
Beauveria bassiana Ribotoxin (BbRib) Induces Silkworm Cell Apoptosis via Activating Ros Stress Response



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Prokaryotic Expression, Purification AMD Identification of Recombinant Target Protein
2.2. The Establishment of a Blood Cavity Injection of BbRib Protein to Kill the Silkworm
2.3. Preparation and Staining of Insect Paraffin Sections
2.4. ROS Activity Level and Apoptosis Level in Insect Cells or Tissues
2.5. Statistical Analysis
3. Results
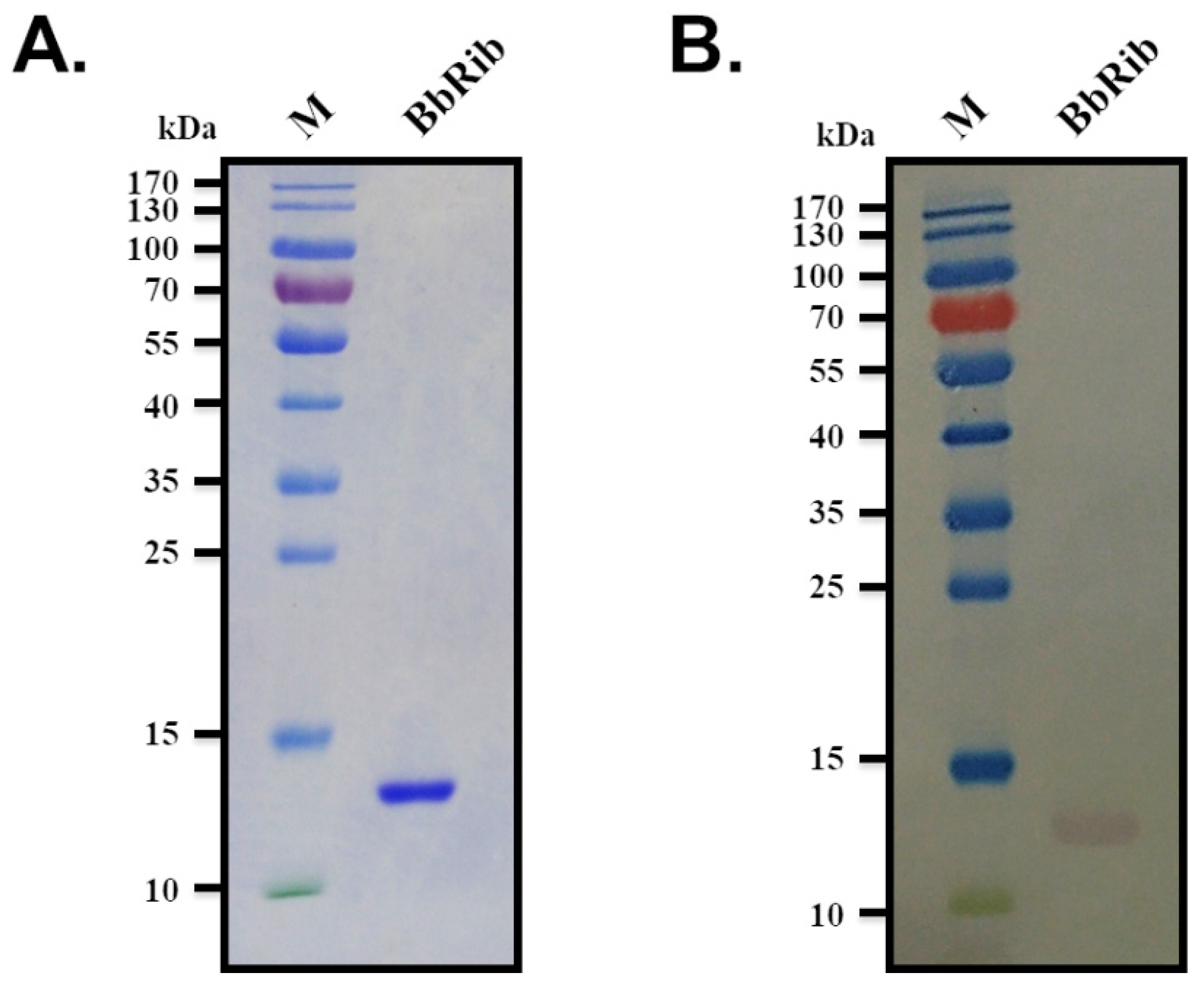
3.1. Prokaryotic Expression, Purification and Mass Spectrometry Identification of BbRibProtein
3.2. Protein Profile Identification
3.3. Changes of Physiological Indexes and Histopathological Phases of the Silkworm after BbRib Injection into the Haemocoel
3.4. Effect of BbRib Protein on Pathological Changes of Silkworm
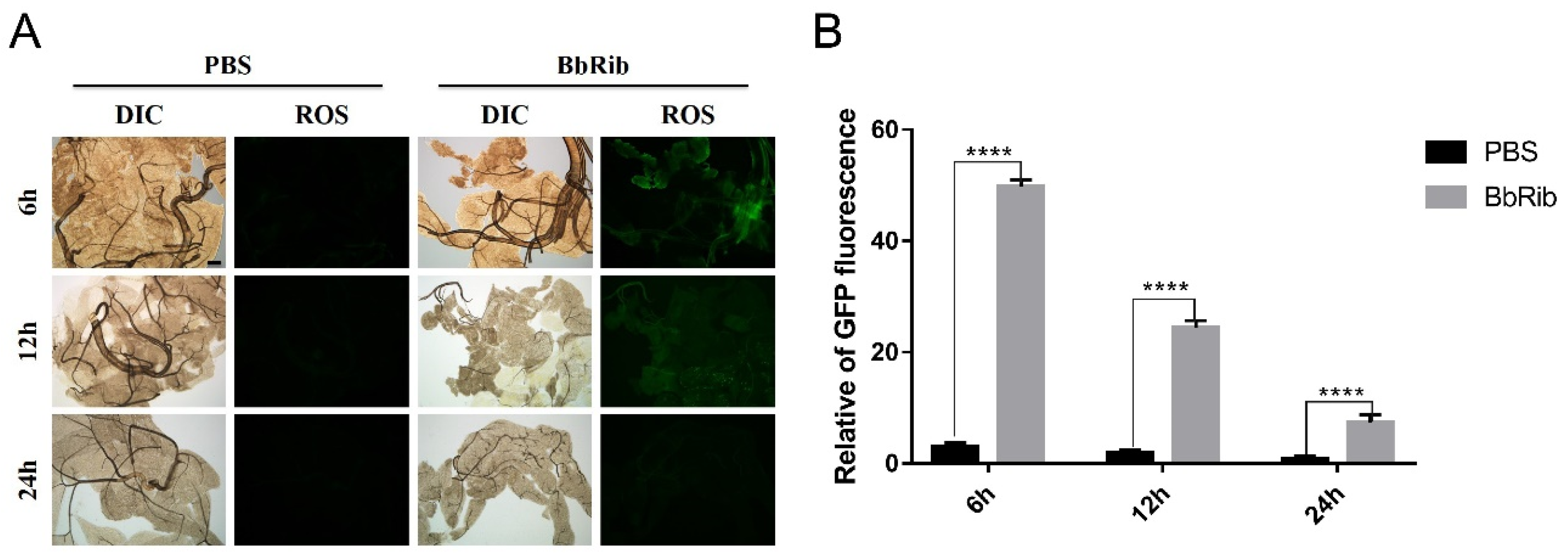
3.5. Stress Changes of ROS Activity in the Fat Body of Silkworm after Haemocoel Injection of BbRib
3.6. Injecting BbRib into the Haemocoel Can Induce Apoptosis of Adipose Tissue in the Silkworm
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, G.; Ying, S.H.; Hu, Y.; Fang, X.; Feng, M.G.; Wang, J. Roles of Three HSF Domain-Containing Proteins in Mediating Heat-Shock Protein Genes and Sustaining Asexual Cycle, Stress Tolerance, and Virulence in Beauveria bassiana. Front. Microbiol. 2018, 9, 1677. [Google Scholar] [CrossRef]
- Luo, Z.; Zhang, Y.; Jin, K.; Ma, J.; Wang, X.; Pei, Y. Construction of Beauveria bassiana T-DNA insertion mutant collections and identification of thermosensitive and osmosensitive mutants. Wei Sheng Wu Xue Bao Acta Microbiol. Sin. 2009, 49, 1301–1305. [Google Scholar]
- Matsuda, K. Okaramines and other plant fungal products as new insecticide leads. Curr. Opin. Insect Sci. 2018, 30, 67–72. [Google Scholar] [CrossRef]
- Shikano, I.; Gomez, L.; Bellicanta, G.S.; Jenkins, N.E. Persistence and Lethality of a Fungal Biopesticide (Aprehend) Applied to Insecticide-Impregnated and Encasement-Type Box Spring Covers for Bed Bug Management. J. Econ. Entomol. 2019, 112, 2489–2492. [Google Scholar] [CrossRef]
- Mkiga, A.M.; Mohamed, S.A.; du Plessis, H.; Khamis, F.M.; Akutse, K.S.; Ekesi, S. Metarhizium anisopliae and Beauveria bassiana: Pathogenicity, Horizontal Transmission, and Their Effects on Reproductive Potential of Thaumatotibia leucotreta (Lepidoptera: Tortricidae). J. Econ. Entomol. 2020, 113, 660–668. [Google Scholar] [CrossRef]
