
Carotenoid Extraction from Locally and Organically Produced Cereals Using Saponification Method



Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Samples
2.3. Extraction by Saponification
2.4. HPLC Analysis
2.5. Data Analysis
3. Results
3.1. Grain Carotenoid Content in Locally and Organically Produced Cereals and Genotype X Environment Effects
3.2. Percentage of Explanation for Variation in Carotenoids Content
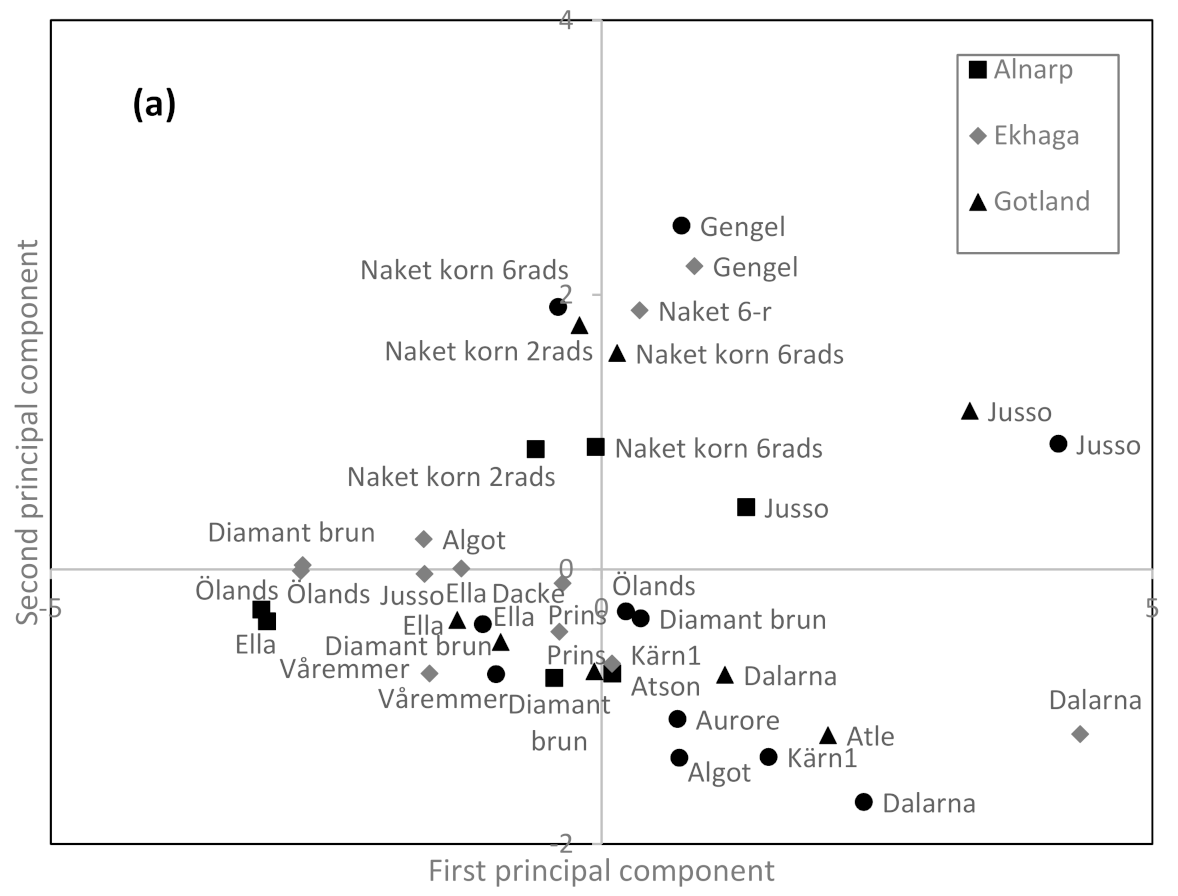
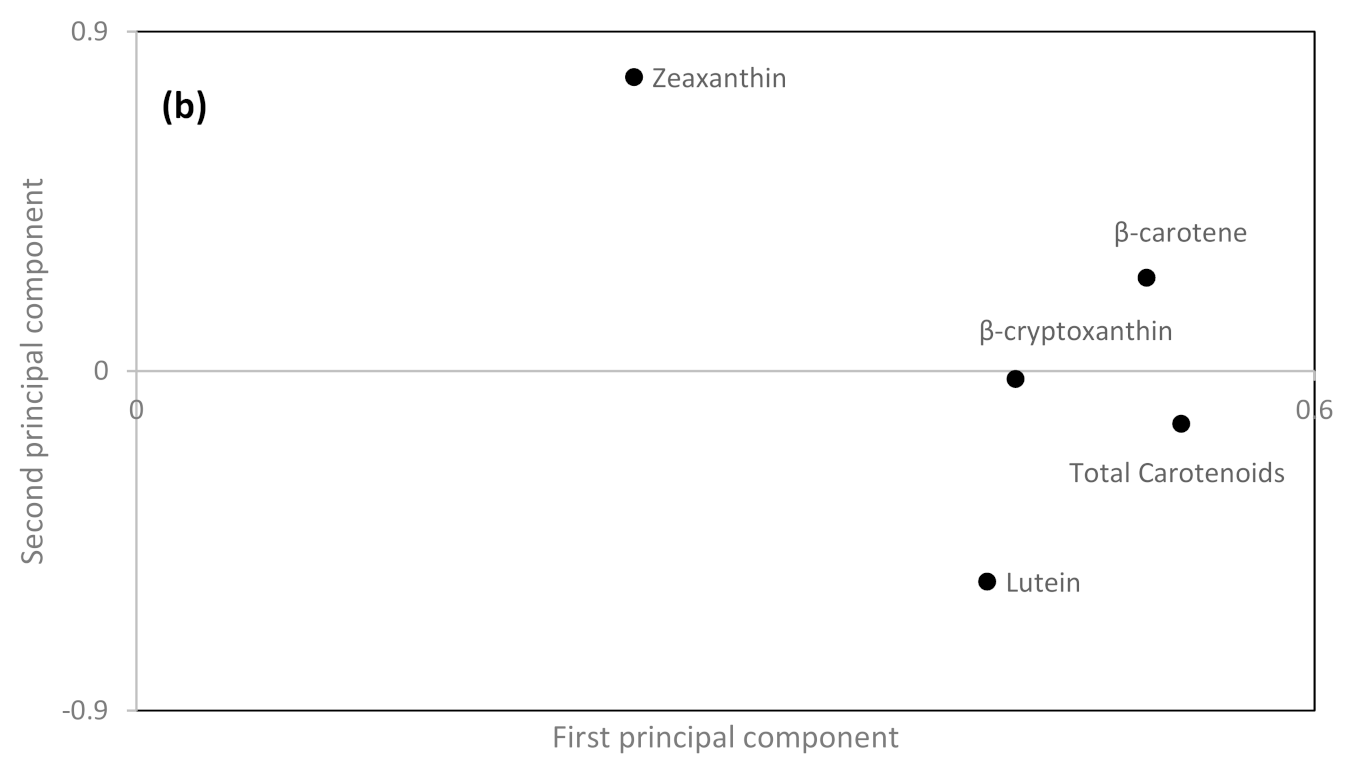
3.3. Relationship between Genotype, Environment and Contents of Different Carotenoids
