Bioactive Compounds in Wild Nettle (Urtica dioica L.) Leaves and Stalks: Polyphenols and Pigments upon Seasonal and Habitat Variations

 ,
,  ,
,  ,
,  ,
, 

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Plant Material
2.3. Extraction Conditions
2.4. UPLC-MS/MS Conditions
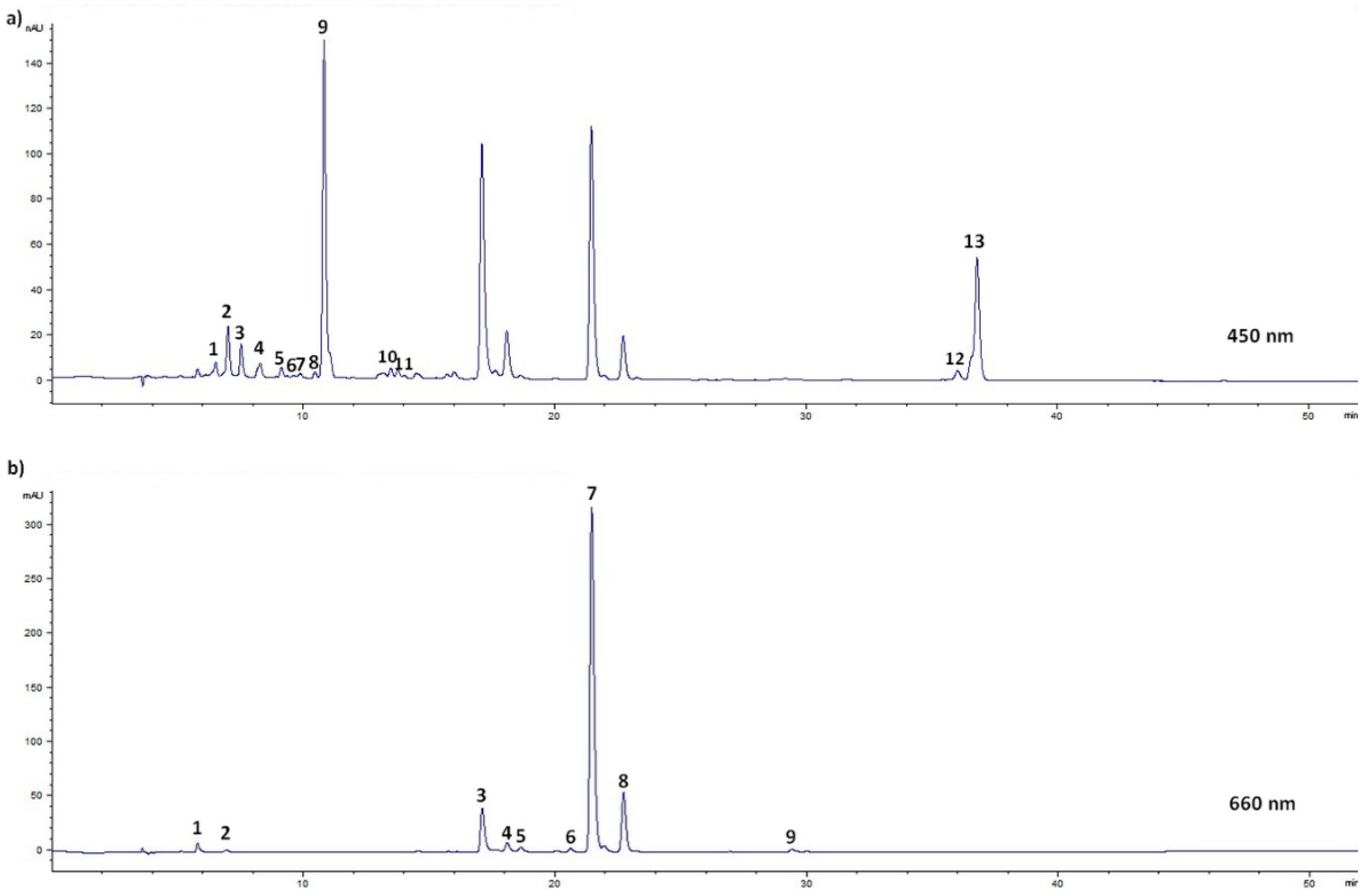
2.5. HPLC-UV-VIS/PDA Conditions
2.6. ORAC Determination
2.7. Statistical Analysis
3. Results and Discussion
3.1. Influence of Phenological Stage and Habitat on Polyphenols in Nettle Leaves and Stalks
3.2. Influence of Phenological Stage and Habitat on Pigments in Nettle Leaves and Stalks
3.3. Influence of Phenological Stage and Habitat on Antioxidant Capacity in Nettle Leaves and Stalks
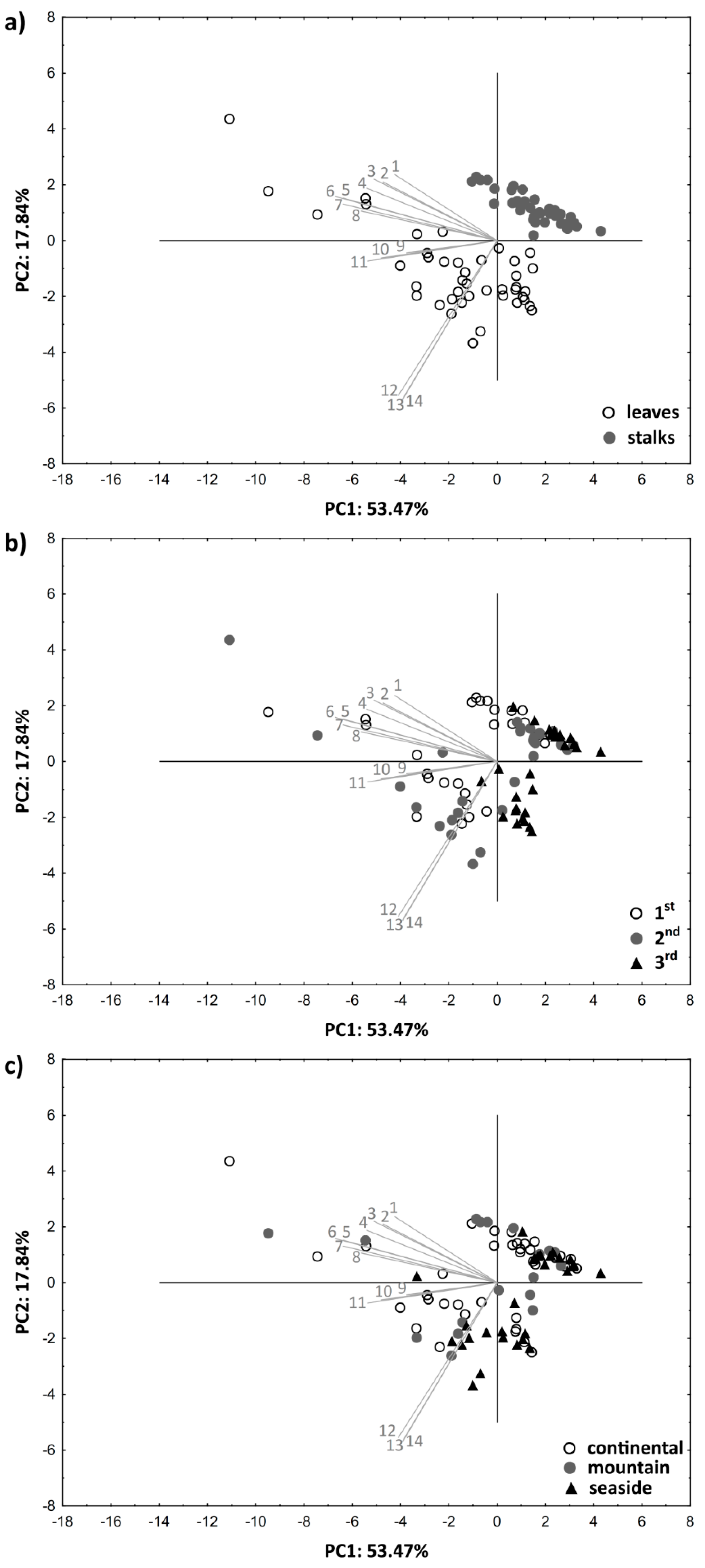
3.4. PCA Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Di Virgilio, N.; Papazoglou, E.G.; Jankauskiene, Z.; Di Lonardo, S.; Praczyk, M.; Wielgusz, K. The potential of stinging nettle (Urtica dioica L.) as a crop with multiple uses. Ind. Crop. Prod. 2015, 68, 42–49. [Google Scholar] [CrossRef]
- Kregiel, D.; Pawlikowska, E.; Antolak, H. Urtica spp.: Ordinary plants with extraordinary properties. Molecules 2018, 23, 1664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upton, R. Stinging nettles leaf (Urtica dioica L.): Extraordinary vegetable medicine. J. Herb. Med. 2013, 3, 9–38. [Google Scholar] [CrossRef]
- Guil-Guerrero, J.L.; Rebolloso-Fuentes, M.M.; Isasa, M.E.T. Fatty acids and carotenoids from Stinging Nettle (Urtica dioica L.). J. Food Compos. Anal. 2003, 16, 111–119. [Google Scholar] [CrossRef]
- Sajfrtová, M.; Sovová, H.; Opletal, L.; Bártlová, M. Near-critical extraction of β-sitosterol and scopoletin from stinging nettle roots. J. Supercrit. Fluids 2005, 35, 111–118. [Google Scholar] [CrossRef]
- Pinelli, P.; Ieri, F.; Vignolini, P.; Bacci, L.; Baronti, S.; Romani, A. Extraction and HPLC analysis of phenolic compounds in leaves, stalks, and textile fibers of Urtica dioica L. J. Agric. Food Chem. 2008, 56, 9127–9132. [Google Scholar] [CrossRef]
