |
| Wetland B | 15.4 | 99 | 55 | 56 | - | - | - | - | - | - | - | - | 22.7 | [19] |
| Waikato | 10.9 a | 295 | 97 | 33 | 0.0 a | 2 | −2 | −90 | 0.7 a | 360 | 357 | 99 | 4.4 b | [20,42] |
| Northland | 8.4 a | 474 | 277 | 58 | 0.2 a | 30 | −100 | −334 | 1.6 a | 119 | 15 | 13 | - | [20] |
| - | - | - | - | 68 | - | - | - | - | - | - | - | - | - | [31] |
| Wetland A | 13 c | 104 | 42 | 41 | 1 | 8 | 4 | 54 | 0 | 0 | -1 | - | 41.1 | [39] |
| Wetland B | 10.3 c | 55 | 25 | 45 | 0.1 | 1 | 0 | 45 | 0 | 0 | -1 | - | 27.5 | [39] |
| Wetland D | 8.5 c | 87 | 30 | 34 | 0.2 | 2 | 1 | 42 | 0 | 0 | -3 | - | 37.9 | [39] |
| Walbridge | 3.4 | 263 | 49 | 19 | 0.0 | 0.41 | 0.11 | 27 | - | - | - | - | - | [40] |
| Titoki | 8.1 | 429 | 239 | 56 | 2.0 | 107 | −8 | −7 | 0.6 | 29 | −75 | −263 | - | [41] |
| Toenepi | 11.1 | 282 | 84 | 30 | 0.1 | 3 | −8 | −267 | 2.2 | 58 | 43 | 74 | - | [41] |
| Bog Burn | 4.2 | 164 | 78 | 47 | 0.1 | 5 | −2 | −50 | 1.4 | 54 | −6 | −10 | - | [41] |
| Wetland 1 | 8.6 | 34 | 9 | 25 | 0.1 | 0 | 0 | 0 | 1.1 | 4 | 3 | 73 | 38.1 | [43] |
| Wetland 2 | 12.4 | 92 | 35 | 37 | 0.1 | 0 | 0 | −42 | 0.8 | 3 | 2 | 50 | 24.8 | [43] |
| Boden | 2.6 | 79 | 23 | 29 | 0.3 | 8 | −1 | −7 | 2.6 | 79 | 22 | 28 | 11.7 | [45] |
| - | 14.1 | 2 | 1 | 50 | 0.1 | - | - | - | - | - | - | - | 7240.1 | [22] |
| - | 1.4 a | 35 | - | - | 0.0 a | - | - | - | 1.0 a | - | - | 80 | - | [23] |
| - | 12.8 | 48 | 32 | 67 | 0.1 | 0.3 | 0.1 | 38 | 2.2 | 9 | 5 | 54 | 58.8 | [24] |
| Wetland B | 8.1 | 71 | 35 | 50 | 0.0 | 0.1 | −0.2 | −125 | 0.0 | 0 | −1 | - | 19.4 | [25] |
| - | 8.0 a | 161 | - | - | 0.0 a | 0.2 | - | - | 1.0 a | 20 | - | - | - | [26] |
| - | 4.3 a | 40 | 33 | 84 | - | - | - | - | - | - | - | - | - | [28] |
| Toenepi | 10.0 | 253 | 24 | 9 | 0.1 | 1 | −1 | −100 | 0.3 | 3 | −3 | −100 | - | [29] |
| Bog Burn | 5.8 | 221 | 81 | 37 | 0.3 | 10 | 7 | 70 | 1.4 | 53 | 31 | 58 | - | [29] |
| Wetland A | 11.2 c | 106 | 58 | 55 | - | - | - | - | - | - | - | - | 35.8 | [35] |
| Wetland D | 7.1 c | 74 | 24 | 33 | - | - | - | - | - | - | - | - | 23.3 | [35] |
2.1.2. Nitrogen Removal Efficiency
2.2. Effect of Hydrology
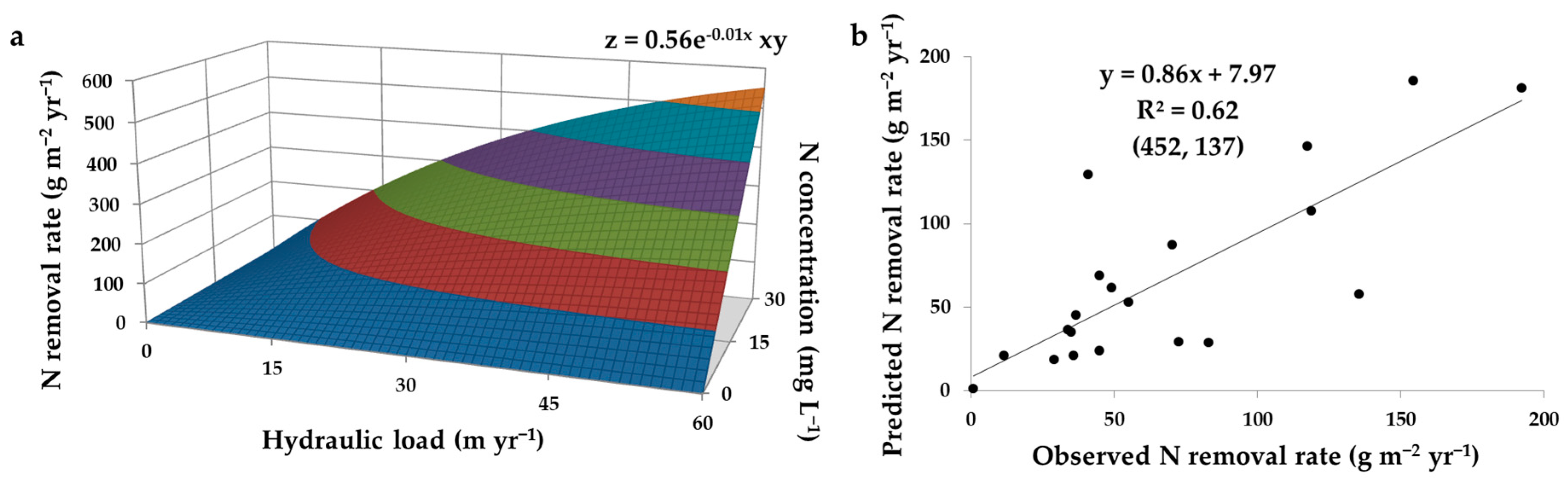
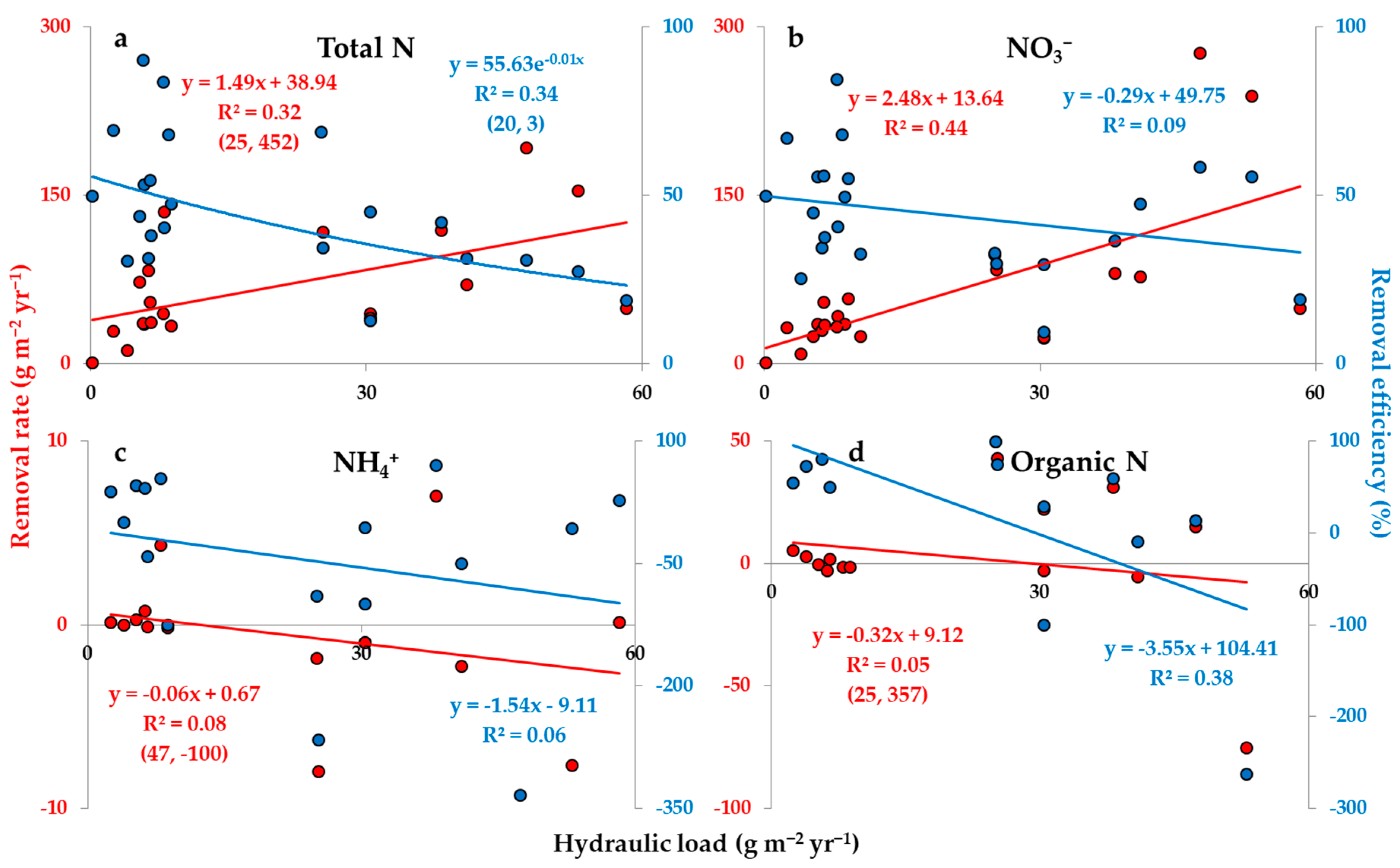
2.2.1. Hydraulic Load
2.2.2. Hydraulic Residence Time
2.2.3. Seasonality
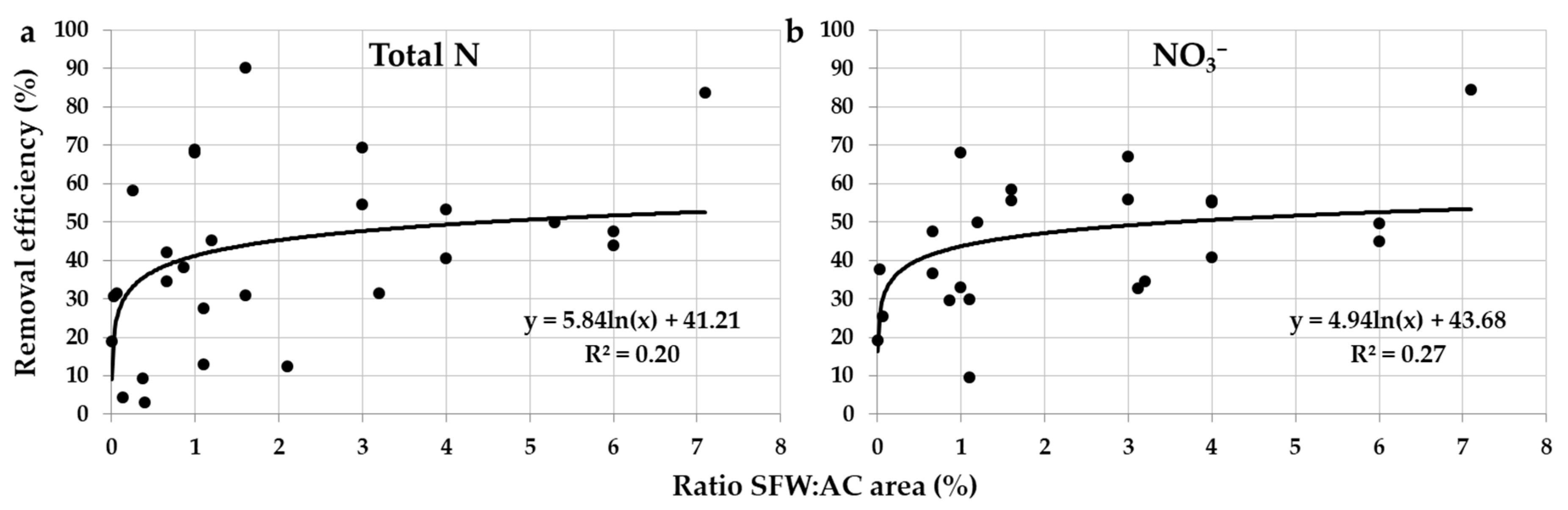
2.2.4. Area Ratio of Surface-Flow Constructed Wetland for the Agricultural Catchment
2.2.5. Water Flow Dynamics
2.2.6. Design Aspects
2.2.7. Final Remarks
2.3. Nitrogen Removal Mechanisms and Biogeochemical Factors
2.3.1. Overview
2.3.2. Denitrification
2.3.3. Biological Uptake
2.3.4. Settling
2.3.5. Nitrogen Transformation Processes
2.3.6. Seasonality
2.3.7. Final Remarks
3. Summary, Conclusions and Final Remarks
3.1. Nitrogen Inputs and Incoming Loads
3.2. Design
3.3. Removal Processes and Factors
3.4. Climate
Funding
Conflicts of Interest
References
