
Performance of Single and Two-Stage Cross-Flow Ultrafiltration Membrane in Fractionation of Peptide from Microalgae Protein Hydrolysate (Nannochloropsis gaditana)

 ,
,
Abstract
:1. Introduction
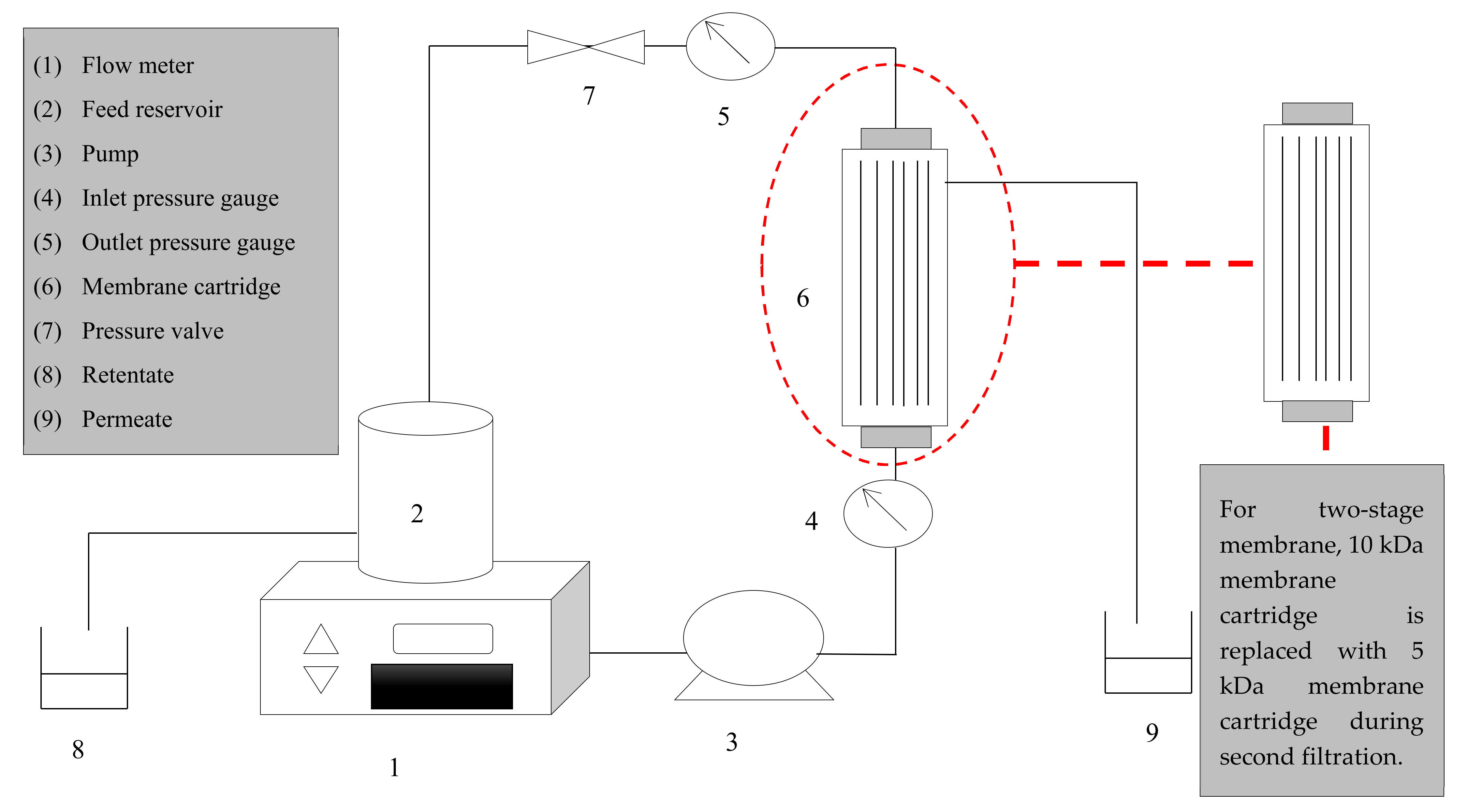
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Preparation of Microalgae Protein Hydrolysate
2.3. Preparation of Microalgae Protein Hydrolysate
2.4. Membrane Sanitization
2.5. Evaluation of Membrane Performance (Permeate Flux and Peptide Transmission)
2.5.1. Permeate Flux
2.5.2. Peptide Transmission
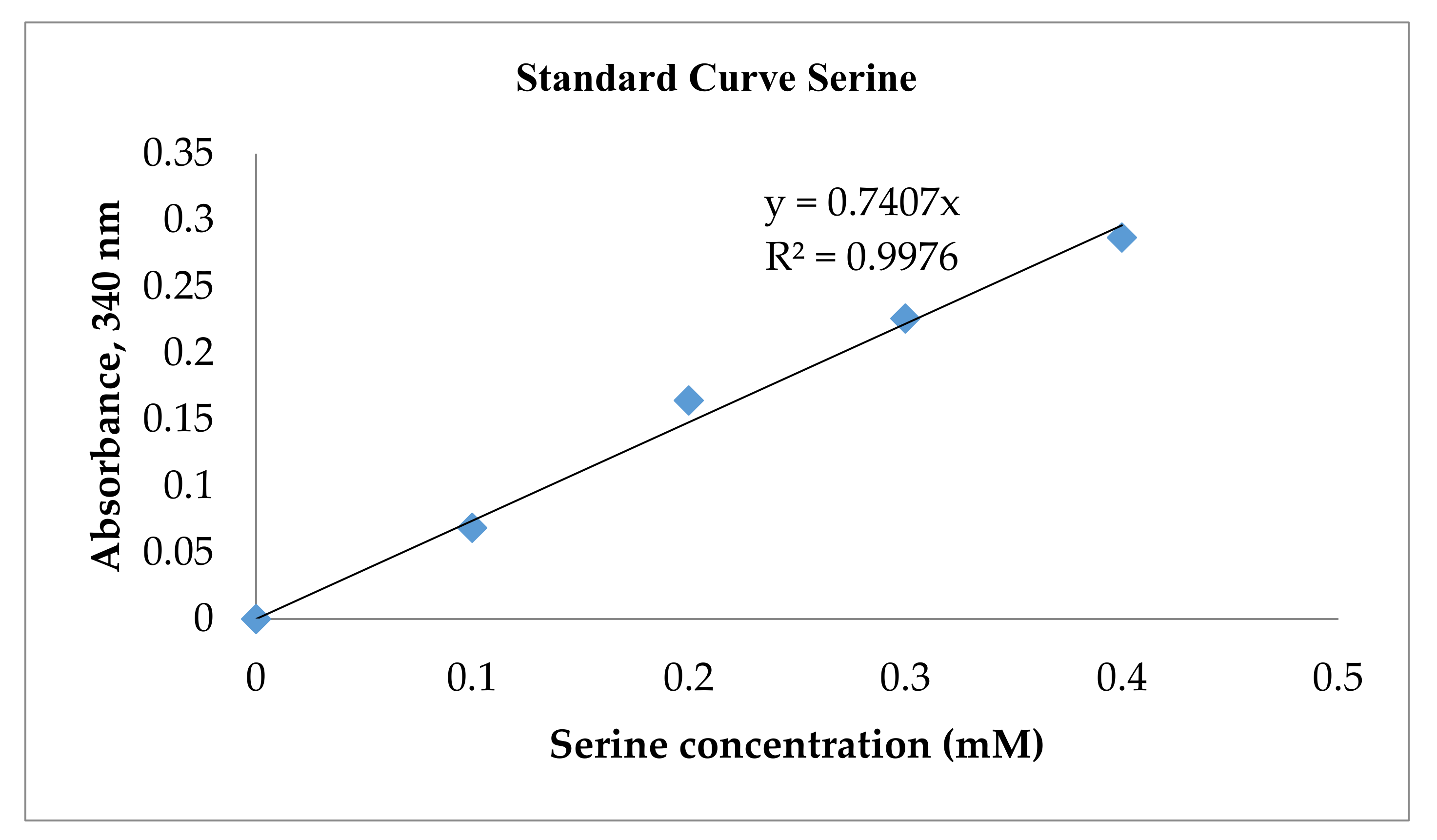
2.6. Measurement of Peptide Content
2.7. Fractionation Process
2.8. Peptide Selectivity
3. Results and Discussion
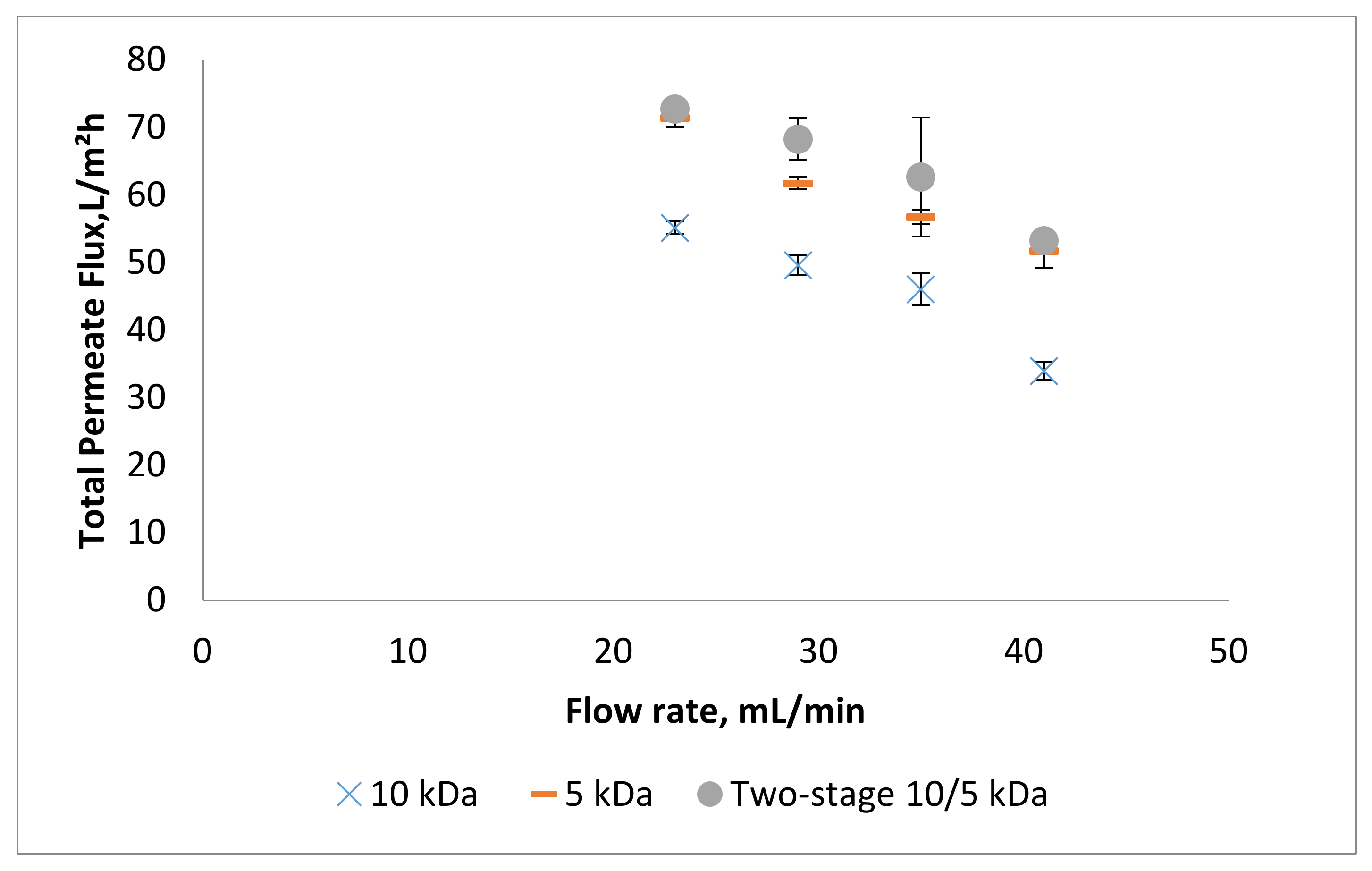
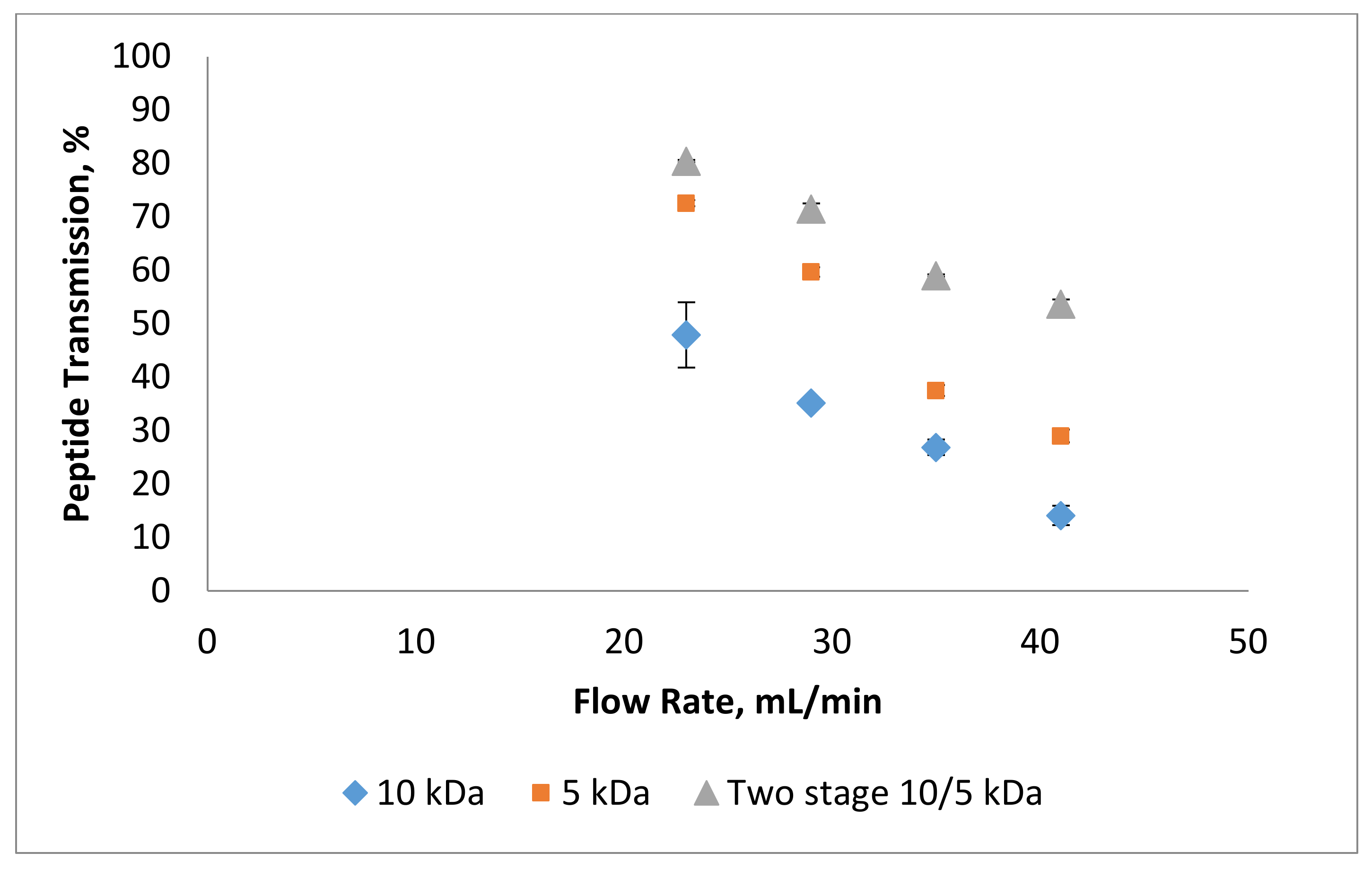
3.1. Effect of Flow Rate on Permeate Flux and Peptide Transmission in Fractionating MPH
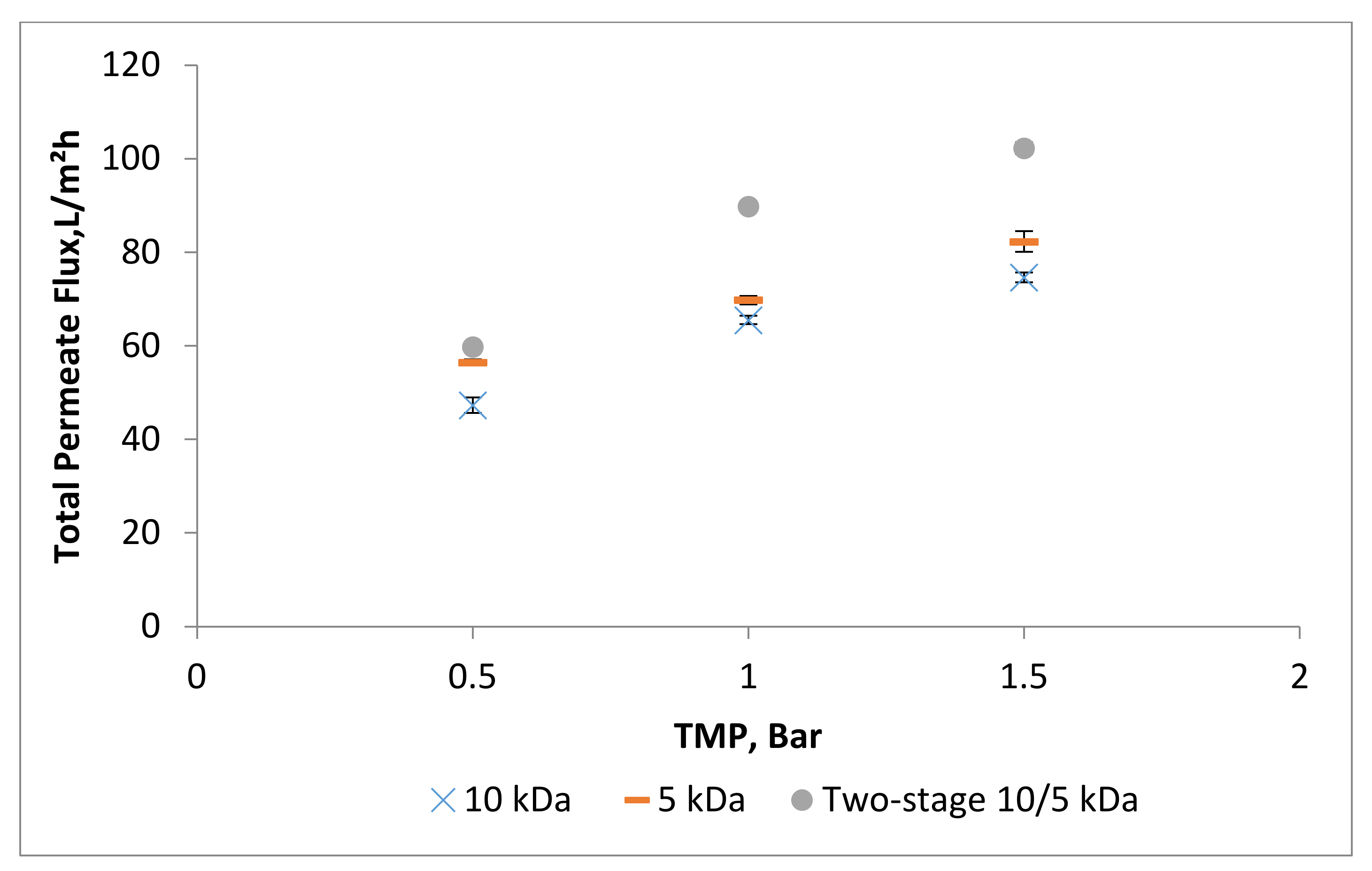
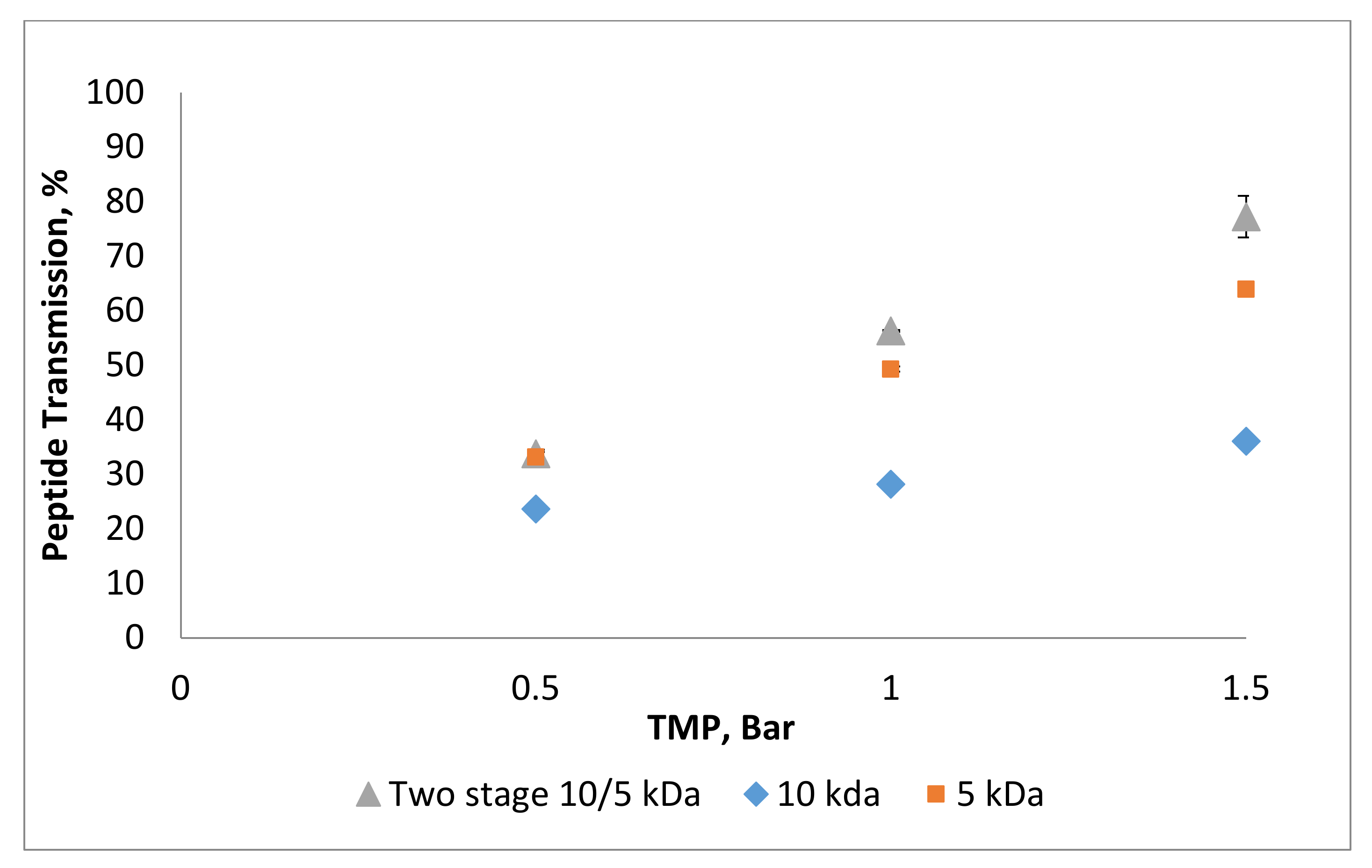
3.2. Effect of Trans-Membrane Pressure on Permeate Flux and Peptide Transmission in Fractionating MPH
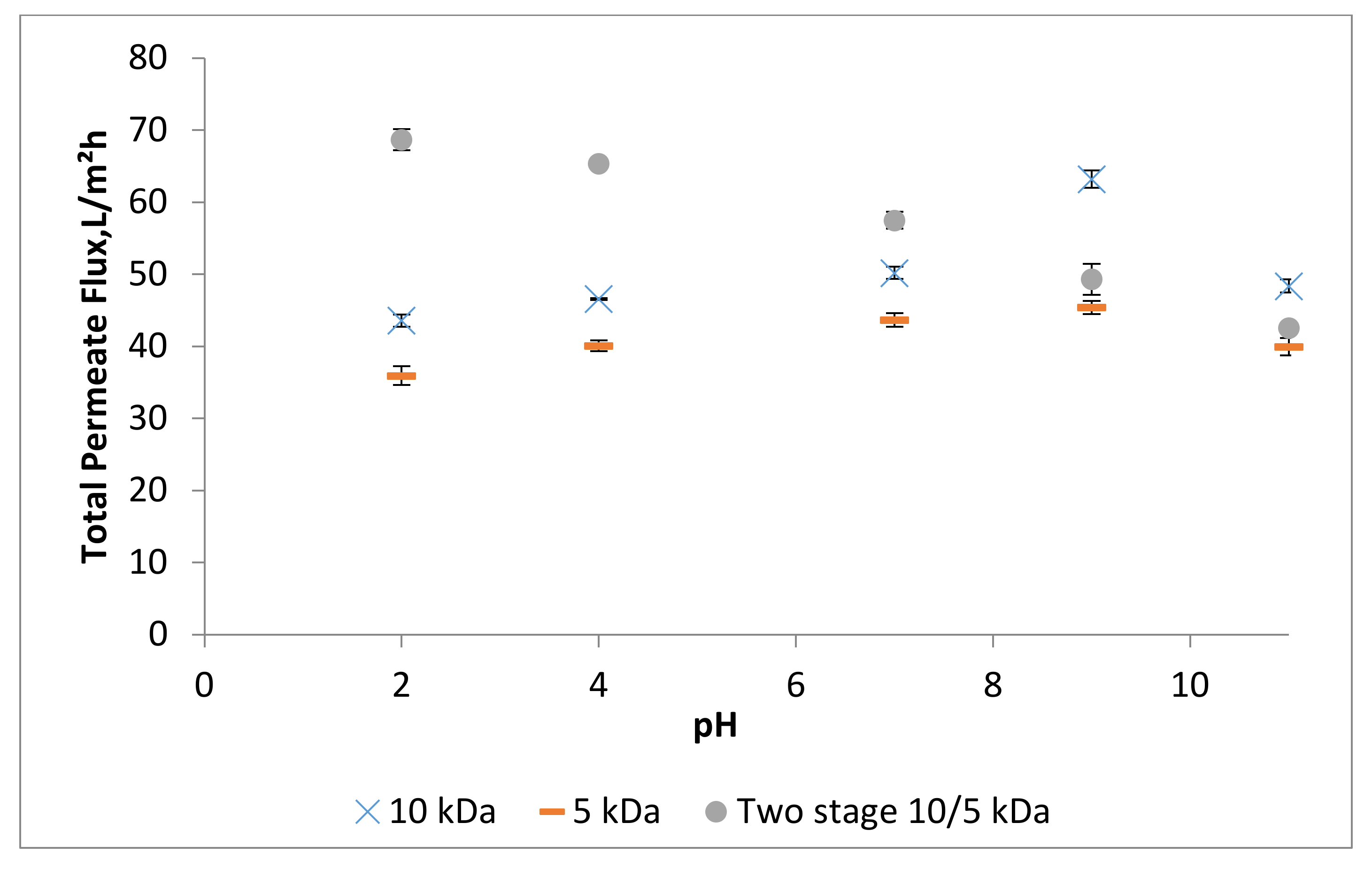
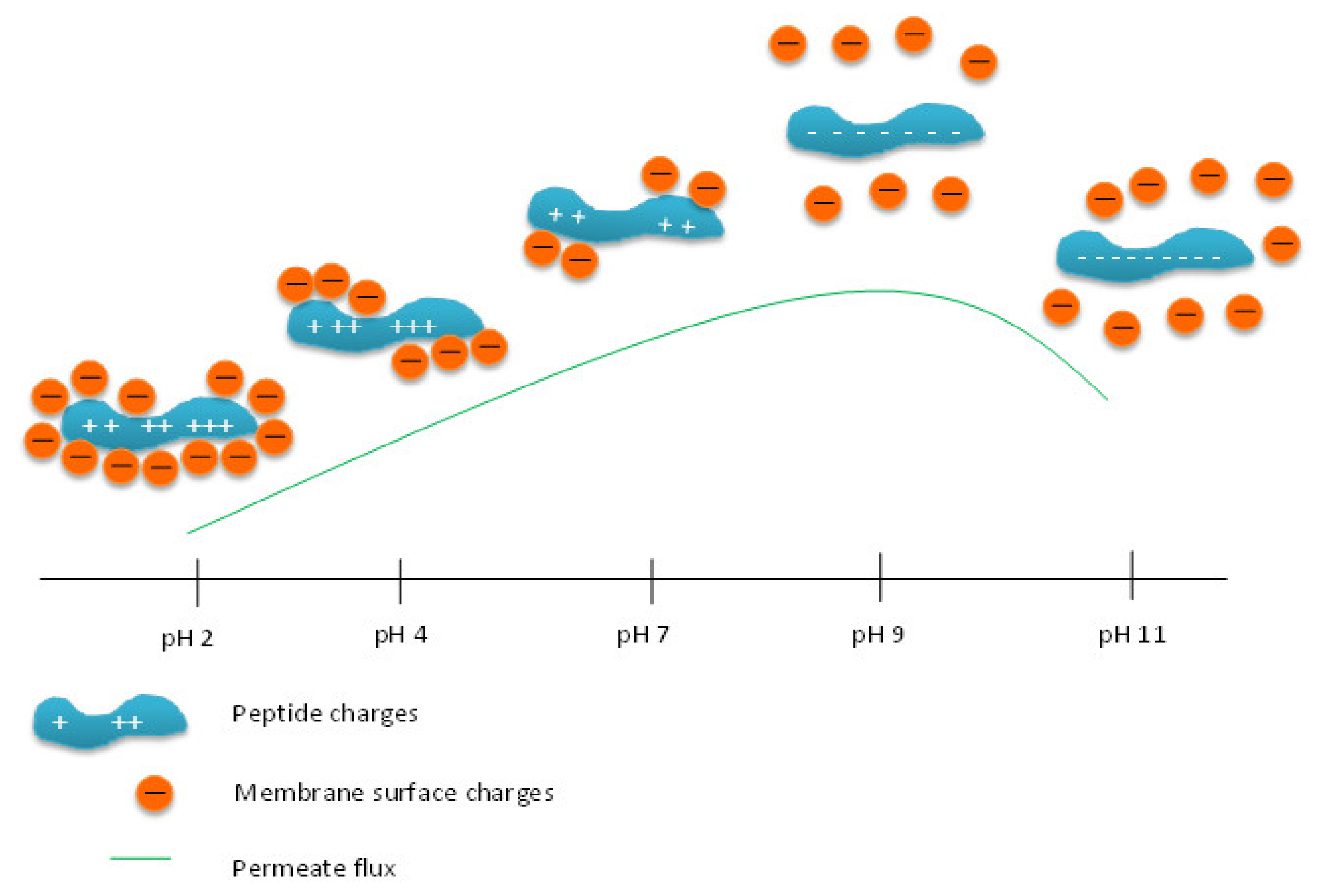
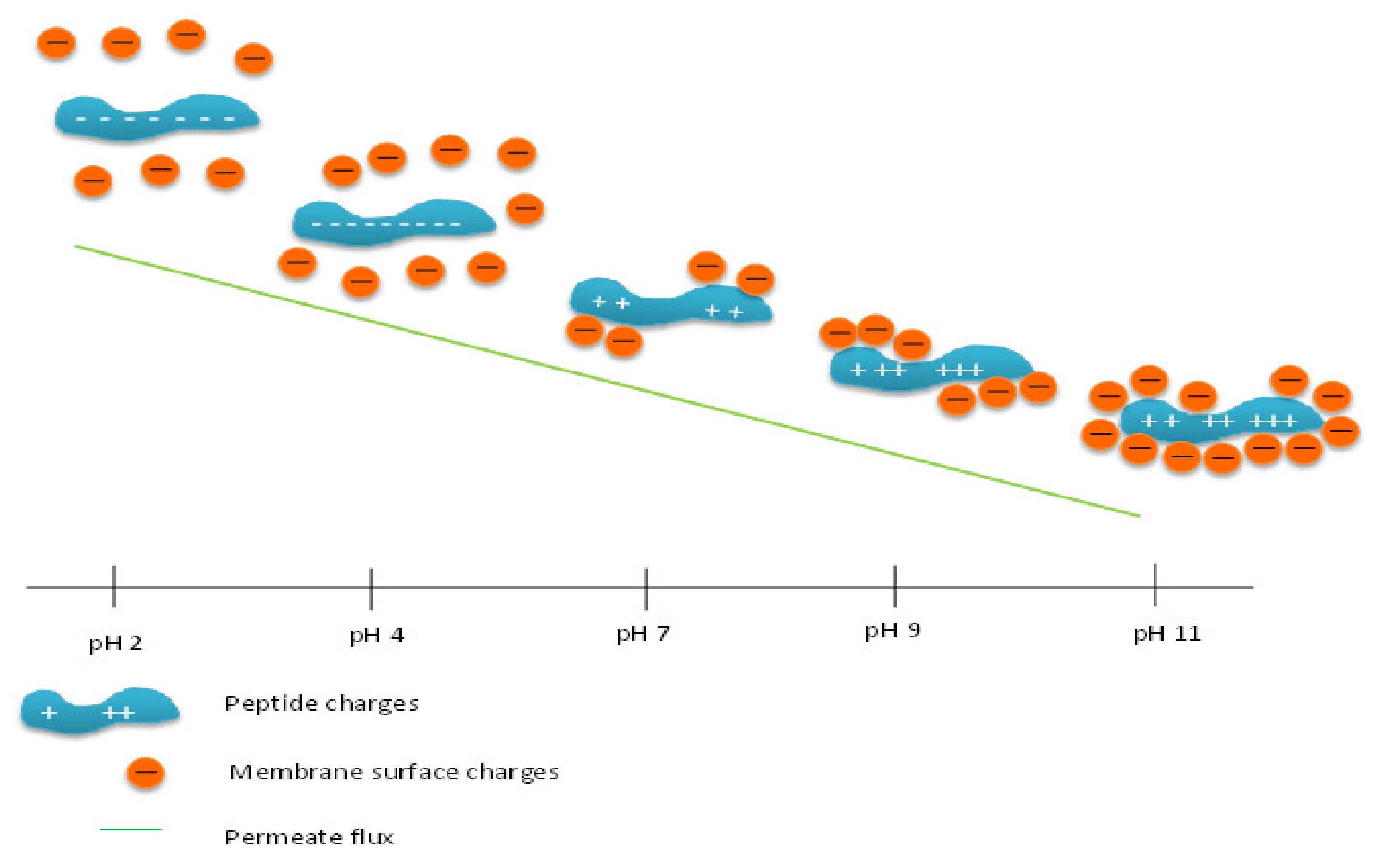
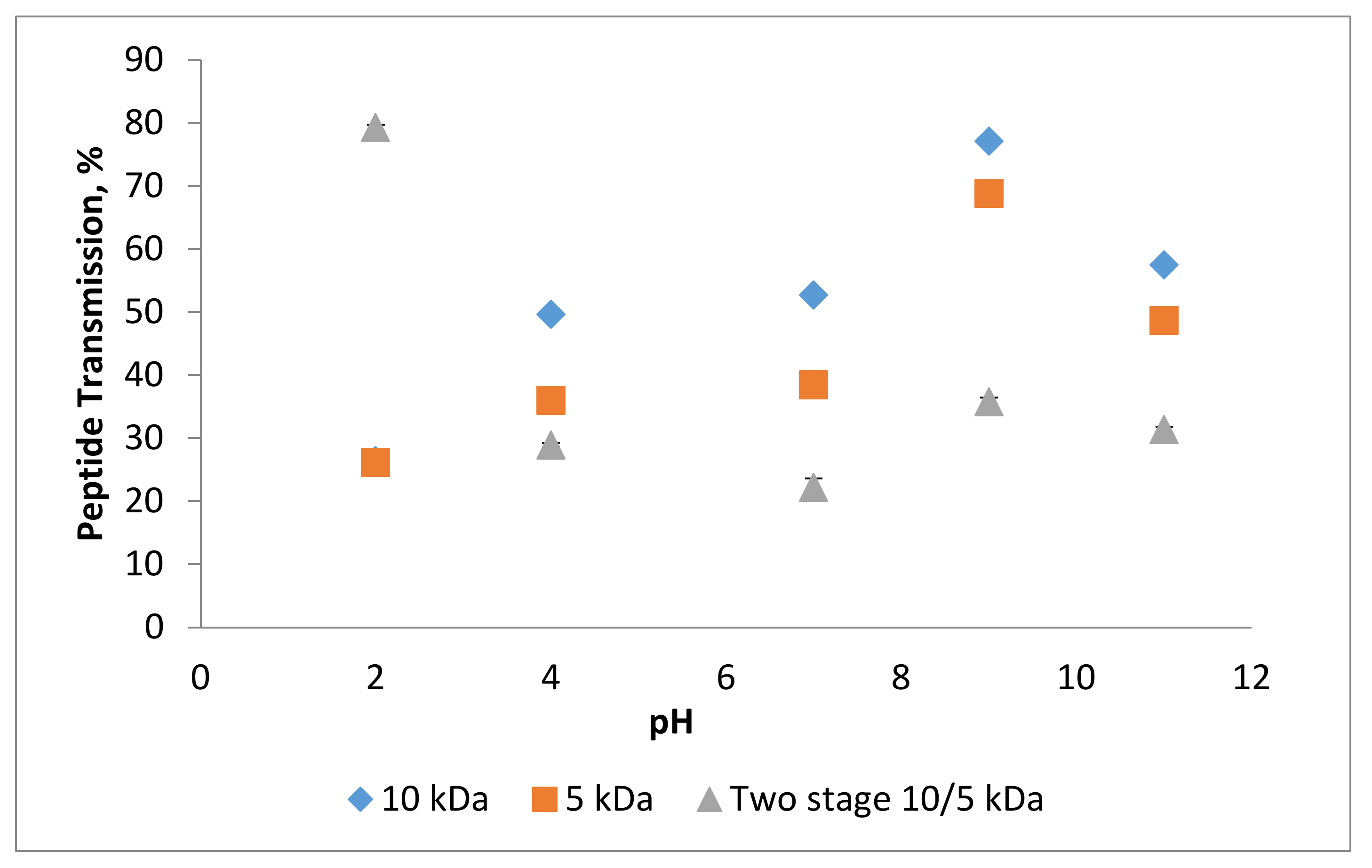
3.3. Effect of pH on Permeate Flux and Peptide Transmission in Fractionating MPH
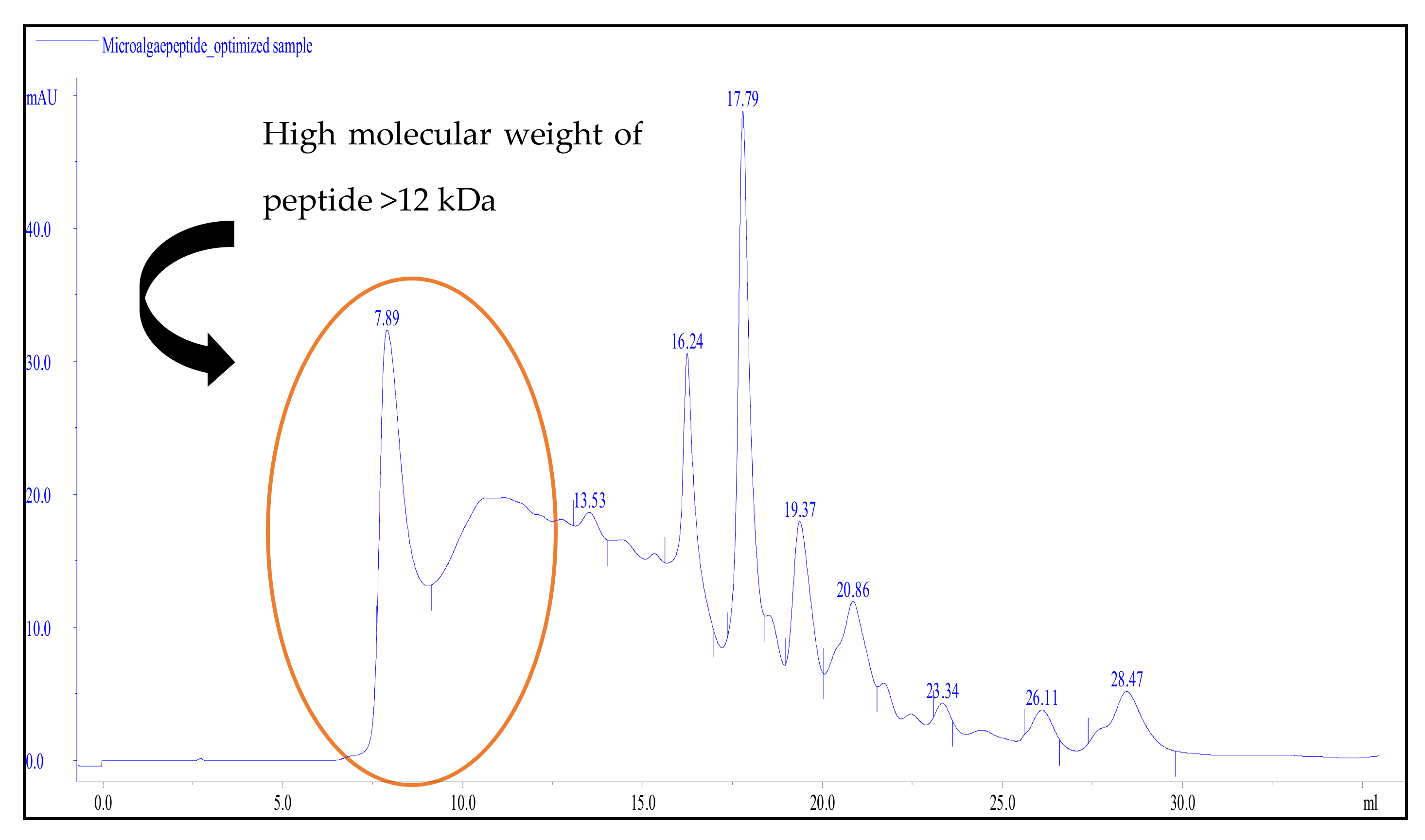
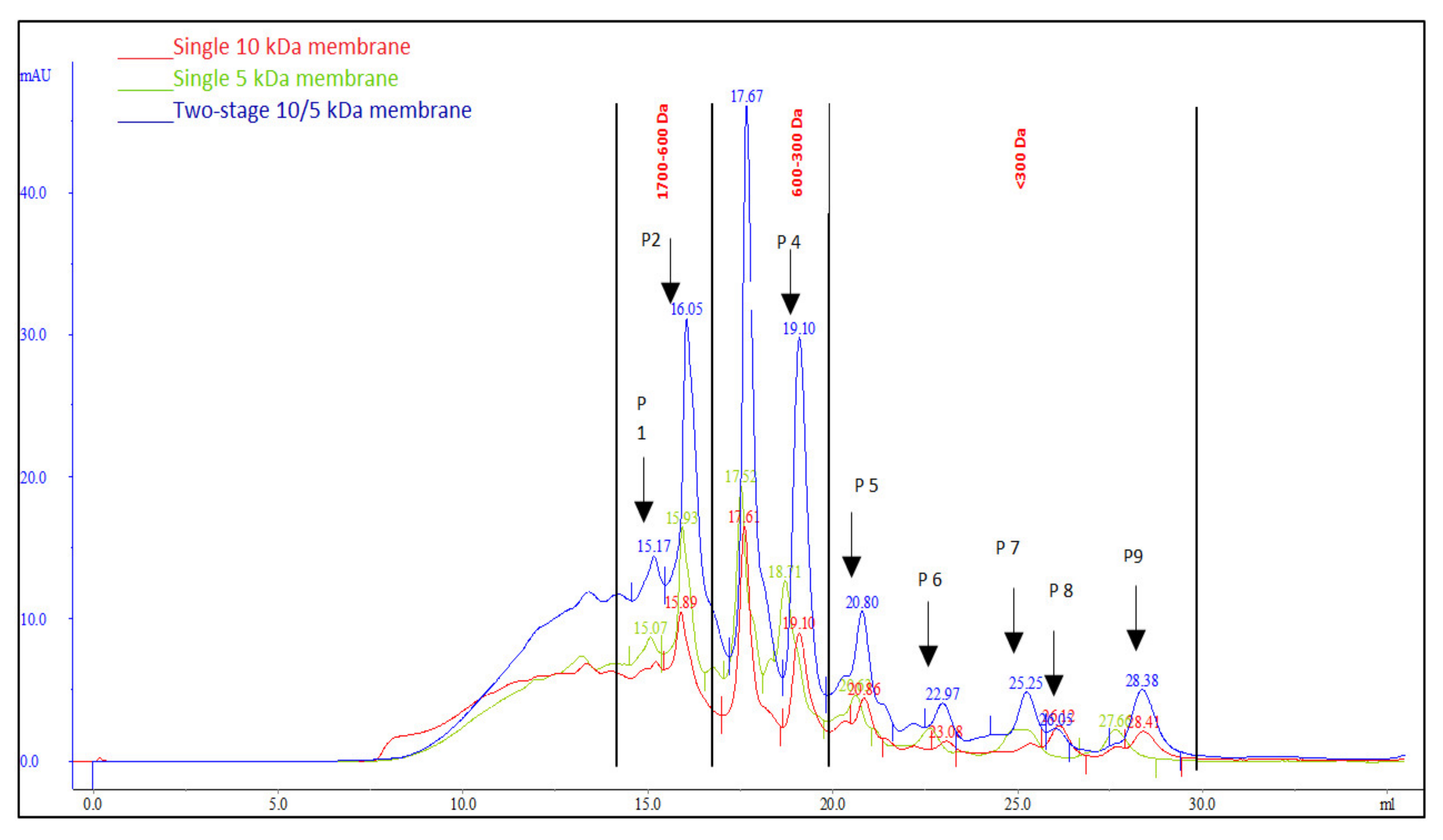
3.4. Peptide Selectivity of MPH Fractionation
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chen, C.Y.; Zhao, X.Q.; Yen, H.W.; Ho, S.H.; Cheng, C.L.; Lee, D.J.; Bai, F.W.; Chang, J.S. Microalgae-based carbohydrates for biofuel production. Biochem. Eng. J. 2013, 78, 1–10. [Google Scholar] [CrossRef]
- Demirbas, A. Use of algae as biofuel sources. Energy Convers. Manag. 2010, 51, 2738–2749. [Google Scholar] [CrossRef]
- Shebis, Y.; Iluz, D.; Kinel-Tahan, Y.; Dubinsky, Z.; Yehoshua, Y. Natural Antioxidants: Function and Sources. Food Nutr. Sci. 2013, 4, 643–649. [Google Scholar] [CrossRef] [Green Version]
- De Jesus Raposo, M.F.; de Morais, R.M.S.C.; de Morais, A.M.M.B. Health applications of bioactive compounds from marine microalgae. Life Sci. 2013, 93, 479–486. [Google Scholar] [CrossRef] [PubMed]
- Munir, N.; Sharif, N.; Naz, S.; Manzoor, F. Algae: A potent antioxidant source. Sky J. Microbiol. Res. 2013, 1, 22–31. [Google Scholar]