- Otles, S.; Yalcin, B. Phenolic compounds analysis of root, stalk, and leaves of nettle. Sci. World J. 2012, 2012. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, A.R.; Costa, G.; Figueirinha, A.; Liberal, J.; Prior, J.A.V.; Lopes, M.C.; Cruz, M.T.; Batista, M.T. Urtica spp.: Phenolic composition, safety, antioxidant and anti-inflammatory activities. Food Res. Int. 2017, 99, 485–494. [Google Scholar] [CrossRef]
- Đurović, S.; Pavlić, B.; Šorgić, S.; Popov, S.; Savić, S.; Pertonijević, M.; Radojković, M.; Cvetanović, A.; Zeković, Z. Chemical composition of stinging nettle leaves obtained by different analytical approaches. J. Funct. Foods 2017, 32, 18–26. [Google Scholar] [CrossRef]
- Marchetti, N.; Bonetti, G.; Brandolini, V.; Cavazzini, A.; Maietti, A.; Meca, G.; Mañes, J. Stinging nettle (Urtica dioica L.) as a functional food additive in egg pasta: Enrichment and bioaccessibility of Lutein and β-carotene. J. Funct. Foods 2018, 47, 547–553. [Google Scholar] [CrossRef]
- Repajić, M.; Cegledi, E.; Kruk, V.; Pedisić, S.; Çınar, F.; Bursać Kovačević, D.; Žutić, I.; Dragović-Uzelac, V. Accelerated Solvent Extraction as a Green Tool for the Recovery of Polyphenols and Pigments from Wild Nettle Leaves. Processes 2020, 8, 803. [Google Scholar] [CrossRef]
- Dhouibi, R.; Affes, H.; Salem, M.B.; Hammami, S.; Sahnoun, Z.; Zeghal, K.M.; Ksouda, K. Screening of pharmacological uses of Urtica dioica and others benefits. Prog. Biophys. Mol. Biol. 2020, 150, 67–77. [Google Scholar] [CrossRef] [PubMed]
- Wagner, H.; Willer, F.; Samtleben, R.; Boos, G. Search for the antiprostatic principle of stinging nettle (Urtica dioica) roots. Phytomedicine 1994, 1, 213–224. [Google Scholar] [CrossRef]
