
Unlocking the Treasure Box: The Role of HEPES Buffer in Disassembling an Uncommon Ferritin Nanoparticle

 , ,
, ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mutagenesis
2.2. Protein Expression and Purification
2.3. Chromatographic Analyses
2.4. Transmission Electron Microscopy Negative Staining
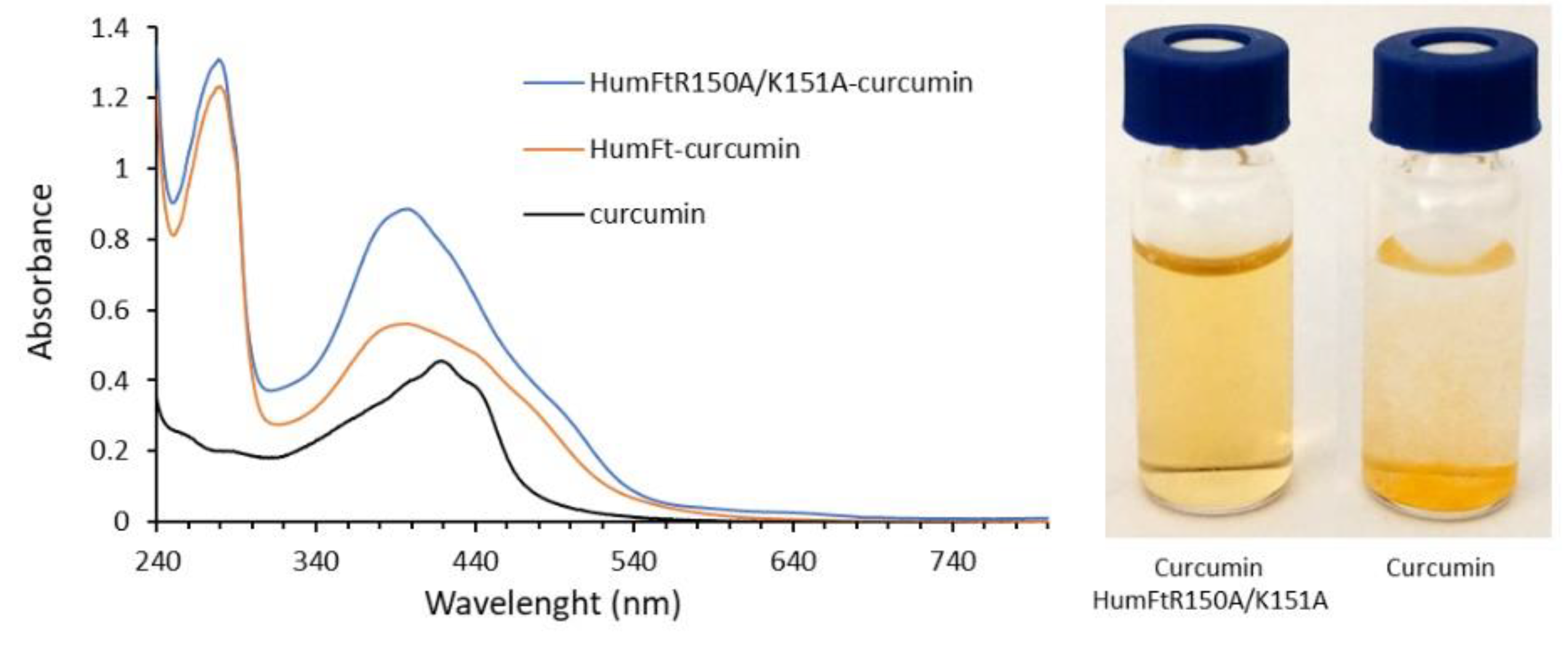
2.5. Curcumin-Loaded Nanoparticles
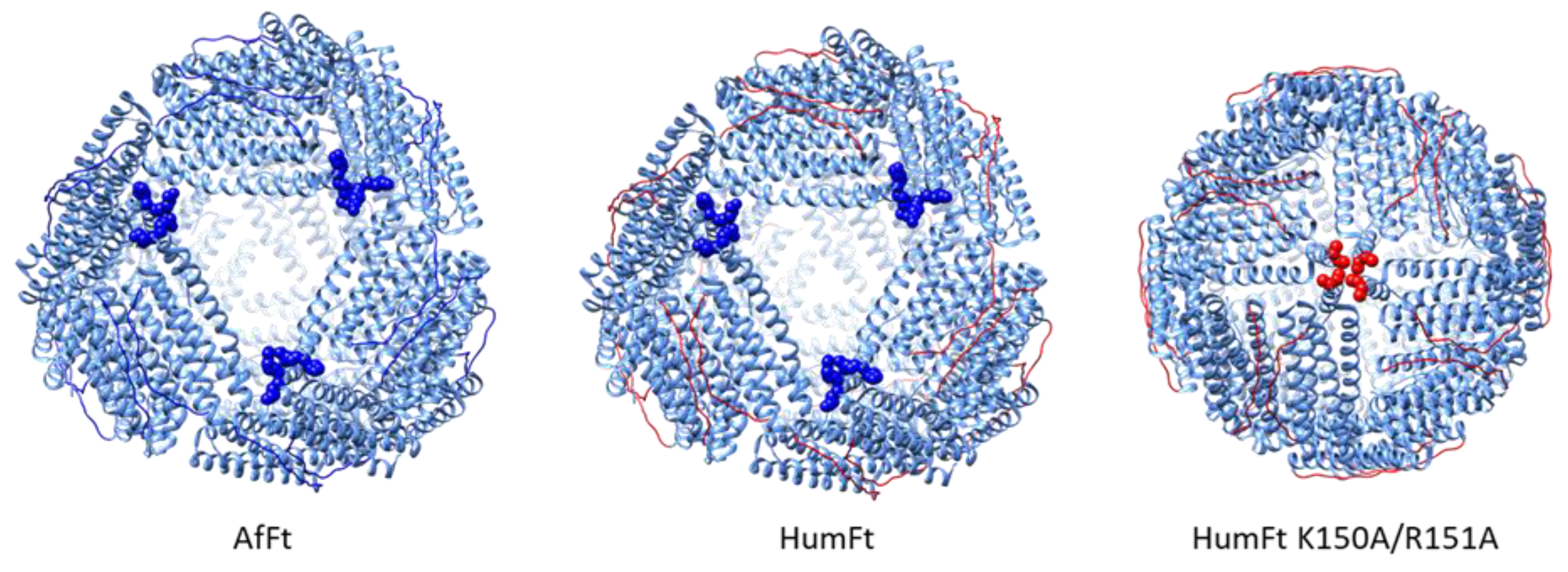
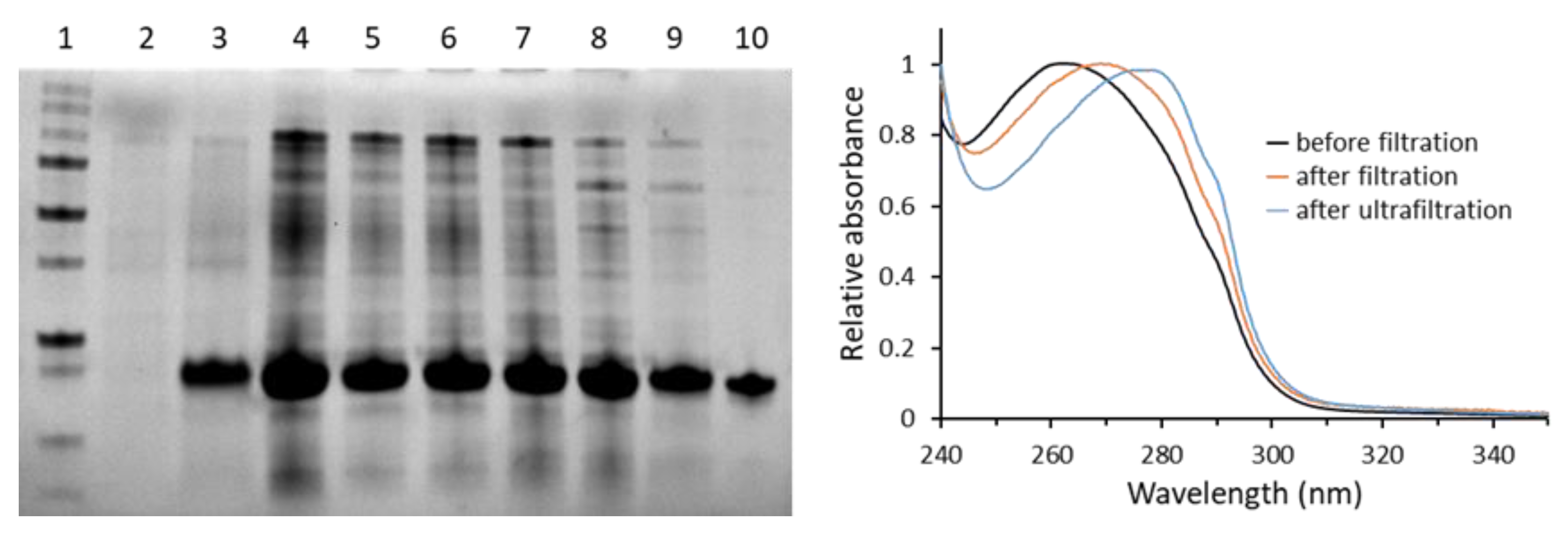
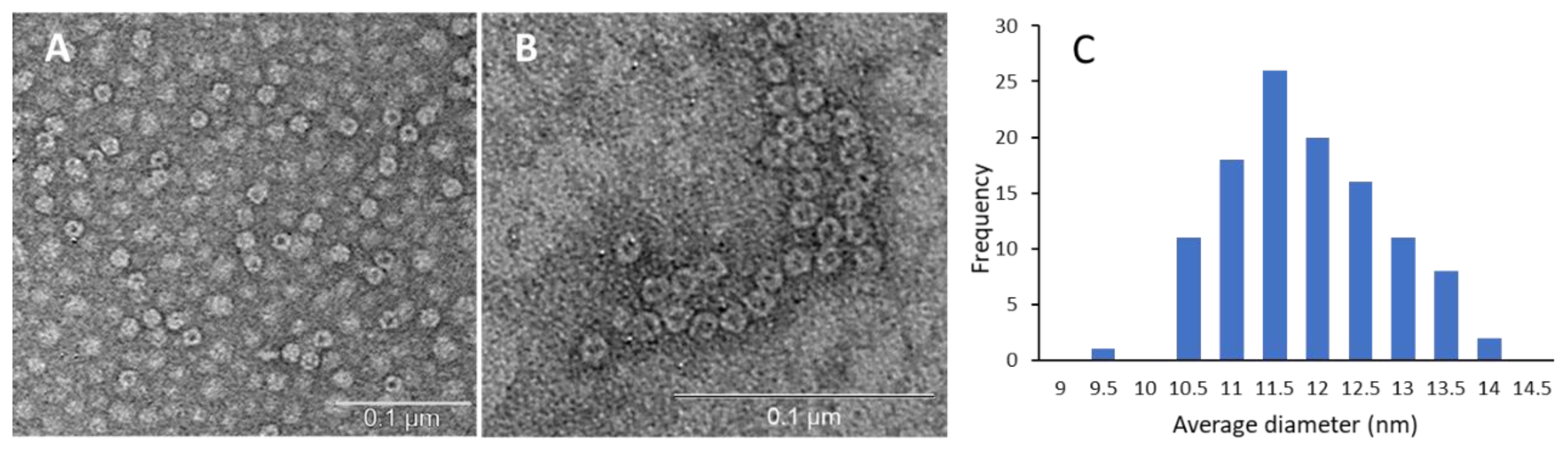
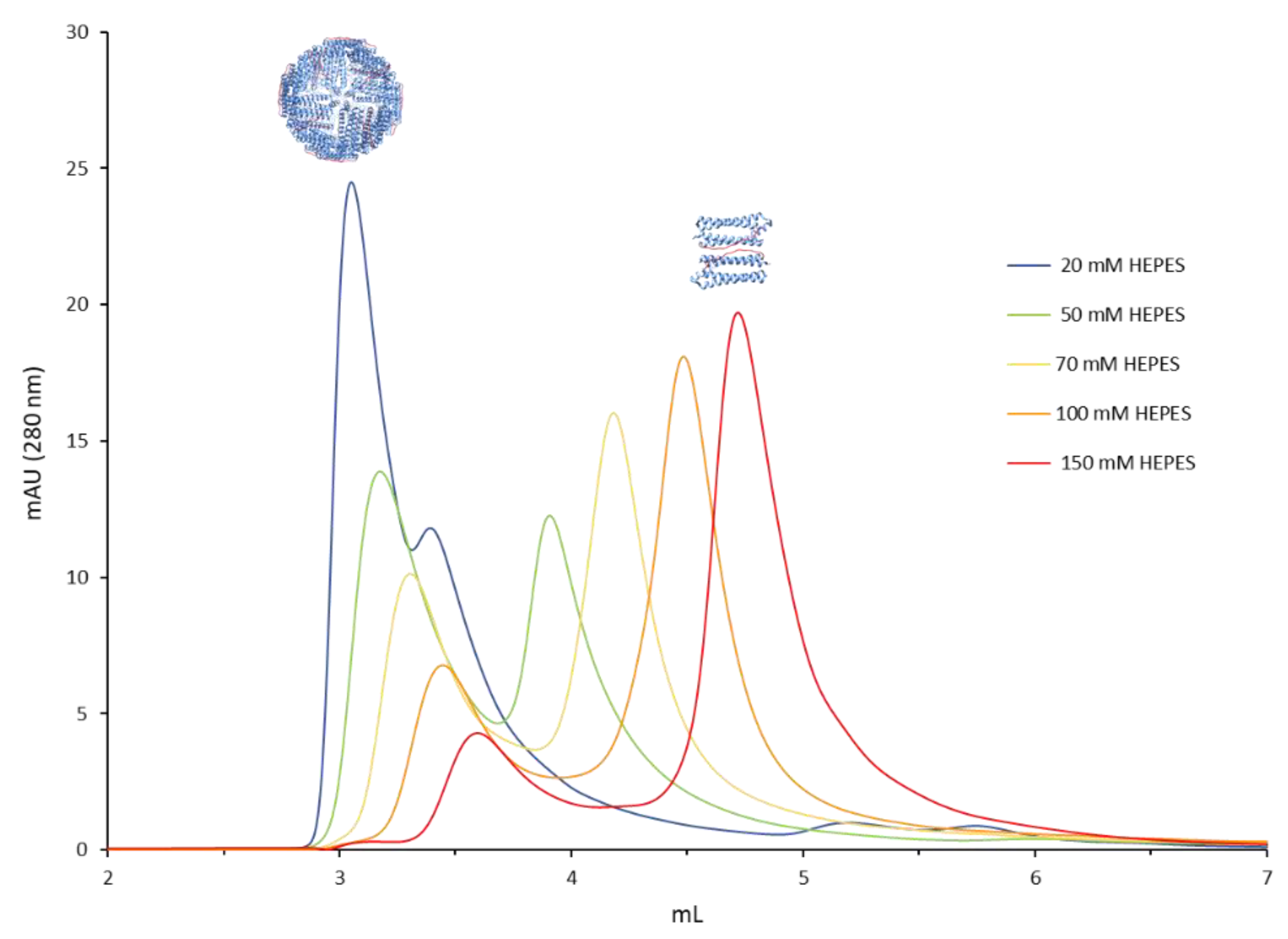
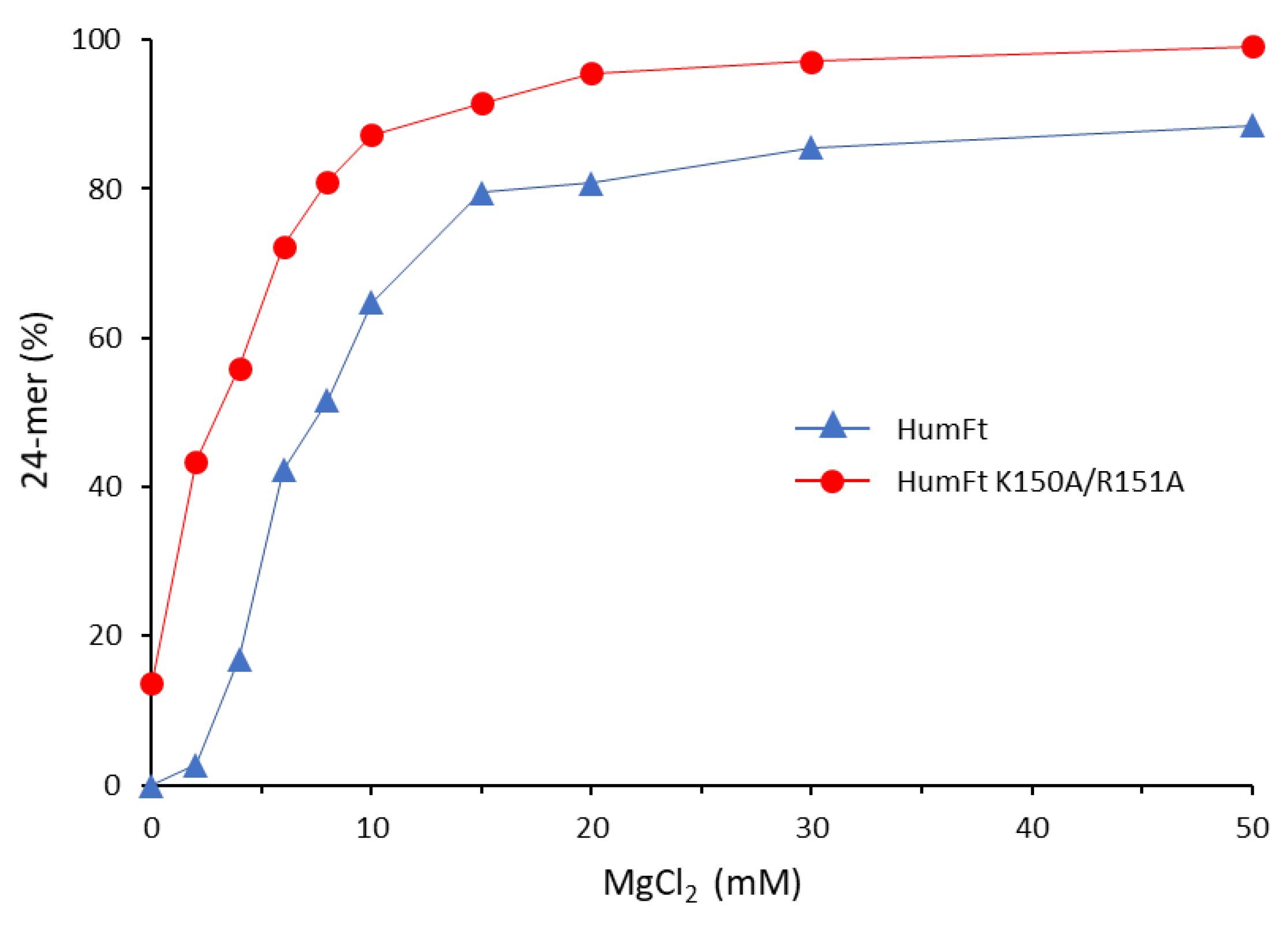
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yao, Y.; Zhou, Y.; Liu, L.; Xu, Y.; Chen, Q.; Wang, Y.; Wu, S.; Deng, Y.; Zhang, J.; Shao, A. Nanoparticle-Based Drug Delivery in Cancer Therapy and Its Role in Overcoming Drug Resistance. Front. Mol. Biosci. 2020, 7, 193. [Google Scholar] [CrossRef] [PubMed]
- Van der Meel, R.; Sulheim, E.; Shi, Y.; Kiessling, F.; Mulder, W.J.M.; Lammers, T. Smart Cancer Nanomedicine. Nat. Nanotechnol. 2019, 14, 1007–1017. [Google Scholar] [CrossRef]
- Noireaux, J.; Grall, R.; Hullo, M.; Chevillard, S.; Oster, C.; Brun, E.; Sicard-Roselli, C.; Loeschner, K.; Fisicaro, P. Gold Nanoparticle Uptake in Tumor Cells: Quantification and Size Distribution by Sp-ICPMS. Separations 2019, 6, 3. [Google Scholar] [CrossRef]
