Kinetic Studies of Sodium and Metforminium Decavanadates Decomposition and In Vitro Cytotoxicity and Insulin- Like Activity


 , ,
, , 
Abstract
:1. Introduction
2. Results and Discussion
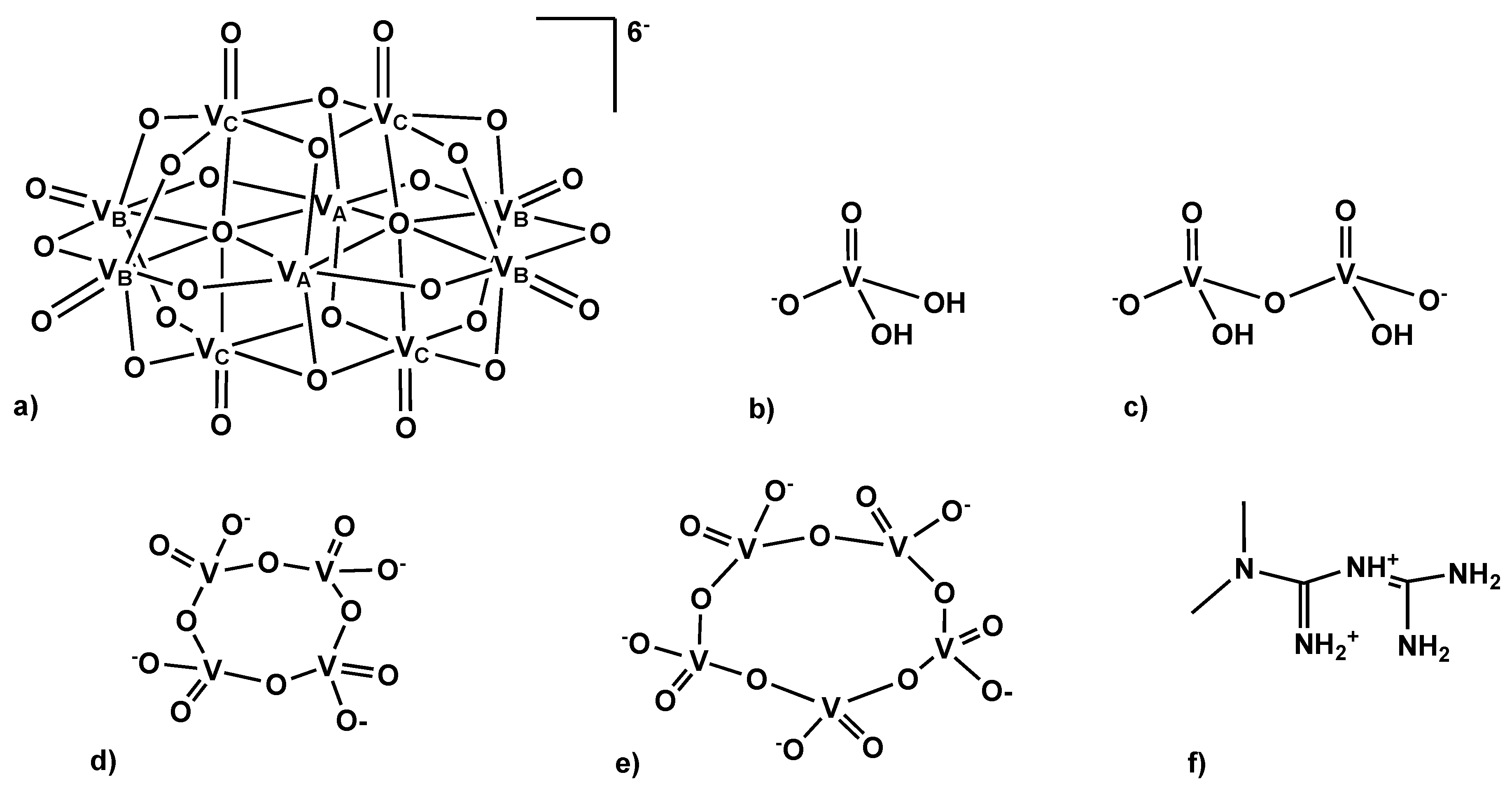
2.1. Characterization of the Sodium and Metforminium Decavanadate Solutions
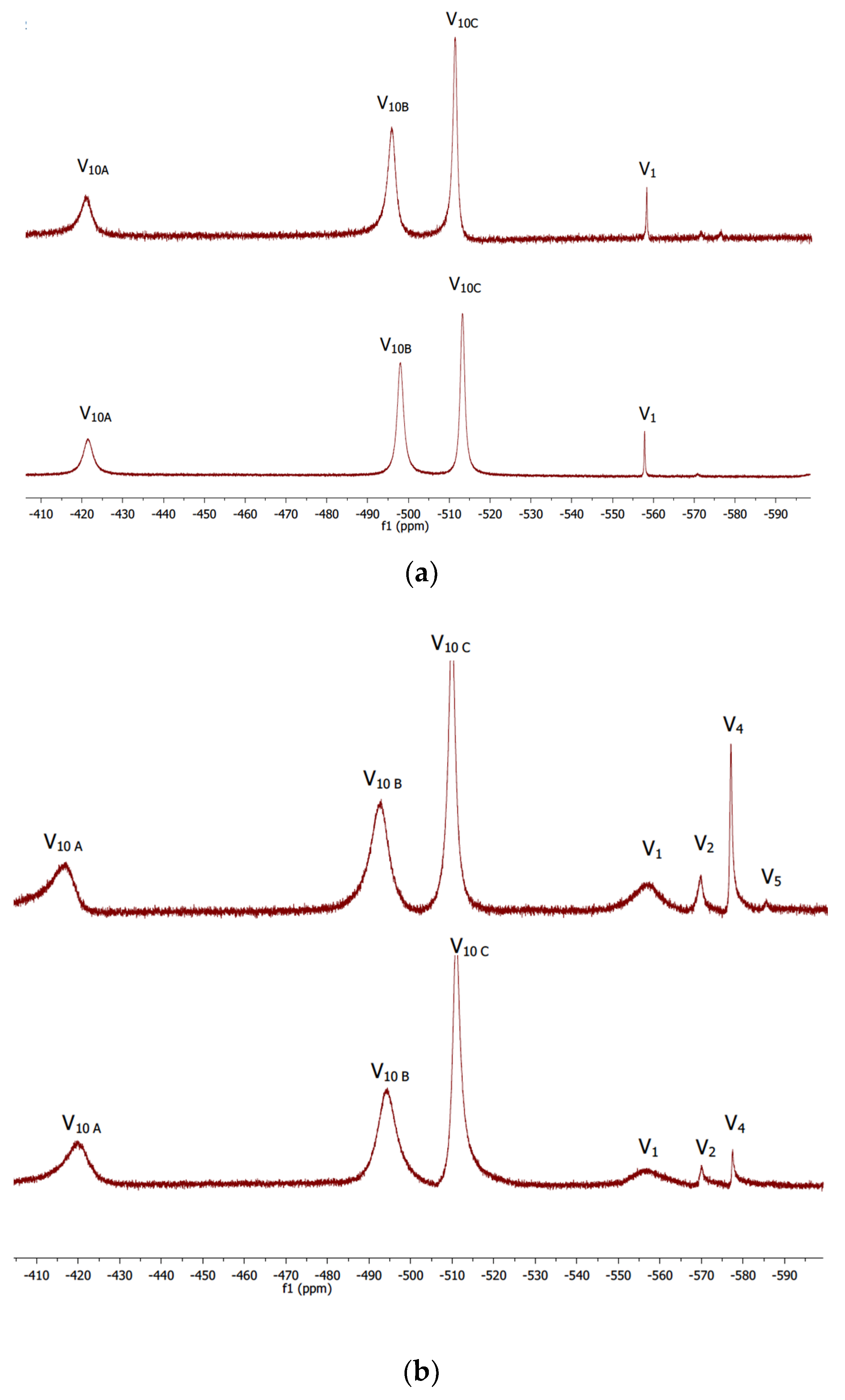
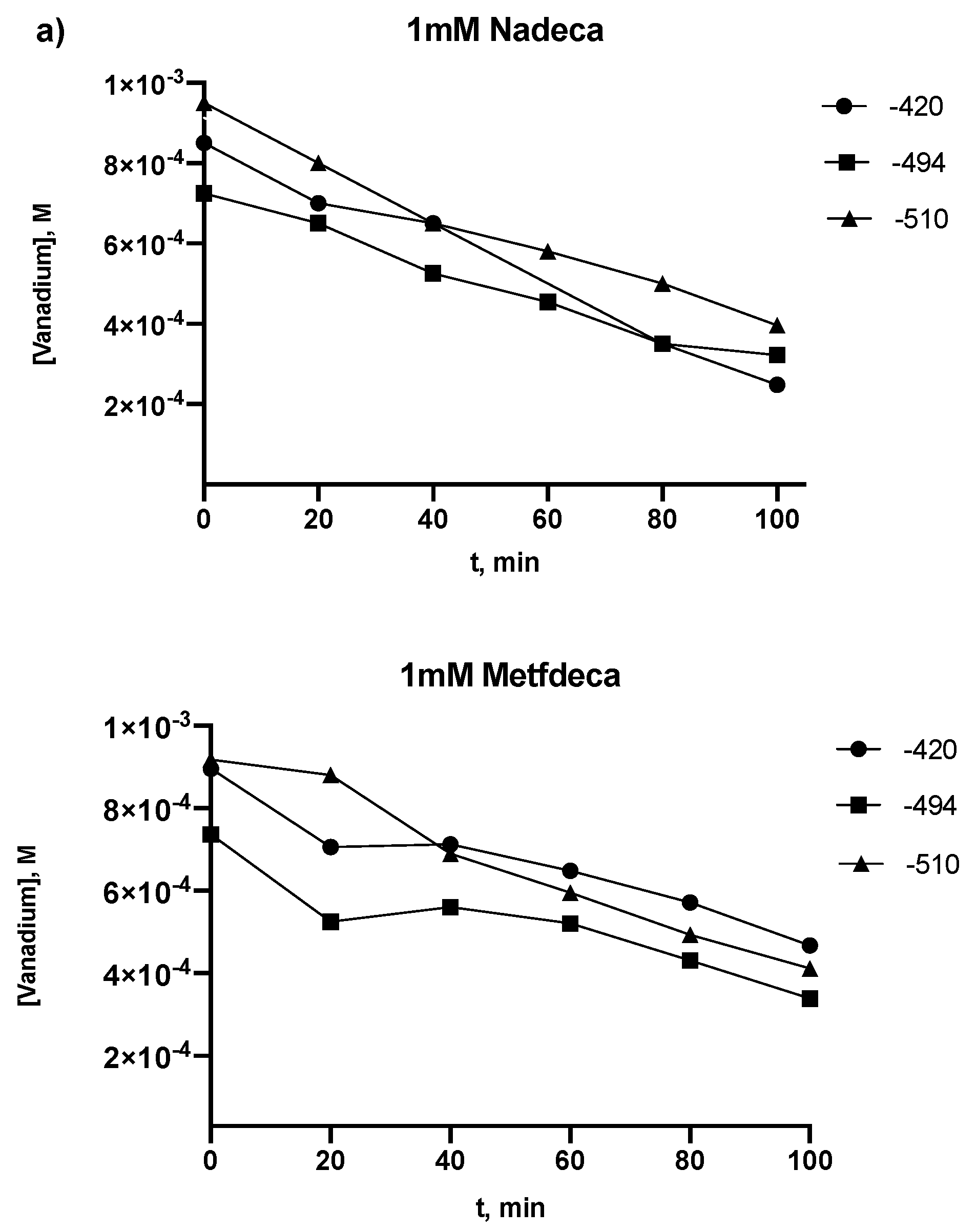
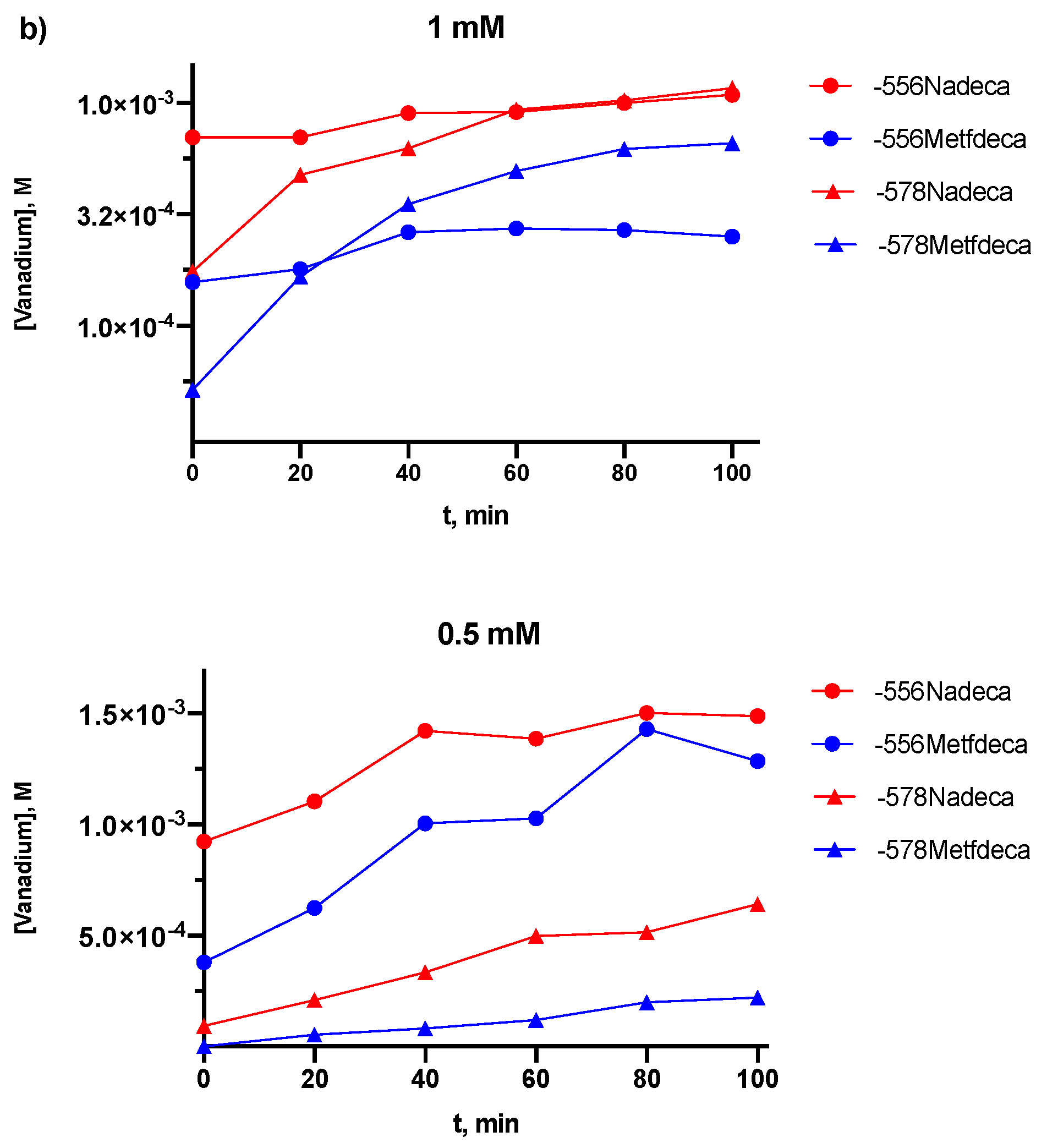
2.2. Kinetic Studies by 51V NMR
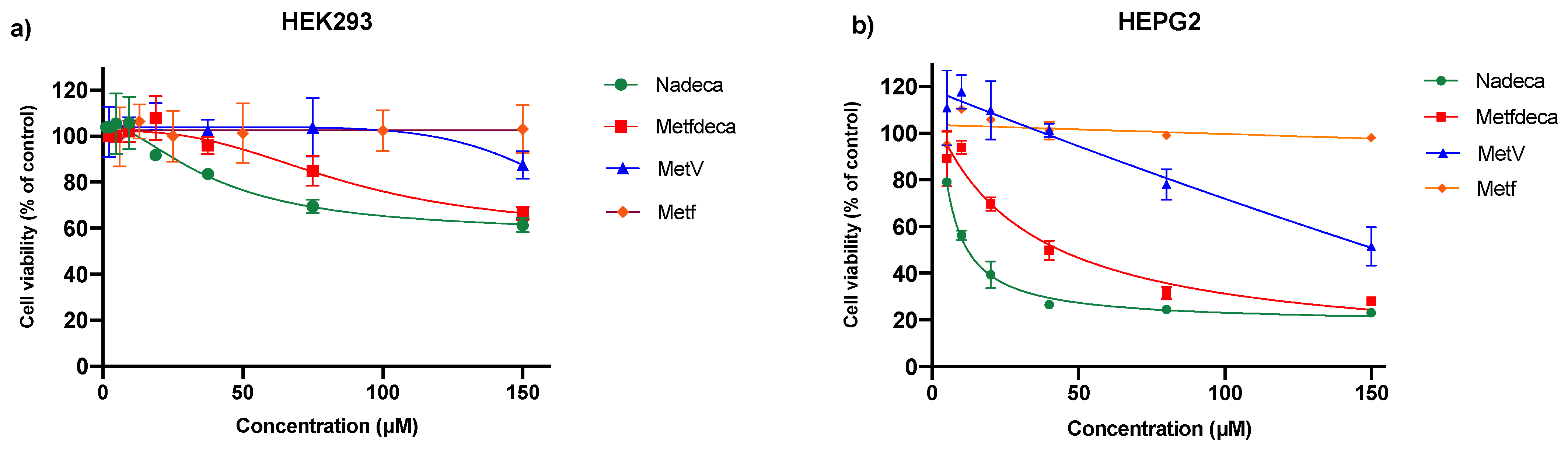
2.3. Cell Viability
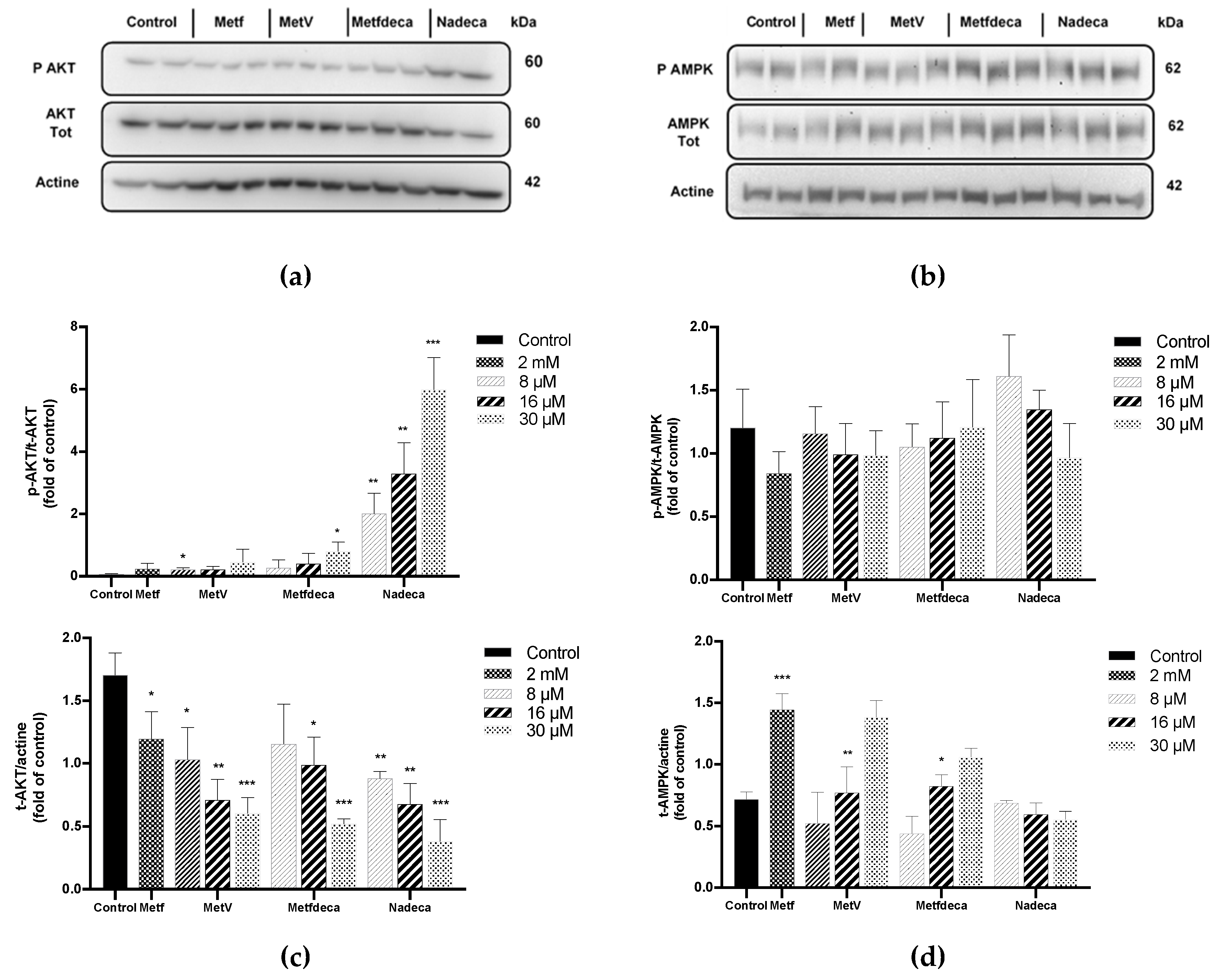
2.4. Proteins Expression
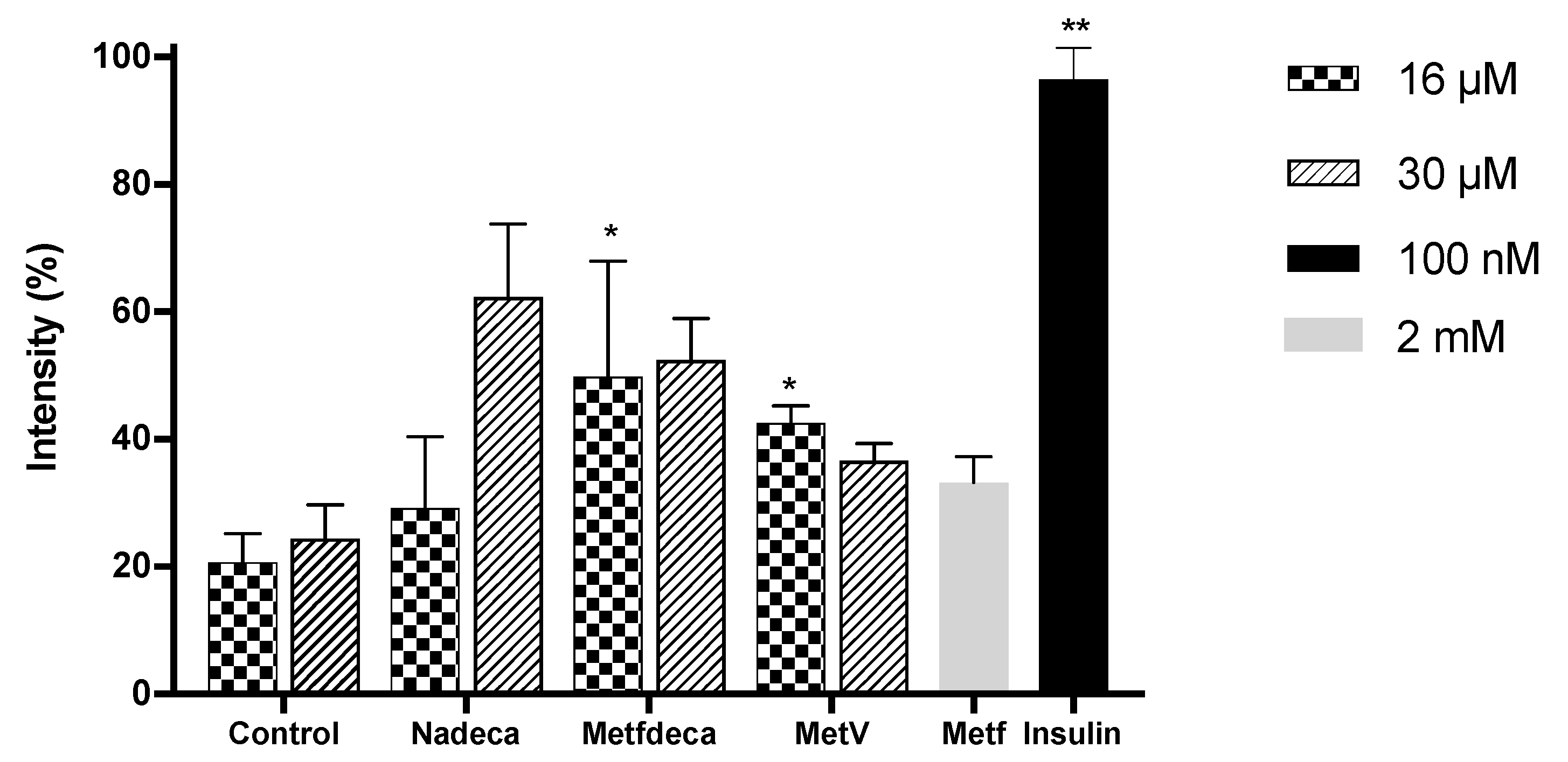
2.5. Glucose Uptake Assay
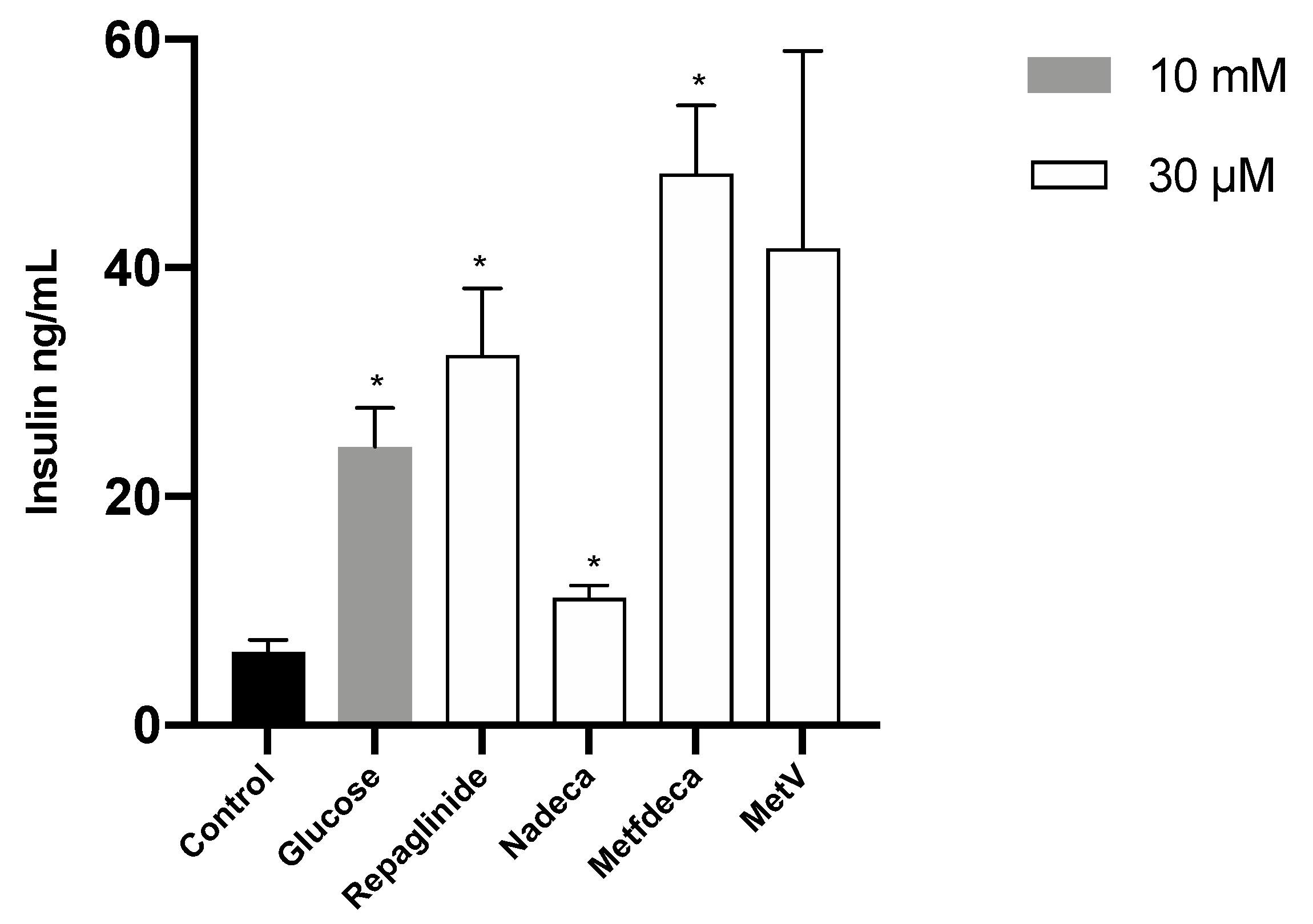
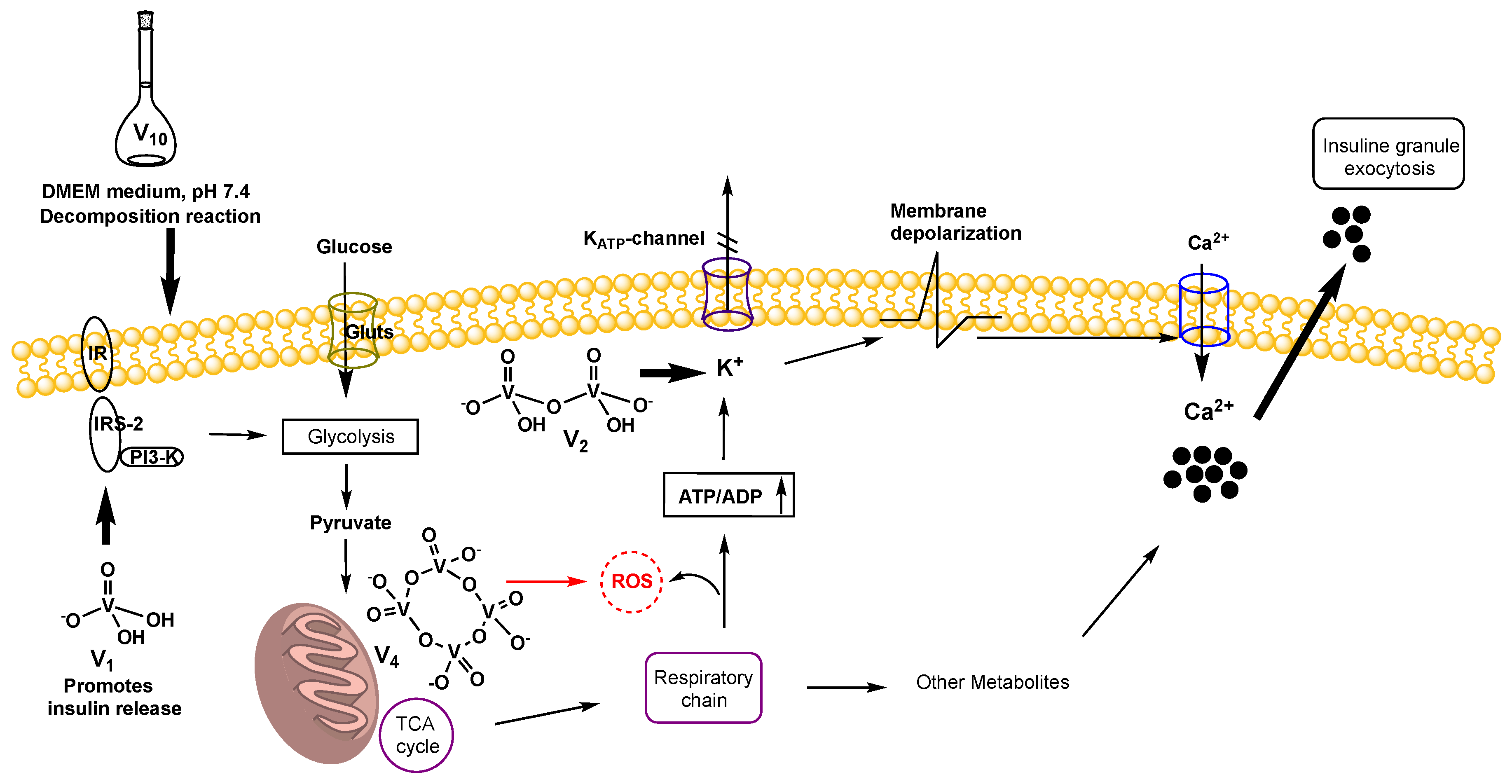
2.6. Insulin Release Assay
3. Experimental
3.1. Chemicals and Reagents
3.2. Kinetic Studies
3.3. Cell Viability Assay
3.4. Western Blot Analysis
3.5. Adipocyte Differentiation
3.6. Glucose Uptake Assay
3.7. Insulin Release Assay
3.8. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Rompuy, L.S.; Parac-Vogt, T.N. Interactions between polyoxometalates and biological systems: From drug design to artificial enzymes. Curr. Opin. Biotechnol. 2019, 58, 92–99. [Google Scholar] [CrossRef]
- Čolović, M.B.; Lacković, M.; Lalatović, J.; Mougharbel, A.S.; Kortz, U.; Krstić, D.Z. Polyoxometalates in Biomedicine: Update and Overview. Curr. Med. Chem. 2020, 27, 362–379. [Google Scholar] [CrossRef]
- Aureliano, M. Decavanadate contribution to vanadium biochemistry: In vitro and in vivo studies. Inorg. Chim. Acta 2014, 420, 4–7. [Google Scholar] [CrossRef]
- Bijelic, A.; Rompel, A. The use of polyoxometalates in protein crystallography—An attempt to widen a well-known bottleneck. Coord. Chem. Rev. 2015, 299, 22–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crans, D.C.; Sánchez-Lombardo, I.; McLauchlan, C.C. Exploring Wells-Dawson Clusters Associated with the Small Ribosomal Subunit. Front. Chem. 2019, 7, 462–476. [Google Scholar] [CrossRef] [PubMed]
- Aureliano, M.; Crans, D.C. Decavanadate (V10O286−) and oxovanadates: Oxometalates with many biological activities. J. Inorg. Biochem. 2009, 103, 536–546. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Ozeki, T. Hydrogen-bonded aggregates of protonated decavanadate anions in their tetraalkylammonium salts. J. Chem. Soc. Dalton Trans. 2001, 472–480. [Google Scholar] [CrossRef]
- Ferreira da Silva, J.L.; Fátima Minas da Piedade, M.; Teresa Duarte, M. Decavanadates: A building-block for supramolecular assemblies. Inorg. Chim. Acta 2003, 356, 222–242. [Google Scholar] [CrossRef]
