Mechanism of Iron–Sulfur Cluster Assembly: In the Intimacy of Iron and Sulfur Encounter


Abstract
:1. Introduction
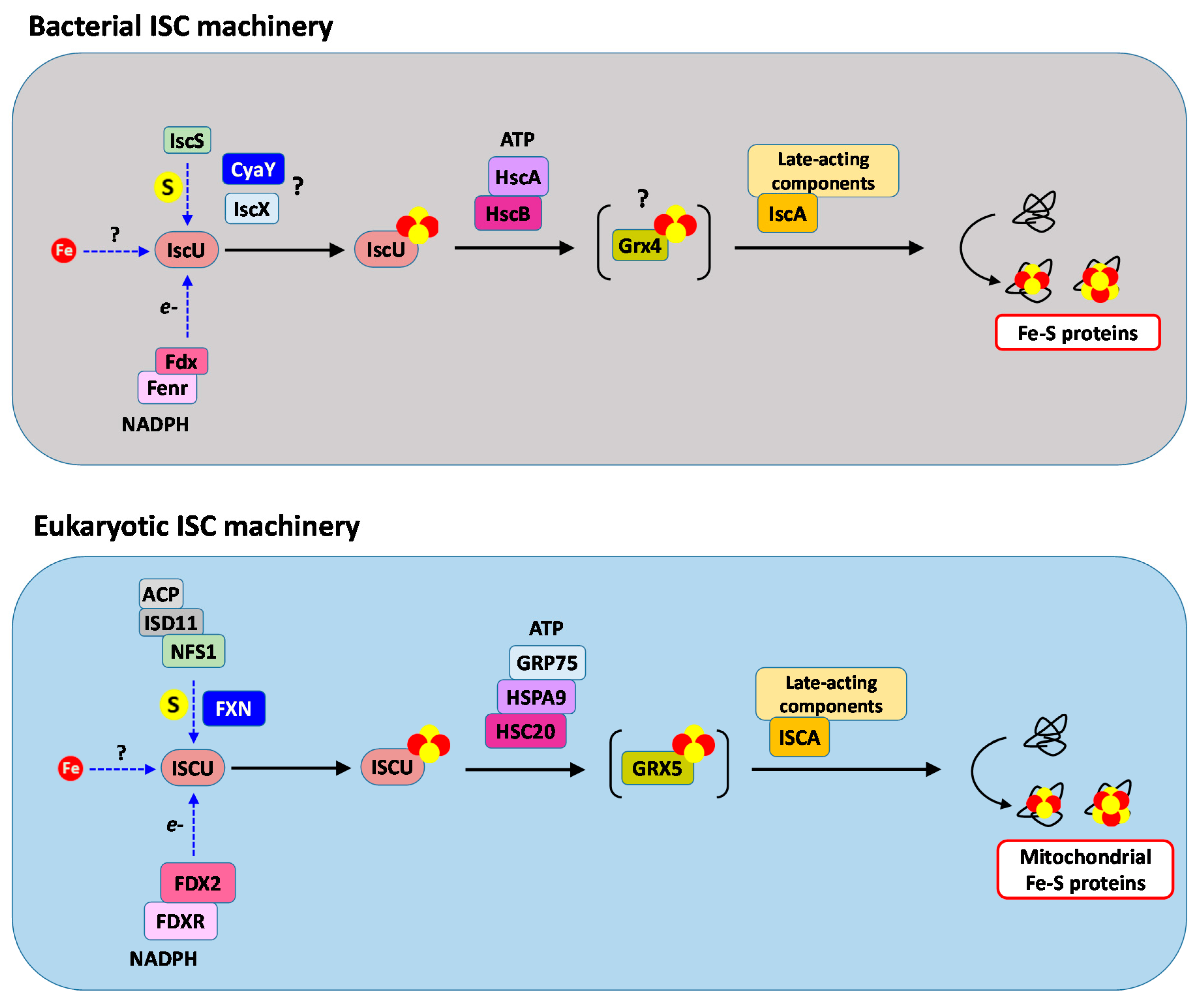
2. Overview of the Fe–S Cluster Assembly Machineries
3. Mechanism of Assembly by the ISC Core Machinery
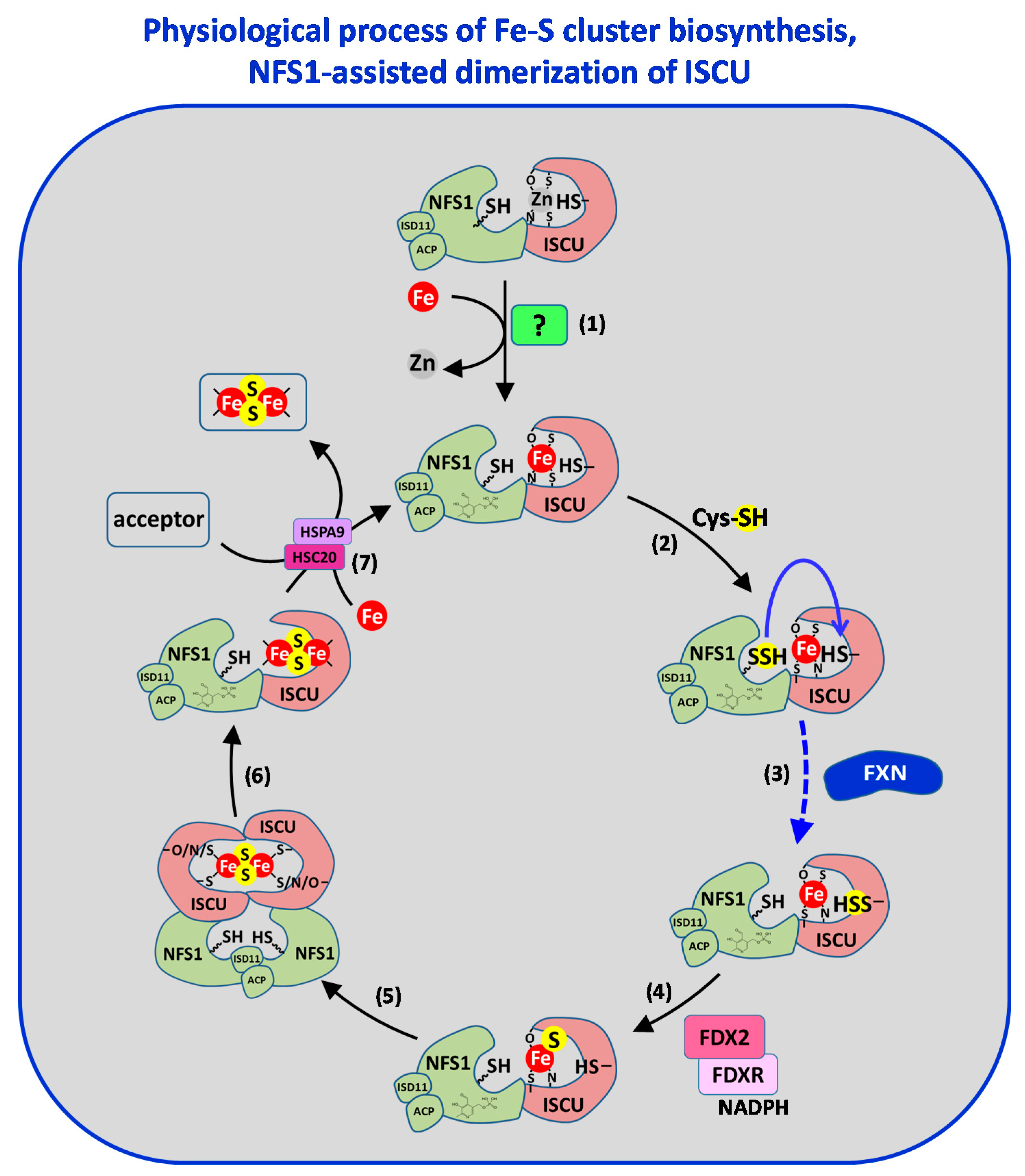
3.1. Step 1: Iron Insertion
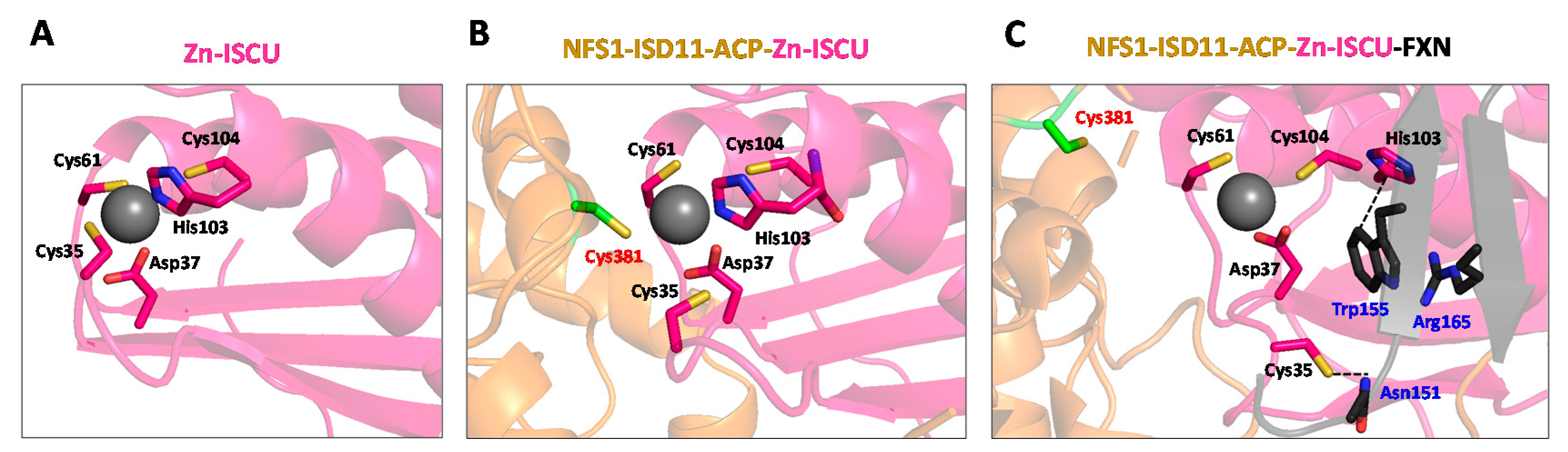
3.1.1. A Mononuclear Ferrous Iron-Binding Site in IscU/ISCU
3.1.2. Iron Donor to IscU/ISCU
Frataxin
ISCA
Connection with the Labile Iron Pool
3.2. Step 2: Sulfur Insertion
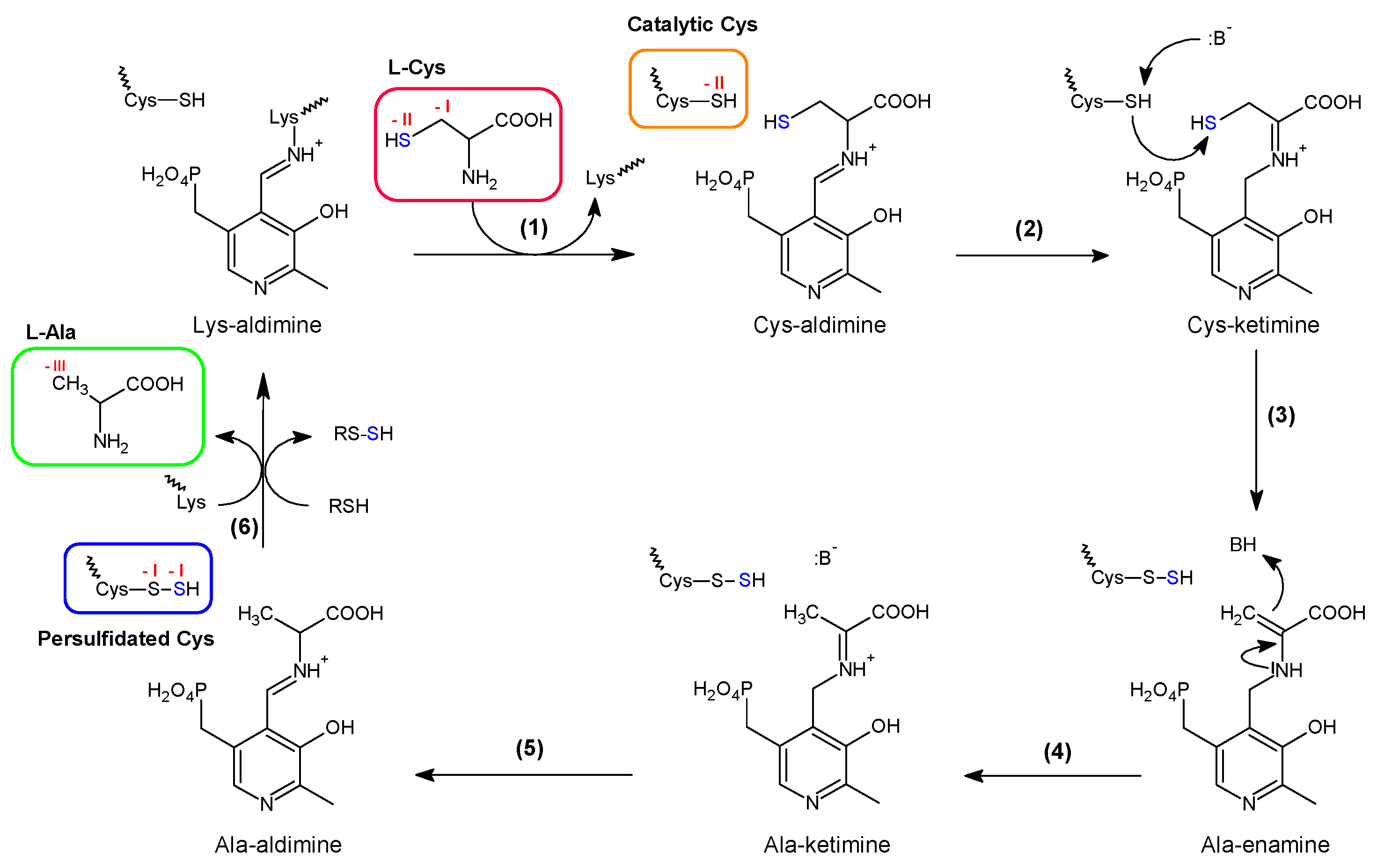
3.2.1. Two Different Classes of Cysteine Desulfurase
3.2.2. Mechanism Generating Persulfide in Class I Cysteine Desulfurase
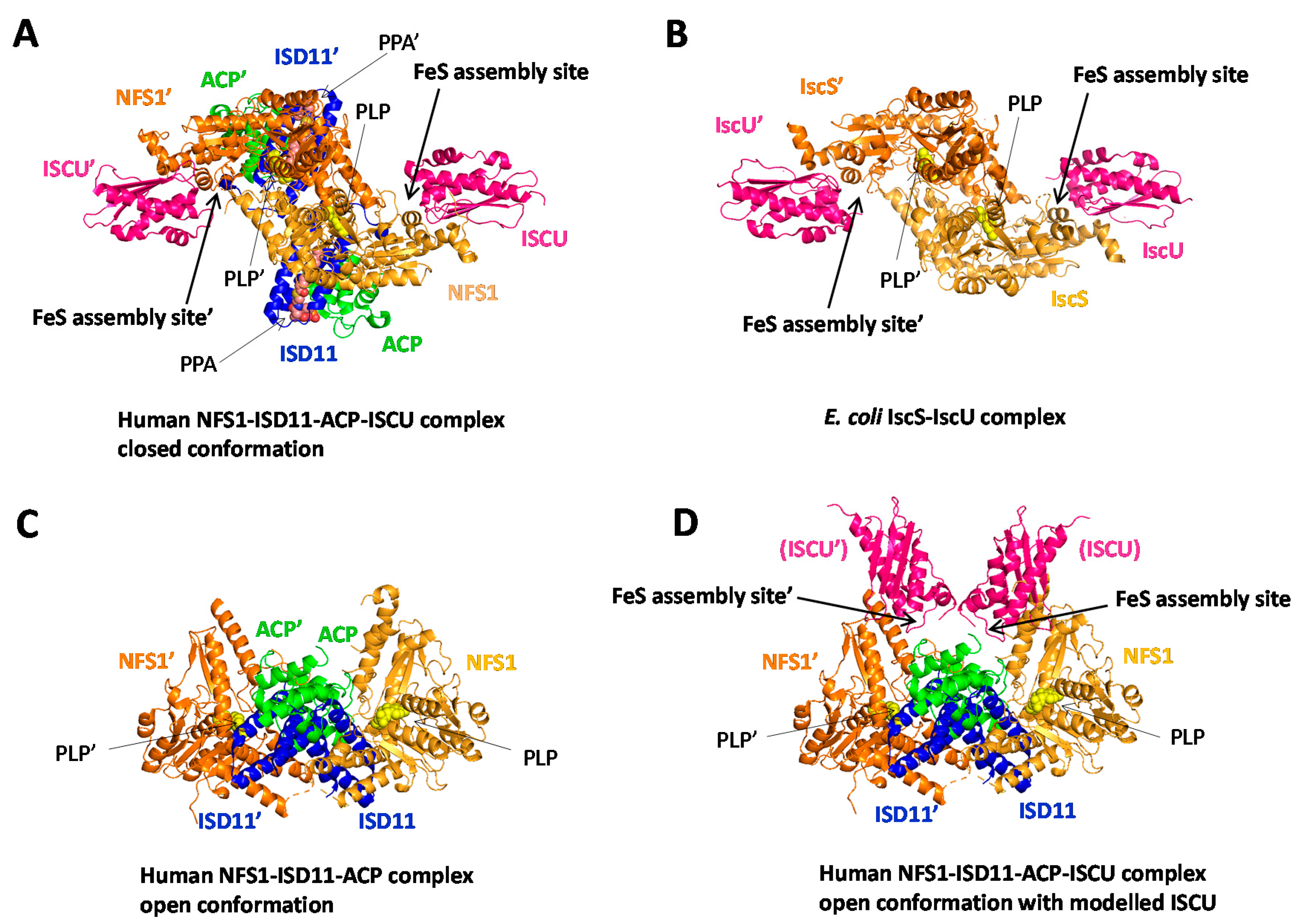
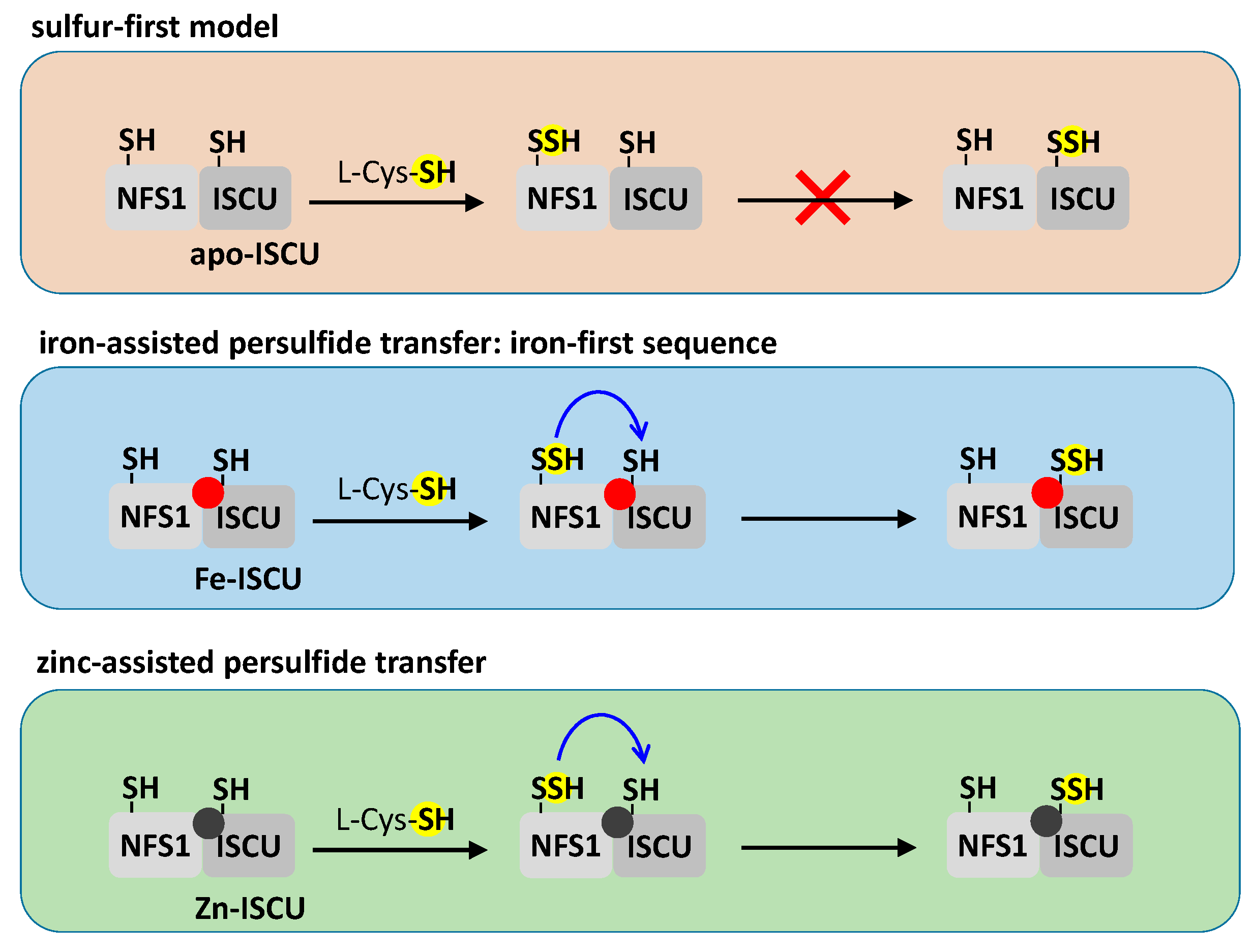
3.2.3. Sulfur Transfer to the IscU/ISCU Scaffold, a Metal-Driven Process
3.3. Step 3: Persulfide Reduction
3.3.1. Reduction by Iron
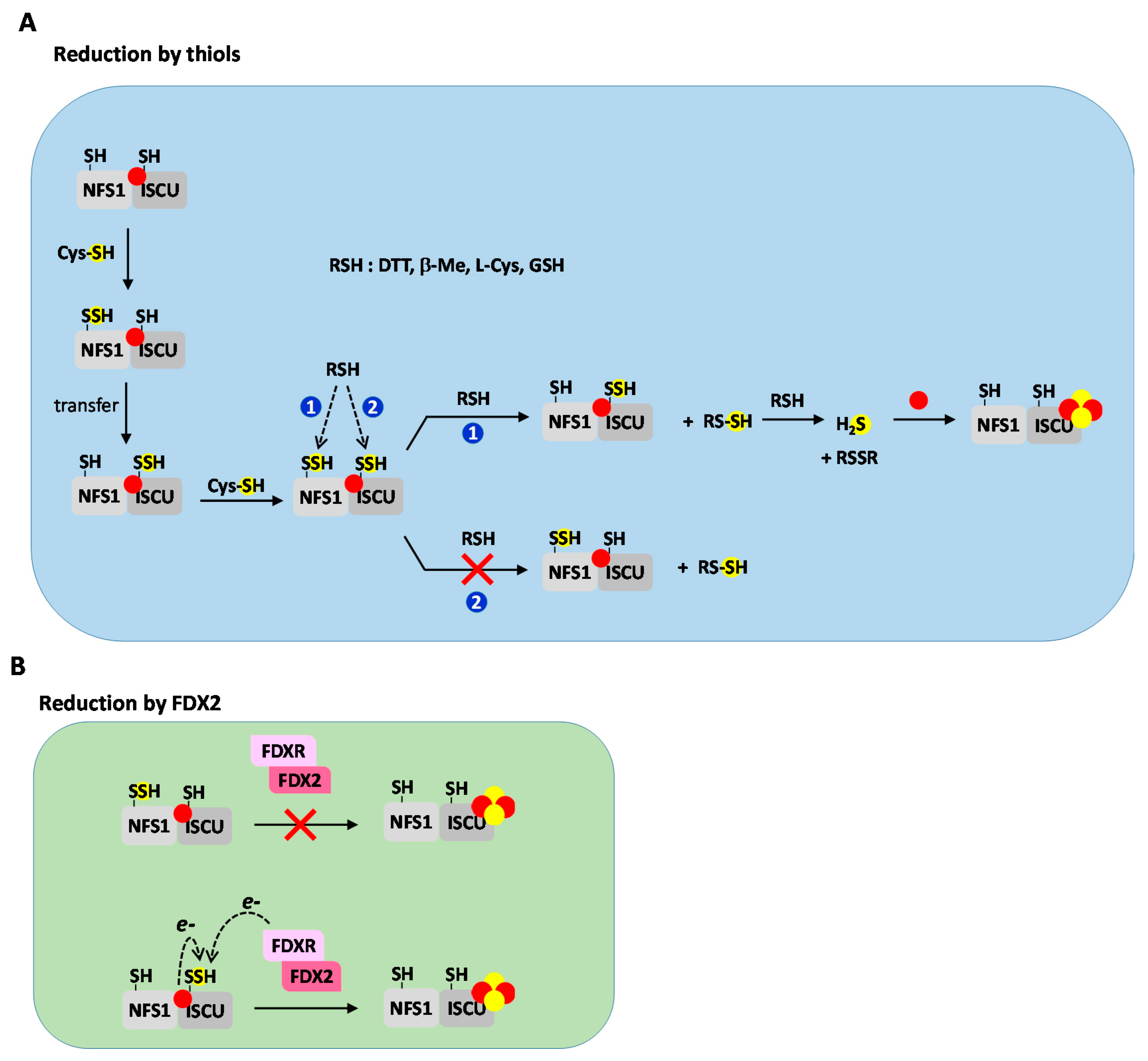
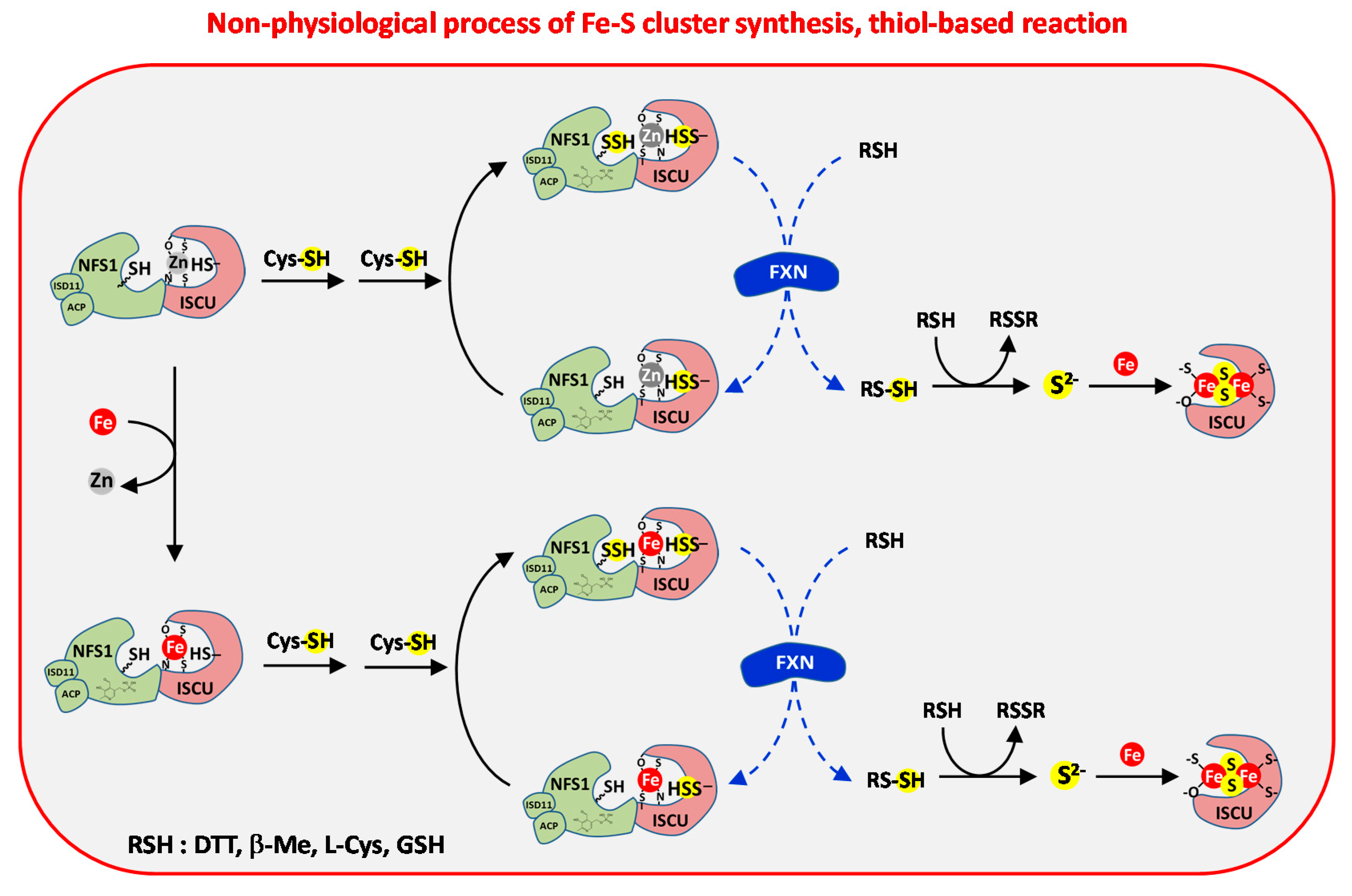
3.3.2. Reduction by Thiols
3.3.3. Ferredoxin and Iron Mediated Reduction of Persulfides
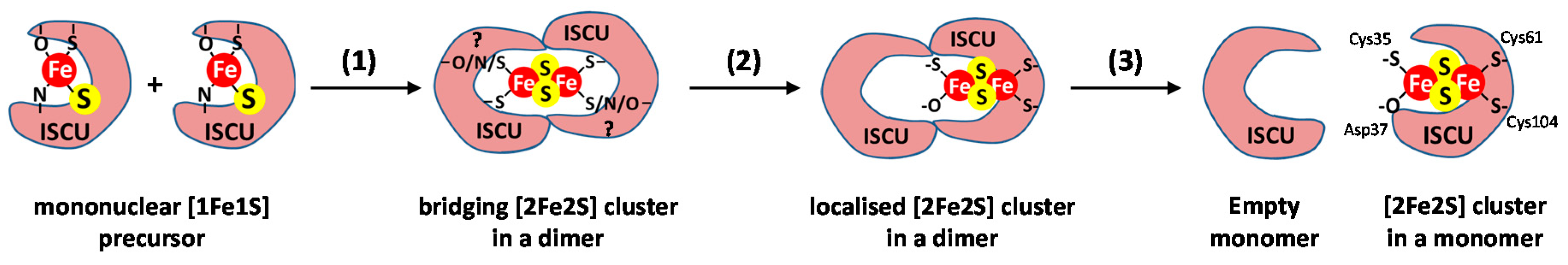
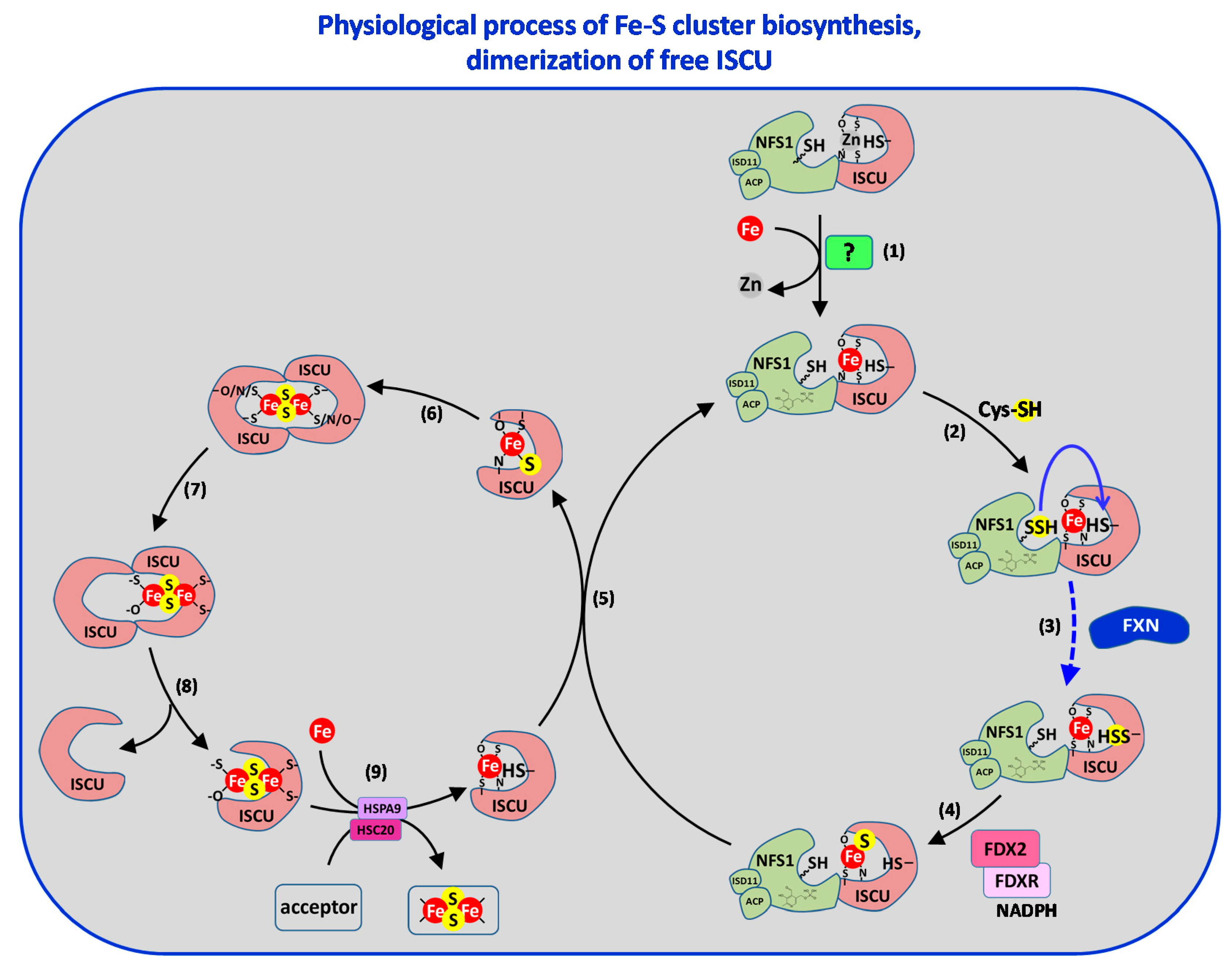
3.4. Step 4: Fe–S Cluster Formation
3.5. Regulation of Fe–S Cluster Assembly by Frataxin
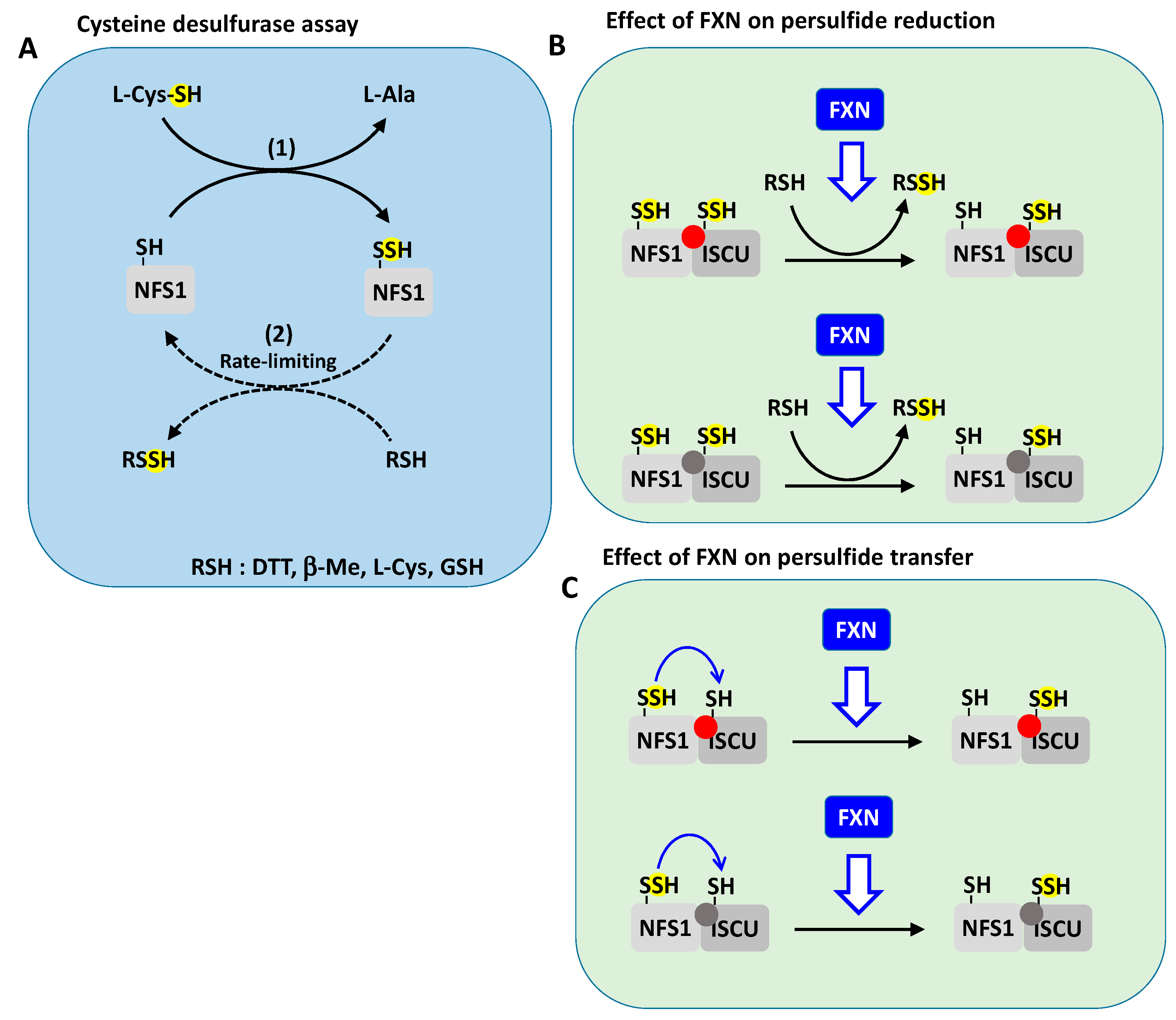
3.5.1. Effect of Frataxin on Cysteine Desulfurase Activity: The Key Question of the Rate-Limiting Step
3.5.2. Effect of Frataxin on Persulfide Transfer
3.6. Toward a Model of [2Fe2S] Cluster Biosynthesis by the ISC Machinery
4. Fe–S Assembly by the NIF Machinery
4.1. Iron Insertion in NifU
4.2. Still Elusive Steps Leading to Formation of the [2Fe2S] and [4Fe4S] Clusters
5. Fe–S Assembly by the SUF Machinery
5.1. Overall Description of the SUF Machinery
5.2. Iron Insertion in the SufBC2D Scaffold
5.3. Sulfur Insertion in the SufBC2D Complex
5.3.1. SufS, the Source of Sulfur
5.3.2. Sulfur Transfer via SufE
5.3.3. Sulfur Transfer via SufU
5.4. Persulfide Transfer to SufBC2D and Reduction via FADH2?
6. Concluding Remarks
Author Contributions
Funding
Conflicts of Interest
References
- Camprubi, E.; Jordan, S.F.; Vasiliadou, R.; Lane, N. Iron catalysis at the origin of life. IUBMB Life 2017, 69, 373–381. [Google Scholar] [CrossRef]
- Huber, C.; Wachtershauser, G. Alpha-Hydroxy and alpha-amino acids under possible Hadean, volcanic origin-of-life conditions. Science 2006, 314, 630–632. [Google Scholar] [CrossRef]
- Cody, G. Transition metal sulfides and the origin of metabolism. Annu. Rev. Earth Planet. Sci. 2004, 32, 569–599. [Google Scholar] [CrossRef]
- Martin, W.; Russell, M.J. On the origins of cells: A hypothesis for the evolutionary transitions from abiotic geochemistry to chemoautotrophic prokaryotes, and from prokaryotes to nucleated cells. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2003, 358, 59–83, discussion 55–83. [Google Scholar] [CrossRef] [Green Version]
- Chahal, H.K.; Boyd, J.M.; Outten, F.W. Iron–Sulfur Cluster Biogenesis in Archaea and Bacteria. In Encyclopedia of Inorganic and Bioinorganic Chemistry; Scott, R.A., Ed.; Wiley: Hoboken, NJ, USA, 2013. [Google Scholar] [CrossRef]
- Lill, R.; Freibert, S.A. Mechanisms of Mitochondrial Iron-Sulfur Protein Biogenesis. Annu. Rev. Biochem. 2020, 89, 471–499. [Google Scholar] [CrossRef]
- Paul, V.D.; Lill, R. Biogenesis of cytosolic and nuclear iron-sulfur proteins and their role in genome stability. Biochim. Biophys. Acta 2015, 1853, 1528–1539. [Google Scholar] [CrossRef] [Green Version]
- Przybyla-Toscano, J.; Roland, M.; Gaymard, F.; Couturier, J.; Rouhier, N. Roles and maturation of iron-sulfur proteins in plastids. J. Biol. Inorg. Chem. 2018, 23, 545–566. [Google Scholar] [CrossRef] [Green Version]
- Roche, B.; Aussel, L.; Ezraty, B.; Mandin, P.; Py, B.; Barras, F. Iron/sulfur proteins biogenesis in prokaryotes: Formation, regulation and diversity. Biochim. Biophys. Acta 2013, 1827, 455–469. [Google Scholar] [CrossRef] [Green Version]
- Martin, D.; Charpilienne, A.; Parent, A.; Boussac, A.; D’Autreaux, B.; Poupon, J.; Poncet, D. The rotavirus nonstructural protein NSP5 coordinates a [2Fe–2S] iron-sulfur cluster that modulates interaction to RNA. FASEB J. 2013, 27, 1074–1083. [Google Scholar] [CrossRef]
- Tam, W.; Pell, L.G.; Bona, D.; Tsai, A.; Dai, X.X.; Edwards, A.M.; Hendrix, R.W.; Maxwell, K.L.; Davidson, A.R. Tail tip proteins related to bacteriophage lambda gpL coordinate an iron-sulfur cluster. J. Mol. Biol. 2013, 425, 2450–2462. [Google Scholar] [CrossRef] [Green Version]
- Tsang, S.H.; Wang, R.; Nakamaru-Ogiso, E.; Knight, S.A.; Buck, C.B.; You, J. The Oncogenic Small Tumor Antigen of Merkel Cell Polyomavirus Is an Iron-Sulfur Cluster Protein That Enhances Viral DNA Replication. J. Virol. 2016, 90, 1544–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estellon, J.; Ollagnier de Choudens, S.; Smadja, M.; Fontecave, M.; Vandenbrouck, Y. An integrative computational model for large-scale identification of metalloproteins in microbial genomes: A focus on iron-sulfur cluster proteins. Metallomics 2014, 6, 1913–1930. [Google Scholar] [CrossRef]
- Liu, J.; Chakraborty, S.; Hosseinzadeh, P.; Yu, Y.; Tian, S.; Petrik, I.; Bhagi, A.; Lu, Y. Metalloproteins containing cytochrome, iron-sulfur, or copper redox centers. Chem. Rev. 2014, 114, 4366–4469. [Google Scholar] [CrossRef] [PubMed]
- Stegmaier, K.; Blinn, C.M.; Bechtel, D.F.; Greth, C.; Auerbach, H.; Muller, C.S.; Jakob, V.; Reijerse, E.J.; Netz, D.J.A.; Schunemann, V.; et al. Apd1 and Aim32 Are Prototypes of Bishistidinyl-Coordinated Non-Rieske [2Fe–2S] Proteins. J. Am. Chem. Soc. 2019, 141, 5753–5765. [Google Scholar] [CrossRef] [Green Version]
- Berkovitch, F.; Nicolet, Y.; Wan, J.T.; Jarrett, J.T.; Drennan, C.L. Crystal Structure of Biotin Synthase, an S-Adenosylmethionine-Dependent Radical Enzyme. Science 2004, 303, 76. [Google Scholar] [CrossRef] [Green Version]
- Bak, D.W.; Elliott, S.J. Alternative FeS cluster ligands: Tuning redox potentials and chemistry. Curr. Opinion Chem. Biol. 2014, 19, 50–58. [Google Scholar] [CrossRef]
- Hu, Y.; Ribbe, M.W. Biosynthesis of the Metalloclusters of Nitrogenases. Annu. Rev. Biochem. 2016, 85, 455–483. [Google Scholar] [CrossRef] [PubMed]
- Dobbek, H.; Svetlitchnyi, V.; Gremer, L.; Huber, R.; Meyer, O. Crystal Structure of a Carbon Monoxide Dehydrogenase Reveals a [Ni–4Fe–5S] Cluster. Science 2001, 293, 1281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, S.J.; Garner, C.D.; Hagen, W.R.; Lindley, P.F.; Bailey, S. Hybrid-cluster protein (HCP) from Desulfovibrio vulgaris (Hildenborough) at 1.6 A resolution. Biochemistry 2000, 39, 15044–15054. [Google Scholar] [CrossRef] [PubMed]
- Beinert, H. Iron-sulfur proteins: Ancient structures, still full of surprises. J. Biol. Inorg. Chem. 2000, 5, 2–15. [Google Scholar] [CrossRef]
- Blanc, B.; Gerez, C.; Ollagnier de Choudens, S. Assembly of Fe/S proteins in bacterial systems: Biochemistry of the bacterial ISC system. Biochim. Biophys. Acta 2015, 1853, 1436–1447. [Google Scholar] [CrossRef] [PubMed]
- Gervason, S.; Larkem, D.; Mansour, A.B.; Botzanowski, T.; Muller, C.S.; Pecqueur, L.; Le Pavec, G.; Delaunay-Moisan, A.; Brun, O.; Agramunt, J.; et al. Physiologically relevant reconstitution of iron-sulfur cluster biosynthesis uncovers persulfide-processing functions of ferredoxin-2 and frataxin. Nat. Commun. 2019, 10, 3566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webert, H.; Freibert, S.A.; Gallo, A.; Heidenreich, T.; Linne, U.; Amlacher, S.; Hurt, E.; Muhlenhoff, U.; Banci, L.; Lill, R. Functional reconstitution of mitochondrial Fe/S cluster synthesis on Isu1 reveals the involvement of ferredoxin. Nat. Commun. 2014, 5, 5013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciofi-Baffoni, S.; Nasta, V.; Banci, L. Protein networks in the maturation of human iron–sulfur proteins. Metallomics 2018, 10, 49–72. [Google Scholar] [CrossRef]