- Orcic, D.; Franciškovic, M.; Bekvalac, K.; Svircev, E.; Beara, I.; Lesjak, M.; Mimica-Dukic, N. Quantitative determination of plant phenolics in Urtica dioica extracts by high-performance liquid chromatography coupled with tandem mass spectrometric detection. Food Chem. 2014, 143, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Chrubasik, J.E.; Roufogalis, B.D.; Wagner, H.; Chrubasik, S. A comprehensive review on the stinging nettle effect and efficacy profiles. Part II: Urticae radix. Phytomedicine 2007, 14, 568–579. [Google Scholar] [CrossRef] [PubMed]
- Brown, D. Encyclopedia of herbs and their uses. In Encyclopedia of Common Natural Ingredients Used in Food, Drugs, and Cosmetics; Leung, Y.A., Foster, S., Eds.; Wiley: Hoboken, NJ, USA, 1995. [Google Scholar]
- Mottaleb, M.A.; Sarker, S.D. Accelerated solvent extraction for natural products isolation. In Natural Products Isolation; Springer: Berlin/Heidelberg, Germany, 2012; pp. 75–87. [Google Scholar]
- Hojnik, M.; Škerget, M.; Knez, Ž. Isolation of chlorophylls from stinging nettle (Urtica dioica L.). Sep. Purif. Technol. 2007, 57, 37–46. [Google Scholar] [CrossRef]
- Kőszegi, K.; Békássy-Molnár, E.; Koczka, N.; Kerner, T.; Stefanovits-Bányai, É. Changes in Total Polyphenol Content and Antioxidant Capacity of Stinging Nettle (Urtica dioica L.) from Spring to Autumn. Period. Polytech. Chem. Eng. 2020, 64, 548–554. [Google Scholar] [CrossRef]
- Ncube, B.; Van Staden, J. Tilting plant metabolism for improved metabolite biosynthesis and enhanced human benefit. Molecules 2015, 20, 12698–12731. [Google Scholar] [CrossRef] [Green Version]
- Moore, M. Medicinal Plants of the Mountain West; Museum of New Mexico Press: Santa Fe, NM, USA, 2003; ISBN 0890134545. [Google Scholar]
- Tolić, M.T.; Krbavčić, I.P.; Vujević, P.; Milinović, B.; Jurčević, I.L.; Vahčić, N. Effects of weather conditions on phenolic content and antioxidant capacity in juice of chokeberries (Aronia melanocarpa L.). Pol. J. Food Nutr. Sci. 2017, 67, 67–74. [Google Scholar] [CrossRef]