- Bošnjaković-Pavlović, N.; Prévost, J.; Spasojević-de Biré, A. Crystallographic Statistical Study of Decavanadate Anion Based-Structures: Toward a Prediction of Noncovalent Interactions. Cryst. Growth Des. 2011, 11, 3778–3789. [Google Scholar] [CrossRef]
- Povar, I.; Spinu, O.; Zinicovscaia, I.; Pintilie, B.; Ubaldini, S. Revised Poubaix diagrams for the vanadium–water system. J. Electrochem. Sci. Eng. 2019, 9, 620. [Google Scholar] [CrossRef]
- Kojima, T.; Antonio, M.; Ozeki, T. Solvent-Driven Association and Dissociation of the Hydrogen-Bonded Protonated Decavanadates. J. Am. Chem. Soc. 2011, 133, 7248–7251. [Google Scholar] [CrossRef] [PubMed]
- Ramos, S.; Manuel, M.; Tiago, T.; Duarte, R.; Martins, J.; Gutiérrez-Merino, C.; Moura, J.J.G.; Aureliano, M. Decavanadate interactions with actin: Inhibition of G-actin polymerization and stabilization of decameric vanadate. J. Inorg. Biochem. 2006, 100, 1734–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samart, N.; Arhouma, Z.; Kumar, S.; Murakami, H.A.; Crick, D.C.; Crans, D.C. Decavanadate Inhibits Mycobacterial Growth More Potently Than Other Oxovanadates. Front. Chem. 2018, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krivosudský, L.; Roller, A.; Rompel, A. Tuning the interactions of decavanadate with thaumatin, lysozyme, proteinase K and human serum proteins by its coordination to a pentaaquacobalt(II) complex cation. New J. Chem. 2019, 43, 17863–17871. [Google Scholar] [CrossRef] [Green Version]
- Soares, S.S.; Martins, H.; Duarte, R.O.; Moura, J.J.G.; Coucelo, J.; Gutiérrez-Merino, C.; Aureliano, M. Vanadium distribution, lipid peroxidation and oxidative stress markers upon decavanadate in vivo administration. J. Inorg. Biochem. 2007, 101, 80–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clare, B.W.; Kepert, D.L.; Watts, D.W. Acid decomposition of decavanadate: Specific salt effects. J. Chem. Soc. Dalton Trans. 1973, 2481–2487. [Google Scholar] [CrossRef]
- Druskovich, D.M.; Kepert, D.L. Base decomposition of decavanadate. J. Chem. Soc. Dalton Trans. 1975, 947–951. [Google Scholar] [CrossRef]
- Crans, D.; Rithner, C.; Theisen, L. Application of time-resolved vanadium-51 2D NMR for quantitation of kinetic exchange pathways between vanadate monomer, dimer, tetramer, and pentamer. J. Am. Chem. Soc. 1990, 112, 2901–2908. [Google Scholar] [CrossRef]
- Levina, A.; McLeod, A.I.; Pulte, A.; Aitken, J.B.; Lay, P.A. Biotransformations of Antidiabetic Vanadium Prodrugs in Mammalian Cells and Cell Culture Media: A XANES Spectroscopic Study. Inorg. Chem. 2015, 54, 6707–6718. [Google Scholar] [CrossRef] [Green Version]
