
Affinity-Based Luminescent Iridium(III) Complexes for the Detection of Disease-Related Proteins

 ,
, 
 ,
,  and
and 
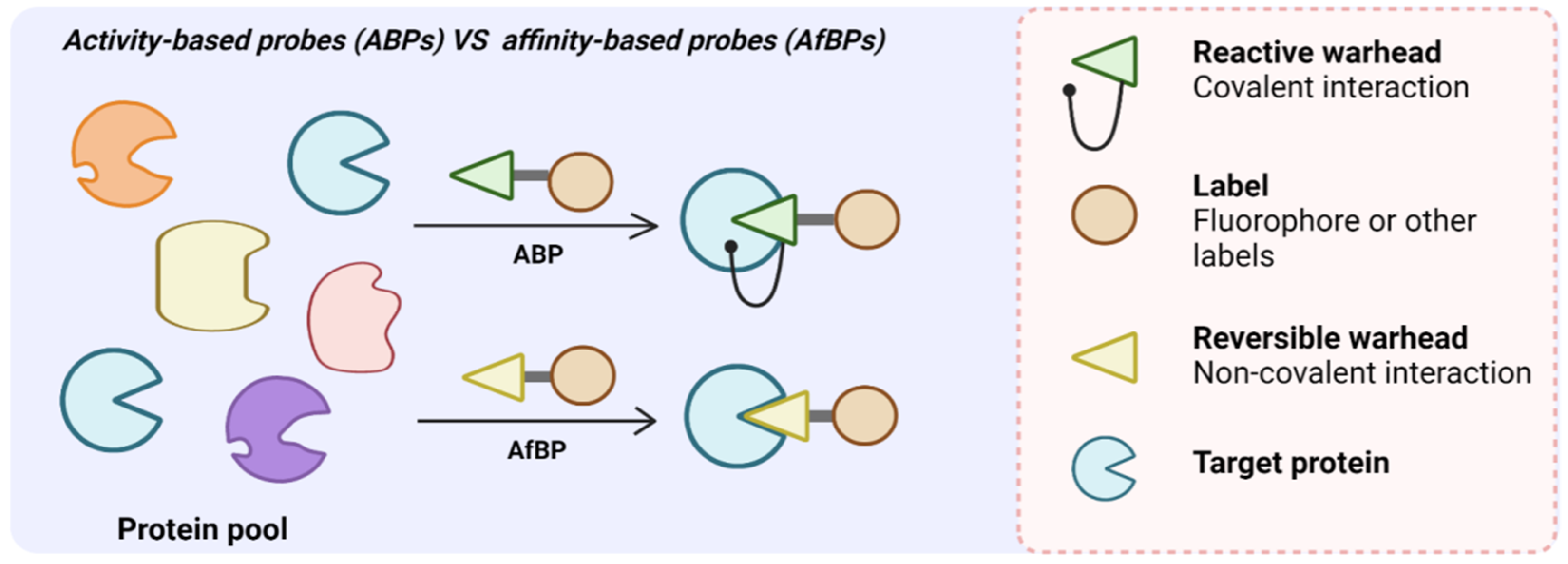{kind=link}
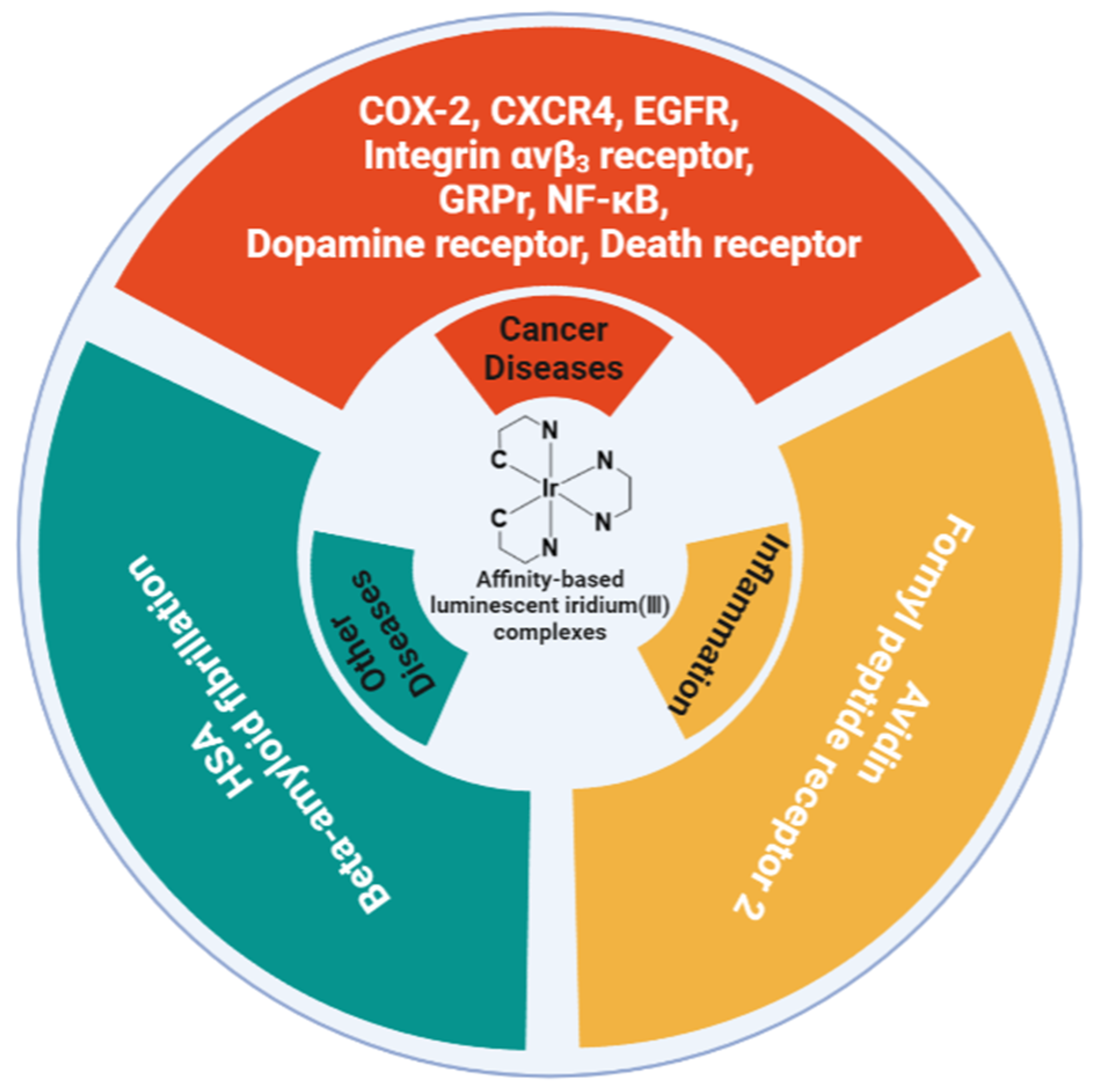{kind=link}
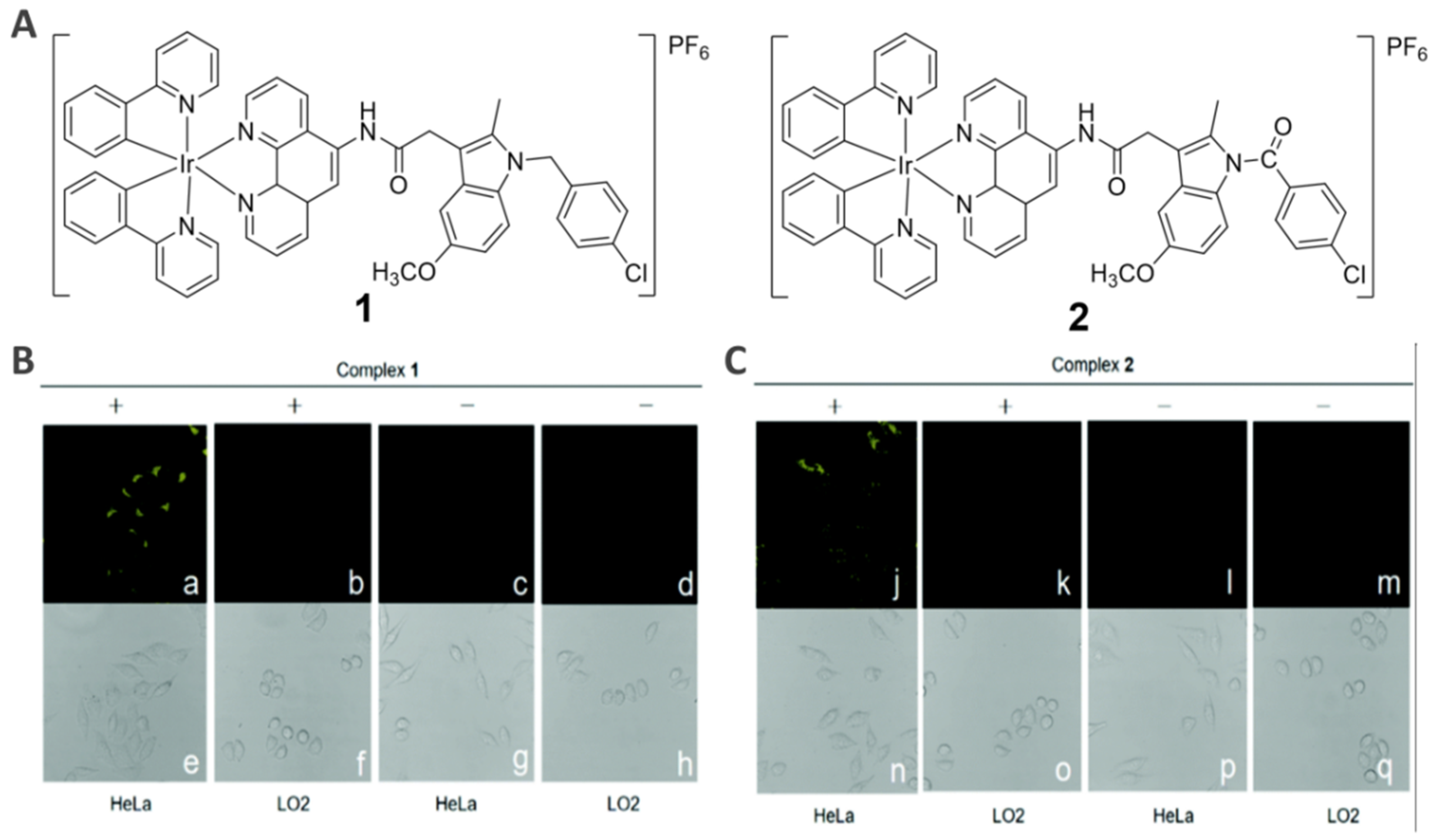{kind=link}
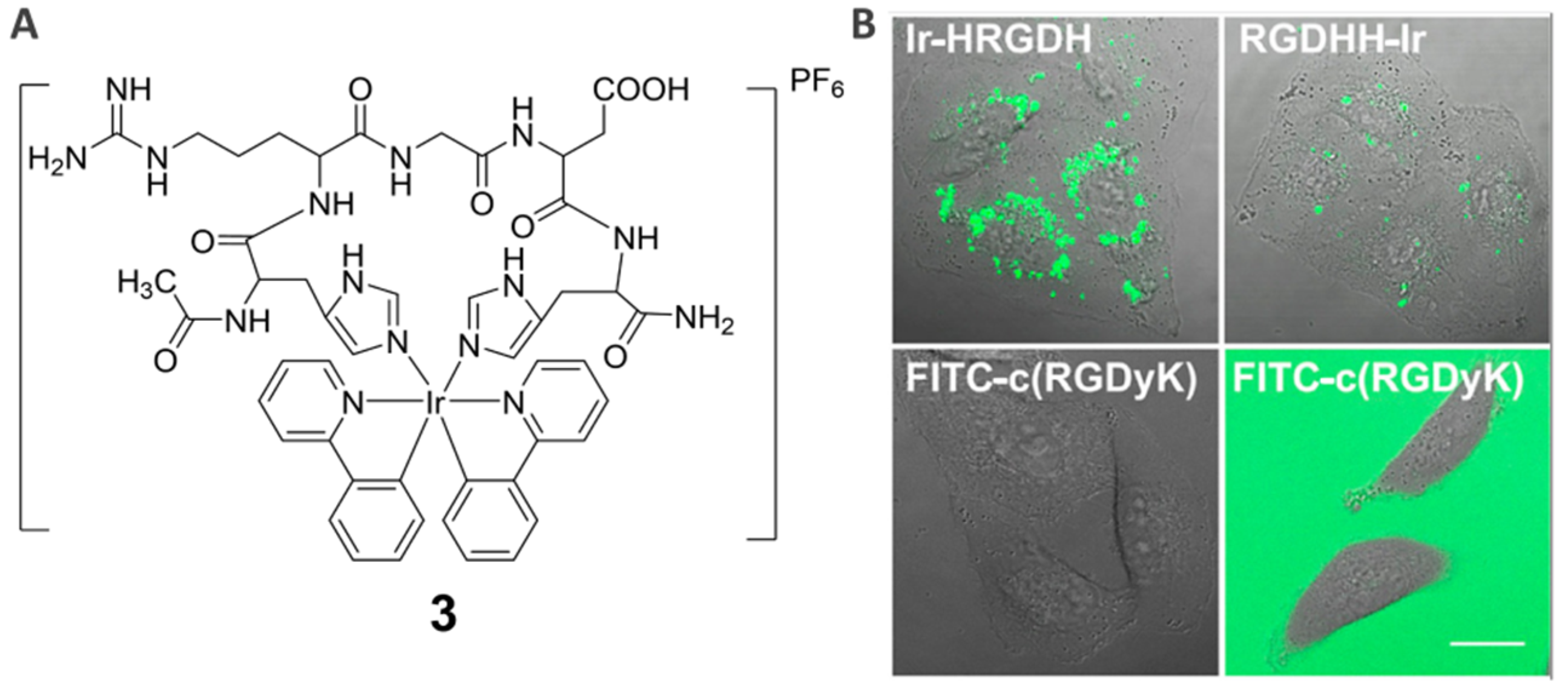{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Cancer-Related Probes
2.1. COX-2-Specific Imaging Agents (1−2)
2.2. Integrin ανβ3 Receptor-Specific Imaging Agents (3)
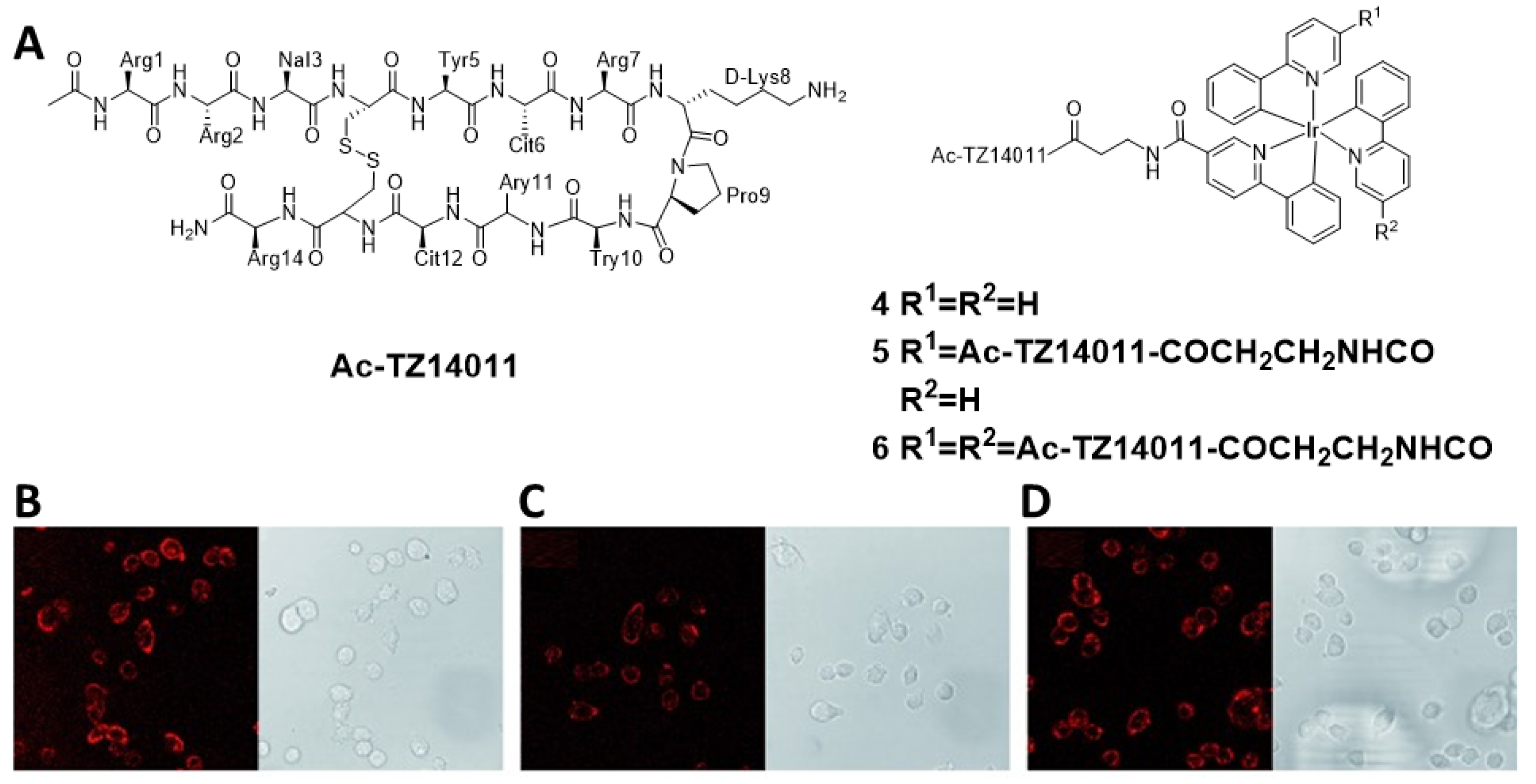
2.3. CXCR4-Specific Imaging Agents (4−6)
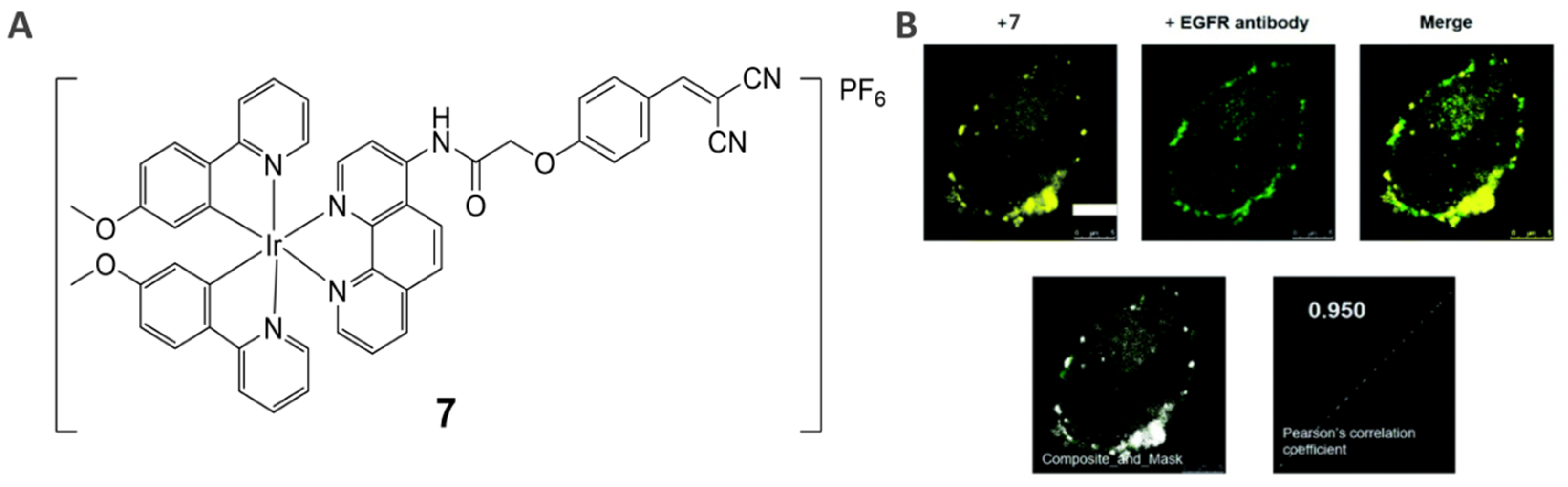
2.4. EGFR-Specific Theranostic Agents (7)
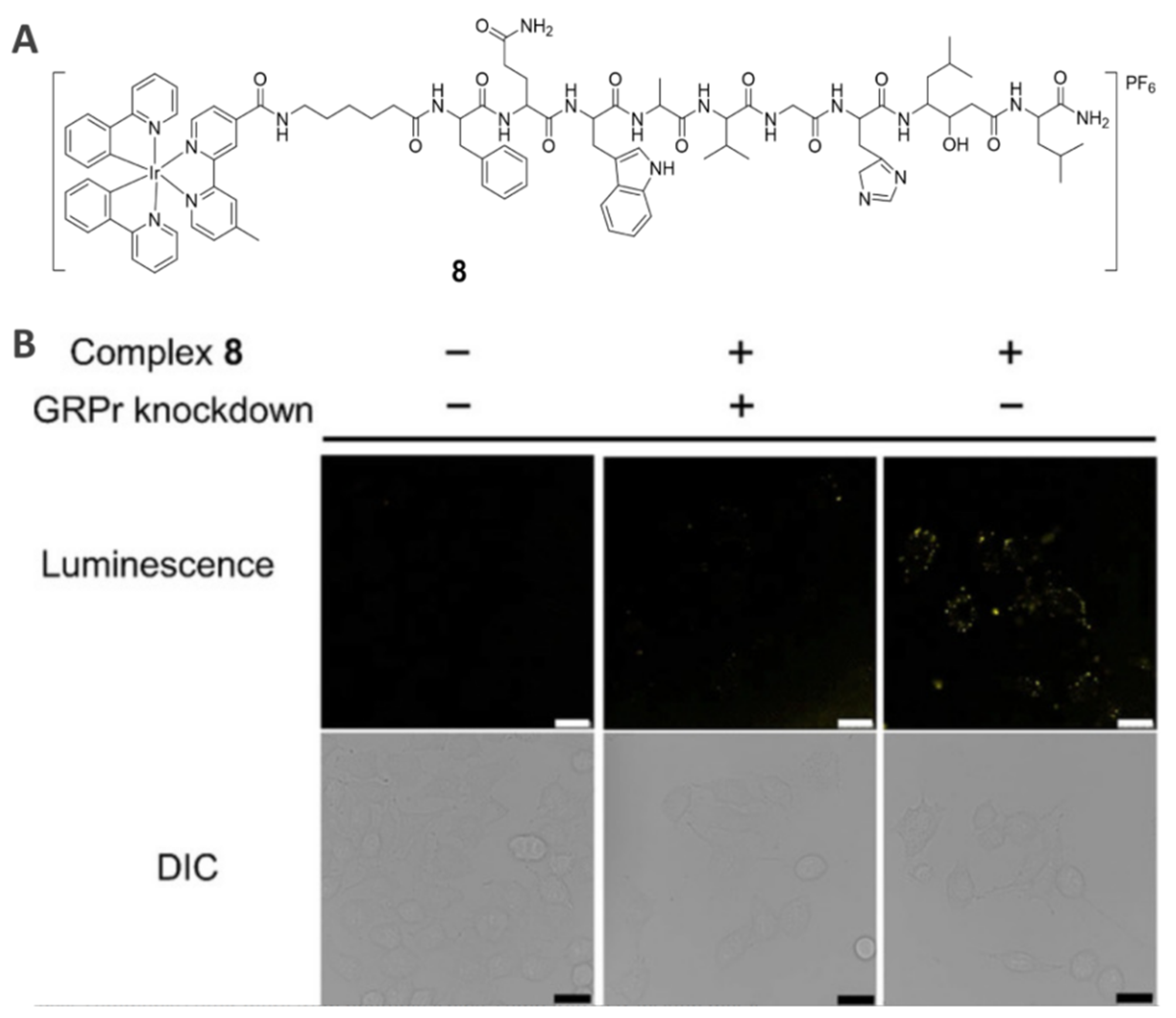
2.5. GRPr-Specific Theranostic Agents (8)
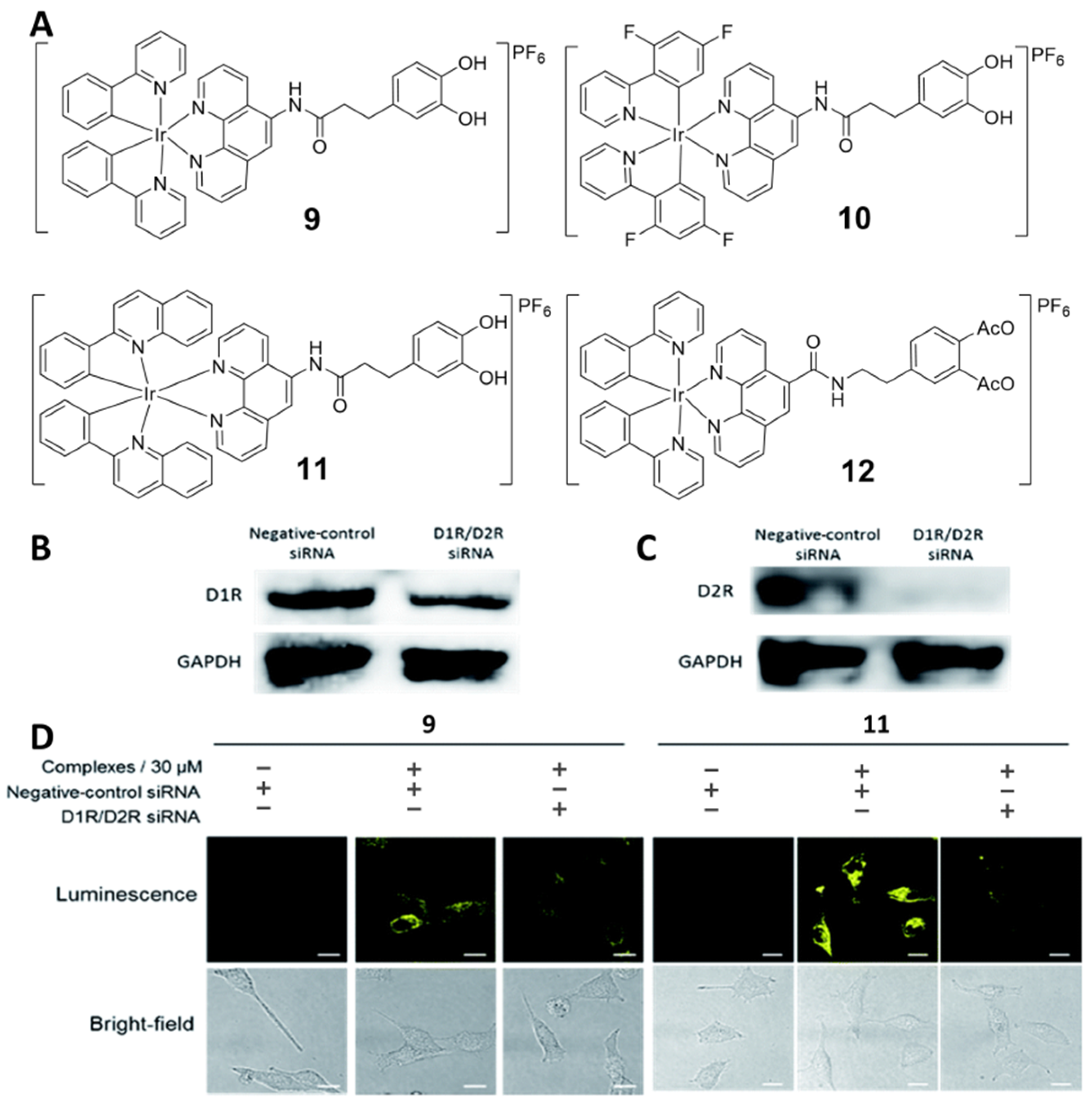
2.6. Dopamine Receptor-Specific Imaging Agents (9−12)
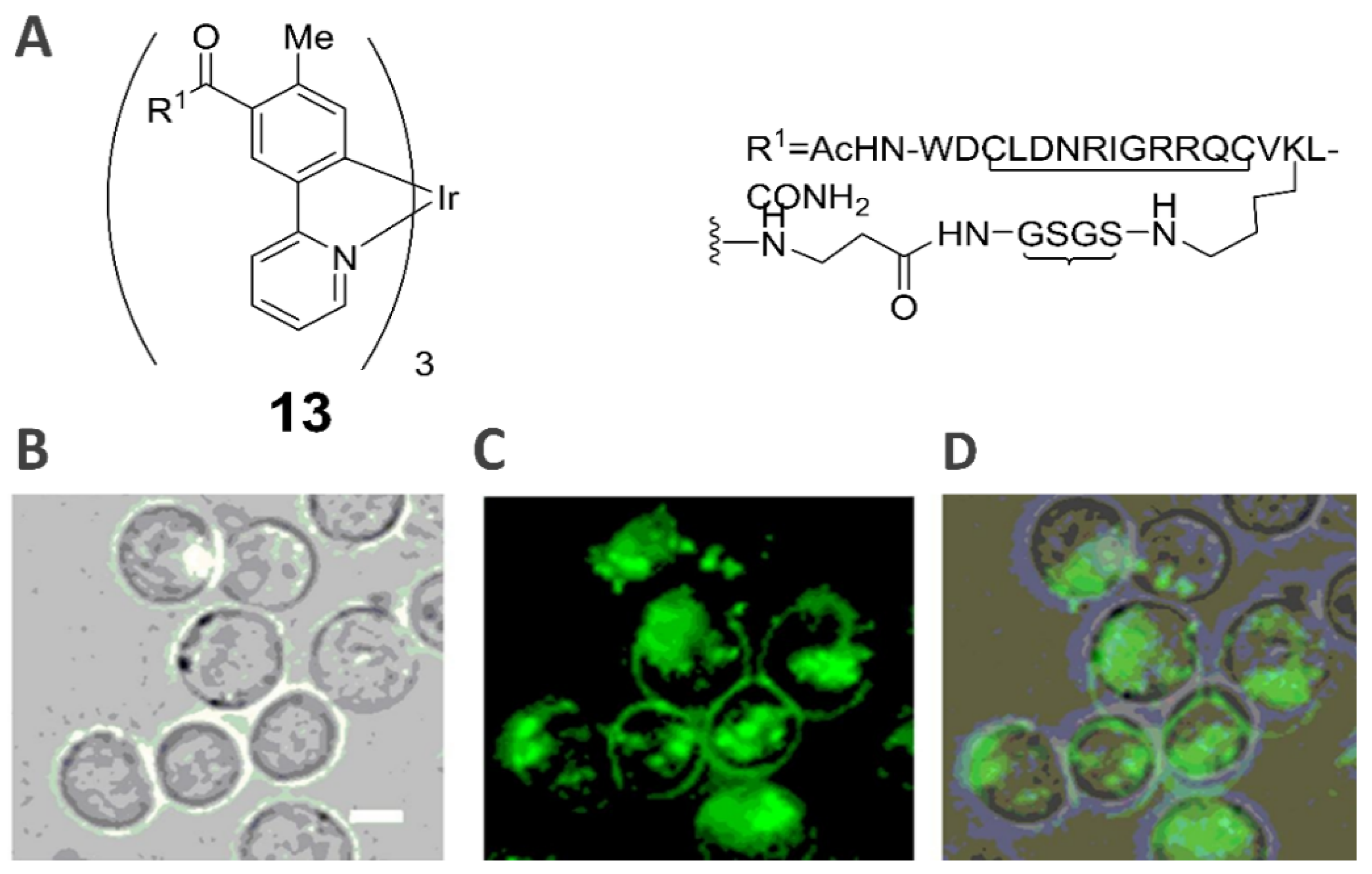
2.7. Death Receptor-Specific Theranostic Probe (13)
3. Inflammation-Related Probes
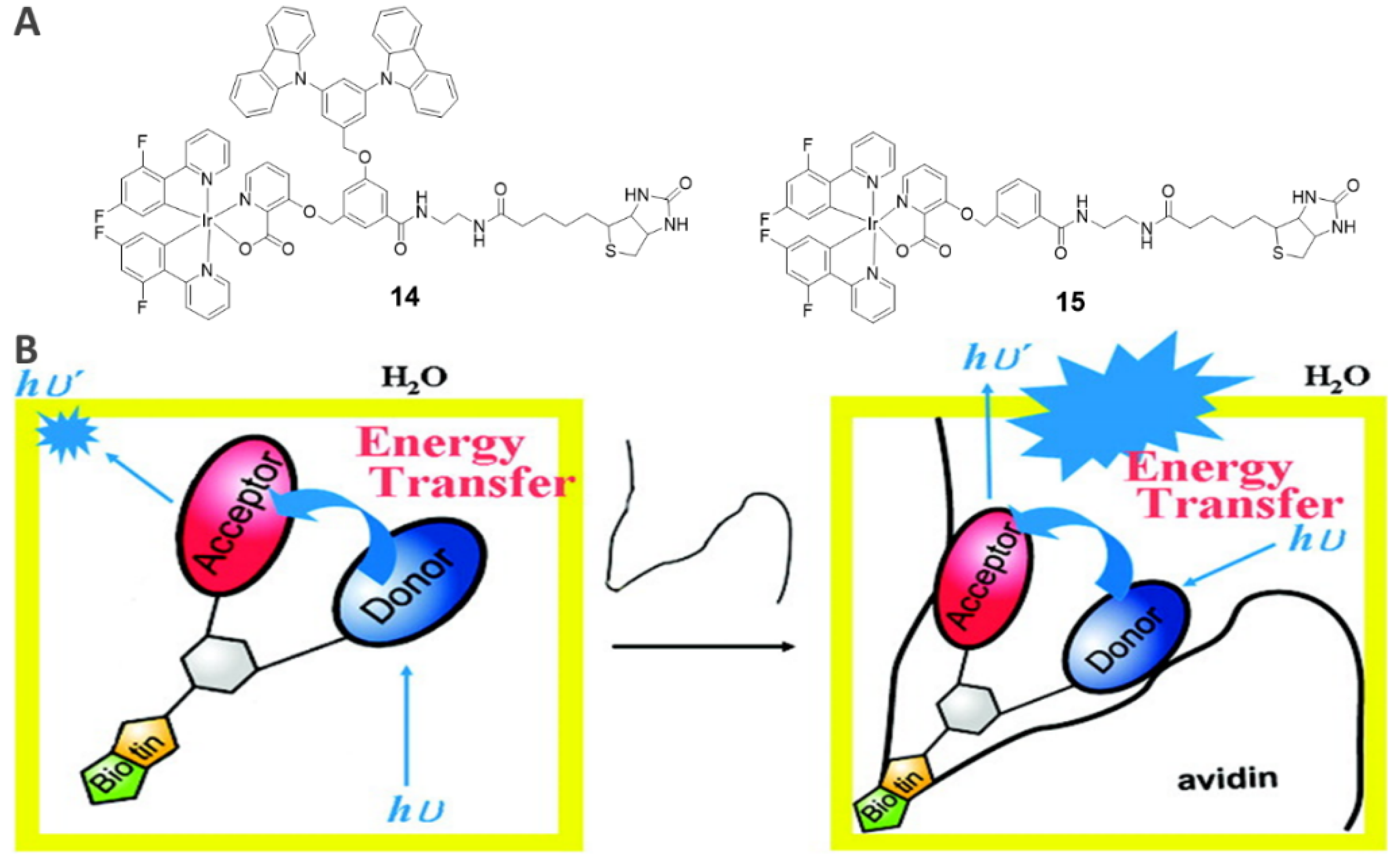
3.1. Avidin-Specific Luminescence Probes (14−15)
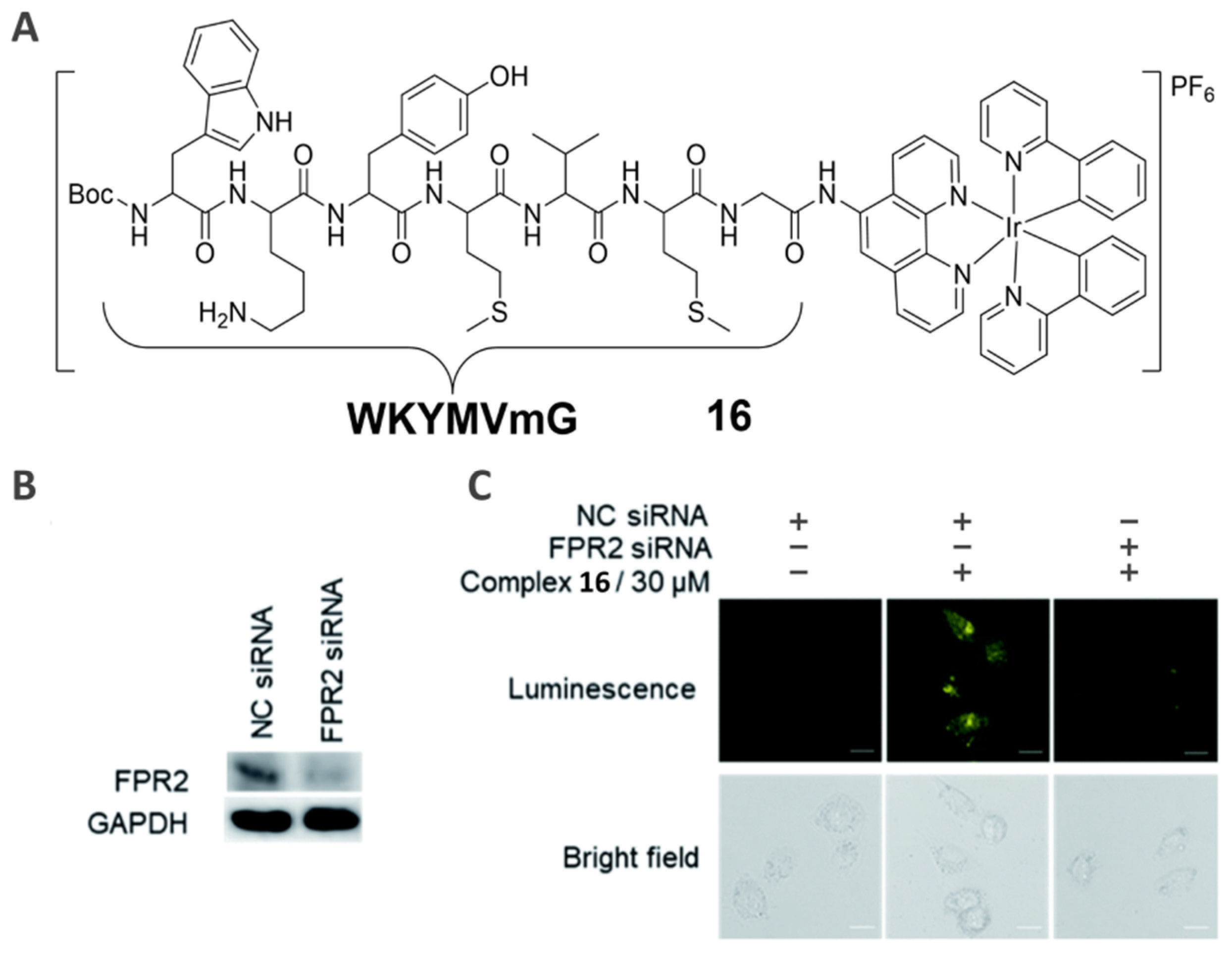
3.2. Formyl Peptide Receptor-Specific Imaging Agent (16)
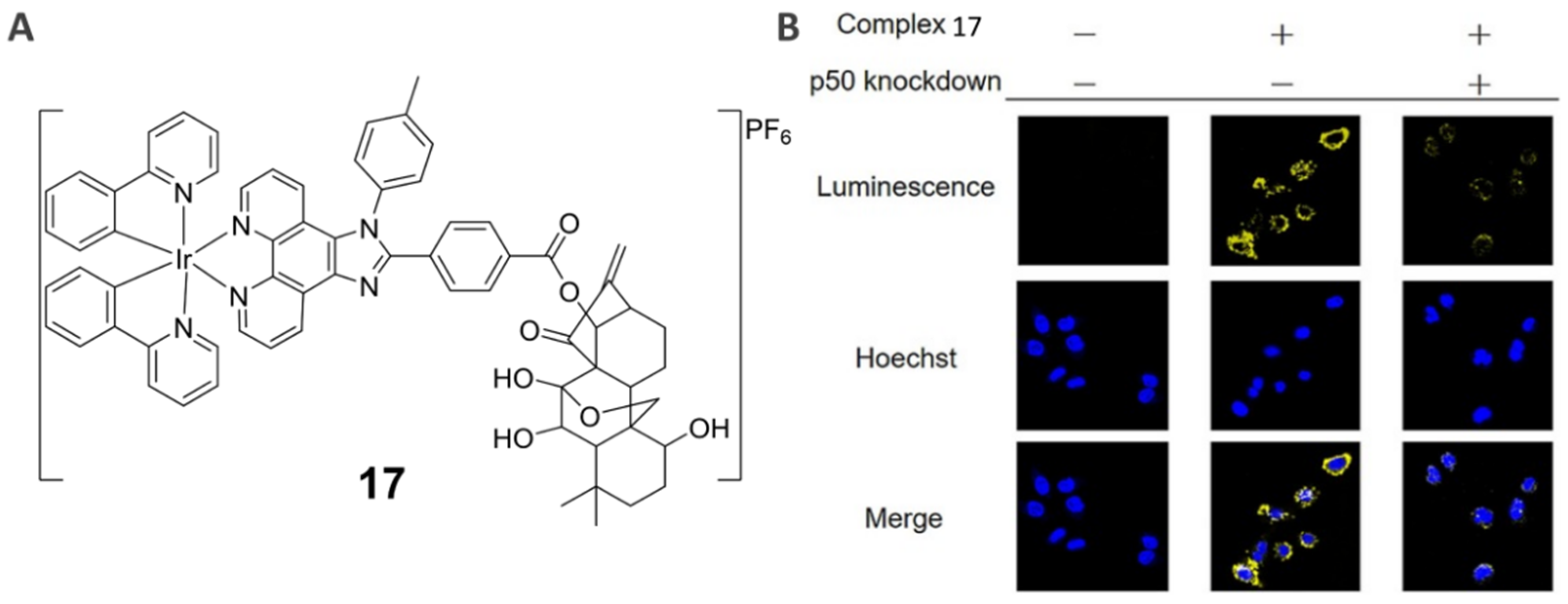
3.3. NF-κB-Specific Imaging Agent (17)
4. Other Disease-Related Protein Probes
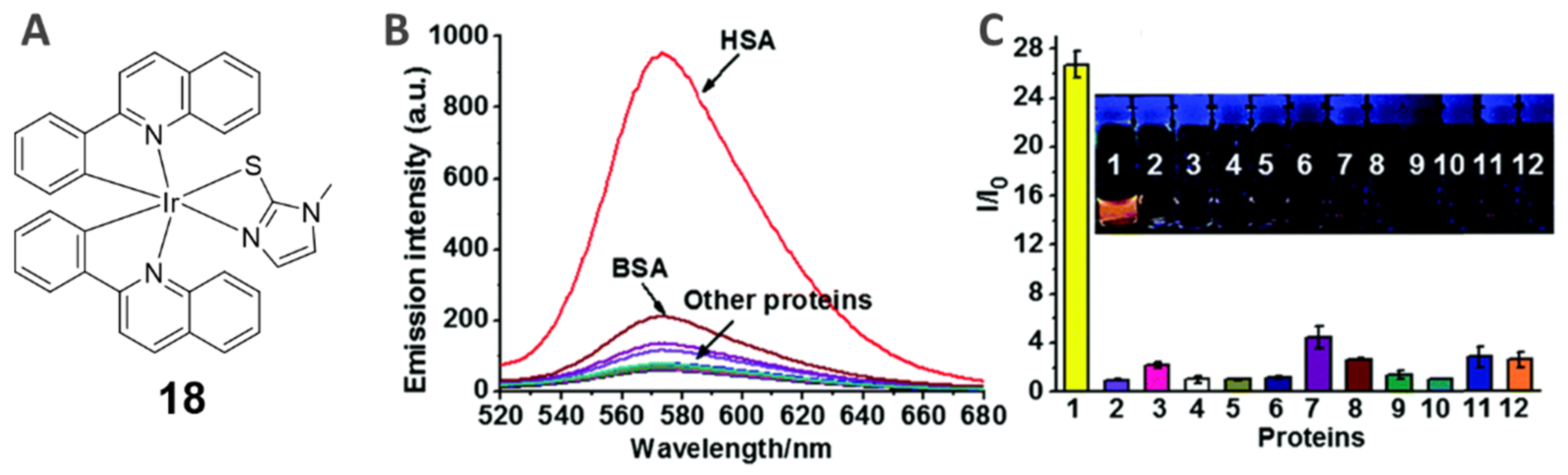
4.1. Human Serum Albumin (HSA)-Specific Staining Probes (18)
4.2. Beta-Amyloid Fibrillation-Specific Theranostic Probes (19−21)
5. Conclusions and Perspective
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Thorp, H.H. Proteins, proteins everywhere. Science 2021, 374, 1415. [Google Scholar] [CrossRef] [PubMed]
- Hipp, M.S.; Park, S.H.; Hartl, F.U. Proteostasis impairment in protein-misfolding and -aggregation diseases. Trends Cell Biol. 2014, 24, 506–514. [Google Scholar] [CrossRef] [PubMed]
- Richards, A.L.; Eckhardt, M.; Krogan, N.J. Mass spectrometry-based protein-protein interaction networks for the study of human diseases. Mol. Syst. Biol. 2021, 17, e8792. [Google Scholar] [CrossRef] [PubMed]
- Vilchez, D.; Saez, I.; Dillin, A. The role of protein clearance mechanisms in organismal ageing and age-related diseases. Nat. Commun. 2014, 5, 5659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, D.; Tsuboi, T.; Qiu, Y.; Duan, L. Recent progress in ionic iridium(III) complexes for organic electronic devices. Adv. Mater. 2017, 29, 1603253. [Google Scholar] [CrossRef]
- Williams, J.A. The coordination chemistry of dipyridylbenzene: N-deficient terpyridine or panacea for brightly luminescent metal complexes? Chem. Soc. Rev. 2009, 38, 1783–1801. [Google Scholar] [CrossRef]
- Ma, D.-L.; Lin, S.; Wang, W.; Yang, C.; Leung, C.-H. Luminescent chemosensors by using cyclometalated iridium(iii) complexes and their applications. Chem. Sci. 2017, 8, 878–889. [Google Scholar] [CrossRef] [Green Version]