- Randall, G.W.; Mulla, D.J. Nitrate nitrogen in surface waters as influenced by climatic conditions and agricultural practices. J. Environ. Qual. 2001, 30, 337–344. [Google Scholar] [CrossRef] [Green Version]
- Sun, B.; Zhang, L.; Yang, L.; Zhang, F.; Norse, D.; Zhu, Z. Agricultural non-point source pollution in China: Causes and mitigation measures. Ambio 2012, 41, 370–379. [Google Scholar] [CrossRef] [Green Version]
- Guo, W.; Fu, Y.; Ruan, B.; Ge, H.; Zhao, N. Agricultural non-point source pollution in the Yongding River Basin. Ecol. Indic. 2014, 36, 254–261. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Day, J.W.; Gilliam, J.W.; Groffman, P.M.; Hey, D.L.; Randall, G.W.; Wang, N. Reducing Nutrient Loads, Especially Nitrate-Nitrogen, to Surface Water, Ground Water, and the Gulf of Mexico: Topic 5 Report for the Integrated Assessment on Hypoxia in the Gulf of Mexico; Decision Analysis Series no. 19; NOAA Coastal Ocean Office: Silver Spring, MD, USA, 1999. [Google Scholar]
- Baker, J.L. Limitations of improved nitrogen management to reduce nitrate leaching and increase use efficiency. In Optimizing Nitrogen Management in Food and Energy Production and Environmental Protection: Proceedings of the 2nd International Nitrogen Conference on Science and Policy; TheScientificWorld, Hindawi: London, UK, 2001; pp. 10–16. [Google Scholar]
- Dinnes, D.L.; Karlen, D.L.; Jaynes, D.B.; Kaspar, T.C.; Hatfield, J.L.; Colvin, T.S.; Cambardella, C.A. Nitrogen management strategies to reduce nitrate leaching in tile-drained midwestern soils. Agron J. 2002, 94, 153–171. [Google Scholar] [CrossRef]
- Baker, J.L.; Melvin, S.W.; Lemke, D.W.; Lawlor, P.A.; Crumpton, W.G.; Helmers, M.J. Subsurface drainage in Iowa and the water quality benefits and problem. In Drainage VIII the Proceedings of the Eighth International Drainage Symposium; Cooke, R., Ed.; American Society of Agricultural and Biological Engineers: Sacramento, CA, USA, 2004; pp. 39–50. [Google Scholar]
- McLellan, E.; Robertson, D.; Schilling, K.; Tomer, M.; Kostel, J.; Smith, D.; King, K. Reducing nitrogen export from the corn belt to the gulf of Mexico: Agricultural strategies for remediating hypoxia. JAWRA J. Am. Water Resour. Assoc. 2014, 51, 263–289. [Google Scholar] [CrossRef]
- Mitsch, W.J.; Day, J.W.; Gilliam, J.W.; Groffman, P.M.; Hey, D.L.; Randall, G.W.; Wang, N. Reducing nitrogen loading to the gulf of Mexico from the Mississippi River Basin: Strategies to counter a persistent ecological problem. Bioscience 2001, 51, 373–388. [Google Scholar] [CrossRef]
- Strock, J.S.; Kleinman, P.J.A.; King, K.W.; Delgado, J.A. Drainage water management for water quality protection. J. Soil Water Conserv. 2010, 65, 131A–136A. [Google Scholar] [CrossRef] [Green Version]
- Valero, C.S.; Madramootoo, C.A.; Stämpfli, N. Water table management impacts on phosphorus loads in tile drainage. Agric. Water Manag. 2007, 89, 71–80. [Google Scholar] [CrossRef]
- Pierce, S.C.; Kröger, R.; Pezeshki, R. Managing artificially drained low-gradient agricultural headwaters for enhanced ecosystem functions. Biology 2012, 1, 794–856. [Google Scholar] [CrossRef]