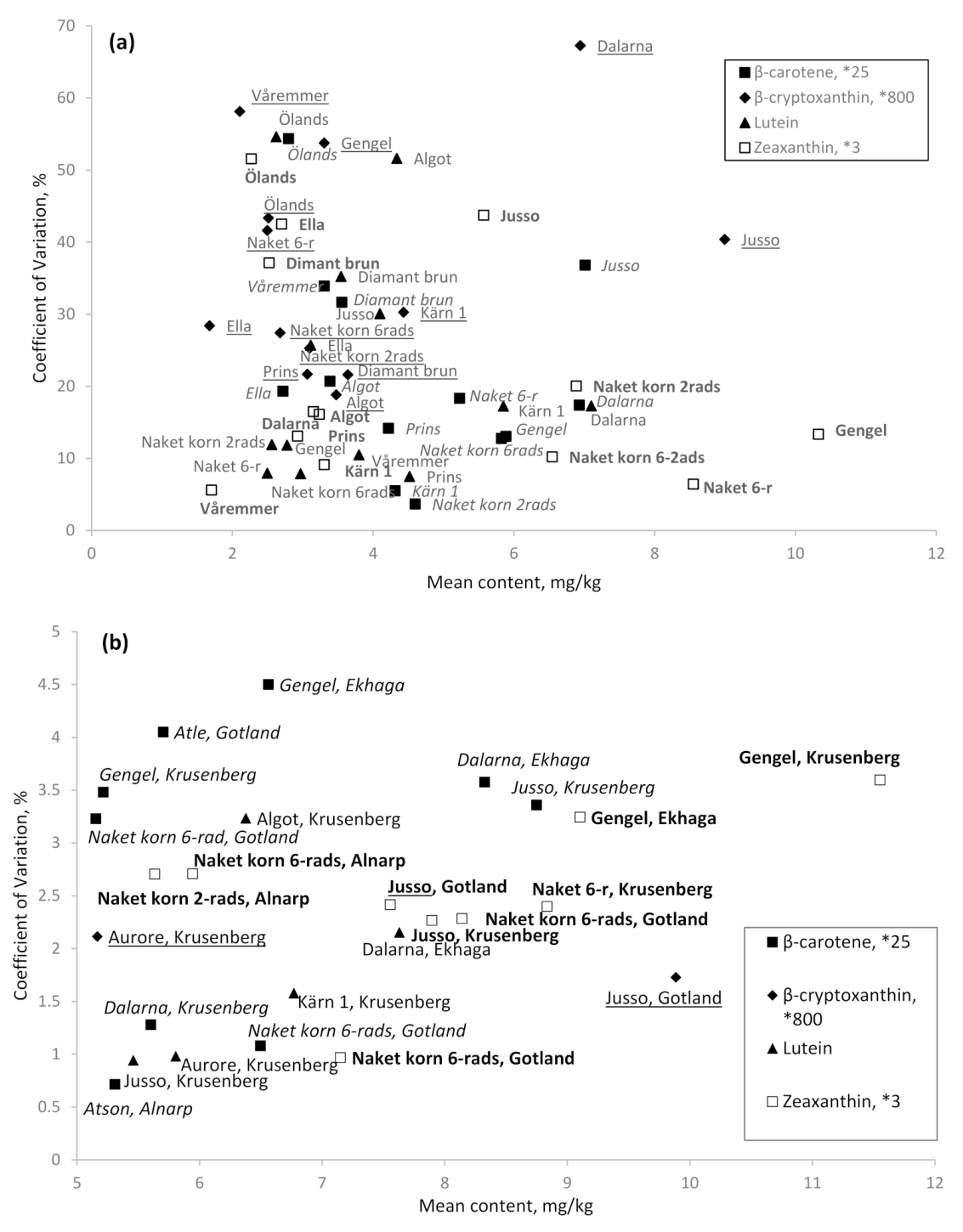
3.4. Stability Over Environments
3.5. High Content and Stability per Cultivar within an Environment
3.6. Correlation among Carotenoids and Plant Characters
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shiferaw, B.; Smale, M.; Braun, H.-J.; Duveiller, E.; Reynolds, M.; Muricho, G. Crops that feed the world 10. Past successes and future challenges to the role played by wheat in global food security. Food Secur. 2013, 5, 291–317. [Google Scholar] [CrossRef] [Green Version]
- FAO Food and Agriculture Organization of the United Nations. FAOSTAT Database; FAO: Rome, Italy, 2019. [Google Scholar]
- Pena-Bautista, R.J.; Hernandez-Espinosa, N.; Jones, J.M.; Guzman, C.; Braun, H.J. Wheat-based foods: Their global and regional importance in the food supply, nutrition and health. Cereal Foods World 2017, 62, 231–249. [Google Scholar] [CrossRef]
- Save and Grow in Practice: Maize Rice Wheat. A Guide to Sustainable Cereal Production. In Food and Agriculture Organization of the United Nations (FAO); FAO: Rome, Italy, 2016; ISBN ISBN 978-92-5-108519-6.