- Saklayen, M.G. The Global Epidemic of the Metabolic Syndrome. Curr. Hypertens. Rep. 2018, 20, 12. [Google Scholar] [CrossRef] [Green Version]
- Giovannucci, E.; Harlan, D.M.; Archer, M.C.; Bergenstal, R.M.; Gapstur, S.M.; Habel, L.A.; Pollak, M.; Regensteiner, J.G.; Yee, D. Diabetes and Cancer: A consensus report. Diabetes Care 2010, 33, 1674–1685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tschöp, M.H. Metabolic Disease Therapies. Cell Metab. 2017, 26, 579–583. [Google Scholar]
- Lee, C.-W.; Wong, L.L.-Y.; Tse, E.Y.-T.; Liu, H.-F.; Leong, V.Y.-L.; Lee, J.M.-F.; Hardie, D.G.; Ng, I.O.-L.; Ching, Y.-P. AMPK Promotes p53 Acetylation via Phosphorylation and Inactivation of SIRT1 in Liver Cancer Cells. Cancer Res. 2012, 72, 4394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koundouros, N.; Poulogiannis, G. Phosphoinositide 3-Kinase/Akt Signaling and Redox Metabolism in Cancer. Front. Oncol. 2018, 8, 160–169. [Google Scholar] [CrossRef]
- Zheng, W.; Feng, Q.; Liu, J.; Guo, Y.; Gao, L.; Li, R.; Xu, M.; Yan, G.; Yin, Z.; Zhang, S.; et al. Inhibition of 6-phosphogluconate Dehydrogenase Reverses Cisplatin Resistance in Ovarian and Lung Cancer. Front. Pharmacol. 2017, 8, 421–432. [Google Scholar] [CrossRef]
- Rehder, D. Implications of vanadium in technical applications and pharmaceutical issues. Inorg. Chim. Acta 2017, 455, 378–389. [Google Scholar] [CrossRef]
- Crans, D.C.; Henry, L.; Cardiff, G.; Posner, B.I. Developing Vanadium as an Antidiabetic or Anticancer Drug: A Clinical and Historical Perspective. In Essential Metals in Medicine: Therapeutic Use and Toxicity of Metal Ions in the Clinic, 1st ed.; Carver, P.L., Ed.; De Gruyter: Berlin, Germany; Boston, MA, USA, 2019; pp. 203–230. [Google Scholar]
- Jakusch, T.; Kiss, T. In vitro study of the antidiabetic behavior of vanadium compounds. Coord. Chem. Rev. 2017, 351, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Thompson, K.H.; Orvig, C. Vanadium in diabetes: 100 years from Phase 0 to Phase I. J. Inorg. Biochem. 2006, 100, 1925–1935. [Google Scholar] [CrossRef]
- Thompson, K.H.; Lichter, J.; LeBel, C.; Scaife, M.C.; McNeill, J.H.; Orvig, C. Vanadium treatment of type 2 diabetes: A view to the future. J. Inorg. Biochem. 2009, 103, 554–558. [Google Scholar] [CrossRef]
- Rehder, D. Transport, Accumulation, and Physiological Effects of Vanadium. In Detoxification of Heavy Metals; Sherameti, I., Varma, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 205–220. [Google Scholar]
- Gruzewska, K.; Michno, A.; Pawelczyk, T.; Bielarczyk, H. Essentiality and toxicity of vanadium supplements in health and pathology. J. Physiol. Pharmacol. 2014, 65, 603–611. [Google Scholar]