- Alspaugh, J.A. Virulence mechanisms and Cryptococcus neoformans pathogenesis. Fungal Genet. Biol. FG B 2015, 78, 55–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Wang, S. Insect Pathogenic Fungi: Genomics, Molecular Interactions, and Genetic Improvements. Annu. Rev. Entomol. 2017, 62, 73–90. [Google Scholar] [CrossRef] [PubMed]
- Mouhoumed, A.Z.; Mou, Y.N.; Tong, S.M.; Ying, S.H.; Feng, M.G. Three proline rotamases involved in calcium homeostasis play differential roles in stress tolerance, virulence and calcineurin regulation of Beauveria bassiana. Cell. Microbiol. 2020, 22, e13239. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Du, Y.; Jin, K.; Xia, Y. The Ste12-like transcription factor MaSte12 is involved in pathogenicity by regulating the appressorium formation in the entomopathogenic fungus, Metarhizium acridum. Appl. Microbiol. Biotechnol. 2017, 101, 8571–8584. [Google Scholar] [CrossRef]
- Wong Sak Hoi, J.; Dumas, B. Ste12 and Ste12-like proteins, fungal transcription factors regulating development and pathogenicity. Eukaryot. Cell 2010, 9, 480–485. [Google Scholar] [CrossRef] [Green Version]
- Wong, J.H.; Bao, H.; Ng, T.B.; Chan, H.H.L.; Ng, C.C.W.; Man, G.C.W.; Wang, H.; Guan, S.; Zhao, S.; Fang, E.F.; et al. New ribosome-inactivating proteins and other proteins with protein synthesis-inhibiting activities. Appl. Microbiol. Biotechnol. 2020, 104, 4211–4226. [Google Scholar] [CrossRef] [PubMed]
- Reyes, A.G.; Anné, J.; Mejía, A. Ribosome-inactivating proteins with an emphasis on bacterial RIPs and their potential medical applications. Future Microbiol. 2012, 7, 705–717. [Google Scholar] [CrossRef] [PubMed]
- Olombrada, M.; Lázaro-Gorines, R.; López-Rodríguez, J.C.; Martínez-Del-Pozo, Á.; Oñaderra, M.; Maestro-López, M.; Lacadena, J.; Gavilanes, J.G.; García-Ortega, L. Fungal Ribotoxins: A Review of Potential Biotechnological Applications. Toxins 2017, 9, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, R.; Martínez-Ruiz, A.; Martínez del Pozo, A.; Crameri, R.; Davies, J. Mitogillin and related fungal ribotoxins. Methods Enzymol. 2001, 341, 324–335. [Google Scholar] [CrossRef]