- Ma, D.-L.; Wang, W.; Mao, Z.; Kang, T.-S.; Han, Q.-B.; Chan, P.W.H.; Leung, C.-H. Utilization of G-quadruplex-forming aptamers for the construction of luminescence sensing platforms. ChemPlusChem 2017, 82, 8–17. [Google Scholar] [CrossRef]
- Caporale, C.; Ranieri, A.M.; Paternoster, S.; Bader, C.A.; Falasca, M.; Plush, S.E.; Brooks, D.A.; Stagni, S.; Massi, M. Photophysical and biological properties of iridium tetrazolato complexes functionalised with fatty acid chains. Inorganics 2020, 8, 23. [Google Scholar] [CrossRef] [Green Version]
- Bokareva, O.S.; Möhle, T.; Neubauer, A.; Bokarev, S.I.; Lochbrunner, S.; Kühn, O. Chemical tuning and absorption properties of iridium photosensitizers for photocatalytic applications. Inorganics 2017, 5, 23. [Google Scholar] [CrossRef]
- Chi, Y.; Chou, P.-T. Transition-metal phosphors with cyclometalating ligands: Fundamentals and applications. Chem. Soc. Rev. 2010, 39, 638–655. [Google Scholar] [CrossRef]
- Zhu, X.; Su, Q.; Feng, W.; Li, F. Anti-Stokes shift luminescent materials for bio-applications. Chem. Soc. Rev. 2017, 46, 1025–1039. [Google Scholar] [CrossRef]
- Chelushkin, P.S.; Shakirova, J.R.; Kritchenkov, I.S.; Baigildin, V.A.; Tunik, S.P. Phosphorescent NIR emitters for biomedicine: Applications, advances and challenges. Dalton Trans. 2022, 51, 1257–1280. [Google Scholar] [CrossRef]
- Zhou, J.; Li, J.; Zhang, K.Y.; Liu, S.; Zhao, Q. Phosphorescent iridium(III) complexes as lifetime-based biological sensors for photoluminescence lifetime imaging microscopy. Coord. Chem. Rev. 2022, 453, 214334. [Google Scholar] [CrossRef]
- Wang, W.; Vellaisamy, K.; Li, G.; Wu, C.; Ko, C.-N.; Leung, C.-H.; Ma, D.-L. Development of a long-lived luminescence probe for visualizing β-galactosidase in ovarian carcinoma cells. Anal. Chem. 2017, 89, 11679–11684. [Google Scholar] [CrossRef]
- Wang, W.; Lu, L.; Wu, K.-J.; Liu, J.; Leung, C.-H.; Wong, C.-Y.; Ma, D.-L. Long-lived iridium(III) complexes as luminescent probes for the detection of periodate in living cells. Sens. Actuators B Chem. 2019, 288, 392–398. [Google Scholar] [CrossRef]
- Lo, K.K.-W.; Louie, M.-W.; Zhang, K.Y. Design of luminescent iridium(III) and rhenium(I) polypyridine complexes as in vitro and in vivo ion, molecular and biological probes. Coord. Chem. Rev. 2010, 254, 2603–2622. [Google Scholar] [CrossRef]
- Wu, C.; Wu, K.J.; Liu, J.B.; Zhou, X.M.; Leung, C.H.; Ma, D.L. A dual-functional molecular strategy for in situ suppressing and visualizing of neuraminidase in aqueous solution using iridium(iii) complexes. Chem. Commun. 2019, 55, 6353–6356. [Google Scholar] [CrossRef]
- Wang, W.; Wu, C.; Yang, C.; Li, G.; Han, Q.-B.; Li, S.; Lee, S.M.-Y.; Leung, C.-H.; Ma, D.-L. A dual-functional luminescent probe for imaging H2S in living zebrafish and discrimination hypoxic cells from normoxic cells. Sens. Actuators B Chem. 2018, 255, 1953–1959. [Google Scholar] [CrossRef]