- Pérard, J.; Ollagnier de Choudens, S. Iron–sulfur clusters biogenesis by the SUF machinery: Close to the molecular mechanism understanding. JBIC J. Biol. Inorg. Chem. 2018, 23, 581–596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouault, T.A.; Maio, N. Biogenesis and functions of mammalian iron-sulfur proteins in the regulation of iron homeostasis and pivotal metabolic pathways. J. Biol. Chem. 2017, 292, 12744–12753. [Google Scholar] [CrossRef] [Green Version]
- Delewski, W.; Paterkiewicz, B.; Manicki, M.; Schilke, B.; Tomiczek, B.; Ciesielski, S.J.; Nierzwicki, L.; Czub, J.; Dutkiewicz, R.; Craig, E.A.; et al. Iron-Sulfur Cluster Biogenesis Chaperones: Evidence for Emergence of Mutational Robustness of a Highly Specific Protein-Protein Interaction. Mol. Biol. Evol. 2016, 33, 643–656. [Google Scholar] [CrossRef] [Green Version]
- Beilschmidt, L.K.; Ollagnier de Choudens, S.; Fournier, M.; Sanakis, I.; Hograindleur, M.A.; Clemancey, M.; Blondin, G.; Schmucker, S.; Eisenmann, A.; Weiss, A.; et al. ISCA1 is essential for mitochondrial Fe4S4 biogenesis in vivo. Nat. Commun. 2017, 8, 15124. [Google Scholar] [CrossRef]
- Brancaccio, D.; Gallo, A.; Mikolajczyk, M.; Zovo, K.; Palumaa, P.; Novellino, E.; Piccioli, M.; Ciofi-Baffoni, S.; Banci, L. Formation of [4Fe–4S] clusters in the mitochondrial iron-sulfur cluster assembly machinery. J. Am. Chem. Soc. 2014, 136, 16240–16250. [Google Scholar] [CrossRef]
- Brancaccio, D.; Gallo, A.; Piccioli, M.; Novellino, E.; Ciofi-Baffoni, S.; Banci, L. [4Fe–4S] Cluster Assembly in Mitochondria and Its Impairment by Copper. J. Am. Chem. Soc. 2017, 139, 719–730. [Google Scholar] [CrossRef] [Green Version]
- Muhlenhoff, U.; Richter, N.; Pines, O.; Pierik, A.J.; Lill, R. Specialized function of yeast Isa1 and Isa2 proteins in the maturation of mitochondrial [4Fe–4S] proteins. J. Biol. Chem. 2011, 286, 41205–41216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiler, B.D.; Bruck, M.C.; Kothe, I.; Bill, E.; Lill, R.; Muhlenhoff, U. Mitochondrial [4Fe–4S] protein assembly involves reductive [2Fe–2S] cluster fusion on ISCA1-ISCA2 by electron flow from ferredoxin FDX2. Proc. Natl. Acad. Sci. USA 2020, 117, 20555–20565. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, M.R.; Cash, V.L.; Weiss, M.C.; Laird, N.F.; Newton, W.E.; Dean, D.R. Biochemical and genetic analysis of the nifUSVWZM cluster from Azotobacter vinelandii. Mol. Gen. Genet. 1989, 219, 49–57. [Google Scholar] [CrossRef]
- Zheng, L.; White, R.H.; Cash, V.L.; Jack, R.F.; Dean, D.R. Cysteine desulfurase activity indicates a role for NIFS in metallocluster biosynthesis. Proc. Natl. Acad. Sci. USA 1993, 90, 2754–2758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, W.; Jack, R.F.; Morgan, T.V.; Dean, D.R.; Johnson, M.K. nifU gene product from Azotobacter vinelandii is a homodimer that contains two identical [2Fe–2S] clusters. Biochemistry 1994, 33, 13455–13463. [Google Scholar] [CrossRef]
- Johnson, D.C.; Dos Santos, P.C.; Dean, D.R. NifU and NifS are required for the maturation of nitrogenase and cannot replace the function of isc-gene products in Azotobacter vinelandii. Biochem. Soc. Trans. 2005, 33, 90–93. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.D.; Jameson, G.N.; Dos Santos, P.C.; Agar, J.N.; Naik, S.; Krebs, C.; Frazzon, J.; Dean, D.R.; Huynh, B.H.; Johnson, M.K. NifS-mediated assembly of [4Fe–4S] clusters in the N- and C-terminal domains of the NifU scaffold protein. Biochemistry 2005, 44, 12955–12969. [Google Scholar] [CrossRef]
- Black, K.A.; Dos Santos, P.C. Shared-intermediates in the biosynthesis of thio-cofactors: Mechanism and functions of cysteine desulfurases and sulfur acceptors. Biochim. Biophys. Acta 2015, 1853, 1470–1480. [Google Scholar] [CrossRef] [Green Version]
- Leimkühler, S.; Bühning, M.; Beilschmidt, L. Shared Sulfur Mobilization Routes for tRNA Thiolation and Molybdenum Cofactor Biosynthesis in Prokaryotes and Eukaryotes. Biomolecules 2017, 7, 5. [Google Scholar]
- Zheng, L.; Cash, V.L.; Flint, D.H.; Dean, D.R. Assembly of iron-sulfur clusters. Identification of an iscSUA-hscBA-fdx gene cluster from Azotobacter vinelandii. J. Biol. Chem. 1998, 273, 13264–13272. [Google Scholar] [CrossRef] [Green Version]
- Patzer, S.I.; Hantke, K. SufS is a NifS-like protein, and SufD is necessary for stability of the [2Fe–2S] FhuF protein in Escherichia coli. J. Bacteriol. 1999, 181, 3307–3309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihara, H.; Kurihara, T.; Yoshimura, T.; Esaki, N. Kinetic and mutational studies of three NifS homologs from Escherichia coli: Mechanistic difference between l-cysteine desulfurase and l-selenocysteine lyase reactions. J. Biochem. 2000, 127, 559–567. [Google Scholar] [CrossRef] [PubMed]
- Nachin, L.; El Hassouni, M.; Loiseau, L.; Expert, D.; Barras, F. SoxR-dependent response to oxidative stress and virulence of Erwinia chrysanthemi: The key role of SufC, an orphan ABC ATPase. Mol. Microbiol. 2001, 39, 960–972. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, Y.; Tokumoto, U. A third bacterial system for the assembly of iron-sulfur clusters with homologs in archaea and plastids. J. Biol. Chem. 2002, 277, 28380–28383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyvltova, E.; Sutak, R.; Harant, K.; Sedinova, M.; Hrdy, I.; Paces, J.; Vlcek, C.; Tachezy, J. NIF-type iron-sulfur cluster assembly system is duplicated and distributed in the mitochondria and cytosol of Mastigamoeba balamuthi. Proc. Natl. Acad. Sci. USA 2013, 110, 7371–7376. [Google Scholar] [CrossRef] [Green Version]
- Garcia, P.S.; Gribaldo, S.; Py, B.; Barras, F. The SUF system: An ABC ATPase-dependent protein complex with a role in Fe–S cluster biogenesis. Res. Microbiol. 2019, 170, 426–434. [Google Scholar] [CrossRef] [Green Version]
- Freibert, S.A.; Goldberg, A.V.; Hacker, C.; Molik, S.; Dean, P.; Williams, T.A.; Nakjang, S.; Long, S.; Sendra, K.; Bill, E.; et al. Evolutionary conservation and in vitro reconstitution of microsporidian iron-sulfur cluster biosynthesis. Nat. Commun. 2017, 8, 13932. [Google Scholar] [CrossRef]
- Couturier, J.; Touraine, B.; BRIAT, J.-F.; Gaymard, F.; Rouhier, N. The iron-sulfur cluster assembly machineries in plants: Current knowledge and open questions. Front. Plant Sci. 2013, 4, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Balk, J.; Schaedler, T.A. Iron Cofactor Assembly in Plants. Annu. Rev. Plant Biol. 2014, 65, 125–153. [Google Scholar] [CrossRef]
- Dai, Y.; Outten, F.W. The E. coli SufS-SufE sulfur transfer system is more resistant to oxidative stress than IscS-IscU. FEBS Lett. 2012, 586, 4016–4022. [Google Scholar] [CrossRef] [Green Version]
- Trotter, V.; Vinella, D.; Loiseau, L.; Ollagnier de Choudens, S.; Fontecave, M.; Barras, F. The CsdA cysteine desulphurase promotes Fe/S biogenesis by recruiting Suf components and participates to a new sulphur transfer pathway by recruiting CsdL (ex-YgdL), a ubiquitin-modifying-like protein. Mol. Microbiol. 2009, 74, 1527–1542. [Google Scholar] [CrossRef] [PubMed]
- Fosset, C.; Chauveau, M.J.; Guillon, B.; Canal, F.; Drapier, J.C.; Bouton, C. RNA silencing of mitochondrial m-Nfs1 reduces Fe–S enzyme activity both in mitochondria and cytosol of mammalian cells. J. Biol. Chem. 2006, 281, 25398–25406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kispal, G.; Csere, P.; Prohl, C.; Lill, R. The mitochondrial proteins Atm1p and Nfs1p are essential for biogenesis of cytosolic Fe/S proteins. EMBO J. 1999, 18, 3981–3989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marelja, Z.; Stocklein, W.; Nimtz, M.; Leimkuhler, S. A novel role for human Nfs1 in the cytoplasm: Nfs1 acts as a sulfur donor for MOCS3, a protein involved in molybdenum cofactor biosynthesis. J. Biol. Chem. 2008, 283, 25178–25185. [Google Scholar] [CrossRef] [Green Version]