- Niculescu, A.-G.; Grumezescu, A.M. Novel Tumor-Targeting Nanoparticles for Cancer Treatment—A Review. Int. J. Mol. Sci. 2022, 23, 5253. [Google Scholar] [CrossRef] [PubMed]
- Iranpour, S.; Bahrami, A.R.; Nekooei, S.; Saljooghi, A.S.; Matin, M.M. Improving Anti-Cancer Drug Delivery Performance of Magnetic Mesoporous Silica Nanocarriers for More Efficient Colorectal Cancer Therapy. J. Nanobiotechnol. 2021, 19, 314. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-S. Liposomal Drug Delivery System. J. Pharm. Investig. 2016, 46, 387–392. [Google Scholar] [CrossRef]
- Kianfar, E. Protein Nanoparticles in Drug Delivery: Animal Protein, Plant Proteins and Protein Cages, Albumin Nanoparticles. J. Nanobiotechnol. 2021, 19, 159. [Google Scholar] [CrossRef]
- Patra, J.K.; Das, G.; Fraceto, L.F.; Campos, E.V.R.; del Pilar Rodriguez-Torres, M.; Acosta-Torres, L.S.; Diaz-Torres, L.A.; Grillo, R.; Swamy, M.K.; Sharma, S.; et al. Nano Based Drug Delivery Systems: Recent Developments and Future Prospects. J. Nanobiotechnol. 2018, 16, 71. [Google Scholar] [CrossRef]