- Sánchez Lombardo, I.; Sánchez Lara, E.; Pérez Benítez, A.; Mendoza, Á.; Bernès, S.; González Vergara, E. Synthesis of Metforminium(2+) Decavanadates-Crystal Structures and Solid-State Characterization. Eur. J. Inorg. Chem. 2014, 2014, 4581–4588. [Google Scholar] [CrossRef]
- Treviño, S.; González-Vergara, E. Metformin-decavanadate treatment ameliorates hyperglycemia and redox balance of the liver and muscle in a rat model of alloxan-induced diabetes. New J. Chem. 2019, 43, 17850–17862. [Google Scholar] [CrossRef]
- Treviño, S.; Sánchez-Lara, E.; Sarmiento-Ortega, V.E.; Sánchez-Lombardo, I.; Flores-Hernández, J.Á.; Pérez-Benítez, A.; Brambila-Colombres, E.; González-Vergara, E. Hypoglycemic, lipid-lowering and metabolic regulation activities of metforminium decavanadate (H2Metf)3 [V10O28]·8H2O using hypercaloric-induced carbohydrate and lipid deregulation in Wistar rats as biological model. J. Inorg. Biochem. 2015, 147, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Durif, A.; Averbuch-Pouchot, M.T.; Guitel, J.C. Structure d’un Decavanadate d’Hexasodium Hydrate. Acta Crystallogr. Sect. B Struct. Sci. 1980, 36, 680–682. [Google Scholar] [CrossRef] [Green Version]
- Howarth, O.W.; Jarrold, M. Protonation of the decavanadate(6-) ion: A vanadium-51 nuclear magnetic resonance study. J. Chem. Soc. Dalton Trans. 1978, 5, 503–506. [Google Scholar] [CrossRef]
- Goddard, J.B.; Gonas, A.M. Kinetics of the dissociation of decavanadate ion in basic solutions. Inorg. Chem. 1973, 12, 574–579. [Google Scholar] [CrossRef]
- Schwarzenbach, G.; Geier, G. 97. Die Raschacidifierung und -alkalisierung von Vanadaten. Helv. Chim. Acta 1963, 46, 906–926. [Google Scholar] [CrossRef]
- Pascale, F.; Bedouet, L.; Baylatry, M.; Namur, J.; Laurent, A. Comparative Chemosensitivity of VX2 and HCC Cell Lines to Drugs Used in TACE. Anticancer Res. 2015, 35, 6497–6503. [Google Scholar]
- Mohamadi, M.; Yousef Ebrahimipour, S.; Torkzadeh-Mahani, M.; Foro, S.; Akbari, A. A mononuclear diketone-based oxido-vanadium(IV) complex: Structure, DNA and BSA binding, molecular docking and anticancer activities against MCF-7, HPG-2, and HT-29 cell lines. RSC Adv. 2015, 5, 101063–101075. [Google Scholar] [CrossRef]
- Dhahagani, K.; Mathan Kumar, S.; Chakkaravarthi, G.; Anitha, K.; Rajesh, J.; Ramu, A.; Rajagopal, G. Synthesis and spectral characterization of Schiff base complexes of Cu(II), Co(II), Zn(II) and VO(IV) containing 4-(4-aminophenyl)morpholine derivatives: Antimicrobial evaluation and anticancer studies. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2014, 117, 87–94. [Google Scholar] [CrossRef]