- Muhlenhoff, U.; Balk, J.; Richhardt, N.; Kaiser, J.T.; Sipos, K.; Kispal, G.; Lill, R. Functional characterization of the eukaryotic cysteine desulfurase Nfs1p from Saccharomyces cerevisiae. J. Biol. Chem. 2004, 279, 36906–36915. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.K.; Pain, J.; Dancis, A.; Pain, D. Mitochondria export iron-sulfur and sulfur intermediates to the cytoplasm for iron-sulfur cluster assembly and tRNA thiolation in yeast. J. Biol. Chem. 2019, 294, 9489–9502. [Google Scholar] [CrossRef]
- Pastore, A.; Puccio, H. Frataxin: A protein in search for a function. J. Neurochem. 2013, 126 (Suppl. S1), 43–52. [Google Scholar] [CrossRef]
- Roche, B.; Huguenot, A.; Barras, F.; Py, B. The iron-binding CyaY and IscX proteins assist the ISC-catalyzed Fe–S biogenesis in Escherichia coli. Mol. Microbiol. 2015, 95, 605–623. [Google Scholar] [CrossRef] [Green Version]
- Van Vranken, J.G.; Nowinski, S.M.; Clowers, K.J.; Jeong, M.Y.; Ouyang, Y.; Berg, J.A.; Gygi, J.P.; Gygi, S.P.; Winge, D.R.; Rutter, J. ACP Acylation Is an Acetyl-CoA-Dependent Modification Required for Electron Transport Chain Assembly. Mol. Cell 2018, 71, 567–580 e564. [Google Scholar] [CrossRef] [Green Version]
- Agar, J.N.; Krebs, C.; Frazzon, J.; Huynh, B.H.; Dean, D.R.; Johnson, M.K. IscU as a scaffold for iron-sulfur cluster biosynthesis: Sequential assembly of [2Fe–2S] and [4Fe–4S] clusters in IscU. Biochemistry 2000, 39, 7856–7862. [Google Scholar] [CrossRef]
- Mansy, S.S.; Wu, G.; Surerus, K.K.; Cowan, J.A. Iron-sulfur cluster biosynthesis. Thermatoga maritima IscU is a structured iron-sulfur cluster assembly protein. J. Biol. Chem. 2002, 277, 21397–21404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raulfs, E.C.; O’Carroll, I.P.; Dos Santos, P.C.; Unciuleac, M.C.; Dean, D.R. In vivo iron-sulfur cluster formation. Proc. Natl. Acad. Sci. USA 2008, 105, 8591–8596. [Google Scholar] [CrossRef] [Green Version]
- Shimomura, Y.; Wada, K.; Fukuyama, K.; Takahashi, Y. The asymmetric trimeric architecture of [2Fe–2S] IscU: Implications for its scaffolding during iron-sulfur cluster biosynthesis. J. Mol. Biol. 2008, 383, 133–143. [Google Scholar] [CrossRef] [PubMed]
- Bonomi, F.; Iametti, S.; Morleo, A.; Ta, D.; Vickery, L.E. Facilitated transfer of IscU-[2Fe2S] clusters by chaperone-mediated ligand exchange. Biochemistry 2011, 50, 9641–9650. [Google Scholar] [CrossRef] [PubMed]
- Marinoni, E.N.; de Oliveira, J.S.; Nicolet, Y.; Amara, P.; Dean, D.R.; Fontecilla-Camps, J.C. (IscS-IscU)2 complex structures provide insights into Fe2S2 biogenesis and transfer. Angew. Chem. Int. Ed. Engl. 2012, 51, 5439–5442. [Google Scholar] [CrossRef] [PubMed]
- Boniecki, M.T.; Freibert, S.A.; Muhlenhoff, U.; Lill, R.; Cygler, M. Structure and functional dynamics of the mitochondrial Fe/S cluster synthesis complex. Nat. Commun. 2017, 8, 1287. [Google Scholar] [CrossRef] [Green Version]
- Fox, N.G.; Martelli, A.; Nabhan, J.F.; Janz, J.; Borkowska, O.; Bulawa, C.; Yue, W.W. Zinc(II) binding on human wild-type ISCU and Met140 variants modulates NFS1 desulfurase activity. Biochimie 2018, 152, 211–218. [Google Scholar] [CrossRef]
- Iannuzzi, C.; Adrover, M.; Puglisi, R.; Yan, R.; Temussi, P.A.; Pastore, A. The role of zinc in the stability of the marginally stable IscU scaffold protein. Protein Sci. 2014, 23, 1208–1219. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Fuzery, A.K.; Tonelli, M.; Ta, D.T.; Westler, W.M.; Vickery, L.E.; Markley, J.L. Structure and dynamics of the iron-sulfur cluster assembly scaffold protein IscU and its interaction with the cochaperone HscB. Biochemistry 2009, 48, 6062–6071. [Google Scholar] [CrossRef] [Green Version]
- Ramelot, T.A.; Cort, J.R.; Goldsmith-Fischman, S.; Kornhaber, G.J.; Xiao, R.; Shastry, R.; Acton, T.B.; Honig, B.; Montelione, G.T.; Kennedy, M.A. Solution NMR structure of the iron-sulfur cluster assembly protein U (IscU) with zinc bound at the active site. J. Mol. Biol. 2004, 344, 567–583. [Google Scholar] [CrossRef]
- Yan, R.; Kelly, G.; Pastore, A. The scaffold protein IscU retains a structured conformation in the Fe–S cluster assembly complex. Chembiochem 2014, 15, 1682–1686. [Google Scholar] [CrossRef] [PubMed]
- Fox, N.G.; Yu, X.; Feng, X.; Bailey, H.J.; Martelli, A.; Nabhan, J.F.; Strain-Damerell, C.; Bulawa, C.; Yue, W.W.; Han, S. Structure of the human frataxin-bound iron-sulfur cluster assembly complex provides insight into its activation mechanism. Nat. Commun. 2019, 10, 2210. [Google Scholar] [CrossRef] [Green Version]
- Bertini, I.; Cowan, J.A.; Del Bianco, C.; Luchinat, C.; Mansy, S.S. Thermotoga maritima IscU. Structural characterization and dynamics of a new class of metallochaperone. J. Mol. Biol. 2003, 331, 907–924. [Google Scholar] [CrossRef]
- Cai, K.; Frederick, R.O.; Kim, J.H.; Reinen, N.M.; Tonelli, M.; Markley, J.L. Human mitochondrial chaperone (mtHSP70) and cysteine desulfurase (NFS1) bind preferentially to the disordered conformation, whereas co-chaperone (HSC20) binds to the structured conformation of the iron-sulfur cluster scaffold protein (ISCU). J. Biol. Chem. 2013, 288, 28755–28770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansy, S.S.; Wu, S.P.; Cowan, J.A. Iron-sulfur cluster biosynthesis: Biochemical characterization of the conformational dynamics of Thermotoga maritima IscU and the relevance for cellular cluster assembly. J. Biol. Chem. 2004, 279, 10469–10475. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.H.; Tonelli, M.; Markley, J.L. Disordered form of the scaffold protein IscU is the substrate for iron-sulfur cluster assembly on cysteine desulfurase. Proc. Natl. Acad. Sci. USA 2012, 109, 454–459. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.W.; McCabe, J.W.; Russell, D.H.; Barondeau, D.P. Molecular Mechanism of ISC Iron-Sulfur Cluster Biogenesis Revealed by High-Resolution Native Mass Spectrometry. J. Am. Chem. Soc. 2020, 142, 6018–6029. [Google Scholar] [CrossRef]
- Lewis, B.E.; Mason, Z.; Rodrigues, A.V.; Nuth, M.; Dizin, E.; Cowan, J.A.; Stemmler, T.L. Unique roles of iron and zinc binding to the yeast Fe–S cluster scaffold assembly protein “Isu1”. Metallomics 2019, 11, 1820–1835. [Google Scholar] [CrossRef]
- Rodrigues, A.V.; Kandegedara, A.; Rotondo, J.A.; Dancis, A.; Stemmler, T.L. Iron loading site on the Fe–S cluster assembly scaffold protein is distinct from the active site. Biometals 2015, 28, 567–576. [Google Scholar] [CrossRef] [Green Version]
- Dzul, S.P.; Rocha, A.G.; Rawat, S.; Kandegedara, A.; Kusowski, A.; Pain, J.; Murari, A.; Pain, D.; Dancis, A.; Stemmler, T.L. In vitro characterization of a novel Isu homologue from Drosophila melanogaster for de novo FeS-cluster formation. Metallomics 2017, 9, 48–60. [Google Scholar] [CrossRef] [Green Version]
- Adamec, J.; Rusnak, F.; Owen, W.G.; Naylor, S.; Benson, L.M.; Gacy, A.M.; Isaya, G. Iron-dependent self-assembly of recombinant yeast frataxin: Implications for Friedreich ataxia. Am. J. Hum. Genet. 2000, 67, 549–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cavadini, P.; O’Neill, H.A.; Benada, O.; Isaya, G. Assembly and iron-binding properties of human frataxin, the protein deficient in Friedreich ataxia. Hum. Mol. Genet. 2002, 11, 217–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, H.; Yang, J.; Coleman, L.C.; Yeung, S. Distinct iron binding property of two putative iron donors for the iron-sulfur cluster assembly: IscA and the bacterial frataxin ortholog CyaY under physiological and oxidative stress conditions. J. Biol. Chem. 2007, 282, 7997–8004. [Google Scholar] [CrossRef] [Green Version]
- Gakh, O.; Adamec, J.; Gacy, A.M.; Twesten, R.D.; Owen, W.G.; Isaya, G. Physical evidence that yeast frataxin is an iron storage protein. Biochemistry 2002, 41, 6798–6804. [Google Scholar] [CrossRef] [PubMed]
- Layer, G.; Ollagnier-de Choudens, S.; Sanakis, Y.; Fontecave, M. Iron-sulfur cluster biosynthesis: Characterization of Escherichia coli CYaY as an iron donor for the assembly of [2Fe–2S] clusters in the scaffold IscU. J. Biol. Chem. 2006, 281, 16256–16263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, T.; Cowan, J.A. Iron-sulfur cluster biosynthesis. Characterization of frataxin as an iron donor for assembly of [2Fe–2S] clusters in ISU-type proteins. J. Am. Chem. Soc. 2003, 125, 6078–6084. [Google Scholar] [CrossRef]