- Frey, S.K.; Hwang, H.-T.; Park, Y.-J.; Hussain, S.I.; Gottschall, N.; Edwards, M.; Lapen, D.R. Dual permeability modeling of tile drain management influences on hydrologic and nutrient transport characteristics in macroporous soil. J. Hydrol. 2016, 535, 392–406. [Google Scholar] [CrossRef]
- Hussain, S.I.; Frey, S.K.; Blowes, D.W.; Ptacek, C.J.; Wilson, D.; Mayer, K.U.; Su, D.; Gottschall, N.; Edwards, M.; Lapen, D.R. Reactive Transport of Manure-Derived Nitrogen in the Vadose Zone: Consideration of Macropore Connectivity to Subsurface Receptors. Vadose Zone J. 2019, 18, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Stålnacke, P.; Aakerøy, P.A.; Blicher-Mathiesen, G.; Iital, A.; Jansons, V.; Koskiaho, J.; Kyllmar, K.; Lagzdins, A.; Pengerud, A.; Povilaitis, A. Temporal trends in nitrogen concentrations and losses from agricultural catchments in the Nordic and Baltic countries. Agric. Ecosyst. Environ. 2014, 198, 94–103. [Google Scholar] [CrossRef]
- Mendes, L.R.D. Edge-of-Field technologies for phosphorus retention from agricultural drainage discharge. Appl. Sci. 2020, 10, 634. [Google Scholar] [CrossRef] [Green Version]
- Weisner, S.E.B.; Johannesson, K.; Thiere, G.; Svengren, H.; Ehde, P.M.; Tonderski, K.S. National large-scale wetland creation in agricultural areas—Potential versus realized effects on nutrient transports. Water 2016, 8, 544. [Google Scholar] [CrossRef]
- Hoffmann, C.C.; Zak, D.; Kronvang, B.; Kjaergaard, C.; Carstensen, M.V.; Audet, J. An overview of nutrient transport mitigation measures for improvement of water quality in Denmark. Ecol. Eng. 2020, 155, 105863. [Google Scholar] [CrossRef]
- Groh, T.A.; Gentry, L.E.; David, M.B. Nitrogen removal and greenhouse gas emissions from constructed wetlands receiving tile drainage water. J. Environ. Qual. 2015, 44, 1001–1010. [Google Scholar] [CrossRef] [Green Version]
- Tanner, C.C.; Nguyen, M.; Sukias, J. Constructed wetland attenuation of nitrogen exported in subsurface drainage from irrigated and rain-fed dairy pastures. Water Sci. Technol. 2005, 51, 55–61. [Google Scholar] [CrossRef]
- Kang, H.; Freeman, C.; Lee, D.; Mitsch, W.J. Enzyme activities in constructed wetlands: Implication for water quality amelioration. Hydrobiologia 1998, 368, 231–235. [Google Scholar] [CrossRef]
- Tournebize, J.; Chaumont, C.; Fesneau, C.; Guenne, A.; Vincent, B.; Garnier, J.; Mander, Ü. Long-term nitrate removal in a buffering pond-reservoir system receiving water from an agricultural drained catchment. Ecol. Eng. 2015, 80, 32–45. [Google Scholar] [CrossRef]
- Borin, M.; Tocchetto, D. Five year water and nitrogen balance for a constructed surface flow wetland treating agricultural drainage waters. Sci. Total. Environ. 2007, 380, 38–47. [Google Scholar] [CrossRef]