- Andersen, V.; Bar, E.; Wirtanen, G. Nutritional and Health Aspects of Food in Nordic Countries; Academic Press: Berlin/Heidelberg, Germany, 2018; ISBN 978-0-12-809416-7. [Google Scholar]
- Pinheiro, M.A.A.; Bebeli, P.J.; Bettencourt, E.; Costa, G.; Dias, S.; Dos Santos, T.M.M.; Slaski, J.J. Cereal landraces genetic resources in worldwide GeneBanks. A review. Agron. Sust. Dev. 2013, 33, 177–203. [Google Scholar]
- Schmidt, S.B.; George, T.S.; Brown, L.K.; Booth, A.; Wishart, J.; Hedley, P.E.; Martin, P.; Russell, J.; Husted, S. Ancient barley landraces adapted to marginal soils demonstrate exceptional tolerance to manganese limitation. Ann. Bot. 2019, 123, 831–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hussain, A.; Larsson, H.; Kuktaite, R.; Johansson, E. Mineral composition of organically grown wheat genotypes: Contribution to daily minerals intake. Int. J. Environ. Res. Public Health 2010, 7, 3442–3456. [Google Scholar] [CrossRef] [PubMed]
- Moreira-Ascarrunz, S.G.; Larsson, H.; Prieto-Linde, M.L.; Johansson, E. Mineral nutritional yield and nutrient density of locally adapted wheat genotypes under organic production. Foods 2016, 5, 89. [Google Scholar] [CrossRef] [Green Version]
- Shafie, F.A.; Rennie, D. Consumer perceptions towards organic food. Procedia Soc. Behav. Sci. 2012, 49, 360–367. [Google Scholar] [CrossRef] [Green Version]
- Johansson, E.; Hussain, A.; Kuktaite, R.; Andersson, S.C.; Olsson, M.E. Contribution of organically grown crops to human health. Int. J. Environ. Res. Public Health 2014, 11, 3870–3893. [Google Scholar] [CrossRef] [PubMed]
- Underwood, T.; McCullum-Gomez, C.; Harmon, A.; Roberts, S. Organic agriculture supports biodiversity and sustainable food production. J. Hunger Environ. Nutr. 2011, 6, 398–423. [Google Scholar] [CrossRef]
- Liu, R.H. Whole grain phytochemicals and health. J. Cereal Sci. 2007, 46, 2017–2219. [Google Scholar] [CrossRef]
- Ndolo, V.U.; Beta, T. Distribution of carotenoids in endosperm, germ and aleurone fractions of cereal grain kernels. Food Chem. 2013, 139, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Mellado-Ortega, E.; Hornero-Méndez, D. Carotenoids in cereals: An ancient resource with present and future applications. Phytochem. Rev. 2015, 14, 873–890. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; Larsson, H.; Kuktaite, R.; Olsson, M.E.; Johansson, E. Carotenoid content in organically produced wheat: Relevance for human nutritional health on consumption. Int. J. Environ. Res. Pub. Health 2015, 12, 14068–14083. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, K.; Yin, J.J.; Yu, L. Phenolic acid, tocopherol and carotenoid compositions, and antioxidant functions of hard red winter wheat bran. J. Agric. Food Chem. 2005, 53, 3916–3922. [Google Scholar] [CrossRef]
- Moore, J.; Hao, Z.; Zhou, K.; Luther, M.; Costa, J.; Yu, L. Carotenoid, tocopherol, phenolic acid and antioxidant properties of Maryland-grown soft wheat. J. Agric. Food Chem. 2005, 53, 6649–6657. [Google Scholar] [CrossRef] [PubMed]
- Leenhardt, F.; Lyan, B.; Rock, E.; Boussard, A.; Potus, J.; Chanliaud, E.; Remesy, C. Genetic variability of carotenoid concentration, and lipoxygenase and perioxidase activities among cultivated wheat species and bread wheat varieties. Eur. J. Agron. 2006, 25, 170–176. [Google Scholar] [CrossRef]