- Molino, A.; Iovine, A.; Casella, P.; Mehariya, S.; Chianese, S.; Cerbone, A.; Rimauro, J.; Musmarra, D. Microalgae Characterization for Consolidated and New Application in Human Food, Animal Feed and Nutraceuticals. Int. J. Environ. Res. Public Health 2018, 15, 2436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Morais, M.G.; Vaz, B.D.S.; de Morais, E.G.; Costa, J.A.V. Biologically Active Metabolites Synthesized by Microalgae. BioMed Res. Int. 2015, 2015, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Korhonen, H.; Pihlanto, A. Bioactive peptides: Production and functionality. Int. Dairy J. 2006, 16, 945–960. [Google Scholar] [CrossRef]
- Liu, Y.; Wan, S.; Liu, J.; Zou, Y.; Liao, S. Antioxidant Activity and Stability Study of Peptides from Enzymatically Hydrolyzed Male Silkmoth. J. Food Process. Preserv. 2017, 41, e13081. [Google Scholar] [CrossRef]
- Wijesinghe, W.A.J.P.; Jeon, Y.J. Fitoterapia Enzyme-assistant extraction (EAE) of bioactive components: A useful approach for recovery of industrially important metabolites from seaweeds: A review. Fitoterapia 2012, 83, 6–12. [Google Scholar] [CrossRef]
- Bazinet, L.; Firdaous, L. Membrane Processes and Devices for Separation of Bioactive Peptides. Recent Pat. Biotechnol. 2009, 3, 61–72. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; He, J.; Zhuang, Y.; Sun, L. Purification and Identification of Antioxidant Peptides from Enzymatic Hydrolysates of Tilapia (Oreochromis niloticus) Frame Protein. Molecules 2012, 17, 12836–12850. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Sharma, N.; Ranjan, R.; Kumar, S.; Bhat, Z.F.; Jeong, D.K. Perspective of Membrane Technology in Dairy Industry: A Review. J. Anim. Sci. 2013, 26, 1347–1358. [Google Scholar] [CrossRef] [Green Version]
- Firdaous, L.; Dhulster, P.; Amiot, J.; Gaudreau, A.; Lecouturier, D.; Kapel, R.; Lutin, F.; Vézina, L.-P.; Bazinet, L. Concentration and selective separation of bioactive peptides from an alfalfa white protein hydrolysate by electrodialysis with ultrafiltration membranes. J. Membr. Sci. 2009, 329, 60–67. [Google Scholar] [CrossRef]
- Microdyn-Nadir. Membrane Filtration Proceses Dead-End vs Cross-Flow Bulletin; Microdyn-Nadir: Wiesbaden, Germany, 2017. [Google Scholar]