- Lavrnić, S.; Braschi, I.; Anconelli, S.; Blasioli, S.; Solimando, D.; Mannini, P.; Toscano, A. Long-term monitoring of a surface flow constructed wetland treating agricultural drainage water in northern Italy. Water 2018, 10, 644. [Google Scholar] [CrossRef] [Green Version]
- Hoagland, C.R.; Gentry, L.E.; David, M.B.; Kovacic, D.A. Plant nutrient uptake and biomass accumulation in a constructed wetland. J. Freshw. Ecol. 2001, 16, 527–540. [Google Scholar] [CrossRef]
- Steidl, J.; Kalettka, T.; Bauwe, A. Nitrogen retention efficiency of a surface-flow constructed wetland receiving tile drainage water: A case study from north-eastern Germany. Agric. Ecosyst. Environ. 2019, 283. [Google Scholar] [CrossRef]
- de Haan, J.; van der Schoot, J.R.; Verstegen, H.; Clevering, O. Removal of nitrogen leaching from vegetable crops in constructed wetlands. Acta Hortic. 2010, 852, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Tolomio, M.; Ferro, N.D.; Borin, M. Multi-Year N and P removal of a 10-year-old surface flow constructed wetland treating agricultural drainage waters. Agronomy 2019, 9, 170. [Google Scholar] [CrossRef] [Green Version]
- Sukias, J.; Tanner, C.C. Surface flow constructed wetlands as a drainage management tool–long term performance. In Adding to the Knowledge Base for the Nutrient Manager; Currie, L.D., Christensen, C.L., Eds.; Fertilizer & Lime Research Centre, Occasional Report No 24. Massey University: Palmerston North, New Zealand, 2011; pp. 1–16. [Google Scholar]
- Strand, J.A.; Weisner, S.E.B. Effects of wetland construction on nitrogen transport and species richness in the agricultural landscape—Experiences from Sweden. Ecol. Eng. 2013, 56, 14–25. [Google Scholar] [CrossRef]
- Lenhart, C.; Gordon, B.; Gamble, J.; Current, D.; Ross, N.; Herring, L.; Nieber, J.; Peterson, H. Design and hydrologic performance of a tile drainage treatment wetland in Minnesota, USA. Water 2016, 8, 549. [Google Scholar] [CrossRef] [Green Version]
- Tournebize, J.; Chaumont, C.; Mander, Ü. Implications for constructed wetlands to mitigate nitrate and pesticide pollution in agricultural drained watersheds. Ecol. Eng. 2017, 103, 415–425. [Google Scholar] [CrossRef]
- Vymazal, J. The use of constructed wetlands for nitrogen removal from agricultural drainage: A review. Sci. Agric. Bohem. 2017, 48, 82–91. [Google Scholar] [CrossRef] [Green Version]
- Maxwell, E.; Peterson, E.W.; O’Reilly, C.M. Enhanced nitrate reduction within a constructed wetland system: Nitrate removal within groundwater flow. Wetlands 2017, 37, 413–422. [Google Scholar] [CrossRef]
- Larson, A.C.; Gentry, L.E.; David, M.B.; Cooke, R.A.; Kovacic, D.A. The role of seepage in constructed wetlands receiving agricultural tile drainage. Ecol. Eng. 2000, 15, 91–104. [Google Scholar] [CrossRef]
- Carstensen, M.V.; Hashemi, F.; Hoffmann, C.C.; Zak, D.; Audet, J.; Kronvang, B. Efficiency of mitigation measures targeting nutrient losses from agricultural drainage systems: A review. Ambio 2020, 49, 1820–1837. [Google Scholar] [CrossRef]