- Parent, A.; Elduque, X.; Cornu, D.; Belot, L.; Le Caer, J.P.; Grandas, A.; Toledano, M.B.; D’Autreaux, B. Mammalian frataxin directly enhances sulfur transfer of NFS1 persulfide to both ISCU and free thiols. Nat. Commun. 2015, 6, 5686. [Google Scholar] [CrossRef] [Green Version]
- Patra, S.; Barondeau, D.P. Mechanism of activation of the human cysteine desulfurase complex by frataxin. Proc. Natl. Acad. Sci. USA 2019, 116, 19421–19430. [Google Scholar] [CrossRef] [Green Version]
- Armas, A.M.; Balparda, M.; Terenzi, A.; Busi, M.V.; Pagani, M.A.; Gomez-Casati, D.F. Ferrochelatase activity of plant frataxin. Biochimie 2019, 156, 118–122. [Google Scholar] [CrossRef]
- Bellanda, M.; Maso, L.; Doni, D.; Bortolus, M.; De Rosa, E.; Lunardi, F.; Alfonsi, A.; Noguera, M.E.; Herrera, M.G.; Santos, J.; et al. Exploring iron-binding to human frataxin and to selected Friedreich ataxia mutants by means of NMR and EPR spectroscopies. Biochim. Biophys. Acta Proteins Proteom. 2019, 1867, 140254. [Google Scholar] [CrossRef]
- Cai, K.; Frederick, R.O.; Tonelli, M.; Markley, J.L. Interactions of iron-bound frataxin with ISCU and ferredoxin on the cysteine desulfurase complex leading to Fe–S cluster assembly. J. Inorg. Biochem. 2018, 183, 107–116. [Google Scholar] [CrossRef]
- Castro, I.H.; Pignataro, M.F.; Sewell, K.E.; Espeche, L.D.; Herrera, M.G.; Noguera, M.E.; Dain, L.; Nadra, A.D.; Aran, M.; Smal, C.; et al. Frataxin Structure and Function. Subcell Biochem. 2019, 93, 393–438. [Google Scholar]
- Gakh, O.; Ranatunga, W.; Galeano, B.K.; Smith, D.S.T.; Thompson, J.R.; Isaya, G. Defining the Architecture of the Core Machinery for the Assembly of Fe–S Clusters in Human Mitochondria. Methods Enzymol. 2017, 595, 107–160. [Google Scholar]
- Galeano, B.K.; Ranatunga, W.; Gakh, O.; Smith, D.Y.; Thompson, J.R.; Isaya, G. Zinc and the iron donor frataxin regulate oligomerization of the scaffold protein to form new Fe–S cluster assembly centers. Metallomics 2017, 9, 773–801. [Google Scholar] [CrossRef] [Green Version]
- Ranatunga, W.; Gakh, O.; Galeano, B.K.; Smith, D.Y.T.; Soderberg, C.A.; Al-Karadaghi, S.; Thompson, J.R.; Isaya, G. Architecture of the Yeast Mitochondrial Iron-Sulfur Cluster Assembly Machinery: The Sub-Complex Formed by the Iron Donor, Yfh1 Protein, and the Scaffold, Isu1 Protein. J. Biol. Chem. 2016, 291, 10378–10398. [Google Scholar] [CrossRef] [Green Version]
- Babcock, M.; de Silva, D.; Oaks, R.; Davis-Kaplan, S.; Jiralerspong, S.; Montermini, L.; Pandolfo, M.; Kaplan, J. Regulation of mitochondrial iron accumulation by Yfh1p, a putative homolog of frataxin. Science 1997, 276, 1709–1712. [Google Scholar] [CrossRef] [PubMed]
- Duby, G.; Foury, F.; Ramazzotti, A.; Herrmann, J.; Lutz, T. A non-essential function for yeast frataxin in iron-sulfur cluster assembly. Hum. Mol. Genet. 2002, 11, 2635–2643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foury, F.; Cazzalini, O. Deletion of the yeast homologue of the human gene associated with Friedreich’s ataxia elicits iron accumulation in mitochondria. FEBS Lett. 1997, 411, 373–377. [Google Scholar] [CrossRef] [Green Version]
- Puccio, H.; Simon, D.; Cossee, M.; Criqui-Filipe, P.; Tiziano, F.; Melki, J.; Hindelang, C.; Matyas, R.; Rustin, P.; Koenig, M. Mouse models for Friedreich ataxia exhibit cardiomyopathy, sensory nerve defect and Fe–S enzyme deficiency followed by intramitochondrial iron deposits. Nat. Genet. 2001, 27, 181–186. [Google Scholar] [CrossRef]
- Rotig, A.; de Lonlay, P.; Chretien, D.; Foury, F.; Koenig, M.; Sidi, D.; Munnich, A.; Rustin, P. Aconitase and mitochondrial iron-sulphur protein deficiency in Friedreich ataxia. Nat. Genet. 1997, 17, 215–217. [Google Scholar] [CrossRef] [PubMed]
- Gerber, J.; Muhlenhoff, U.; Lill, R. An interaction between frataxin and Isu1/Nfs1 that is crucial for Fe/S cluster synthesis on Isu1. EMBO Rep. 2003, 4, 906–911. [Google Scholar] [CrossRef] [Green Version]
- Ramazzotti, A.; Vanmansart, V.; Foury, F. Mitochondrial functional interactions between frataxin and Isu1p, the iron-sulfur cluster scaffold protein, in Saccharomyces cerevisiae. FEBS Lett. 2004, 557, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Schmucker, S.; Martelli, A.; Colin, F.; Page, A.; Wattenhofer-Donze, M.; Reutenauer, L.; Puccio, H. Mammalian frataxin: An essential function for cellular viability through an interaction with a preformed ISCU/NFS1/ISD11 iron-sulfur assembly complex. PLoS ONE 2011, 6, e16199. [Google Scholar] [CrossRef] [PubMed]
- Prischi, F.; Konarev, P.V.; Iannuzzi, C.; Pastore, C.; Adinolfi, S.; Martin, S.R.; Svergun, D.I.; Pastore, A. Structural bases for the interaction of frataxin with the central components of iron-sulphur cluster assembly. Nat. Commun. 2010, 1, 95. [Google Scholar] [CrossRef] [Green Version]
- Campuzano, V.; Montermini, L.; Molto, M.D.; Pianese, L.; Cossee, M.; Cavalcanti, F.; Monros, E.; Rodius, F.; Duclos, F.; Monticelli, A.; et al. Friedreich’s ataxia: Autosomal recessive disease caused by an intronic GAA triplet repeat expansion. Science 1996, 271, 1423–1427. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Casis, G.; Cote, M.; Barbeau, A. Pathology of the heart in Friedreich’s ataxia: Review of the literature and report of one case. Can. J. Neurol. Sci. 1976, 3, 349–354. [Google Scholar] [CrossRef] [Green Version]
- Cook, J.D.; Bencze, K.Z.; Jankovic, A.D.; Crater, A.K.; Busch, C.N.; Bradley, P.B.; Stemmler, A.J.; Spaller, M.R.; Stemmler, T.L. Monomeric yeast frataxin is an iron-binding protein. Biochemistry 2006, 45, 7767–7777. [Google Scholar] [CrossRef] [Green Version]
- O’Neill, H.A.; Gakh, O.; Isaya, G. Supramolecular assemblies of human frataxin are formed via subunit-subunit interactions mediated by a non-conserved amino-terminal region. J. Mol. Biol. 2005, 345, 433–439. [Google Scholar] [CrossRef]
- Aloria, K.; Schilke, B.; Andrew, A.; Craig, E.A. Iron-induced oligomerization of yeast frataxin homologue Yfh1 is dispensable in vivo. EMBO Rep. 2004, 5, 1096–1101. [Google Scholar] [CrossRef] [Green Version]
- Schmucker, S.; Argentini, M.; Carelle-Calmels, N.; Martelli, A.; Puccio, H. The in vivo mitochondrial two-step maturation of human frataxin. Hum. Mol. Genet. 2008, 17, 3521–3531. [Google Scholar] [CrossRef] [Green Version]
- Seguin, A.; Sutak, R.; Bulteau, A.L.; Garcia-Serres, R.; Oddou, J.L.; Lefevre, S.; Santos, R.; Dancis, A.; Camadro, J.M.; Latour, J.M.; et al. Evidence that yeast frataxin is not an iron storage protein in vivo. Biochim. Biophys. Acta 2010, 1802, 531–538. [Google Scholar] [CrossRef] [PubMed]
- Muhlenhoff, U.; Richhardt, N.; Ristow, M.; Kispal, G.; Lill, R. The yeast frataxin homolog Yfh1p plays a specific role in the maturation of cellular Fe/S proteins. Hum. Mol. Genet. 2002, 11, 2025–2036. [Google Scholar] [CrossRef] [Green Version]
- Stehling, O.; Wilbrecht, C.; Lill, R. Mitochondrial iron-sulfur protein biogenesis and human disease. Biochimie 2014, 100, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Adinolfi, S.; Nair, M.; Politou, A.; Bayer, E.; Martin, S.; Temussi, P.; Pastore, A. The factors governing the thermal stability of frataxin orthologues: How to increase a protein’s stability. Biochemistry 2004, 43, 6511–6518. [Google Scholar] [CrossRef] [PubMed]
- Bou-Abdallah, F.; Adinolfi, S.; Pastore, A.; Laue, T.M.; Dennis Chasteen, N. Iron binding and oxidation kinetics in frataxin CyaY of Escherichia coli. J. Mol. Biol. 2004, 341, 605–615. [Google Scholar] [CrossRef] [PubMed]
- Yoon, T.; Dizin, E.; Cowan, J.A. N-terminal iron-mediated self-cleavage of human frataxin: Regulation of iron binding and complex formation with target proteins. J. Biol. Inorg. Chem. 2007, 12, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Dizin, E.; Cowan, J.A. Mapping iron binding sites on human frataxin: Implications for cluster assembly on the ISU Fe–S cluster scaffold protein. J. Biol. Inorg. Chem. 2008, 13, 825–836. [Google Scholar] [CrossRef]