- Correia, I.; Adão, P.; Roy, S.; Wahba, M.; Matos, C.; Maurya, M.R.; Marques, F.; Pavan, F.R.; Leite, C.Q.F.; Avecilla, F.; et al. Hydroxyquinoline derived vanadium(IV and V) and copper(II) complexes as potential anti-tuberculosis and anti-tumor agents. J. Inorg. Biochem. 2014, 141, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Amuthan, V.D.A.; Chandrashekara, S.S.; Venkata, R.; Richard, L. Cytoprotective Activity of Neichitti (Vernonia cinerea) in Human Embryonic Kidney (HEK293) Normal Cells and Human Cervix Epitheloid Carcinoma (HeLa) Cells against Cisplatin Induced Toxicity: A Comparative Study. J. Clin. Diagn. Res. 2019, 13, KC01–KC06. [Google Scholar] [CrossRef]
- Fujita, H.; Hirose, K.; Sato, M.; Fujioka, I.; Fujita, T.; Aoki, M.; Takai, Y. Metformin attenuates hypoxia-induced resistance to cisplatin in the HepG2 cell line. Oncol. Lett. 2019, 17, 2431–2440. [Google Scholar] [CrossRef] [PubMed]
- Cheng, M.; Li, N.; Wang, N.; Hu, K.; Xiao, Z.; Wu, P.; Wei, Y. Synthesis, structure and antitumor studies of a novel decavanadate complex with a wavelike two-dimensional network. Polyhedron 2018, 155, 313–319. [Google Scholar] [CrossRef]
- Li, Y.-T.; Zhu, C.-Y.; Wu, Z.-Y.; Jiang, M.; Yan, C.-W. Synthesis, crystal structures and anticancer activities of two decavanadate compounds. Transit. Met. Chem. 2010, 35, 597–603. [Google Scholar] [CrossRef]
- Missina, J.M.; Gavinho, B.; Postal, K.; Santana, F.S.; Valdameri, G.; de Souza, E.M.; Hughes, D.L.; Ramirez, M.I.; Soares, J.F.; Nunes, G.G. Effects of Decavanadate Salts with Organic and Inorganic Cations on Escherichia coli, Giardia intestinalis, and Vero Cells. Inorg. Chem. 2018, 57, 11930–11941. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, T.-T.; Fu, Y.; Wang, K.; Yang, X.-G. Vanadium compounds discriminate hepatoma and normal hepatic cells by differential regulation of reactive oxygen species. J. Biol. Inorg. Chem. 2010, 15, 1087–1097. [Google Scholar] [CrossRef]
- Hao, J.; Huang, K.; Chen, C.; Liang, Y.; Wang, Y.; Zhang, X.; Huang, H. Polydatin Improves Glucose and Lipid Metabolisms in Insulin-Resistant HepG2 Cells through the AMPK Pathway. Biol. Pharm. Bull. 2018, 41, 891–898. [Google Scholar] [CrossRef] [Green Version]
- Ferretti, A.C.; Hidalgo, F.; Tonucci, F.M.; Almada, E.; Pariani, A.; Larocca, M.C.; Favre, C. Metformin and glucose starvation decrease the migratory ability of hepatocellular carcinoma cells: Targeting AMPK activation to control migration. Sci. Rep. 2019, 9, 2815. [Google Scholar] [CrossRef]
- Hardie, D.G. Molecular Pathways: Is AMPK a Friend or a Foe in Cancer? Clin. Cancer Res. 2015, 21, 3836–3840. [Google Scholar] [CrossRef] [Green Version]