- Drake, C.; Jones, C.; Schilling, K.; Amado, A.A.; Weber, L. Estimating nitrate-nitrogen retention in a large constructed wetland using high-frequency, continuous monitoring and hydrologic modeling. Ecol. Eng. 2018, 117, 69–83. [Google Scholar] [CrossRef]
- Ulén, B.; Geranmayeh, P.; Blomberg, M.; Bieroza, M. Seasonal variation in nutrient retention in a free water surface constructed wetland monitored with flow-proportional sampling and optical sensors. Ecol. Eng. 2019, 139, 105588. [Google Scholar] [CrossRef]
- Kovacic, D.A.; David, M.B.; Gentry, L.E.; Starks, K.M.; Cooke, R.A. Effectiveness of constructed wetlands in reducing nitrogen and phosphorus export from agricultural tile drainage. J. Environ. Qual. 2000, 29, 1262–1274. [Google Scholar] [CrossRef] [Green Version]
- Kroeger, A.C.; Madramootoo, C.A.; Enright, P.; Laflamme, C. Efficiency of a small constructed wetland in southern Québec for treatment of agricultural runoff waters. In IWA Specialist Conference: Wastewater Biosolids Sustainability: Technical, Managerial, and Public Synergy; ResearchGate: Moncton, NB, Canada, 2007; pp. 1057–1062. [Google Scholar]
- Tanner, C.C.; Sukias, J.P.S. Multiyear nutrient removal performance of three constructed wetlands intercepting tile drain flows from grazed pastures. J. Environ. Qual. 2011, 40, 620–633. [Google Scholar] [CrossRef]
- Tanner, C.C.; Nguyen, M.; Sukias, J. Nutrient removal by a constructed wetland treating subsurface drainage from grazed dairy pasture. Agric. Ecosyst. Environ. 2005, 105, 145–162. [Google Scholar] [CrossRef]
- Kovacic, D.A.; Twait, R.M.; Wallace, M.P.; Bowling, J.M. Use of created wetlands to improve water quality in the Midwest—Lake Bloomington case study. Ecol. Eng. 2006, 28, 258–270. [Google Scholar] [CrossRef]
- Tanner, C.C.; Nguyen, M.L.; Sukias, J. Using constructed wetlands to treat subsurface drainage from intensively grazed dairy pastures in New Zealand. Water Sci. Technol. 2003, 48, 207–213. [Google Scholar] [CrossRef]
- Reinhardt, M.; Müller, B.; Gächter, R.; Wehrli, B. Nitrogen removal in a small constructed wetland: An isotope mass balance approach. Environ. Sci. Technol. 2006, 40, 3313–3319. [Google Scholar] [CrossRef]
- Kadlec, R.H. Constructed wetlands to remove nitrate. In Nutrient Management in Agricultural Watersheds: A Wetlands Solution; Dunne, E.J., Reddy, K.R., Carton, O.T., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2005; pp. 132–143. [Google Scholar]
- Kadlec, R.H. Nitrogen farming for pollution control. J. Environ. Sci. Heal. Part A 2005, 40, 1307–1330. [Google Scholar] [CrossRef] [PubMed]
- O’Geen, A.; Budd, R.; Gan, J.; Maynard, J.; Parikh, S.; Dahlgren, R. Mitigating nonpoint source pollution in agriculture with constructed and restored wetlands. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2010; pp. 1–76. [Google Scholar]