- Montemiglio, L.C.; Testi, C.; Ceci, P.; Falvo, E.; Pitea, M.; Savino, C.; Arcovito, A.; Peruzzi, G.; Baiocco, P.; Mancia, F.; et al. Cryo-EM Structure of the Human Ferritin–Transferrin Receptor 1 Complex. Nat. Commun. 2019, 10, 1121. [Google Scholar] [CrossRef]
- Wang, Z.; Gao, H.; Zhang, Y.; Liu, G.; Niu, G.; Chen, X. Functional Ferritin Nanoparticles for Biomedical Applications. Front. Chem. Sci. Eng. 2017, 11, 633–646. [Google Scholar] [CrossRef]
- Khoshnejad, M.; Parhiz, H.; Shuvaev, V.V.; Dmochowski, I.J.; Muzykantov, V.R. Ferritin-Based Drug Delivery Systems: Hybrid Nanocarriers for Vascular Immunotargeting. J. Control. Release 2018, 282, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Mainini, F.; Bonizzi, A.; Sevieri, M.; Sitia, L.; Truffi, M.; Corsi, F.; Mazzucchelli, S. Protein-Based Nanoparticles for the Imaging and Treatment of Solid Tumors: The Case of Ferritin Nanocages, a Narrative Review. Pharmaceutics 2021, 13, 2000. [Google Scholar] [CrossRef] [PubMed]
- Truffi, M.; Fiandra, L.; Sorrentino, L.; Monieri, M.; Corsi, F.; Mazzucchelli, S. Ferritin Nanocages: A Biological Platform for Drug Delivery, Imaging and Theranostics in Cancer. Pharmacol. Res. 2016, 107, 57–65. [Google Scholar] [CrossRef] [PubMed]