- Lu, B.; Ennis, D.; Lai, R.; Bogdanovic, E.; Nikolov, R.; Salamon, L.; Fantus, C.; Le-Tien, H.; Fantus, I.G. Enhanced sensitivity of insulin-resistant adipocytes to vanadate is associated with oxidative stress and decreased reduction of vanadate (+5) to vanadyl (+4). J. Biol. Chem. 2001, 276, 35589–35598. [Google Scholar] [CrossRef] [Green Version]
- Tardif, A.; Julien, N.; Chiasson, J.-L.; Coderre, L. Stimulation of glucose uptake by chronic vanadate pretreatment in cardiomyocytes requires PI 3-kinase and p38 MAPK activation. Am. J. Physiol. Endocrinol. Metab. 2003, 284, E1055–E1064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Žižić, M.; Miladinović, Z.; Stanić, M.; Hadžibrahimović, M.; Živić, M.; Zakrzewska, J. 51V NMR investigation of cell-associated vanadate species in Phycomyces blakesleeanus mycelium. Res. Microbiol. 2016, 167, 521–528. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Chen, D.; Liu, P.; Wei, T.; Zhang, F.; Ding, W. Oxidovanadium(IV) sulfate-induced glucose uptake in HepG2 cells through IR/Akt pathway and hydroxyl radicals. J. Inorg. Biochem. 2015, 149, 39–44. [Google Scholar] [CrossRef]
- Seale, A.P.; de Jesus, L.A.; Park, M.C.; Kim, Y.S. Vanadium and insulin increase adiponectin production in 3T3-L1 adipocytes. Pharmacol. Res. 2006, 54, 30–38. [Google Scholar] [CrossRef] [PubMed]
- Yraola, F.; García-Vicente, S.; Marti, L.; Albericio, F.; Zorzano, A.; Royo, M. Understanding the mechanism of action of the novel SSAO substrate (C7NH10)6(V10O28)∙2H2O, a prodrug of peroxovanadate insulin mimetics. Chem. Biol. Drug. Des. 2007, 69, 423–428. [Google Scholar] [CrossRef]
- Marín-Peñalver, J.J.; Martín-Timón, I.; Sevillano-Collantes, C.; Del Cañizo-Gómez, F.J. Update on the treatment of type 2 diabetes mellitus. World J. Diabetes 2016, 7, 354–395. [Google Scholar] [CrossRef]
- Skelin, M.; Rupnik, M.; Cencic, A. Pancreatic beta cell lines and their applications in diabetes mellitus research. Altex 2010, 27, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Zhang, A.Q.; Gao, Z.Y.; Gilon, P.; Nenquin, M.; Drews, G.; Henquin, J.C. Vanadate stimulation of insulin release in normal mouse islets. J. Biol. Chem. 1991, 266, 21649–21656. [Google Scholar]
- Proks, P.; Ashfield, R.; Ashcroft, F.M. Interaction of vanadate with the cloned beta cell K(ATP) channel. J. Biol. Chem. 1999, 274, 25393–25397. [Google Scholar] [CrossRef] [Green Version]