- Wang, W.; Yung, T.-L.; Cheng, S.-S.; Chen, F.; Liu, J.-B.; Leung, C.-H.; Ma, D.-L. A long-lived luminogenic iridium(III) complex for acetylacetone detection in environmental samples. Sens. Actuators B Chem. 2020, 321, 128486. [Google Scholar] [CrossRef]
- Wang, W.; Dong, Z.-Z.; Yang, C.; Li, G.; Tse, Y.-C.; Leung, C.-H.; Ma, D.-L. An iridium(III) complex-based chemosensor for the detection of thiourea in living cells. Sens. Actuators B Chem. 2017, 251, 374–379. [Google Scholar] [CrossRef]
- Mao, Z.; Liu, J.; Kang, T.-S.; Wang, W.; Han, Q.-B.; Wang, C.-M.; Leung, C.-H.; Ma, D.-L. An Ir(III) complex chemosensor for the detection of thiols. Sci. Technol. Adv. Mater. 2016, 17, 109–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Mao, Z.; Wang, M.; Liu, L.-J.; Kwong, D.W.J.; Leung, C.-H.; Ma, D.-L. A long lifetime luminescent iridium(iii) complex chemosensor for the selective switch-on detection of Al3+ ions. Chem. Commun. 2016, 52, 3611–3614. [Google Scholar] [CrossRef] [PubMed]
- Lo, K.K.-W. Molecular design of bioorthogonal probes and imaging reagents derived from photofunctional transition metal complexes. Acc. Chem. Res. 2020, 53, 32–44. [Google Scholar] [CrossRef] [PubMed]
- Heal, W.P.; Dang, T.H.T.; Tate, E.W. Activity-based probes: Discovering new biology and new drug targets. Chem. Soc. Rev. 2011, 40, 246–257. [Google Scholar] [CrossRef]
- Fang, H.; Peng, B.; Ong, S.Y.; Wu, Q.; Li, L.; Yao, S.Q. Recent advances in activity-based probes (ABPs) and affinity-based probes (AfBPs) for profiling of enzymes. Chem. Sci. 2021, 12, 8288–8310. [Google Scholar] [CrossRef]
- Heinzlmeir, S.; Müller, S. Selectivity aspects of activity-based (chemical) probes. Drug Discov. Today 2022, 27, 519–528. [Google Scholar] [CrossRef]
- Yu, W.; Baskin, J.M. Photoaffinity labeling approaches to elucidate lipid–protein interactions. Curr. Opin. Chem. Biol. 2022, 69, 102173. [Google Scholar] [CrossRef]
- Liu, Y.; Patricelli, M.P.; Cravatt, B.F. Activity-based protein profiling: The serine hydrolases. Proc. Natl. Acad. Sci. USA. 1999, 96, 14694–14699. [Google Scholar] [CrossRef] [Green Version]
- Zuhl, A.M.; Mohr, J.T.; Bachovchin, D.A.; Niessen, S.; Hsu, K.L.; Berlin, J.M.; Dochnahl, M.; Lopez-Alberca, M.P.; Fu, G.C.; Cravatt, B.F. Competitive activity-based protein profiling identifies aza-beta-lactams as a versatile chemotype for serine hydrolase inhibition. J. Am. Chem. Soc. 2012, 134, 5068–5071. [Google Scholar] [CrossRef]
- Wang, Z.; Guo, Z.; Song, T.; Zhang, X.; He, N.; Liu, P.; Wang, P.; Zhang, Z. Proteome-wide identification of on- and off-targets of Bcl-2 inhibitors in native biological systems by using affinity-based probes (AfBPs). ChemBioChem 2018, 19, 2312–2320. [Google Scholar] [CrossRef]
- Yang, X.; Michiels, T.J.M.; de Jong, C.; Soethoudt, M.; Dekker, N.; Gordon, E.; van der Stelt, M.; Heitman, L.H.; van der Es, D.; AP, I.J. An affinity-based probe for the human adenosine A2A receptor. J. Med. Chem. 2018, 61, 7892–7901. [Google Scholar] [CrossRef] [Green Version]