- Calisti, L.; Trabuco, M.C.; Boffi, A.; Testi, C.; Montemiglio, L.C.; des Georges, A.; Benni, I.; Ilari, A.; Taciak, B.; Białasek, M.; et al. Engineered Ferritin for Lanthanide Binding. PLoS ONE 2018, 13, e0201859. [Google Scholar] [CrossRef]
- Zhang, Y.; Orner, B.P. Self-Assembly in the Ferritin Nano-Cage Protein Superfamily. Int. J. Mol. Sci. 2011, 12, 5406–5421. [Google Scholar] [CrossRef]
- Lawson, D.M.; Artymiuk, P.J.; Yewdall, S.J.; Smith, J.M.A.; Livingstone, J.C.; Treffry, A.; Luzzago, A.; Levi, S.; Arosio, P.; Cesareni, G.; et al. Solving the Structure of Human H Ferritin by Genetically Engineering Intermolecular Crystal Contacts. Nature 1991, 349, 541–544. [Google Scholar] [CrossRef]
- Palombarini, F.; Di Fabio, E.; Boffi, A.; Macone, A.; Bonamore, A. Ferritin Nanocages for Protein Delivery to Tumor Cells. Molecules 2020, 25, 825. [Google Scholar] [CrossRef]
- Kim, M.; Rho, Y.; Jin, K.S.; Ahn, B.; Jung, S.; Kim, H.; Ree, M. PH-Dependent Structures of Ferritin and Apoferritin in Solution: Disassembly and Reassembly. Biomacromolecules 2011, 12, 1629–1640. [Google Scholar] [CrossRef]
- Stühn, L.; Auernhammer, J.; Dietz, C. PH-Depended Protein Shell Dis- and Reassembly of Ferritin Nanoparticles Revealed by Atomic Force Microscopy. Sci. Rep. 2019, 9, 17755. [Google Scholar] [CrossRef]