- Gogg, S.; Chen, J.; Efendic, S.; Smith, U.; Ostenson, C. Effects of phosphotyrosine phosphatase inhibition on insulin secretion and intracellular signaling events in rat pancreatic islets. Biochem. Biophys. Res. Commun. 2001, 280, 1161–1168. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ostenson, C.G. Inhibition of protein-tyrosine phosphatases stimulates insulin secretion in pancreatic islets of diabetic Goto-Kakizaki rats. Pancreas 2005, 30, 314–317. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-K.; Green, B.D.; McClenaghan, N.H.; McCluskey, J.T.; Flatt, P.R. Long-Term Beneficial Effects of Vanadate, Tungstate, and Molybdate on Insulin Secretion and Function of Cultured Beta Cells. Pancreas 2004, 28, 264–268. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Ye, L.; Liu, H.; Xia, Q.; Zhang, Y.; Yang, X.; Wang, K. Vanadium compounds induced mitochondria permeability transition pore (PTP) opening related to oxidative stress. J. Inorg. Biochem. 2010, 104, 371–378. [Google Scholar] [CrossRef]
- Hosseini, M.J.; Shaki, F.; Ghazi-Khansari, M.; Pourahmad, J. Toxicity of vanadium on isolated rat liver mitochondria: A new mechanistic approach. Metallomics 2013, 5, 152–166. [Google Scholar] [CrossRef]
- Soares, S.S.; Gutiérrez-Merino, C.; Aureliano, M. Mitochondria as a target for decavanadate toxicity in Sparus aurata heart. Aquat. Toxicol. 2007, 83, 1–9. [Google Scholar] [CrossRef]
- Crans, D.C.; Bunch, R.L.; Theisen, L.A. Interaction of trace levels of vanadium(IV) and vanadium(V) in biological systems. J. Am. Chem. Soc. 1989, 111, 7597–7607. [Google Scholar] [CrossRef]
- Griffin, E.; Levina, A.; Lay, P.A. Vanadium(V) tris-3,5-di-tert-butylcatecholato complex: Links between speciation and anti-proliferative activity in human pancreatic cancer cells. J. Inorg. Biochem. 2019, 201, 110815. [Google Scholar] [CrossRef]
- Aureliano, M.; Tiago, T.; Gândara, R.M.C.; Sousa, A.; Moderno, A.; Kaliva, M.; Salifoglou, A.; Duarte, R.O.; Moura, J.J.G. Interactions of vanadium(V)–citrate complexes with the sarcoplasmic reticulum calcium pump. J. Inorg. Biochem. 2005, 99, 2355–2361. [Google Scholar] [CrossRef] [Green Version]
- Sánchez-Lombardo, I.; Baruah, B.; Alvarez, S.; Werst, K.R.; Segaline, N.A.; Levinger, N.E.; Crans, D.C. Size and shape trump charge in interactions of oxovanadates with self-assembled interfaces: Application of continuous shape measure analysis to the decavanadate anion. New J. Chem. 2016, 40, 962–975. [Google Scholar] [CrossRef]
- Student, A.K.; Hsu, R.Y.; Lane, M.D. Induction of fatty acid synthetase synthesis in differentiating 3T3-L1 preadipocytes. J. Biol. Chem. 1980, 255, 4745–4750. [Google Scholar] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
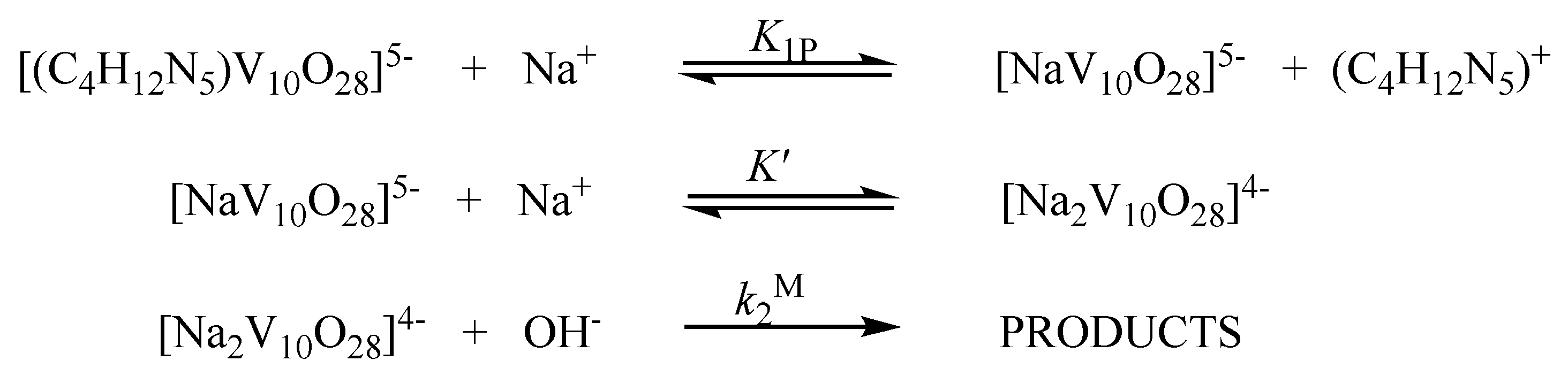{kind=link}