- Sheinenzon, A.; Shehadeh, M.; Michelis, R.; Shaoul, E.; Ronen, O. Serum albumin levels and inflammation. Int. J. Biol. Macromol. 2021, 184, 857–862. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Li, J.; Wu, Z.; Zhang, H.; Zhao, Y.; Yang, R.; Lu, L. Curcumin ligand based iridium(III) complexes as inhibition and visualization agent of beta-amyloid fibrillation. Microchem. J. 2021, 160, 105721. [Google Scholar] [CrossRef]
- Srivastava, S.; Koay, E.J.; Borowsky, A.D.; De Marzo, A.M.; Ghosh, S.; Wagner, P.D.; Kramer, B.S. Cancer overdiagnosis: A biological challenge and clinical dilemma. Nat. Rev. Cancer 2019, 19, 349–358. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: Globocan estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Bogani, G.; Ditto, A.; Raspagliesi, F. Early diagnosis in endometrial cancer minimizes the impact of treatments. Am. J. Obstet. Gynecol. 2018, 219, 631–632. [Google Scholar] [CrossRef] [Green Version]
- Whitaker, K. Earlier diagnosis: The importance of cancer symptoms. Lancet Oncol. 2020, 21, 6–8. [Google Scholar] [CrossRef] [Green Version]
- Lo, K.K.-W.; Zhang, K.Y.; Chung, C.-K.; Kwok, K.Y. Synthesis, photophysical and electrochemical properties, and protein-binding studies of luminescent cyclometalated iridium(III) bipyridine estradiol conjugates. Chem. Eur. J. 2007, 13, 7110–7120. [Google Scholar] [CrossRef]
- Sun, T.; Liu, Z.; Yang, Q. The role of ubiquitination and deubiquitination in cancer metabolism. Mol. Cancer 2020, 19, 146. [Google Scholar] [CrossRef]
- Caon, I.; Bartolini, B.; Parnigoni, A.; Carava, E.; Moretto, P.; Viola, M.; Karousou, E.; Vigetti, D.; Passi, A. Revisiting the hallmarks of cancer: The role of hyaluronan. Semin. Cancer Biol. 2020, 62, 9–19. [Google Scholar] [CrossRef]
- Smith, W.L.; Langenbach, R. Why there are two cyclooxygenase isozymes. J. Clin. Investig. 2001, 107, 1491–1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uchida, K. HNE as an inducer of COX-2. Free Radic. Biol. Med. 2017, 111, 169–172. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.J.; Crews, B.C.; Xu, S.; Ghebreselasie, K.; Daniel, C.K.; Kingsley, P.J.; Banerjee, S.; Marnett, L.J. Antitumor activity of cytotoxic cyclooxygenase-2 Inhibitors. ACS Chem. Biol. 2016, 11, 3052–3060. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, C.; Yang, C.; Lu, L.; Wang, W.; Tan, W.; Leung, C.H.; Ma, D.L. Luminescent iridium(iii) complexes as COX-2-specific imaging agents in cancer cells. Chem. Commun. 2017, 53, 2822–2825. [Google Scholar] [CrossRef] [Green Version]
- Slack, R.J.; Macdonald, S.J.F.; Roper, J.A.; Jenkins, R.G.; Hatley, R.J.D. Emerging therapeutic opportunities for integrin inhibitors. Nat. Rev. Drug Discov. 2022, 21, 60–78. [Google Scholar] [CrossRef]
- Ma, X.; Jia, J.; Cao, R.; Wang, X.; Fei, H. Histidine-iridium(III) coordination-based peptide luminogenic cyclization and cyclo-RGD peptides for cancer-cell targeting. J. Am. Chem. Soc. 2014, 136, 17734–17737. [Google Scholar] [CrossRef]