- Lacadena, J.; Alvarez-García, E.; Carreras-Sangrà, N.; Herrero-Galán, E.; Alegre-Cebollada, J.; García-Ortega, L.; Oñaderra, M.; Gavilanes, J.G.; Martínez del Pozo, A. Fungal ribotoxins: Molecular dissection of a family of natural killers. FEMS Microbiol. Rev. 2007, 31, 212–237. [Google Scholar] [CrossRef] [Green Version]
- Swale, D.R. Perspectives on new strategies for the identification and development of insecticide targets. Pestic. Biochem. Physiol. 2019, 161, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Zhong, G.; Cui, G.; Yi, X.; Sun, R.; Zhang, J. Insecticide cytotoxicology in China: Current status and challenges. Pestic. Biochem. Physiol. 2016, 132, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Thapa, S.; Lv, M.; Xu, H. Acetylcholinesterase: A Primary Target for Drugs and Insecticides. Mini Rev. Med. Chem. 2017, 17, 1665–1676. [Google Scholar] [CrossRef]
- Han, P.; Han, J.; Zhang, M.; Fan, J.; Gong, Q.; Ma, E.; Zhang, J. 20-Hydroxyecdysone enhances Immulectin-1 mediated immune response against entomogenous fungus in Locusta migratoria. Pest Manag. Sci. 2020, 76, 304–313. [Google Scholar] [CrossRef]
- Wichadakul, D.; Kobmoo, N.; Ingsriswang, S.; Tangphatsornruang, S.; Chantasingh, D.; Luangsa-ard, J.J.; Eurwilaichitr, L. Insights from the genome of Ophiocordyceps polyrhachis-furcata to pathogenicity and host specificity in insect fungi. BMC Genom. 2015, 16, 881. [Google Scholar] [CrossRef] [Green Version]
- Kurata, S. Fly immunity: Recognition of pathogens and induction of immune responses. Adv. Exp. Med. Biol. 2010, 708, 205–217. [Google Scholar] [CrossRef]
- Kurata, S. Recognition and elimination of diversified pathogens in insect defense systems. Mol. Divers. 2006, 10, 599–605. [Google Scholar] [CrossRef]
- Gottar, M.; Gobert, V.; Matskevich, A.A.; Reichhart, J.M.; Wang, C.; Butt, T.M.; Belvin, M.; Hoffmann, J.A.; Ferrandon, D. Dual detection of fungal infections in Drosophila via recognition of glucans and sensing of virulence factors. Cell 2006, 127, 1425–1437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Keyhani, N.O.; Yu, X.; He, Z.; Luo, Z.; Pei, Y.; Zhang, Y. The MAP kinase Bbslt2 controls growth, conidiation, cell wall integrity, and virulence in the insect pathogenic fungus Beauveria bassiana. Fungal Genet. Biol. FG B 2012, 49, 544–555. [Google Scholar] [CrossRef]
- Fang, W.; Leng, B.; Xiao, Y.; Jin, K.; Ma, J.; Fan, Y.; Feng, J.; Yang, X.; Zhang, Y.; Pei, Y. Cloning of Beauveria bassiana chitinase gene Bbchit1 and its application to improve fungal strain virulence. Appl. Environ. Microbiol. 2005, 71, 363–370. [Google Scholar] [CrossRef] [Green Version]