- Ding, B.; Smith, E.S.; Ding, H. Mobilization of the iron centre in IscA for the iron-sulphur cluster assembly in IscU. Biochem. J. 2005, 389, 797–802. [Google Scholar] [CrossRef] [Green Version]
- Ding, H.; Clark, R.J.; Ding, B. IscA mediates iron delivery for assembly of iron-sulfur clusters in IscU under the limited accessible free iron conditions. J. Biol. Chem. 2004, 279, 37499–37504. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Bitoun, J.P.; Tan, G.; Wang, W.; Min, W.; Ding, H. Iron-binding activity of human iron-sulfur cluster assembly protein hIscA1. Biochem. J. 2010, 428, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Lu, J.; Yang, J.; Tan, G.; Ding, H. Complementary roles of SufA and IscA in the biogenesis of iron-sulfur clusters in Escherichia coli. Biochem. J. 2008, 409, 535–543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Bitoun, J.P.; Ding, H. Interplay of IscA and IscU in biogenesis of iron-sulfur clusters. J. Biol. Chem. 2006, 281, 27956–27963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, H.; Clark, R.J. Characterization of iron binding in IscA, an ancient iron-sulphur cluster assembly protein. Biochem. J. 2004, 379, 433–440. [Google Scholar] [CrossRef]
- Mapolelo, D.T.; Zhang, B.; Naik, S.G.; Huynh, B.H.; Johnson, M.K. Spectroscopic and functional characterization of iron-bound forms of Azotobacter vinelandii (Nif)IscA. Biochemistry 2012, 51, 8056–8070. [Google Scholar] [CrossRef] [Green Version]
- Gupta, V.; Sendra, M.; Naik, S.G.; Chahal, H.K.; Huynh, B.H.; Outten, F.W.; Fontecave, M.; Ollagnier de Choudens, S. Native Escherichia coli SufA, coexpressed with SufBCDSE, purifies as a [2Fe–2S] protein and acts as an Fe–S transporter to Fe–S target enzymes. J. Am. Chem. Soc. 2009, 131, 6149–6153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morimoto, K.; Yamashita, E.; Kondou, Y.; Lee, S.J.; Arisaka, F.; Tsukihara, T.; Nakai, M. The asymmetric IscA homodimer with an exposed [2Fe–2S] cluster suggests the structural basis of the Fe–S cluster biosynthetic scaffold. J. Mol. Biol. 2006, 360, 117–132. [Google Scholar] [CrossRef]
- Wang, W.; Huang, H.; Tan, G.; Si, F.; Liu, M.; Landry, A.P.; Lu, J.; Ding, H. In vivo evidence for the iron-binding activity of an iron-sulfur cluster assembly protein IscA in Escherichia coli. Biochem. J. 2010, 432, 429–436. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.; Geng, M.; Jiang, H.; Liu, Y.; Liu, J.; Qiu, G. The IscA from Acidithiobacillus ferrooxidans is an iron-sulfur protein which assemble the [Fe4S4] cluster with intracellular iron and sulfur. Arch. Biochem. Biophys. 2007, 463, 237–244. [Google Scholar] [CrossRef]
- Mapolelo, D.T.; Zhang, B.; Randeniya, S.; Albetel, A.N.; Li, H.; Couturier, J.; Outten, C.E.; Rouhier, N.; Johnson, M.K. Monothiol glutaredoxins and A-type proteins: Partners in Fe–S cluster trafficking. Dalton Trans. 2013, 42, 3107–3115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, K.D.; Chung, W.H.; Kim, H.J.; Lee, K.C.; Roe, J.H. Monothiol glutaredoxin Grx5 interacts with Fe–S scaffold proteins Isa1 and Isa2 and supports Fe–S assembly and DNA integrity in mitochondria of fission yeast. Biochem. Biophys. Res. Commun. 2010, 392, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Gelling, C.; Dawes, I.W.; Richhardt, N.; Lill, R.; Muhlenhoff, U. Mitochondrial Iba57p is required for Fe/S cluster formation on aconitase and activation of radical SAM enzymes. Mol. Cell Biol. 2008, 28, 1851–1861. [Google Scholar] [CrossRef] [Green Version]
- Tokumoto, U.; Nomura, S.; Minami, Y.; Mihara, H.; Kato, S.; Kurihara, T.; Esaki, N.; Kanazawa, H.; Matsubara, H.; Takahashi, Y. Network of protein-protein interactions among iron-sulfur cluster assembly proteins in Escherichia coli. J. Biochem. 2002, 131, 713–719. [Google Scholar] [CrossRef] [PubMed]
- Al-Karadaghi, S.; Franco, R.; Hansson, M.; Shelnutt, J.A.; Isaya, G.; Ferreira, G.C. Chelatases: Distort to select? Trends Biochem. Sci. 2006, 31, 135–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foster, A.W.; Osman, D.; Robinson, N.J. Metal preferences and metallation. J. Biol. Chem. 2014, 289, 28095–28103. [Google Scholar] [CrossRef] [Green Version]
- Capdevila, D.A.; Edmonds, K.A.; Giedroc, D.P. Metallochaperones and metalloregulation in bacteria. Essays Biochem. 2017, 61, 177–200. [Google Scholar]
- Philpott, C.C.; Jadhav, S. The ins and outs of iron: Escorting iron through the mammalian cytosol. Free Radic Biol. Med. 2019, 133, 112–117. [Google Scholar] [CrossRef]
- Kruszewski, M. The role of labile iron pool in cardiovascular diseases. Acta Biochim. Pol. 2004, 51, 471–480. [Google Scholar] [CrossRef] [Green Version]
- Wofford, J.D.; Bolaji, N.; Dziuba, N.; Outten, F.W.; Lindahl, P.A. Evidence that a respiratory shield in Escherichia coli protects a low-molecular-mass Fe(II) pool from O2-dependent oxidation. J. Biol. Chem. 2019, 294, 50–62. [Google Scholar] [CrossRef] [Green Version]
- Lindahl, P.A.; Moore, M.J. Labile Low-Molecular-Mass Metal Complexes in Mitochondria: Trials and Tribulations of a Burgeoning Field. Biochemistry 2016, 55, 4140–4153. [Google Scholar] [CrossRef]
- McCormick, S.P.; Moore, M.J.; Lindahl, P.A. Detection of Labile Low-Molecular-Mass Transition Metal Complexes in Mitochondria. Biochemistry 2015, 54, 3442–3453. [Google Scholar] [CrossRef]
- Moore, M.J.; Wofford, J.D.; Dancis, A.; Lindahl, P.A. Recovery of mrs3∆mrs4∆ Saccharomyces cerevisiae Cells under Iron-Sufficient Conditions and the Role of Fe580. Biochemistry 2018, 57, 672–683. [Google Scholar] [CrossRef] [PubMed]
- Hider, R.C.; Kong, X.L. Glutathione: A key component of the cytoplasmic labile iron pool. Biometals 2011, 24, 1179–1187. [Google Scholar] [CrossRef] [PubMed]
- Cupp-Vickery, J.R.; Urbina, H.; Vickery, L.E. Crystal structure of IscS, a cysteine desulfurase from Escherichia coli. J. Mol. Biol. 2003, 330, 1049–1059. [Google Scholar] [CrossRef]
- Fujii, T.; Maeda, M.; Mihara, H.; Kurihara, T.; Esaki, N.; Hata, Y. Structure of a NifS homologue: X-ray structure analysis of CsdB, an Escherichia coli counterpart of mammalian selenocysteine lyase. Biochemistry 2000, 39, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, J.T.; Clausen, T.; Bourenkow, G.P.; Bartunik, H.D.; Steinbacher, S.; Huber, R. Crystal structure of a NifS-like protein from Thermotoga maritima: Implications for iron sulphur cluster assembly. J. Mol. Biol. 2000, 297, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, R.; Hikita, M.; Ogawa, S.; Takahashi, Y.; Fujishiro, T. Snapshots of PLP-substrate and PLP-product external aldimines as intermediates in two types of cysteine desulfurase enzymes. FEBS J. 2020, 287, 1138–1154. [Google Scholar] [CrossRef]
- Roret, T.; Pegeot, H.; Couturier, J.; Mulliert, G.; Rouhier, N.; Didierjean, C. X-ray structures of Nfs2, the plastidial cysteine desulfurase from Arabidopsis thaliana. Acta Crystallogr. F Struct. Biol. Commun. 2014, 70, 1180–1185. [Google Scholar] [CrossRef] [Green Version]
- Rybniker, J.; Pojer, F.; Marienhagen, J.; Kolly, G.S.; Chen, J.M.; van Gumpel, E.; Hartmann, P.; Cole, S.T. The cysteine desulfurase IscS of Mycobacterium tuberculosis is involved in iron-sulfur cluster biogenesis and oxidative stress defence. Biochem. J. 2014, 459, 467–478. [Google Scholar] [CrossRef] [Green Version]
- Shi, R.; Proteau, A.; Villarroya, M.; Moukadiri, I.; Zhang, L.; Trempe, J.F.; Matte, A.; Armengod, M.E.; Cygler, M. Structural basis for Fe–S cluster assembly and tRNA thiolation mediated by IscS protein-protein interactions. PLoS Biol. 2010, 8, e1000354. [Google Scholar] [CrossRef]