- Cancilla, D.; Rettig, M.P.; DiPersio, J.F. Targeting CXCR4 in AML and ALL. Front. Oncol. 2020, 10, 1672. [Google Scholar] [CrossRef]
- Bhandari, D.; Robia, S.L.; Marchese, A. The E3 ubiquitin ligase atrophin interacting protein 4 binds directly to the chemokine receptor CXCR4 via a novel WW domain-mediated interaction. Mol. Biol. Cell 2009, 20, 1324–1339. [Google Scholar] [CrossRef]
- Furusato, B.; Mohamed, A.; Uhlen, M.; Rhim, J.S. CXCR4 and cancer. Pathol. Int. 2010, 60, 497–505. [Google Scholar] [CrossRef]
- Kang, H.; Watkins, G.; Douglas-Jones, A.; Mansel, R.E.; Jiang, W.G. The elevated level of CXCR4 is correlated with nodal metastasis of human breast cancer. Breast 2005, 14, 360–367. [Google Scholar] [CrossRef]
- Kuil, J.; Steunenberg, P.; Chin, P.T.; Oldenburg, J.; Jalink, K.; Velders, A.H.; van Leeuwen, F.W. Peptide-functionalized luminescent iridium complexes for lifetime imaging of CXCR4 expression. ChemBioChem 2011, 12, 1897–1903. [Google Scholar] [CrossRef]
- Nayak, R.T.; Hong, H.; Zhang, Y.; Cai, W. Multimodality imaging of CXCR4 in cancer: Current status towards clinical translation. Curr. Mol. Med. 2013, 13, 1538–1548. [Google Scholar] [CrossRef]
- Chen, J.; Zeng, F.; Forrester, S.J.; Eguchi, S.; Zhang, M.Z.; Harris, R.C. Expression and function of the epidermal growth factor receptor in physiology and disease. Plant Physiol. 2016, 96, 1025–1069. [Google Scholar] [CrossRef]
- Li, Z.; Tyrpak, D.R.; Park, M.; Okamoto, C.T.; MacKay, J.A. A new temperature-dependent strategy to modulate the epidermal growth factor receptor. Biomaterials 2018, 183, 319–330. [Google Scholar] [CrossRef]
- Gazit, A.; Yaish, P.; Gilon, C.; Levitzki, A. Tyrphostins I: Synthesis and biological activity of protein tyrosine kinase inhibitors. J. Med. Chem. 1989, 32, 2344–2352. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.; Wu, K.-J.; Liu, J.-B.; Wang, W.; Leung, C.-H.; Ma, D.-L. Structure-guided discovery of a luminescent theranostic toolkit for living cancer cells and the imaging behavior effect. Chem. Sci. 2020, 11, 11404–11412. [Google Scholar] [CrossRef]
- Baratto, L.; Duan, H.; Macke, H.; Iagaru, A. Imaging the distribution of gastrin-releasing peptide receptors in cancer. J. Nucl. Med. 2020, 61, 792–798. [Google Scholar] [CrossRef]
- Constantinides, C.; Lazaris, A.C.; Haritopoulos, K.N.; Pantazopoulos, D.; Chrisofos, M.; Giannopoulos, A. Immunohistochemical detection of gastrin releasing peptide in patients with prostate cancer. World J. Urol. 2003, 21, 183–187. [Google Scholar] [CrossRef]
- Sano, H.; Feighner, S.D.; Hreniuk, D.L.; Iwaasa, H.; Sailer, A.W.; Pan, J.; Reitman, M.L.; Kanatani, A.; Howard, A.D.; Tan, C.P. Characterization of the bombesin-like peptide receptor family in primates. Genomics 2004, 84, 139–346. [Google Scholar] [CrossRef]