- Ortiz-Urquiza, A.; Keyhani, N.O. Molecular Genetics of Beauveria bassiana Infection of Insects. Adv. Genet. 2016, 94, 165–249. [Google Scholar] [CrossRef]
- Zhang, S.; Widemann, E.; Bernard, G.; Lesot, A.; Pinot, F.; Pedrini, N.; Keyhani, N.O. CYP52X1, representing new cytochrome P450 subfamily, displays fatty acid hydroxylase activity and contributes to virulence and growth on insect cuticular substrates in entomopathogenic fungus Beauveria bassiana. J. Biol. Chem. 2012, 287, 13477–13486. [Google Scholar] [CrossRef] [Green Version]
- Fan, Y.; Ortiz-Urquiza, A.; Kudia, R.A.; Keyhani, N.O. A fungal homologue of neuronal calcium sensor-1, Bbcsa1, regulates extracellular acidification and contributes to virulence in the entomopathogenic fungus Beauveria bassiana. Microbiology 2012, 158, 1843–1851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, S.H.; Feng, M.G.; Keyhani, N.O. A carbon responsive G-protein coupled receptor modulates broad developmental and genetic networks in the entomopathogenic fungus, Beauveria bassiana. Environ. Microbiol. 2013, 15, 2902–2921. [Google Scholar] [CrossRef] [PubMed]
- Lapadula, W.J.; Ayub, M.J. Ribosome Inactivating Proteins from an evolutionary perspective. Toxicon Off. J. Int. Soc. Toxinol. 2017, 136, 6–14. [Google Scholar] [CrossRef] [PubMed]
- Barbier, J.; Gillet, D. Ribosome Inactivating Proteins: From Plant Defense to Treatments against Human Misuse or Diseases. Toxins 2018, 10, 160. [Google Scholar] [CrossRef] [Green Version]
- Walsh, M.J.; Dodd, J.E.; Hautbergue, G.M. Ribosome-inactivating proteins: Potent poisons and molecular tools. Virulence 2013, 4, 774–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kao, R.; Davies, J. Molecular dissection of mitogillin reveals that the fungal ribotoxins are a family of natural genetically engineered ribonucleases. J. Biol. Chem. 1999, 274, 12576–12582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myers, A.L.; Harris, C.M.; Choe, K.M.; Brennan, C.A. Inflammatory production of reactive oxygen species by Drosophila hemocytes activates cellular immune defenses. Biochem. Biophys. Res. Commun. 2018, 505, 726–732. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C.; Cunningham-Bussel, A. Beyond oxidative stress: An immunologist’s guide to reactive oxygen species. Nat. Rev. Immunol. 