- Hasan, A.; Peluso, C.R.; Hull, T.S.; Fieschko, J.; Chatterjee, S.G. A surface-renewal model of cross-flow microfiltration. Braz. J. Chem. Eng. 2013, 30, 167–186. [Google Scholar] [CrossRef]
- Poulin, J.-F.; Amiot, J.; Bazinet, L. Impact of Feed Solution Flow Rate on Peptide Fractionation by Electrodialysis with Ultrafiltration Membrane. J. Agric. Food Chem. 2008, 56, 2007–2011. [Google Scholar] [CrossRef]
- Poulin, J.-F.; Amiot, J.; Bazinet, L. Improved peptide fractionation by electrodialysis with ultrafiltration membrane: Influence of ultrafiltration membrane stacking and electrical field strength. J. Membr. Sci. 2007, 299, 83–90. [Google Scholar] [CrossRef]
- Atkinson, A.J.; Wang, J.; Grzebyk, K.; Zhang, Z.; Jung, D.; Zeng, D.; Pollard, A.; Gold, A.; Coronell, O. Scalable fabrication of anti-biofouling membranes through 2-aminoimidazole incorporation during polyamide casting. J. Membr. Sci. 2019, 579, 151–161. [Google Scholar] [CrossRef]
- Khairkar, S.R.; Pansare, A.V.; Shedge, A.A.; Chhatre, S.Y.; Suresh, A.K.; Chakrabarti, S.; Patil, V.R.; Nagarkar, A.A. Chemosphere Hydrophobic interpenetrating polyamide-PDMS membranes for desalination, pesticides removal and enhanced chlorine tolerance. Chemosphere 2020, 258, 127179. [Google Scholar] [CrossRef] [PubMed]
- Firouzjaei, M.D.; Seyedpour, S.F.; Aktij, S.A.; Giagnorio, M.; Bazrafshan, N.; Mollahosseini, A.; Samadi, F.; Ahmadalipour, S.; Firouzjaei, F.D.; Esfahani, M.R.; et al. Recent advances in functionalized polymer membranes for biofouling control and mitigation in forward osmosis. J. Membr. Sci. 2020, 596, 117604. [Google Scholar] [CrossRef]
- Roslan, J.; Kamal, S.M.M.; Yunos, K.F.M.; Abdullah, N. Assessment on multilayer ultrafiltration membrane for fractionation of tilapia by-product protein hydrolysate with angiotensin I-converting enzyme (ACE) inhibitory activity. Sep. Purif. Technol. 2017, 173, 250–257. [Google Scholar] [CrossRef]
- Cheang, B.; Zydney, A.L. A two-stage ultrafiltration process for fractionation of whey protein isolate. J. Membr. Sci. 2004, 231, 159–167. [Google Scholar] [CrossRef]
- Nor, M.Z.M.; Ramchandran, L.; Duke, M.; Vasiljevic, T. Performance of a Two-Stage Membrane System for Bromelain Separation from Pineapple Waste Mixture as Impacted by Enzymatic Pretreatment and Diafiltration. Food Technol. Biotechnol. 2017, 56, 218–227. [Google Scholar] [CrossRef]