- Konopka, I.; Czaplicki, S.; Rotkiewicz, D. Differences in content and composition of free lipids and carotenoids in flour of spring and winter wheat cultivated in Poland. Food Chem. 2006, 95, 290–300. [Google Scholar] [CrossRef]
- Granado, F.; Olmedilla, B.; Blanco, I. Nutritional and clinical relevance of lutein in human health. Br. J. Nutr. 2003, 90, 487–502. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Lin, X.-M. Effects of lutein and zeaxanthin on aspects of eye health. J. Sci. Food Agric. 2010, 90, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Keum, Y.S. Carotenoid extraction methods: A review of recent developments. Food Chem. 2018, 240, 90–103. [Google Scholar] [CrossRef]
- Kopec, R.E.; Cooperstone, J.L.; Cichon, M.J.; Schwartz, S.J. Analysis Methods of Carotenoids. In Analysis of Antioxidant-Rich Phytochemicals; Wiley-Blackwell: Oxford, UK, 2012; pp. 105–148. ISBN 9780813823911. [Google Scholar]
- Granado, F.; Olmedilla, B.; Gil-Martinez, E.; Blanco, I. A Fast, Reliable and Low-cost Saponification Protocol for Analysis of Carotenoids in Vegetables. J. Food Comp. Anal. 2001, 14, 479–489. [Google Scholar] [CrossRef]
- Abdel-Aal, E.-S.M.; Young, J.C.; Rabalski, I.; Hucl, P.; Frgeau-Reid, J. Identification and quantification of seed carotenois in selected wheat species. J. Agric. Food Chem. 2007, 55, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, A.; Brandolini, A.; Pompei, C.; Piscozzi, R. Carotenoids and tocols of einkorn wheat (Triticum monococcum ssp. monococcum L.). J. Cereal Sci. 2006, 44, 182–193. [Google Scholar] [CrossRef]
- Paznocht, L.; Kotíková, Z.; Sulc, M.; Lachman, J.; Orsák, M.; Eliásová, M.; Martinek, P. Free and esterified carotenoids in pigmented wheat. Tritordeum and barley grains. Food Chem. 2018, 240, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Yerke, A.; Sang, S. Oats Whole Grains and Their Bioactives: Composition and Health; Johnson, J., Wallace, T., Eds.; John Wiley & Sons Ltd.: Hoboken, NJ, USA, 2019. [Google Scholar]
- Adams, D.C.; Salois, M.J. Local versus organic: A consumer preferences and willingness-to-pay. Renew. Agric. Food Syst. 2010, 25, 331–341. [Google Scholar] [CrossRef] [Green Version]
- Klopcic, M.; Verhees, F.J.H.M.; Kuipers, A.; Kos-Skubic, M. Consumer preceptions of home made, organic, EU certified, and traditional local products in Slovenia. In Consumer Attitudes to Food Quality Products; Klopcic, M., Kuipers, A., Hocquette, J.F., Eds.; EAAP—European Federation of Animal Science; Wageningen Academic Press: Wageningen, The Netherlands, 2013; Volume 133. [Google Scholar]
- Fratianni, A.; Caboni, M.F.; Irano, M.; Panfili, G.A. Critical comparison between traditional methods and supercritical carbon dioxide extraction for the determination of tocochromanols in cereals. Eur. Food Res. Technol. 2002, 215, 353–358. [Google Scholar] [CrossRef]
- Hussain, A.; Larsson, H.; Olsson, M.E.; Kuktaite, R.; Grausgruber, H.; Johansson, E. Is organically produced wheat a source of tocopherols and tocotrienols for health food? Food Chem. 2012, 132, 1789–1795. [Google Scholar] [CrossRef]
- Panfili, G.; Fratianni, A.; Irano, M. Improved normal-phase high-performance liquid chromatography procedure for the determination of carotenoids in cereals. J. Agric. Food Chem. 2004, 52, 6373–6377. [Google Scholar] [CrossRef]