- Mohanty, A.; K, M.; Jena, S.S.; Behera, R.K. Kinetics of Ferritin Self-Assembly by Laser Light Scattering: Impact of Subunit Concentration, PH, and Ionic Strength. Biomacromolecules 2021, 22, 1389–1398. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, S.; Xu, C.; Zhao, G. Engineering Protein Interfaces Yields Ferritin Disassembly and Reassembly under Benign Experimental Conditions. Chem. Commun. 2016, 52, 7402–7405. [Google Scholar] [CrossRef] [PubMed]
- Calisti, L.; Benni, I.; Cardoso Trabuco, M.; Baiocco, P.; Ruzicka, B.; Boffi, A.; Falvo, E.; Malatesta, F.; Bonamore, A. Probing Bulky Ligand Entry in Engineered Archaeal Ferritins. Biochim. Biophys. Acta BBA-Gen. Subj. 2017, 1861, 450–456. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.; Cascio, D.; Sawaya, M.R.; Gingery, M.; Schröder, I. Crystal Structures of a Tetrahedral Open Pore Ferritin from the Hyperthermophilic Archaeon Archaeoglobus Fulgidus. Structure 2005, 13, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Swift, J.; Butts, C.A.; Cheung-Lau, J.; Yerubandi, V.; Dmochowski, I.J. Efficient Self-Assembly of Archaeoglobus Fulgidus Ferritin around Metallic Cores. Langmuir 2009, 25, 5219–5225. [Google Scholar] [CrossRef]