- Cory, S.A.; Van Vranken, J.G.; Brignole, E.J.; Patra, S.; Winge, D.R.; Drennan, C.L.; Rutter, J.; Barondeau, D.P. Structure of human Fe–S assembly subcomplex reveals unexpected cysteine desulfurase architecture and acyl-ACP-ISD11 interactions. Proc. Natl. Acad. Sci. USA 2017, 114, E5325–E5334. [Google Scholar] [CrossRef] [Green Version]
- Turowski, V.R.; Busi, M.V.; Gomez-Casati, D.F. Structural and functional studies of the mitochondrial cysteine desulfurase from Arabidopsis thaliana. Mol. Plant 2012, 5, 1001–1010. [Google Scholar] [CrossRef] [Green Version]
- Herrera, M.G.; Pignataro, M.F.; Noguera, M.E.; Cruz, K.M.; Santos, J. Rescuing the Rescuer: On the Protein Complex between the Human Mitochondrial Acyl Carrier Protein and ISD11. ACS Chem. Biol. 2018, 13, 1455–1462. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.D.; Agar, J.N.; Johnson, K.A.; Frazzon, J.; Amster, I.J.; Dean, D.R.; Johnson, M.K. Sulfur transfer from IscS to IscU: The first step in iron-sulfur cluster biosynthesis. J. Am. Chem. Soc. 2001, 123, 11103–11104. [Google Scholar] [CrossRef]
- Urbina, H.D.; Silberg, J.J.; Hoff, K.G.; Vickery, L.E. Transfer of sulfur from IscS to IscU during Fe/S cluster assembly. J. Biol. Chem. 2001, 276, 44521–44526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.D.; Frazzon, J.; Dean, D.R.; Johnson, M.K. Role of conserved cysteines in mediating sulfur transfer from IscS to IscU. FEBS Lett. 2005, 579, 5236–5240. [Google Scholar] [CrossRef] [Green Version]
- Nuth, M.; Yoon, T.; Cowan, J.A. Iron-sulfur cluster biosynthesis: Characterization of iron nucleation sites for assembly of the [2Fe–2S]2+ cluster core in IscU proteins. J. Am. Chem. Soc. 2002, 124, 8774–8775. [Google Scholar] [CrossRef]
- Bridwell-Rabb, J.; Fox, N.G.; Tsai, C.L.; Winn, A.M.; Barondeau, D.P. Human frataxin activates Fe–S cluster biosynthesis by facilitating sulfur transfer chemistry. Biochemistry 2014, 53, 4904–4913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
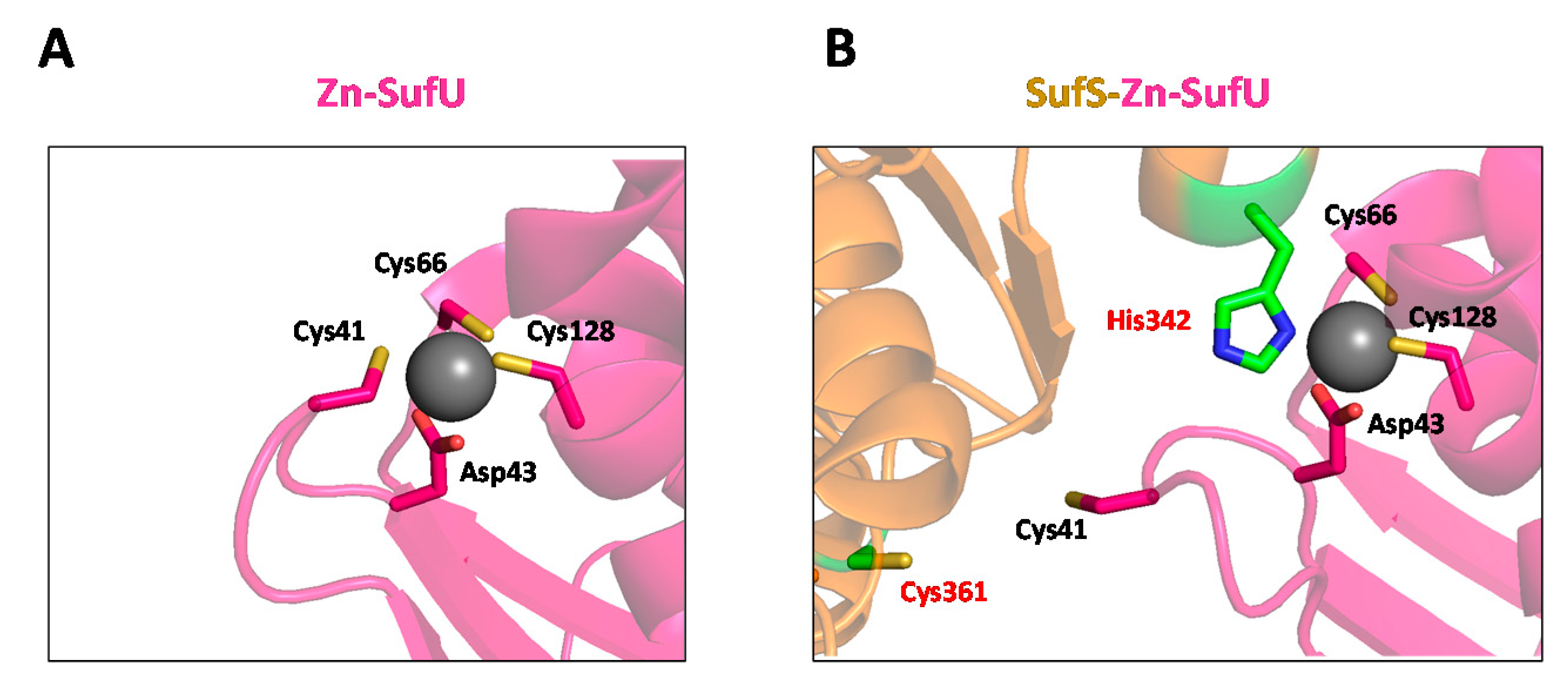
- Selbach, B.P.; Chung, A.H.; Scott, A.D.; George, S.J.; Cramer, S.P.; Dos Santos, P.C. Fe–S Cluster Biogenesis in Gram-Positive Bacteria: SufU Is a Zinc-Dependent Sulfur Transfer Protein. Biochemistry 2014, 53, 152–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adinolfi, S.; Iannuzzi, C.; Prischi, F.; Pastore, C.; Iametti, S.; Martin, S.R.; Bonomi, F.; Pastore, A. Bacterial frataxin CyaY is the gatekeeper of iron-sulfur cluster formation catalyzed by IscS. Nat. Struct. Mol. Biol. 2009, 16, 390–396. [Google Scholar] [CrossRef]
- Nuth, M.; Cowan, J.A. Iron-sulfur cluster biosynthesis: Characterization of IscU-IscS complex formation and a structural model for sulfide delivery to the [2Fe–2S] assembly site. J. Biol. Inorg. Chem. 2009, 14, 829–839. [Google Scholar] [CrossRef]
- Tsai, C.L.; Barondeau, D.P. Human frataxin is an allosteric switch that activates the Fe–S cluster biosynthetic complex. Biochemistry 2010, 49, 9132–9139. [Google Scholar] [CrossRef] [PubMed]
- Bridwell-Rabb, J.; Iannuzzi, C.; Pastore, A.; Barondeau, D.P. Effector role reversal during evolution: The case of frataxin in Fe–S cluster biosynthesis. Biochemistry 2012, 51, 2506–2514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colin, F.; Martelli, A.; Clemancey, M.; Latour, J.M.; Gambarelli, S.; Zeppieri, L.; Birck, C.; Page, A.; Puccio, H.; Ollagnier de Choudens, S. Mammalian frataxin controls sulfur production and iron entry during de novo Fe4S4 cluster assembly. J. Am. Chem. Soc. 2013, 135, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Unciuleac, M.C.; Chandramouli, K.; Naik, S.; Mayer, S.; Huynh, B.H.; Johnson, M.K.; Dean, D.R. In vitro activation of apo-aconitase using a [4Fe–4S] cluster-loaded form of the IscU [Fe-S] cluster scaffolding protein. Biochemistry 2007, 46, 6812–6821. [Google Scholar] [CrossRef]
- Iannuzzi, C.; Adinolfi, S.; Howes, B.D.; Garcia-Serres, R.; Clemancey, M.; Latour, J.M.; Smulevich, G.; Pastore, A. The role of CyaY in iron sulfur cluster assembly on the E. coli IscU scaffold protein. PLoS ONE 2011, 6, e21992. [Google Scholar] [CrossRef] [Green Version]
- Fox, N.G.; Chakrabarti, M.; McCormick, S.P.; Lindahl, P.A.; Barondeau, D.P. The Human Iron-Sulfur Assembly Complex Catalyzes the Synthesis of [2Fe–2S] Clusters on ISCU2 That Can Be Transferred to Acceptor Molecules. Biochemistry 2015, 54, 3871–3879. [Google Scholar] [CrossRef] [Green Version]
- Tokumoto, U.; Takahashi, Y. Genetic analysis of the isc operon in Escherichia coli involved in the biogenesis of cellular iron-sulfur proteins. J. Biochem. 2001, 130, 63–71. [Google Scholar] [CrossRef]
- Lange, H.; Kaut, A.; Kispal, G.; Lill, R. A mitochondrial ferredoxin is essential for biogenesis of cellular iron-sulfur proteins. Proc. Natl. Acad. Sci. USA 2000, 97, 1050–1055. [Google Scholar] [CrossRef] [Green Version]
- Djaman, O.; Outten, F.W.; Imlay, J.A. Repair of oxidized iron-sulfur clusters in Escherichia coli. J. Biol. Chem. 2004, 279, 44590–44599. [Google Scholar] [CrossRef] [Green Version]
- Muhlenhoff, U.; Gerber, J.; Richhardt, N.; Lill, R. Components involved in assembly and dislocation of iron-sulfur clusters on the scaffold protein Isu1p. EMBO J. 2003, 22, 4815–4825. [Google Scholar] [CrossRef] [Green Version]
- Sheftel, A.D.; Stehling, O.; Pierik, A.J.; Elsasser, H.P.; Muhlenhoff, U.; Webert, H.; Hobler, A.; Hannemann, F.; Bernhardt, R.; Lill, R. Humans possess two mitochondrial ferredoxins, Fdx1 and Fdx2, with distinct roles in steroidogenesis, heme, and Fe/S cluster biosynthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 11775–11780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Ghosh, M.; Kovtunovych, G.; Crooks, D.R.; Rouault, T.A. Both human ferredoxins 1 and 2 and ferredoxin reductase are important for iron-sulfur cluster biogenesis. Biochim. Biophys. Acta 2012, 1823, 484–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Saxena, S.; Pain, D.; Dancis, A. Adrenodoxin reductase homolog (Arh1p) of yeast mitochondria required for iron homeostasis. J. Biol. Chem. 2001, 276, 1503–1509. [Google Scholar] [CrossRef] [Green Version]
- Wan, J.T.; Jarrett, J.T. Electron acceptor specificity of ferredoxin (flavodoxin): NADP+ oxidoreductase from Escherichia coli. Arch. Biochem. Biophys. 2002, 406, 116–126. [Google Scholar] [CrossRef]
- Yan, R.; Adinolfi, S.; Pastore, A. Ferredoxin, in conjunction with NADPH and ferredoxin-NADP reductase, transfers electrons to the IscS/IscU complex to promote iron-sulfur cluster assembly. Biochim. Biophys. Acta 2015, 1854, 1113–1117. [Google Scholar] [CrossRef] [Green Version]
- Cai, K.; Tonelli, M.; Frederick, R.O.; Markley, J.L. Human Mitochondrial Ferredoxin 1 (FDX1) and Ferredoxin 2 (FDX2) Both Bind Cysteine Desulfurase and Donate Electrons for Iron-Sulfur Cluster Biosynthesis. Biochemistry 2017, 56, 487–499. [Google Scholar] [CrossRef]
- Kim, J.H.; Frederick, R.O.; Reinen, N.M.; Troupis, A.T.; Markley, J.L. [2Fe–2S]-ferredoxin binds directly to cysteine desulfurase and supplies an electron for iron-sulfur cluster assembly but is displaced by the scaffold protein or bacterial frataxin. J. Am. Chem. Soc. 2013, 135, 8117–8120. [Google Scholar] [CrossRef]
- Yan, R.; Konarev, P.V.; Iannuzzi, C.; Adinolfi, S.; Roche, B.; Kelly, G.; Simon, L.; Martin, S.R.; Py, B.; Barras, F.; et al. Ferredoxin competes with bacterial frataxin in binding to the desulfurase IscS. J. Biol. Chem. 2013, 288, 24777–24787. [Google Scholar] [CrossRef] [Green Version]
- Galardon, E.; Roger, T.; Deschamps, P.; Roussel, P.; Tomas, A.; Artaud, I. Synthesis of a Fe(II)SH complex stabilized by an intramolecular N-H...S hydrogen bond, which acts as a H2S donor. Inorg. Chem. 2012, 51, 10068–10070. [Google Scholar] [CrossRef]
- Tsou, C.C.; Chiu, W.C.; Ke, C.H.; Tsai, J.C.; Wang, Y.M.; Chiang, M.H.; Liaw, W.F. Iron(III) bound by hydrosulfide anion ligands: NO-promoted stabilization of the [Fe(III)-SH] motif. J. Am. Chem. Soc. 2014, 136, 9424–9433. [Google Scholar] [CrossRef]