- Wang, W.; Wu, K.J.; Vellaisamy, K.; Leung, C.H.; Ma, D.L. Peptide-conjugated long-lived theranostic imaging for targeting GRPr in cancer and immune cells. Angew. Chem. Int. Ed. 2020, 59, 17897–17902. [Google Scholar] [CrossRef] [PubMed]
- Sobczuk, P.; Lomiak, M.; Cudnoch-Jedrzejewska, A. Dopamine D1 receptor in cancer. Cancers 2020, 12, 3232. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, V.V.; Madhukar, N.S.; Gilvary, C.; Kline, C.L.B.; Oster, S.; El-Deiry, W.S.; Elemento, O.; Doherty, F.; VanEngelenburg, A.; Durrant, J.; et al. Dopamine receptor D5 is a modulator of tumor response to dopamine receptor D2 antagonism. Clin. Cancer Res. 2019, 25, 2305–2313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butini, S.; Nikolic, K.; Kassel, S.; Bruckmann, H.; Filipic, S.; Agbaba, D.; Gemma, S.; Brogi, S.; Brindisi, M.; Campiani, G.; et al. Polypharmacology of dopamine receptor ligands. Prog. Neurobiol. 2016, 142, 68–103. [Google Scholar] [CrossRef] [PubMed]
- Vellaisamy, K.; Li, G.; Ko, C.N.; Zhong, H.J.; Fatima, S.; Kwan, H.Y.; Wong, C.Y.; Kwong, W.J.; Tan, W.; Leung, C.H.; et al. Cell imaging of dopamine receptor using agonist labeling iridium(iii) complex. Chem. Sci. 2018, 9, 1119–1125. [Google Scholar] [CrossRef] [Green Version]
- Burke, P.J. Mitochondria, bioenergetics and apoptosis in cancer. Trends Cancer 2017, 3, 857–870. [Google Scholar] [CrossRef]
- Nagata, S. Apoptosis and clearance of apoptotic cells. Annu. Rev. Immunol. 2018, 36, 489–517. [Google Scholar] [CrossRef]
- Tang, D.; Lotze, M.T.; Kang, R.; Zeh, H.J. Apoptosis promotes early tumorigenesis. Oncogene 2011, 30, 1851–1854. [Google Scholar] [CrossRef] [Green Version]
- Vunnam, N.; Szymonski, S.; Hirsova, P.; Gores, G.J.; Sachs, J.N.; Hackel, B.J. Noncompetitive allosteric antagonism of death receptor 5 by a synthetic affibody ligand. Biochemistry 2020, 59, 3856–3868. [Google Scholar] [CrossRef]
- Zhang, B.; van Roosmalen, I.A.M.; Reis, C.R.; Setroikromo, R.; Quax, W.J. Death receptor 5 is activated by fucosylation in colon cancer cells. FEBS J. 2019, 286, 555–571. [Google Scholar] [CrossRef]
- Masum, A.A.; Yokoi, K.; Hisamatsu, Y.; Naito, K.; Shashni, B.; Aoki, S. Design and synthesis of a luminescent iridium complex-peptide hybrid (IPH) that detects cancer cells and induces their apoptosis. Bioorg. Med. Chem. 2018, 26, 4804–4816. [Google Scholar] [CrossRef]
- Lesch, H.P.; Kaikkonen, M.U.; Pikkarainen, J.T.; Ylä-Herttuala, S. Avidin-biotin technology in targeted therapy. Expert Opin. Drug Deliv. 2010, 7, 551–564. [Google Scholar] [CrossRef]
- Luong, J.H.T.; Male, K.B.; Glennon, J.D. Biotin interference in immunoassays based on biotin-strept(avidin) chemistry: An emerging threat. Biotechnol. Adv. 2019, 37, 634–641. [Google Scholar] [CrossRef]
- Lo, K.K.-W.; Chan, J.S.-W.; Lui, L.-H.; Chung, C.-K. Novel luminescent cyclometalated iridium(III) diimine complexes that contain a biotin moiety. Organometallics 2004, 23, 3108–3116. [Google Scholar] [CrossRef]