- Safi, C.; Liu, D.Z.; Yap, B.H.J.; Martin, G.J.O.; Vaca-Garcia, C.; Pontalier, P.Y. A two-stage ultrafiltration process for separating multiple components of Tetraselmis suecica after cell disruption. J. Appl. Phycol. 2014, 26, 2379–2387. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, H.; Joshi, R.; Gupta, M. Isolation and characterisation of enzymatic hydrolysed peptides of green tender sorghum and their antioxidant activities. LWT Food Sci. Technol. 2017, 84, 1–9. [Google Scholar] [CrossRef]
- Cheung, R.C.F.; Ng, T.B.; Wong, J.H. Marine Peptides: Bioactivities and Applications. Mar. Drugs 2015, 13, 4006–4043. [Google Scholar] [CrossRef]
- Wang, Y.-N.; Tang, C.Y. Fouling of Nanofiltration, Reverse Osmosis, and Ultrafiltration Membranes by Protein Mixtures: The Role of Inter-Foulant-Species Interaction. Environ. Sci. Technol. 2011, 45, 6373–6379. [Google Scholar] [CrossRef]
- Zain, M.M.; Mohammad, A.W.; Hairom, N.H.H. Flux and Permeation Behaviour of Ultrafiltration in Sugaring Out Cellulose Hydrolysate Solution: A Membrane Screening. J. Phys. Sci. 2017, 28, 25–38. [Google Scholar] [CrossRef] [Green Version]
- Roslan, J.; Kamal, S.M.M.; Yunos, K.F.M.; Abdullah, N. Evaluation on performance of dead-end ultrafiltration membrane in fractionating tilapia by-product protein hydrolysate. Sep. Purif. Technol. 2017, 195, 21–29. [Google Scholar] [CrossRef]
- Yunos, K.M.; Field, R. Rejection amplification in the ultrafiltration of binary protein mixtures using sandwich configurations. Chem. Eng. Process. Process Intensif. 2008, 47, 1053–1060. [Google Scholar] [CrossRef]
- Church, F.C.; Swaisgood, H.E.; Porter, D.H.; Catignani, G.L. Spectrophotometric Assay Using o-Phthaldialdehyde for Determination of Proteolysis in Milk and Isolated Milk Proteins. J. Dairy Sci. 1983, 66, 1219–1227. [Google Scholar] [CrossRef]
- Hernández, M.J.M.; Camañas, R.M.V.; Cuenca, E.M.; Alvarez-Coque, M.C.G. Determination of the protein and free amino acid content in a sample using o-phthalaldehyde and N-acetyl-L-cysteine. Analyst 1990, 115, 1125–1128. [Google Scholar] [CrossRef] [PubMed]