- Malik, A.H.; Kuktaite, R.; Johansson, E. Combined effect of genetic and environmental factors on the accumulation of proteins in the wheat grain and their relationship to bread-making quality. J. Cereal Sci. 2013, 57, 170–174. [Google Scholar] [CrossRef]
- Vazquez, D.; Berger, A.; Prieto-Linde, M.L.; Johansson, E. Can nitrogen fertilization be used to modulate yield, protein content and bread-making quality in Uruguayan wheat? J. Cereal Sci. 2019, 85, 153–161. [Google Scholar] [CrossRef]
- Mukamuhirwa, A.; Persson Hovmalm, H.; Ortiz, R.; Nyamangyoku, O.; Prieto-Linde, M.L.; Ekholm, A.; Johansson, E. Effect of intermittent drought on grain yield and quality of rice (Oryza sativa L.) grown in Rwanda. J. Agron. Crop Sci. 2020, 206, 252–262. [Google Scholar] [CrossRef]
- Ross, S.M. Introductory Statistics; Elsevier Inc.: Oxford, UK, 2010. [Google Scholar]
- Okarter, N.; Liu, C.S.; Sorrells, M.E.; Liu, R.H. Phytochemical content and antioxidant activity of six diverse varieties of whole wheat. Food Chem. 2010, 119, 249–257. [Google Scholar] [CrossRef]
- Adom, K.K.; Sorrells, M.E.; Liu, R.H. Phytochemicals and antioxidant activity of wheat varieties. J. Agric. Food Chem. 2003, 51, 7825–7834. [Google Scholar] [CrossRef] [PubMed]
- Lachman, J.; Hejtmánková, K.; Kotíková, Z. Tocols and carotenoids of einkorn, emmer and spring wheat varieties: Selection for breeding and production. J. Cereal Sci. 2013, 57, 207–214. [Google Scholar] [CrossRef]
- Fratianni, A.; Giuzio, L.; Di Criscio, T.; Zina, F.; Panfili, G. Response of carotenoids and tocols of durum wheat in relation to water stress and sulfur fertilization. Food Chem. 2013, 61, 2583–2590. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, S. Wide adaptation: How wide? Euphytica 1989, 40, 197–205. [Google Scholar]
- Wolfe, M.S.; Baresel, J.P.; Desclaux, D.; Goldringer, I.; Hoad, S.; Kovacs, G.; Löschenberger, F.; Miedaner, T.; Østergård, H.; Van Bueren, E.L. Developments inbreeding cereals for organic agriculture. Euphytica 2008, 163, 323–346. [Google Scholar] [CrossRef] [Green Version]
- Van Bueren, E.L.; Jones, S.S.; Tamm, L.; Murphy, K.M.; Myers, J.R.; Leifert, C.; Messmer, M.M. The need to breed crop varieties suitable for organic farming using wheat, tomato and broccoli as examples: A review. Njas–Wagen J. Life Sci. 2010. [Google Scholar] [CrossRef]
- Johansson, E.; Prieto-Linde, M.L.; Larsson, H. Locally adapted and organically grown landrace and ancient spring cereals—A unique source of minerals in the human diet. Foods 2021, 10, 393. [Google Scholar] [CrossRef]
- Rao, A.V.; Rao, L.G. Carotenoids and human health. Pharmacol. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef]
- Zile, M.H. Vitamin A and embryonic development: An overview. J. Nutr. 1998, 128, 455s–458s. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, S.C.; Olsson, M.E.; Johansson, E.; Rumpunen, K. Carotenoids in Sea Buckthorn (Hippophae rhamnoides L.) Berries during Ripening and Use of Pheophytin a as a Maturity Marker. J. Agric. Food Chem. 2009, 57, 250–258. [Google Scholar] [CrossRef] [PubMed]