- Lo, K.K.-W.; Li, C.-K.; Lau, J.S.-Y. Luminescent cyclometalated iridium(III) arylbenzothiazole biotin complexes. Organometallics 2005, 24, 4594–4601. [Google Scholar] [CrossRef]
- Lo, K.K.-W.; Chung, C.-K.; Zhu, N. Nucleic acid intercalators and avidin probes derived from luminescent cyclometalated iridium(III)–dipyridoquinoxaline and –dipyridophenazine complexes. Chem. Eur. J. 2006, 12, 1500–1512. [Google Scholar] [CrossRef]
- Lo, K.K.-W.; Lau, J.S.-Y. Cyclometalated iridium(III) diimine bis(biotin) complexes as the first luminescent biotin-based cross-linkers for avidin. Inorg. Chem. 2007, 46, 700–709. [Google Scholar] [CrossRef]
- Zhang, K.Y.; Lo, K.K.-W. Synthesis, properties, and live-cell imaging studies of luminescent cyclometalated iridium(III) polypyridine complexes containing two or three biotin pendants. Inorg. Chem. 2009, 48, 6011–6025. [Google Scholar] [CrossRef]
- Zhang, K.Y.; Liu, H.-W.; Tang, M.-C.; Choi, A.W.-T.; Zhu, N.; Wei, X.-G.; Lau, K.-C.; Lo, K.K.-W. Dual-emissive cyclometalated iridium(III) polypyridine complexes as ratiometric biological probes and organelle-selective bioimaging reagents. Inorg. Chem. 2015, 54, 6582–6593. [Google Scholar] [CrossRef]
- Kwon, T.-H.; Kwon, J.; Hong, J.-I. Signal amplification via intramolecular energy transfer using tripodal neutral iridium(III) complexes upon binding to avidin. J. Am. Chem. Soc. 2008, 130, 3726–3727. [Google Scholar] [CrossRef]
- Raabe, C.A.; Groper, J.; Rescher, U. Biased perspectives on formyl peptide receptors. BBA-Mol. Cell Res. 2019, 1866, 305–316. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, Y.; Han, J.; Zhu, Z.; Li, M.; Liu, Q.; Wang, Y.; Shi, F.-D. Formyl peptide receptor 1 signaling potentiates inflammatory brain injury. Sci. Transl. Med. 2021, 13, eabe9890. [Google Scholar] [CrossRef] [PubMed]
- Vellaisamy, K.; Li, G.; Wang, W.; Leung, C.H.; Ma, D.L. A long-lived peptide-conjugated iridium(iii) complex as a luminescent probe and inhibitor of the cell migration mediator, formyl peptide receptor 2. Chem. Sci. 2018, 9, 8171–8177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, H.; Guo, X.; Wang, Z.; Han, M.; Liu, H. Therapeutic potential of WKYMVm in diseases. Front. Pharmacol. 2022, 13, 986963. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-kappaB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Imbert, V.; Peyron, J.F. NF-kappaB in hematological malignancies. Biomedicines 2017, 5, 27. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Yang, C.; Lin, S.; Vellaisamy, K.; Li, G.; Tan, W.; Leung, C.H.; Ma, D.L. First synthesis of an oridonin-conjugated iridium(III) complex for the intracellular tracking of NF-kappaB in living cells. Chem. Eur. J. 2017, 23, 4929–4935. [Google Scholar] [CrossRef]