- Dey, A.; Hocking, R.K.; Larsen, P.; Borovik, A.S.; Hodgson, K.O.; Hedman, B.; Solomon, E.I. X-ray absorption spectroscopy and density functional theory studies of [(H3buea)FeIII−X]n− (X = S2−, O2−, OH−): Comparison of bonding and hydrogen bonding in oxo and sulfido complexes. J. Am. Chem. Soc. 2006, 128, 9825–9833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conradie, J.; Tangen, E.; Ghosh, A. Trigonal bipyramidal iron(III) and manganese(III) oxo, sulfido, and selenido complexes. An electronic-structural overview. J. Inorg. Biochem. 2006, 100, 707–715. [Google Scholar] [CrossRef] [PubMed]
- Larsen, P.L.; Gupta, R.; Powell, D.R.; Borovik, A.S. Chalcogens as terminal ligands to iron: Synthesis and structure of complexes with Fe(III)-S and Fe(III)-Se motifs. J. Am. Chem. Soc. 2004, 126, 6522–6523. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Outten, C.E. Monothiol CGFS glutaredoxins and BolA-like proteins: [2Fe–2S] binding partners in iron homeostasis. Biochemistry 2012, 51, 4377–4389. [Google Scholar] [CrossRef] [Green Version]
- Rouhier, N.; Couturier, J.; Johnson, M.K.; Jacquot, J.P. Glutaredoxins: Roles in iron homeostasis. Trends Biochem. Sci. 2010, 35, 43–52. [Google Scholar] [CrossRef] [Green Version]
- Banci, L.; Brancaccio, D.; Ciofi-Baffoni, S.; Del Conte, R.; Gadepalli, R.; Mikolajczyk, M.; Neri, S.; Piccioli, M.; Winkelmann, J. [2Fe–2S] cluster transfer in iron-sulfur protein biogenesis. Proc. Natl. Acad. Sci. USA 2014, 111, 6203–6208. [Google Scholar] [CrossRef] [Green Version]
- Shakamuri, P.; Zhang, B.; Johnson, M.K. Monothiol glutaredoxins function in storing and transporting [Fe2S2] clusters assembled on IscU scaffold proteins. J. Am. Chem. Soc. 2012, 134, 15213–15216. [Google Scholar] [CrossRef]
- Agar, J.N.; Zheng, L.; Cash, V.L.; Dean, D.R.; Johnson, M.K. Role of the IscU Protein in Iron−Sulfur Cluster Biosynthesis: IscS-mediated Assembly of a [Fe2S2] Cluster in IscU. J. Am. Chem. Soc. 2000, 122, 2136–2137. [Google Scholar] [CrossRef]
- Wu, G.; Mansy, S.S.; Wu Sp, S.P.; Surerus, K.K.; Foster, M.W.; Cowan, J.A. Characterization of an iron-sulfur cluster assembly protein (ISU1) from Schizosaccharomyces pombe. Biochemistry 2002, 41, 5024–5032. [Google Scholar] [CrossRef]
- Tsai, C.L.; Bridwell-Rabb, J.; Barondeau, D.P. Friedreich’s ataxia variants I154F and W155R diminish frataxin-based activation of the iron-sulfur cluster assembly complex. Biochemistry 2011, 50, 6478–6487. [Google Scholar] [CrossRef] [Green Version]
- Bridwell-Rabb, J.; Winn, A.M.; Barondeau, D.P. Structure-function analysis of Friedreich’s ataxia mutants reveals determinants of frataxin binding and activation of the Fe–S assembly complex. Biochemistry 2011, 50, 7265–7274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, N.G.; Das, D.; Chakrabarti, M.; Lindahl, P.A.; Barondeau, D.P. Frataxin Accelerates [2Fe–2S] Cluster Formation on the Human Fe–S Assembly Complex. Biochemistry 2015, 54, 3880–3889. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandey, A.; Gordon, D.M.; Pain, J.; Stemmler, T.L.; Dancis, A.; Pain, D. Frataxin directly stimulates mitochondrial cysteine desulfurase by exposing substrate-binding sites, and a mutant Fe–S cluster scaffold protein with frataxin-bypassing ability acts similarly. J. Biol. Chem. 2013, 288, 36773–36786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Behshad, E.; Parkin, S.E.; Bollinger, J.M., Jr. Mechanism of cysteine desulfurase Slr0387 from Synechocystis sp. PCC 6803: Kinetic analysis of cleavage of the persulfide intermediate by chemical reductants. Biochemistry 2004, 43, 12220–12226. [Google Scholar] [CrossRef]
- Lacourciere, G.M.; Stadtman, T.C. The NIFS protein can function as a selenide delivery protein in the biosynthesis of selenophosphate. J. Biol. Chem. 1998, 273, 30921–30926. [Google Scholar] [CrossRef] [Green Version]
- Zheng, L.; White, R.H.; Cash, V.L.; Dean, D.R. Mechanism for the desulfurization of L-cysteine catalyzed by the nifS gene product. Biochemistry 1994, 33, 4714–4720. [Google Scholar] [CrossRef]
- Ponsero, A.J.; Igbaria, A.; Darch, M.A.; Miled, S.; Outten, C.E.; Winther, J.R.; Palais, G.; D’Autreaux, B.; Delaunay-Moisan, A.; Toledano, M.B. Endoplasmic Reticulum Transport of Glutathione by Sec61 Is Regulated by Ero1 and Bip. Mol. Cell 2017, 67, 962–973 e965. [Google Scholar] [CrossRef] [Green Version]
- Furne, J.; Saeed, A.; Levitt, M.D. Whole tissue hydrogen sulfide concentrations are orders of magnitude lower than presently accepted values. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R1479–R1485. [Google Scholar] [CrossRef] [Green Version]
- Ross-Inta, C.; Tsai, C.Y.; Giulivi, C. The mitochondrial pool of free amino acids reflects the composition of mitochondrial DNA-encoded proteins: Indication of a post-translational quality control for protein synthesis. Biosci. Rep. 2008, 28, 239–249. [Google Scholar] [CrossRef] [Green Version]
- Agar, J.N.; Yuvaniyama, P.; Jack, R.F.; Cash, V.L.; Smith, A.D.; Dean, D.R.; Johnson, M.K. Modular organization and identification of a mononuclear iron-binding site within the NifU protein. J. Biol. Inorg. Chem. 2000, 5, 167–177. [Google Scholar] [CrossRef]
- Yuvaniyama, P.; Agar, J.N.; Cash, V.L.; Johnson, M.K.; Dean, D.R. NifS-directed assembly of a transient [2Fe–2S] cluster within the NifU protein. Proc. Natl. Acad. Sci. USA 2000, 97, 599–604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandramouli, K.; Unciuleac, M.C.; Naik, S.; Dean, D.R.; Huynh, B.H.; Johnson, M.K. Formation and properties of [4Fe–4S] clusters on the IscU scaffold protein. Biochemistry 2007, 46, 6804–6811. [Google Scholar] [CrossRef] [PubMed]
- Outten, F.W. Recent advances in the Suf Fe–S cluster biogenesis pathway: Beyond the Proteobacteria. Biochim. Biophys. Acta 2015, 1853, 1464–1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wollers, S.; Layer, G.; Garcia-Serres, R.; Signor, L.; Clemancey, M.; Latour, J.M.; Fontecave, M.; Ollagnier de Choudens, S. Iron–sulfur (Fe–S) cluster assembly: The SufBC2D complex is a new type of Fe–S scaffold with a flavin redox cofactor. J. Biol. Chem. 2010, 285, 23331–23341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirabayashi, K.; Yuda, E.; Tanaka, N.; Katayama, S.; Iwasaki, K.; Matsumoto, T.; Kurisu, G.; Outten, F.W.; Fukuyama, K.; Takahashi, Y.; et al. Functional Dynamics Revealed by the Structure of the SufBC2D Complex, a Novel ATP-binding Cassette (ABC) Protein That Serves as a Scaffold for Iron-Sulfur Cluster Biogenesis. J. Biol. Chem. 2015, 290, 29717–29731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riboldi, G.P.; Verli, H.; Frazzon, J. Structural studies of the Enterococcus faecalis SufU [Fe-S] cluster protein. BMC Biochem. 2009, 10, 3. [Google Scholar] [CrossRef] [Green Version]
- Selbach, B.; Earles, E.; Dos Santos, P.C. Kinetic analysis of the bisubstrate cysteine desulfurase SufS from Bacillus subtilis. Biochemistry 2010, 49, 8794–8802. [Google Scholar] [CrossRef] [PubMed]
- Albrecht, A.G.; Netz, D.J.; Miethke, M.; Pierik, A.J.; Burghaus, O.; Peuckert, F.; Lill, R.; Marahiel, M.A. SufU is an essential iron-sulfur cluster scaffold protein in Bacillus subtilis. J. Bacteriol. 2010, 192, 1643–1651. [Google Scholar] [CrossRef] [Green Version]
- Yokoyama, N.; Nonaka, C.; Ohashi, Y.; Shioda, M.; Terahata, T.; Chen, W.; Sakamoto, K.; Maruyama, C.; Saito, T.; Yuda, E.; et al. Distinct roles for U-type proteins in iron-sulfur cluster biosynthesis revealed by genetic analysis of the Bacillus subtilis sufCDSUB operon. Mol. Microbiol. 2018, 107, 688–703. [Google Scholar] [CrossRef] [Green Version]
- Saini, A.; Mapolelo, D.T.; Chahal, H.K.; Johnson, M.K.; Outten, F.W. SufD and SufC ATPase activity are required for iron acquisition during in vivo Fe–S cluster formation on SufB. Biochemistry 2010, 49, 9402–9412. [Google Scholar] [CrossRef] [Green Version]