- Saini, R.K.; Nile, S.H.; Park, S.W. Carotenoids from fruits and vegetables: Chemistry, analysis, occurrence, bioavailability and biological activities. Food Res. Int. 2015, 76, 735–750. [Google Scholar] [CrossRef] [PubMed]
- Osganian, S.K.; Stampfer, M.J.; Rimm, E.; Spiegelman, D.; Manson, J.E.; Willett, W.C. Dietary carotenoids and risk of coronary artery disease in women. Am. J. Clin. Nutr. 2003, 77, 1390–1399. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Name | Type | Cereal Type | Origin Year Released and NGB Number |
|---|---|---|---|
| Algot | Spring wheat | Old cultivar | 1953, NGB11611 |
| Atle | Spring wheat | Old cultivar | 1936, NGB7455 |
| Atson | Spring wheat | Old cultivar | 1954, NGB7460 |
| Aurore | Spring wheat | Old cultivar | 1929, NGB9690 |
| Dacke | Spring wheat | Old cultivar | 1990, NGB9955 |
| Dalarna | Spring wheat | Landrace | NGB6673, selection |
| Diamant brun | Spring wheat | Old cultivar | 1938, NGB6681, selection |
| Ella | Spring wheat | Old cultivar | 1950, NGB6683 |
| Gengel | Naked 2-row barley | Landrace Czech republic | From Czech growers |
| Jusso | Spring rye | Finnish landrace | From finish growers |
| Kärn 1 | Spring wheat | Old cultivar | 1947, NGB7458 |
| Naket 6-r | Naked 6-row barley | Selection | Diverse origin from Gene bank |
| Naket korn 2rads | Naked 2-row barley | Selection | Diverse origin from Gene bank |
| Naket korn 6rads | Naked 6-row barley | Selection | Diverse origin from Gene bank |
| Ölands | Spring wheat | Landrace | NGB 4798, selection |
| Prins | Spring wheat | Old cultivar | 1962, NGB6688 |
| Våremmer | Spring wheat | Primitive | NGB4499, selection as spring wheat |
| Locality and Cultivar | β-Carotene (10−1) | β-Cryptoxanthin (10−3) | Lutein | Zea-Xanthin | Total Carotenoids |
|---|---|---|---|---|---|
| Ekhaga | |||||
| Algot | 1.13 ± 0.20 | 4.81 ± 0.72 | 2.30 ± 0.06 | 1.12 ± 0.25 | 3.53 ± 0.30 |
| Dacke | 1.94 ± 0.01 | 7.01 ± 0.40 | 3.27 ± 0.21 | 0.90 ± 0.01 | 4.37 ± 0.20 |
| Dalarna | 3.33 ± 0.12 | 16.4 ± 0.91 | 7.63 ± 0.16 | 1.19 ± 0.01 | 9.17 ± 0.16 |
| Diamant brun | 7.93 ± 0.04 | 3.06 ± 0.11 | 1.59 ± 0.04 | 0.67 ± 0.02 | 2.35 ± 0.06 |
| Ella | 1.14 ± 0.03 | 2.80 ± 0.27 | 3.21 ± 0.04 | 1.28 ± 0.03 | 4.61 ± 0.06 |
| Gengel | 2.62 ± 0.12 | 6.14 ± 0.54 | 2.49 ± 0.11 | 3.03 ± 0.10 | 5.79 ± 0.21 |
| Jusso | 1.40 ± 0.22 | 4.79 ± 0.23 | 2.36 ± 0.01 | 0.73 ± 0.06 | 3.24 ± 0.09 |
| Kärn 1 | 1.64 ± 0.03 | 4.32 ± 0.47 | 4.93 ± 0.05 | 1.09 ± 0.02 | 6.19 ± 0.03 |
| Naket 6-r | 2.42 ± 0.22 | 4.27 ± 0.54 | 2.58 ± 0.14 | 2.76 ± 0.23 | 5.58 ± 0.37 |
| Ölands | 0.76 ± 0.02 | 2.91 ± 0.01 | 1.66 ± 0.01 | 0.66 ± 0.01 | 2.40 ± 0.02 |
| Prins | 1.48 ± 0.10 | 4.57 ± 0.01 | 4.21 ± 0.03 | 1.09 ± 0.01 | 5.45 ± 0.03 |
| Våremmer | 0.92 ± 0.05 | 4.02 ± 0.26 | 3.45 ± 0.13 | 0.57 ± 0.04 | 4.12 ± 0.16 |
| Krusenberg | |||||
| Algot | 1.58 ± 0.07 | 3.88 ± 0.72 | 6.38 ± 0.21 | 0.98 ± 0.01 | 7.52 ± 0.22 |
| Aurore | 1.42 ± 0.04 | 6.46 ± 0.14 | 5.80 ± 0.06 | 1.13 ± 0.03 | 7.08 ± 0.08 |
| Dalarna | 2.24 ± 0.03 | 4.94 ± 0.60 | 8.14 ± 0.44 | 1.19 ± 0.05 | 9.56 ± 0.46 |
| Diamant brun | 1.84 ± 0.12 | 4.83 ± 0.22 | 4.77 ± 0.04 | 1.34 ± 0.01 | 6.29 ± 0.06 |
| Ella | 1.19 ± 0.02 | 2.03 ± 0.01 | 3.89 ± 0.12 | 1.09 ± 0.03 | 5.10 ± 0.16 |
| Gengel | 2.09 ± 0.07 | 2.13 ± 0.01 | 3.06 ± 0.12 | 3.85 ± 0.14 | 7.12 ± 0.23 |
| Jusso | 3.50 ± 0.12 | 16.6 ± 2.29 | 5.46 ± 0.05 | 2.63 ± 0.06 | 8.46 ± 0.12 |