- Crumpton, W.G.; Kovacic, D.A.; Hey, D.L.; Kostel, J.A. Potential of restored and constructed wetlands to reduce nutrient export from agricultural watersheds in the corn belt. In Final Report: Gulf Hypoxia and Local Water Quality Concerns Workshop; American Society of Agricultural and Biological Engineers: St. Joseph, MI, USA, 2008; pp. 29–42. [Google Scholar]
- Khan, I. Effects of Hydraulic Load on Nitrate Removal in Surface-Flow Constructed Wetlands; Halmstad University: Halmstad, Sweden, 2011. [Google Scholar]
- Bastviken, S.K.; Weisner, S.E.B.; Thiere, G.; Svensson, J.M.; Ehde, P.M.; Tonderski, K.S. Effects of vegetation and hydraulic load on seasonal nitrate removal in treatment wetlands. Ecol. Eng. 2009, 35, 946–952. [Google Scholar] [CrossRef]
- Kadlec, R.H.; Wallace, S. Treatment Wetlands, 2nd ed.; Taylor & Francis Group: Boca Raton, FL, USA, 2009. [Google Scholar]
- Song, X.; Ehde, P.M.; Weisner, S.E.B. Effects of water depth and phosphorus availability on nitrogen removal in agricultural wetlands. Water 2019, 11, 2626. [Google Scholar] [CrossRef] [Green Version]
- Guo, C.; Cui, Y.; Dong, B.; Luo, Y.; Liu, F.; Zhao, S.; Wu, H. Test study of the optimal design for hydraulic performance and treatment performance of free water surface flow constructed wetland. Bioresour. Technol. 2017, 238, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Cui, Y.; Shi, Y.; Luo, Y.; Liu, F.; Wan, D.; Ma, Z. Improved test to determine design parameters for optimization of free surface flow constructed wetlands. Bioresour. Technol. 2019, 280, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Haverstock, M.J. An Assessment of a Wetland-Reservoir Wastewater Treatment and Reuse System Receiving Agricultural Drainage Water in Nova Scotia; Dalhousie University: Halifax, NS, Canada, 2010. [Google Scholar]
- Tanner, C.C.; Kadlec, R.H. Influence of hydrological regime on wetland attenuation of diffuse agricultural nitrate losses. Ecol. Eng. 2013, 56, 79–88. [Google Scholar] [CrossRef]
- Tanner, C.C.; Sukias, J.P.S.; Yates, C.R. New Zealand Guidelines: Constructed Wetland Treatment of Tile Drainage; NIWA Information Series No. 75; National Institute of Water & Atmospheric Research Ltd.: Hamilton, New Zealand, 2010. [Google Scholar]
- Persson, J.; Somes, N.L.G.; Wong, T.H.F. Hydraulics efficiency of constructed wetlands and ponds. Water Sci. Technol. 1999, 40, 291–300. [Google Scholar] [CrossRef]
- Thackston, E.L.; Shields, F.D.; Schroeder, P.R. Residence time distributions of shallow basins. J. Environ. Eng. 1987, 113, 1319–1332. [Google Scholar] [CrossRef]
- Lavrnić, S.; Alagna, V.; Iovino, M.; Anconelli, S.; Solimando, D.; Toscano, A. Hydrological and hydraulic behaviour of a surface flow constructed wetland treating agricultural drainage water in northern Italy. Sci. Total. Environ. 2020, 702, 134795. [Google Scholar] [CrossRef]
- Su, T.-M.; Yang, S.-C.; Shih, S.-S.; Lee, H.-Y. Optimal design for hydraulic efficiency performance of free-water-surface constructed wetlands. Ecol. Eng. 2009, 35, 1200–1207. [Google Scholar] [CrossRef]