- Spellman, D.; McEvoy, E.; O’Cuinn, G.; Fitzgerald, R. Proteinase and exopeptidase hydrolysis of whey protein: Comparison of the TNBS, OPA and pH stat methods for quantification of degree of hydrolysis. Int. Dairy J. 2003, 13, 447–453. [Google Scholar] [CrossRef]
- Gerardo, M.L.; Oatley-Radcliffe, D.L.; Lovitt, R.W. Integration of membrane technology in microalgae biorefineries Integration of membrane technology in microalgae biore fi neries. J. Membr. Sci. 2015, 464, 86–99. [Google Scholar] [CrossRef]
- Hwang, K.-J.; Liao, C.-Y.; Tung, K.-L. Effect of membrane pore size on the particle fouling in membrane filtration. Desalination 2008, 234, 16–23. [Google Scholar] [CrossRef]
- Siddiqui, M.U.; Arif, A.F.M.; Bashmal, S. Permeability-Selectivity Analysis of Microfiltration and Ultrafiltration Membranes: Effect of Pore Size and Shape Distribution and Membrane Stretching. Membranes 2016, 6, 40. [Google Scholar] [CrossRef] [Green Version]
- Hughes, D.; Field, R. Crossflow filtration of washed and unwashed yeast suspensions at constant shear under nominally sub-critical conditions. J. Membr. Sci. 2006, 280, 89–98. [Google Scholar] [CrossRef]
- D’souza, N.; Wiley, D.E. Whey ultrafiltration: Effect of operating parameters on flux and rejection. In Proceedings of the 5th International Membrane Science and Technology Conference, Sydney, Australia, 10–14 November 2003; pp. 1–6. [Google Scholar]
- Tanaka, T.; Yamagiwa, Y.; Nagano, T.; Taniguchi, M.; Nakanishi, K. Relationship between Cake Structure and Membrane Pore Size in Crossflow Filtration of Microbial Cell Suspension Containing Fine Particles. J. Chem. Eng. Jpn. 2001, 34, 1524–1531. [Google Scholar] [CrossRef]
- Tanaka, T.; Usui, K. Formation of the Gel Layer of Polymers and Its Effect on the Permeation Flux in Crossflow Filtration ofCorynebacterium glutamicumBroth. Sep. Sci. Technol. 2008, 33, 707–722. [Google Scholar] [CrossRef]
- Sofuwani, Z.N.; Aslina, H.S.; Mazlina, M.S. Separation of lactose from raw goat’s milk by cross-flow hollow fiber ultrafiltration membrane. Int. Food Res. J. 2016, 23, 209–219. [Google Scholar]
- Taddei, C.; Aimar, P.; Daufin, G.; Sanchez, V. Factors affecting fouling of an inorganic membrane during sweet whey ultrafiltration. Le Lait 1988, 68, 157–176. [Google Scholar] [CrossRef] [Green Version]