2013, 13, 349–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Christophides, G.K.; Cantera, R.; Charles, B.; Han, Y.S.; Meister, S.; Dimopoulos, G.; Kafatos, F.C.; Barillas-Mury, C. The role of reactive oxygen species on Plasmodium melanotic encapsulation in Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2003, 100, 14139–14144. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Wang, W.; Ma, L.; Cui, X.; Lynch, I.; Wu, G. Acute toxicity of Zinc Oxide nanoparticles to silkworm (Bombyx mori L.). Chemosphere 2020, 259, 127481. [Google Scholar] [CrossRef]
- Ge, Q.; Cao, W.; Zhu, F.; Yuan, Y.; Chen, L.; Xu, J.; Li, J.; Chen, H.; Ma, S.; Sun, L.; et al. Genomics and proteomics combined analysis revealed the toxicity response of silkworm Bombyx mori to the environmental pathogen Bacillus cereus ZJ-4. Ecotoxicol. Environ. Saf. 2021, 222, 112467. [Google Scholar] [CrossRef]
- García-Mayoral, M.F.; Pérez-Cañadillas, J.M.; Santoro, J.; Ibarra-Molero, B.; Sanchez-Ruiz, J.M.; Lacadena, J.; Martínez del Pozo, A.; Gavilanes, J.G.; Rico, M.; Bruix, M. Dissecting structural and electrostatic interactions of charged groups in alpha-sarcin. An NMR study of some mutants involving the catalytic residues. Biochemistry 2003, 42, 13122–13133. [Google Scholar] [CrossRef]
- Narla, A.; Ebert, B.L. Ribosomopathies: Human disorders of ribosome dysfunction. Blood 2010, 115, 3196–3205. [Google Scholar] [CrossRef] [PubMed]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, X.; Ge, Q.; Taha, R.H.; Chen, K.; Yuan, Y. Beauveria bassiana Ribotoxin (BbRib) Induces Silkworm Cell Apoptosis via Activating Ros Stress Response. Processes 2021, 9, 1470. https://doi.org/10.3390/pr9081470
Ma X, Ge Q, Taha RH, Chen K, Yuan Y. Beauveria bassiana Ribotoxin (BbRib) Induces Silkworm Cell Apoptosis via Activating Ros Stress Response. Processes. 2021; 9(8):1470. https://doi.org/10.3390/pr9081470
Chicago/Turabian StyleMa, Xiaoke, Qi Ge, Rehab Hosny Taha, Keping Chen, and Yi Yuan. 2021. "Beauveria bassiana Ribotoxin (BbRib) Induces Silkworm Cell Apoptosis via Activating Ros Stress Response" Processes 9, no. 8: 1470. https://doi.org/10.3390/pr9081470
APA StyleMa, X., Ge, Q., Taha, R. H., Chen, K., & Yuan, Y. (2021). Beauveria bassiana Ribotoxin (BbRib) Induces Silkworm Cell Apoptosis via Activating Ros Stress Response. Processes, 9(8), 1470. https://doi.org/10.3390/pr9081470