- Nachin, L.; Loiseau, L.; Expert, D.; Barras, F. SufC: An unorthodox cytoplasmic ABC/ATPase required for [Fe-S] biogenesis under oxidative stress. EMBO J. 2003, 22, 427–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tirupati, B.; Vey, J.L.; Drennan, C.L.; Bollinger, J.M., Jr. Kinetic and structural characterization of Slr0077/SufS, the essential cysteine desulfurase from Synechocystis sp. PCC 6803. Biochemistry 2004, 43, 12210–12219. [Google Scholar] [CrossRef] [PubMed]
- Outten, F.W.; Wood, M.J.; Munoz, F.M.; Storz, G. The SufE protein and the SufBC2D complex enhance SufS cysteine desulfurase activity as part of a sulfur transfer pathway for Fe–S cluster assembly in Escherichia coli. J. Biol. Chem. 2003, 278, 45713–45719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zafrilla, B.; Martinez-Espinosa, R.M.; Esclapez, J.; Perez-Pomares, F.; Bonete, M.J. SufS protein from Haloferax volcanii involved in Fe–S cluster assembly in haloarchaea. Biochim. Biophys. Acta 2010, 1804, 1476–1482. [Google Scholar] [CrossRef]
- Ollagnier-de-Choudens, S.; Lascoux, D.; Loiseau, L.; Barras, F.; Forest, E.; Fontecave, M. Mechanistic studies of the SufS-SufE cysteine desulfurase: Evidence for sulfur transfer from SufS to SufE. FEBS Lett. 2003, 555, 263–267. [Google Scholar] [CrossRef] [Green Version]
- Singh, H.; Dai, Y.; Outten, F.W.; Busenlehner, L.S. Escherichia coli SufE sulfur transfer protein modulates the SufS cysteine desulfurase through allosteric conformational dynamics. J. Biol. Chem. 2013, 288, 36189–36200. [Google Scholar] [CrossRef] [Green Version]
- Dai, Y.; Kim, D.; Dong, G.; Busenlehner, L.S.; Frantom, P.A.; Outten, F.W. SufE D74R Substitution Alters Active Site Loop Dynamics To Further Enhance SufE Interaction with the SufS Cysteine Desulfurase. Biochemistry 2015, 54, 4824–4833. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Singh, H.; Dai, Y.; Dong, G.; Busenlehner, L.S.; Outten, F.W.; Frantom, P.A. Changes in Protein Dynamics in Escherichia coli SufS Reveal a Possible Conserved Regulatory Mechanism in Type II Cysteine Desulfurase Systems. Biochemistry 2018, 57, 5210–5217. [Google Scholar] [CrossRef]
- Dunkle, J.A.; Bruno, M.R.; Outten, F.W.; Frantom, P.A. Structural Evidence for Dimer-Interface-Driven Regulation of the Type II Cysteine Desulfurase, SufS. Biochemistry 2019, 58, 687–696. [Google Scholar] [CrossRef]
- Blahut, M.; Wise, C.E.; Bruno, M.R.; Dong, G.; Makris, T.M.; Frantom, P.A.; Dunkle, J.A.; Outten, F.W. Direct observation of intermediates in the SufS cysteine desulfurase reaction reveals functional roles of conserved active-site residues. J. Biol. Chem. 2019, 294, 12444–12458. [Google Scholar] [CrossRef]
- Kim, S.; Park, S. Structural changes during cysteine desulfurase CsdA and sulfur acceptor CsdE interactions provide insight into the trans-persulfuration. J. Biol. Chem. 2013, 288, 27172–27180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunkle, J.A.; Bruno, M.R.; Frantom, P.A. Structural evidence for a latch mechanism regulating access to the active site of SufS-family cysteine desulfurases. Acta Crystallogr. D Struct. Biol. 2020, 76, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Fernández, F.J.; Ardá, A.; López-Estepa, M.; Aranda, J.; Peña-Soler, E.; Garces, F.; Round, A.; Campos-Olivas, R.; Bruix, M.; Coll, M.; et al. Mechanism of Sulfur Transfer Across Protein–Protein Interfaces: The Cysteine Desulfurase Model System. ACS Catal. 2016, 6, 3975–3984. [Google Scholar] [CrossRef] [Green Version]
- Goldsmith-Fischman, S.; Kuzin, A.; Edstrom, W.C.; Benach, J.; Shastry, R.; Xiao, R.; Acton, T.B.; Honig, B.; Montelione, G.T.; Hunt, J.F. The SufE sulfur-acceptor protein contains a conserved core structure that mediates interdomain interactions in a variety of redox protein complexes. J. Mol. Biol. 2004, 344, 549–565. [Google Scholar] [CrossRef]
- Liu, G.; Li, Z.; Chiang, Y.; Acton, T.; Montelione, G.T.; Murray, D.; Szyperski, T. High-quality homology models derived from NMR and X-ray structures of E. coli proteins YgdK and Suf E suggest that all members of the YgdK/Suf E protein family are enhancers of cysteine desulfurases. Protein Sci. 2005, 14, 1597–1608. [Google Scholar] [CrossRef] [Green Version]
- Selbach, B.P.; Pradhan, P.K.; Dos Santos, P.C. Protected sulfur transfer reactions by the Escherichia coli Suf system. Biochemistry 2013, 52, 4089–4096. [Google Scholar] [CrossRef]
- Riboldi, G.P.; de Oliveira, J.S.; Frazzon, J. Enterococcus faecalis SufU scaffold protein enhances SufS desulfurase activity by acquiring sulfur from its cysteine-153. Biochim. Biophys. Acta 2011, 1814, 1910–1918. [Google Scholar] [CrossRef] [Green Version]
- Albrecht, A.G.; Peuckert, F.; Landmann, H.; Miethke, M.; Seubert, A.; Marahiel, M.A. Mechanistic characterization of sulfur transfer from cysteine desulfurase SufS to the iron-sulfur scaffold SufU in Bacillus subtilis. FEBS Lett. 2011, 585, 465–470. [Google Scholar] [CrossRef] [Green Version]
- Kornhaber, G.J.; Snyder, D.; Moseley, H.N.; Montelione, G.T. Identification of zinc-ligated cysteine residues based on 13Calpha and 13Cbeta chemical shift data. J. Biomol. NMR 2006, 34, 259–269. [Google Scholar] [CrossRef]
- Liu, J.; Oganesyan, N.; Shin, D.H.; Jancarik, J.; Yokota, H.; Kim, R.; Kim, S.H. Structural characterization of an iron-sulfur cluster assembly protein IscU in a zinc-bound form. Proteins 2005, 59, 875–881. [Google Scholar] [CrossRef]
- Fujishiro, T.; Terahata, T.; Kunichika, K.; Yokoyama, N.; Maruyama, C.; Asai, K.; Takahashi, Y. Zinc-Ligand Swapping Mediated Complex Formation and Sulfur Transfer between SufS and SufU for Iron-Sulfur Cluster Biogenesis in Bacillus subtilis. J. Am. Chem. Soc. 2017, 139, 18464–18467. [Google Scholar] [CrossRef] [PubMed]
- Blauenburg, B.; Mielcarek, A.; Altegoer, F.; Fage, C.D.; Linne, U.; Bange, G.; Marahiel, M.A. Crystal Structure of Bacillus subtilis Cysteine Desulfurase SufS and Its Dynamic Interaction with Frataxin and Scaffold Protein SufU. PLoS ONE 2016, 11, e0158749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuda, E.; Tanaka, N.; Fujishiro, T.; Yokoyama, N.; Hirabayashi, K.; Fukuyama, K.; Wada, K.; Takahashi, Y. Mapping the key residues of SufB and SufD essential for biosynthesis of iron-sulfur clusters. Sci. Rep. 2017, 7, 9387. [Google Scholar] [CrossRef] [PubMed] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Functional Role | Prokaryote | Yeast | Mammal |
|---|---|---|---|
| Mitochondrial iron transporter | - | Mrs3/4 | MFRN1/2 |
| U-type scaffold | IscU | Isu1/2 | ISCU |
| Cysteine desulfurase | IscS | Nfs1 | NFS1 |
| Desulfurase-interacting protein 11 | - | Isd11, Lyrm4 | ISD11, LYRM4 |
| Acyl carrier protein | ACP | ACP | ACP |
| Frataxin | CyaY | Yfh1 | FXN |
| IscX | IscX | - | - |
| Ferredoxin | Fdx | Yah1 | FDX2 |
| Ferredoxin reductase | Fenr | Arh1 | FDXR |
| Hsp70 chaperone | HscA | Ssq1 | HSPA9 |
| J-type co-chaperone | HscB | Jac1 | HSC20 |
| Nucleotide exchanger | - | Mge1 | GRPE |
| Glutaredoxin | Grx4 | Grx5 | GRX5 |
| A-type scaffold | IscA | Isa1/2 | ISCA1/2 |
| Late-acting components | - | Iba57 | IBA57 |
| Bola | Bola1 | BOLA1 | |
| - | Bola3 | BOLA3 | |
| NfuA | Nfu1 | NFU1 | |
| Mrp | Ind1 | IND1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srour, B.; Gervason, S.; Monfort, B.; D’Autréaux, B. Mechanism of Iron–Sulfur Cluster Assembly: In the Intimacy of Iron and Sulfur Encounter. Inorganics 2020, 8, 55. https://doi.org/10.3390/inorganics8100055
Srour B, Gervason S, Monfort B, D’Autréaux B. Mechanism of Iron–Sulfur Cluster Assembly: In the Intimacy of Iron and Sulfur Encounter. Inorganics. 2020; 8(10):55. https://doi.org/10.3390/inorganics8100055
Chicago/Turabian StyleSrour, Batoul, Sylvain Gervason, Beata Monfort, and Benoit D’Autréaux. 2020. "Mechanism of Iron–Sulfur Cluster Assembly: In the Intimacy of Iron and Sulfur Encounter" Inorganics 8, no. 10: 55. https://doi.org/10.3390/inorganics8100055
APA StyleSrour, B., Gervason, S., Monfort, B., & D’Autréaux, B. (2020). Mechanism of Iron–Sulfur Cluster Assembly: In the Intimacy of Iron and Sulfur Encounter. Inorganics, 8(10), 55. https://doi.org/10.3390/inorganics8100055