- De Turris, V.; Cardoso Trabuco, M.; Peruzzi, G.; Boffi, A.; Testi, C.; Vallone, B.; Celeste Montemiglio, L.; Georges, A.D.; Calisti, L.; Benni, I.; et al. Humanized Archaeal Ferritin as a Tool for Cell Targeted Delivery. Nanoscale 2017, 9, 647–655. [Google Scholar] [CrossRef]
- Benni, I.; Trabuco, M.C.; Di Stasio, E.; Arcovito, A.; Boffi, A.; Malatesta, F.; Bonamore, A.; De Panfilis, S.; de Turris, V.; Baiocco, P. Excimer Based Fluorescent Pyrene–Ferritin Conjugate for Protein Oligomerization Studies and Imaging in Living Cells. RSC Adv. 2018, 8, 12815–12822. [Google Scholar] [CrossRef]
- Palombarini, F.; Masciarelli, S.; Incocciati, A.; Liccardo, F.; Di Fabio, E.; Iazzetti, A.; Fabrizi, G.; Fazi, F.; Macone, A.; Bonamore, A.; et al. Self-Assembling Ferritin-Dendrimer Nanoparticles for Targeted Delivery of Nucleic Acids to Myeloid Leukemia Cells. J. Nanobiotechnol. 2021, 19, 172. [Google Scholar] [CrossRef]
- Macone, A.; Masciarelli, S.; Palombarini, F.; Quaglio, D.; Boffi, A.; Trabuco, M.C.; Baiocco, P.; Fazi, F.; Bonamore, A. Ferritin Nanovehicle for Targeted Delivery of Cytochrome C to Cancer Cells. Sci. Rep. 2019, 9, 11749. [Google Scholar] [CrossRef]