- Pugliese, L.; Kusk, M.; Iversen, B.V.; Kjaergaard, C. Internal hydraulics and wind effect in a surface flow constructed wetland receiving agricultural drainage water. Ecol. Eng. 2020, 144, 105661. [Google Scholar] [CrossRef]
- Bodin, H.; Mietto, A.; Ehde, P.M.; Persson, J.; Weisner, S.E.B. Tracer behaviour and analysis of hydraulics in experimental free water surface wetlands. Ecol. Eng. 2012, 49, 201–211. [Google Scholar] [CrossRef] [Green Version]
- Kalcic, M.; Crumpton, W.; Liu, X.; D’Ambrosio, J.; Ward, A.; Witter, J. Assessment of beyond-the-field nutrient management practices for agricultural crop systems with subsurface drainage. J. Soil Water Conserv. 2018, 73, 62–74. [Google Scholar] [CrossRef] [Green Version]
- Grebliunas, B.D.; Perry, W.L. The role of C:N:P stoichiometry in affecting denitrification in sediments from agricultural surface and tile-water wetlands. Springerplus 2016, 5, 359. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Kovacic, D.A.; David, M.B.; Gentry, L.E.; Mulvaney, R.L.; Lindau, C.W. In Situ Measurements of Denitrification in Constructed Wetlands. J. Environ. Qual. 1999, 28, 263–269. [Google Scholar] [CrossRef] [Green Version]
- Matheson, F.E.; Sukias, J.P. Nitrate removal processes in a constructed wetland treating drainage from dairy pasture. Ecol. Eng. 2010, 36, 1260–1265. [Google Scholar] [CrossRef]
- David, M.B.; Gentry, L.E.; Smith, K.M.; Kovacic, D.A. Carbon, plant, and temperature control of nitrate removal from wetland mesocosms. Trans. Ill. State Acad Sci. 1997, 90, 103–112. [Google Scholar]
- Rutherford, K.; Wheeler, D. Wetland nitrogen removal modules in OVERSEER®. In Adding to the Knowledge Base for the Nutrient Manager Occasion Report No 24; Currie, L.D., Christensen, C.L., Eds.; Fertilizer and Lime Research Centre, Massey University: Palmerston North, New Zealand, 2011; p. 12. [Google Scholar]
- Nilsson, J.E.; Liess, A.; Ehde, P.M.; Weisner, S.E. Mature wetland ecosystems remove nitrogen equally well regardless of initial planting. Sci. Total. Environ. 2020, 716, 137002. [Google Scholar] [CrossRef] [PubMed]






| N form | Removal | Number of Observations | R2 | p-Value | p-Value | Coefficient | ||
|---|---|---|---|---|---|---|---|---|
| Hydraulic Load | Concentration | Hydraulic Load | Concentration | |||||
| Total N | Rate | 21 | 0.26 | 0.06 | 0.03 * | 0.07 | 2.76 | 12.33 |
| Efficiency | 22 | 0.25 | 0.06 | 0.02 * | 0.49 | −0.65 | −1.00 | |
| NO3− | Rate | 22 | 0.55 | 0.00 * | 0.00 * | 0.04 * | 3.38 | 8.03 |
| Efficiency | 22 | 0.07 | 0.48 | 0.31 | 0.95 | −0.26 | −0.09 | |
| NH4+ | Rate | 16 | 0.14 | 0.38 | 0.17 | 0.68 | −0.51 | 5.51 |
| Efficiency | 16 | 0.14 | 0.36 | 0.23 | 0.29 | −2.07 | 65.38 | |
| Organic N | Rate | 15 | 0.00 | 0.98 | 0.93 | 0.86 | −0.15 | 5.79 |
| Efficiency | 12 | 0.51 | 0.04 * | 0.03 * | 0.16 | −3.37 | 51.07 | |
| Name (Scale) | Denitrification | Biological Uptake | Settling | Ammonification | Nitrification | DNRA * | NO3− Concentration | Temperature | Carbon in Sediments | Period | Study |