| Kärn 1 | 1.81 ± 0.02 | 6.75 ± 1.55 | 6.77 ± 0.11 | 1.11 ± 0.16 | 9.07 ± 0.06 |
| Naket 6-r | 1.77 ± 0.02 | 1.98 ± 0.01 | 2.41 ± 0.24 | 2.95 ± 0.07 | 5.54 ± 0.31 |
| Ölands | 1.93 ± 0.07 | 4.74 ± 0.82 | 4.52 ± 0.24 | 1.24 ± 0.06 | 5.97 ± 0.31 |
| Våremmer | 1.73 ± 0.07 | 1.24 ± 0.01 | 4.14 ± 0.14 | 0.56 ± 0.03 | 4.88 ± 0.18 |
| Alnarp | |||||
| Atson | 2.12 ± 0.02 | 4.86 ± 0.36 | 4.62 ± 0.01 | 0.67 ± 0.01 | 5.50 ± 0.01 |
| Diamant brun | 1.78 ± 0.04 | 5.06 ± 0.28 | 4.18 ± 0.12 | 0.55 ± 0.01 | 4.91 ± 0.13 |
| Ella | 0.75 ± 0.00 | 1.29 ± 0.01 | 1.86 ± 0.01 | 1.09 ± 0.03 | 2.24 ± 0.01 |
| Jusso | 2.64 ± 0.19 | 11.3 ± 1.15 | 3.74 ± 0.16 | 1.55 ± 0.17 | 5.57 ± 0.16 |
| Naket korn 2rads | 1.84 ± 0.10 | 3.03 ± 0.19 | 2.84 ± 0.06 | 1.88 ± 0.05 | 4.90 ± 0.12 |
| Naket korn 6rads | 2.06 ± 0.07 | 4.16 ± 0.01 | 3.18 ± 0.06 | 1.98 ± 0.05 | 5.37 ± 0.12 |
| Ölands | 0.68 ± 0.06 | 1.78 ± 0.01 | 1.69 ± 0.13 | 0.36 ± 0.03 | 2.12 ± 0.17 |
| Gotland | |||||
| Atle | 2.28 ± 0.09 | 5.51 ± 00.33 | 7.21 ± 0.52 | 1.32 ± 0.07 | 8.76 ± 0.61 |
| Dalarna | 2.74 ± 0.15 | 4.68 ± 0.58 | 5.53 ± 0.32 | 0.85 ± 0.04 | 6.66 ± 0.37 |
| Diamant brun | 1.27 ± 0.12 | 5.25 ± 0.79 | 3.64 ± 0.03 | 0.81 ± 0.01 | 4.58 ± 0.04 |
| Ella | 1.26 ± 0.07 | 2.25 ± 0.32 | 3.50 ± 0.03 | 0.92 ± 0.01 | 4.55 ± 0.04 |
| Jusso | 3.68 ± 0.96 | 12.4 ± 0.21 | 4.83 ± 0.12 | 2.52 ± 0.06 | 7.73 ± 0.09 |
| Naket korn 2rads | 1.84 ± 0.02 | 4.71 ± 0.51 | 2.28 ± 0.03 | 2.71 ± 0.06 | 5.19 ± 0.09 |
| Naket korn 6rads | 2.60 ± 0.03 | 2.54 ± 0.39 | 2.76 ± 0.06 | 2.38 ± 0.02 | 5.41 ± 0.05 |
| Prins | 1.90 ± 0.02 | 3.09 ± 0.28 | 4.83 ± 0.05 | 0.86 ± 0.04 | 5.88 ± 0.06 |
| β-Carotene | β-Cryptoxanthin | Lutein | Zeaxanthin | Total Carotenoids | |
|---|---|---|---|---|---|
| C | 62.8 | 32.3 | 48.1 | 68.6 | 37.4 |
| L | 8.15 | 0.73 | 20.2 | 6.85 | 28.8 |
| CL | 92.6 | 66.1 | 92.4 | 83.5 | 93.5 |
| β-Cryptoxanthin | β-Carotene | Lutein | Zea-Xanthin | Yield (kg/Ha) | TKV (g) | Protein Content (%) | |
|---|---|---|---|---|---|---|---|
| β-carotene | 0.72 *** | ||||||
| Lutein | 0.47 *** | 0.48 ** | |||||
| Zea-xanthin | 0.20 | 0.55 *** | −0.12 | ||||
| Yield (kg/Ha) | 0.14 | 0.09 | 0.06 | −0.16 | |||
| TKV (g) | −0.14 | −0.16 | −0.14 | 0.09 | −0.27 | ||
| Protein Content (%) | −0.17 | −0.09 | −0.38 | 0.06 | −0.11 | 0.29 | |
| Straw Length (cm) | 0.31 | 0.11 | 0.50 ** | −0.28 | 0.24 | −0.17 | −0.69 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, A.; Larsson, H.; Johansson, E. Carotenoid Extraction from Locally and Organically Produced Cereals Using Saponification Method. Processes 2021, 9, 783. https://doi.org/10.3390/pr9050783
Hussain A, Larsson H, Johansson E. Carotenoid Extraction from Locally and Organically Produced Cereals Using Saponification Method. Processes. 2021; 9(5):783. https://doi.org/10.3390/pr9050783
Chicago/Turabian StyleHussain, Abrar, Hans Larsson, and Eva Johansson. 2021. "Carotenoid Extraction from Locally and Organically Produced Cereals Using Saponification Method" Processes 9, no. 5: 783. https://doi.org/10.3390/pr9050783
APA StyleHussain, A., Larsson, H., & Johansson, E. (2021). Carotenoid Extraction from Locally and Organically Produced Cereals Using Saponification Method. Processes, 9(5), 783. https://doi.org/10.3390/pr9050783