- Piry, A.; Heino, A.; Kühnl, W.; Grein, T.; Ripperger, S.; Kulozik, U. Effect of membrane length, membrane resistance, and filtration conditions on the fractionation of milk proteins by microfiltration. J. Dairy Sci. 2012, 95, 1590–1602. [Google Scholar] [CrossRef] [PubMed]
- Carić, M.Đ.; Milanović, S.D.; Krstić, D.M.; Tekic, M.N. Fouling of inorganic membranes by adsorption of whey proteins. J. Membr. Sci. 2000, 165, 83–88. [Google Scholar] [CrossRef]
- Bacchin, P.; Aimar, P.; Field, R. Critical and sustainable fluxes: Theory, experiments and applications. J. Membr. Sci. 2006, 281, 42–69. [Google Scholar] [CrossRef] [Green Version]
- Suwal, S. Fractionation of Peptides from Protein Hydrolysate by Electrodialysis with Filtration Membrane: Process Optimization, Fouling Characterization and Control. Ph.D. Thesis, Laval University, Québec, QC, Canada, 2015. [Google Scholar]
- Cassano, A. Recovery Technologies for Water-Soluble Bioactives: Advances in Membrane-Based Processes. In Engineering Foods for Bioactives Stability and Delivery. Food Engineering Series; Roos, Y.H., Livney, Y.D., Eds.; Springer: New York, NY, USA, 2017; pp. 51–83. [Google Scholar]
- Chabeaud, A.; Vandanjon, L.; Bourseau, P.; Jaouen, P.; Chaplain-Derouiniot, M.; Guérard, F. Performances of ultrafiltration membranes for fractionating a fish protein hydrolysate: Application to the refining of bioactive peptidic fractions. Sep. Purif. Technol. 2009, 66, 463–471. [Google Scholar] [CrossRef]
- Das, R.; Bhattacherjee, C.; Ghosh, S. Effects of operating parameters and nature of fouling behavior in ultrafiltration of sesame protein hydrolysate. Desalination 2009, 237, 268–276. [Google Scholar] [CrossRef]
- Saidi, S.; Deratani, A.; Ben Amar, R.; Belleville, M.-P. Fractionation of a tuna dark muscle hydrolysate by a two-step membrane process. Sep. Purif. Technol. 2013, 108, 28–36. [Google Scholar] [CrossRef]
- Pouliot, Y.; Wijers, M.; Gauthier, S.; Nadeau, L. Fractionation of whey protein hydrolysates using charged UF/NF membranes. J. Membr. Sci. 1999, 158, 105–114. [Google Scholar] [CrossRef]
- Lim, Y.P.; Mohammad, A.W. Influence of pH and Ionic Strength during Food Protein Ultrafiltration: Elucidation of Permeate Flux Behavior, Fouling Resistance, and Mechanism. Sep. Sci. Technol. 2012, 47, 446–454. [Google Scholar] [CrossRef]