|---|---|---|---|---|---|---|---|---|---|---|---|
| mg m−2 h−1 (%) | mg L−1 | °C | g m−2 | d | |||||||
| Gully (Laboratory) a | 38 b | - | - | - | - | - | 2 [10 c] | 25 | - | 20 | [66] |
| Gully (Laboratory) a | 103 b | - | - | - | - | - | 2 [40 c] | 25 | - | 20 | [66] |
| Gully (Laboratory) a | 24 b | - | - | - | - | - | 20 [10 c] | 25 | - | 20 | [66] |
| Gully (Laboratory) a | 221 b | - | - | - | - | - | 20 [40 c] | 25 | - | 20 | [66] |
| Moga, Durbin and Gully (Laboratory) a | 64 b | - | - | - | - | - | 10 [20 c] | 25 | - | 20 | [66] |
| Moga, Durbin and Gully (Laboratory) a | 78 b | - | - | - | - | - | 10 [40 c] | 25 | - | 20 | [66] |
| - (Mesocosm 1) | ≤9.3 (58) | 0.3 (5.5) d+e | - | - | - | - | 15 | - | - | 33 | [67] |
| - (Mesocosm 2) | 0.5 (9.9) d+e | - | - | - | - | [67] | |||||
| Wetland 1 (Mesocosm) | 2.1 | - | - | - | - | - | 9–20 | 24 f | - | 0.7 | [67] |
| Wetland 1 (Mesocosm) | 6.2 | - | - | - | - | - | 9–20 | 18 f | 9.1 e | 0.1 | [67] |
| Wetland 1 (Mesocosm) | 11.8 | - | - | - | - | - | 9–20 | 25 f | 9.5 e | 0.1 | [67] |
| Wetland 2 (Mesocosm) | 2.7 | - | - | - | - | - | 9–20 | 25 f | - | 0.7 | [67] |
| Wetland 2 (Mesocosm) | 2.0 | - | - | - | - | - | 9–20 | 4 f | 2.1 e | 0.1 | [67] |
| Wetland 2 (Mesocosm) | 3.9 | - | - | - | - | - | 9–20 | 17 f | 5.0 e | 0.1 | [67] |
| Wetland 2 (Mesocosm) | 9.0 | - | - | - | - | - | 9–20 | 20 f | 9.3 e | 0.1 | [67] |
| Toenepi (Mesocosm) | 7.2 (39.3) | 1.9 (10.2) d+e+g+h+i | - | - | - | 0.2 (1.0) e | 10.4 | 14 j | - | 6 | [68] |
| Toenepi (Field) | 6.2 b | - | - | - | - | - | 10.8 | - | 61 k | 365 | [42] |
| Boden (Field) | 2.9 | 0.7 i | 4.3 | 1.1 | 3.8 | - | 1.9–3.0 | 10 | - | 210 | [45] |
| Boden (Laboratory) a | ≤28.8 | - | - | - | - | - | 1.9–15.6 | 9 | - | 4.8 | [45] |
| - (Laboratory) a | 3.6 | - | - | - | - | - | 50 | 20 | 9.7 k | <5 | [22] |
| - (Field) | 0.3 (6.3 l) | 2.5 (53.9 l) d+h | 1.3 (27.7 l) m | - | - | - | 1.4 | −1.5–27.2 | - | 1825 | [23] |
| - (Field) | - | 0.0 b, d+h | 0.01 b, n | - | - | - | 11.8 l | 2.7–24.3 | - | 1825 | [24] |
| Wetland B (Field) | - | 0.005 d+h | 0.0 | - | - | - | 7.5 | - | - | 153 | [25] |
| - (Field) | - | - | 2.7 | - | - | - | 7.5 | 9 | 144.2 | 577 | [26] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendes, L.R.D. Nitrogen Removal from Agricultural Subsurface Drainage by Surface-Flow Wetlands: Variability. Processes 2021, 9, 156. https://doi.org/10.3390/pr9010156
Mendes LRD. Nitrogen Removal from Agricultural Subsurface Drainage by Surface-Flow Wetlands: Variability. Processes. 2021; 9(1):156. https://doi.org/10.3390/pr9010156
Chicago/Turabian StyleMendes, Lipe Renato Dantas. 2021. "Nitrogen Removal from Agricultural Subsurface Drainage by Surface-Flow Wetlands: Variability" Processes 9, no. 1: 156. https://doi.org/10.3390/pr9010156
APA StyleMendes, L. R. D. (2021). Nitrogen Removal from Agricultural Subsurface Drainage by Surface-Flow Wetlands: Variability. Processes, 9(1), 156. https://doi.org/10.3390/pr9010156