- Sana, B.; Johnson, E.; Le Magueres, P.; Criswell, A.; Cascio, D.; Lim, S. The Role of Nonconserved Residues of Archaeoglobus Fulgidus Ferritin on Its Unique Structure and Biophysical Properties. J. Biol. Chem. 2013, 288, 32663–32672. [Google Scholar] [CrossRef]
- Sana, B.; Johnson, E.; Lim, S. The Unique Self-Assembly/Disassembly Property of Archaeoglobus Fulgidus Ferritin and Its Implications on Molecular Release from the Protein Cage. Biochim. Biophys. Acta BBA-Gen. Subj. 2015, 1850, 2544–2551. [Google Scholar] [CrossRef]
- Palombarini, F.; Ghirga, F.; Boffi, A.; Macone, A.; Bonamore, A. Application of Crossflow Ultrafiltration for Scaling up the Purification of a Recombinant Ferritin. Protein Expr. Purif. 2019, 163, 105451. [Google Scholar] [CrossRef] [PubMed]
- Brudar, S.; Hribar-Lee, B. Effect of Buffer on Protein Stability in Aqueous Solutions: A Simple Protein Aggregation Model. J. Phys. Chem. B 2021, 125, 2504–2512. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.; Keeling, R.; Tracka, M.; van der Walle, C.F.; Uddin, S.; Warwicker, J.; Curtis, R. Specific Ion and Buffer Effects on Protein–Protein Interactions of a Monoclonal Antibody. Mol. Pharm. 2015, 12, 179–193. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Bai, G.; Yang, S.; Yang, R.; Zhao, G.; Xu, C.; Leung, W. Encapsulation of curcumin in recombinant human H-chain ferritin increases its water-solubility and stability. Food Res. Int. 2014, 62, 1147–1153. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Incocciati, A.; Bertuccini, L.; Boffi, A.; Macone, A.; Bonamore, A. Unlocking the Treasure Box: The Role of HEPES Buffer in Disassembling an Uncommon Ferritin Nanoparticle. Separations 2022, 9, 222. https://doi.org/10.3390/separations9080222
Incocciati A, Bertuccini L, Boffi A, Macone A, Bonamore A. Unlocking the Treasure Box: The Role of HEPES Buffer in Disassembling an Uncommon Ferritin Nanoparticle. Separations. 2022; 9(8):222. https://doi.org/10.3390/separations9080222
Chicago/Turabian StyleIncocciati, Alessio, Lucia Bertuccini, Alberto Boffi, Alberto Macone, and Alessandra Bonamore. 2022. "Unlocking the Treasure Box: The Role of HEPES Buffer in Disassembling an Uncommon Ferritin Nanoparticle" Separations 9, no. 8: 222. https://doi.org/10.3390/separations9080222
APA StyleIncocciati, A., Bertuccini, L., Boffi, A., Macone, A., & Bonamore, A. (2022). Unlocking the Treasure Box: The Role of HEPES Buffer in Disassembling an Uncommon Ferritin Nanoparticle. Separations, 9(8), 222. https://doi.org/10.3390/separations9080222