- Andrade, L.H.; Pires, W.L.; Grossi, L.B.; Aguiar, A.O.; Amaral, M.C. Integration of two-stage nanofiltration with arsenic and calcium intermediate chemical precipitation for gold mining effluent treatment. Environ. Technol. 2019, 40, 1644–1656. [Google Scholar] [CrossRef]
- Robertson, B.C.; Zydney, A.L. Protein adsorption in asymmetric ultrafiltration membranes with highly constricted pores. J. Colloid Interface Sci. 1990, 134, 563–575. [Google Scholar] [CrossRef]
- Fouzia, Y.; Abdelouahab, N.; Amal, K.; Slimane, B. Whey Ultrafiltration: Effect of pH on Permeate Flux and Proteins Retention. World Appl. Sci. J. 2015, 33, 744–751. [Google Scholar]
- Nanda, D.; Tung, K.-L.; Li, Y.-L.; Lin, N.-J.; Chuang, C.-J. Effect of pH on membrane morphology, fouling potential, and filtration performance of nanofiltration membrane for water softening. J. Membr. Sci. 2010, 349, 411–420. [Google Scholar] [CrossRef]
- Muro, C.; Riera, F.A.; Fernandez, A. Advancements in the Fractionation of Milk Biopeptides by Means of Membrane Processes. In Bioactive Food Peptides in Health and Disease; IntechOpen: London, UK, 2013; pp. 241–266. [Google Scholar]
- Collins, L.A.; Mirza, S.P.; Kissebah, A.H.; Olivier, M. Integrated approach for the comprehensive characterization of lipoproteins from human plasma using FPLC and nano-HPLC-tandem mass spectrometry. Physiol. Genom. 2010, 40, 208–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turgeon, S.L.; Gauthier, S.F. Whey Peptide Fractions Obtained with a Two-Step Ultrafiltration Process: Production and Characterization. J. Food Sci. 1990, 55, 106–110. [Google Scholar] [CrossRef]
- Butylina, S.; Luque, S.; Nyström, M. Fractionation of whey-derived peptides using a combination of ultrafiltration and nanofiltration. J. Membr. Sci. 2006, 280, 418–426. [Google Scholar] [CrossRef]
- Brule, G.; Roger, L.; Fauquant, J.; Piot, M. Phosphopeptides from casein-based raw material. United States Patent 4980450, 25 December 1990. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide Compositions (%) | |||
|---|---|---|---|
| Membrane configurations | 10 | 5 | Two-stage 10/5 kDa |
| Peptide fraction (1700–600 Da) | |||
| Peak 1 (1672.92 Da) | ND | 2.25 | 0.73 |
| Peak 2 (1672.92 Da) | 1.68 | 4.82 | 1.80 |
| Peptide fraction (600–300 Da) | |||
| Peak 3 | 19.73 | 12.24 | 18.22 |
| Peak 4 | 3.14 | 5.62 | 1.76 |