| Decavanadate Compound Concentration | 51V NMR Signal a | kobs, min−1 NaDeca | kobs, min−1 MetfDeca |
|---|---|---|---|
| 0.0005 M | −420 (V10A) | (−1.25 ± 0.03) × 10−3 | (−1.40 ± 0.03) × 10−3 |
| −494 (V10B) | (−1.26 ± 0.2) × 10−3 | (−1.48 ± 0.7) × 10−3 | |
| −510 (V10c) | (−1.82 ± 0.4) × 10−3 | (−1.44 ± 0.4) × 10−3 | |
| −556 (V1) | (4.59 ± 0.7) × 10−3 | (2.97 ± 0.4) × 10−3 | |
| −570 (V2) | (7.07 ± 0.7) × 10−4 | (6.17 ± 0.8) × 10−4 | |
| −578 (V4) | (2.28 ± 0.08) × 10−3 | (7.63 ± 0.8) × 10−4 | |
| −586 (V5) | (1.72 ± 0.07) × 10−4 | (4.09 ± 0.3) × 10−5 | |
| 0.001 M | −420 (V10A) | (−1.46 ± 0.7) × 10−3 | (−1.36 ± 0.2) × 10−3 |
| −494 (V10B) | (−1.13 ± 0.1) × 10−3 | (−8.39 ± 0.2) × 10−4 | |
| −510 (V10c) | (−1.16 ± 0.2) × 10−3 | (−9.49 ± 0.5) × 10−4 | |
| −556 (V1) | (2.62 ± 0.6) × 10−3 | (2.00 ± 0.2) × 10−3 | |
| −570 (V2) | (6.81 ± 0.4) × 10−4 | (5.34 ± 0.4) × 10−4 | |
| −578 (V4) | (1.96 ± 0.2) × 10−3 | (1.85 ± 0.09) × 10−3 | |
| −586 (V5) | (1.89 ± 0.1) × 10−4 | (1.38 ± 0.8) × 10−4 |
| Compound | HEK293 Lower Cell Viability (%) | HEK293 IC50 (µM) | HEPG2 Lower Cell Viability (%) | HEPG2 IC50 (µM) | SI (IC50 HEK293/(IC50 HEPG2) |
|---|---|---|---|---|---|
| NaDeca | 61 | 40 ± 4 | 22 | 9 ± 0.7 | 4.4 |
| MetfDeca | 67 | 85 ± 5 | 28 | 29 ± 0.7 | 2.9 |
| MetV | 88 | 181 ± 7 | 51 | 93 ± 5 | 1.9 |
| Metf | 100 | 420 ± 11 | 98 | 540 ± 4 | 0.77 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva-Nolasco, A.M.; Camacho, L.; Saavedra-Díaz, R.O.; Hernández-Abreu, O.; León, I.E.; Sánchez-Lombardo, I. Kinetic Studies of Sodium and Metforminium Decavanadates Decomposition and In Vitro Cytotoxicity and Insulin- Like Activity. Inorganics 2020, 8, 67. https://doi.org/10.3390/inorganics8120067
Silva-Nolasco AM, Camacho L, Saavedra-Díaz RO, Hernández-Abreu O, León IE, Sánchez-Lombardo I. Kinetic Studies of Sodium and Metforminium Decavanadates Decomposition and In Vitro Cytotoxicity and Insulin- Like Activity. Inorganics. 2020; 8(12):67. https://doi.org/10.3390/inorganics8120067
Chicago/Turabian StyleSilva-Nolasco, Aniela M., Luz Camacho, Rafael Omar Saavedra-Díaz, Oswaldo Hernández-Abreu, Ignacio E. León, and Irma Sánchez-Lombardo. 2020. "Kinetic Studies of Sodium and Metforminium Decavanadates Decomposition and In Vitro Cytotoxicity and Insulin- Like Activity" Inorganics 8, no. 12: 67. https://doi.org/10.3390/inorganics8120067
APA StyleSilva-Nolasco, A. M., Camacho, L., Saavedra-Díaz, R. O., Hernández-Abreu, O., León, I. E., & Sánchez-Lombardo, I. (2020). Kinetic Studies of Sodium and Metforminium Decavanadates Decomposition and In Vitro Cytotoxicity and Insulin- Like Activity. Inorganics, 8(12), 67. https://doi.org/10.3390/inorganics8120067