- China, L.; Skene, S.S.; Shabir, Z.; Maini, A.; Sylvestre, Y.; Bennett, K.; Bevan, S.; O’Beirne, J.; Forrest, E.; Portal, J.; et al. Administration of albumin solution increases serum levels of albumin in patients with chronic liver failure in a single-arm feasibility trial. Clin. Gastroenterol. Hepatol. 2018, 16, 748–755.e6. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Huang, H.; Chen, G.; Chen, H.; Xu, T.; Tang, Q.; Zhu, H.; Zhang, Q.; Zhang, P. A novel iridium(iii) complex for sensitive HSA phosphorescence staining in proteome research. Chem. Commun. 2018, 54, 3282–3285. [Google Scholar] [CrossRef]
- Panza, F.; Lozupone, M.; Logroscino, G.; Imbimbo, B.P. A critical appraisal of amyloid-β-targeting therapies for Alzheimer disease. Nat. Rev. Neurol. 2019, 15, 73–88. [Google Scholar] [CrossRef]
- Han, J.; Du, Z.; Lim, M.H. Mechanistic insight into the design of chemical tools to control multiple pathogenic features in Alzheimer’s disease. Acc. Chem. Res. 2021, 54, 3930–3940. [Google Scholar] [CrossRef]
- Fusar-Poli, L.; Vozza, L.; Gabbiadini, A.; Vanella, A.; Concas, I.; Tinacci, S.; Petralia, A.; Signorelli, M.S.; Aguglia, E. Curcumin for depression: A meta-analysis. Crit. Rev. Food Sci. Nutr. 2020, 60, 2643–2653. [Google Scholar] [CrossRef]
- Banik, U.; Parasuraman, S.; Adhikary, A.K.; Othman, N.H. Curcumin: The spicy modulator of breast carcinogenesis. J. Exp. Clin. Canc. Res. 2017, 36, 98. [Google Scholar] [CrossRef] [Green Version]
- McGhie, B.S.; Aldrich-Wright, J.R. Photoactive and luminescent transition metal complexes as anticancer agents: A guiding light in the search for new and improved cancer treatments. Biomedicines 2022, 10, 578. [Google Scholar] [CrossRef]
- Ma, D.-L.; Wu, C.; Cheng, S.-S.; Lee, F.-W.; Han, Q.-B.; Leung, C.-H. Development of natural product-conjugated metal complexes as cancer therapies. Int. J. Mol. Sci. 2019, 20, 341. [Google Scholar] [CrossRef] [Green Version]
- Poursharifi, M.; Wlodarczyk, M.T.; Mieszawska, A.J. Nano-based systems and biomacromolecules as carriers for metallodrugs in anticancer therapy. Inorganics 2019, 7, 2. [Google Scholar] [CrossRef]














Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Liu, J.; Nao, S.-C.; Ma, D.-L.; Wang, J.; Leung, C.-H. Affinity-Based Luminescent Iridium(III) Complexes for the Detection of Disease-Related Proteins. Inorganics 2022, 10, 178. https://doi.org/10.3390/inorganics10110178
Wang W, Liu J, Nao S-C, Ma D-L, Wang J, Leung C-H. Affinity-Based Luminescent Iridium(III) Complexes for the Detection of Disease-Related Proteins. Inorganics. 2022; 10(11):178. https://doi.org/10.3390/inorganics10110178
Chicago/Turabian StyleWang, Wanhe, Jianhua Liu, Sang-Cuo Nao, Dik-Lung Ma, Jing Wang, and Chung-Hang Leung. 2022. "Affinity-Based Luminescent Iridium(III) Complexes for the Detection of Disease-Related Proteins" Inorganics 10, no. 11: 178. https://doi.org/10.3390/inorganics10110178
APA StyleWang, W., Liu, J., Nao, S. -C., Ma, D. -L., Wang, J., & Leung, C. -H. (2022). Affinity-Based Luminescent Iridium(III) Complexes for the Detection of Disease-Related Proteins. Inorganics, 10(11), 178. https://doi.org/10.3390/inorganics10110178