| Peptide fraction (<300 Da) | |||
| Peak 5 | 25.42 | 26.47 | 30.55 |
| Peak 6 | 2.63 | 26.98 | 16.05 |
| Peak 7 | ND | ND | 10.64 |
| peak 8 | 25.90 | ND | 2.40 |
| peak 9 | 21.49 | 21.62 | 17.86 |
| Total (1700–600 Da) | 1.68 | 7.07 | 2.53 |
| Total (600–300 Da) | 22.87 | 17.86 | 19.98 |
| Total (<300 Da) | 75.45 | 75.07 | 77.49 |
| Enrichment of Peptide Using Two-Stage Membrane (%) | ||
|---|---|---|
| Membrane configurations | 10 kDa vs. Two-stage 10/5 kDa | 5 kDa vs. Two-stage 10/5 kDa |
| Peptide fraction (1700–600 Da) | ||
| Peak 1 (1672.92 Da) | - | −67.56 |
| Peak 2 (1672.92 Da) | 7.14 | −62.66 |
| Peptide fraction (600–300 Da) | ||
| Peak 3 | −7.65 | 48.86 |
| Peak 4 | −43.95 | −68.68 |
| Peptide fraction (<300 Da) | ||
| Peak 5 | 20.18 | 15.41 |
| Peak 6 | 510.27 | −40.51 |
| Peak 7 | - | - |
| peak 8 | −90.73 | - |
| peak 9 | −16.89 | −17.39 |
| Total (1700–600 Da) | 50.60 | −64.21 |
| Total (600–300 Da) | −12.64 | 11.87 |
| Total (<300 Da) | 2.68 | 3.22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Md Saleh, N.I.; Wan Ab Karim Ghani, W.A.; Mustapa Kamal, S.M.; Harun, R. Performance of Single and Two-Stage Cross-Flow Ultrafiltration Membrane in Fractionation of Peptide from Microalgae Protein Hydrolysate (Nannochloropsis gaditana). Processes 2021, 9, 610. https://doi.org/10.3390/pr9040610
Md Saleh NI, Wan Ab Karim Ghani WA, Mustapa Kamal SM, Harun R. Performance of Single and Two-Stage Cross-Flow Ultrafiltration Membrane in Fractionation of Peptide from Microalgae Protein Hydrolysate (Nannochloropsis gaditana). Processes. 2021; 9(4):610. https://doi.org/10.3390/pr9040610
Chicago/Turabian StyleMd Saleh, Nur Izzati, Wan Azlina Wan Ab Karim Ghani, Siti Mazlina Mustapa Kamal, and Razif Harun. 2021. "Performance of Single and Two-Stage Cross-Flow Ultrafiltration Membrane in Fractionation of Peptide from Microalgae Protein Hydrolysate (Nannochloropsis gaditana)" Processes 9, no. 4: 610. https://doi.org/10.3390/pr9040610
APA StyleMd Saleh, N. I., Wan Ab Karim Ghani, W. A., Mustapa Kamal, S. M., & Harun, R. (2021). Performance of Single and Two-Stage Cross-Flow Ultrafiltration Membrane in Fractionation of Peptide from Microalgae Protein Hydrolysate (Nannochloropsis gaditana). Processes, 9(4), 610. https://doi.org/10.3390/pr9040610