- González-Talice, J.; Yuri, J.A.; del Pozo, A. Relations among pigments, color and phenolic concentrations in the peel of two Gala apple strains according to canopy position and light environment. Sci. Hortic. 2013, 151, 83–89. [Google Scholar] [CrossRef]
- Saini, R.K.; Nile, S.H.; Park, S.W. Carotenoids from fruits and vegetables: Chemistry, analysis, occurrence, bioavailability and biological activities. Food Res. Int. 2015, 76, 735–750. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis: Changes in Official Methods of Analysis Made at the Annual Meeting. Supplement; Association of Official Analytical Chemists: Rockville, MD, USA, 1990; Volume 15. [Google Scholar]
- Elez Garofulić, I.; Zorić, Z.; Pedisić, S.; Brnčić, M.; Dragović-Uzelac, V. UPLC-MS2 Profiling of Blackthorn Flower Polyphenols Isolated by Ultrasound-Assisted Extraction. J. Food Sci. 2018, 83, 2782–2789. [Google Scholar] [CrossRef] [PubMed]
- Castro-Puyana, M.; Pérez-Sánchez, A.; Valdés, A.; Ibrahim, O.H.M.; Suarez-Álvarez, S.; Ferragut, J.A.; Micol, V.; Cifuentes, A.; Ibáñez, E.; García-Cañas, V. Pressurized liquid extraction of Neochloris oleoabundans for the recovery of bioactive carotenoids with anti-proliferative activity against human colon cancer cells. Food Res. Int. 2017, 99, 1048–1055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sozgen Baskan, K.; Tutem, E.; Ozer, N.; Apak, R. Spectrophotometric and chromatographic assessment of contributions of carotenoids and chlorophylls to the total antioxidant capacities of plant foods. J. Agric. Food Chem. 2013, 61, 11371–11381. [Google Scholar] [CrossRef]
- Gupta, P.; Sreelakshmi, Y.; Sharma, R. A rapid and sensitive method for determination of carotenoids in plant tissues by high performance liquid chromatography. Plant Methods 2015, 11, 5. [Google Scholar] [CrossRef] [Green Version]
- Prior, R.L.; Wu, X.; Schaich, K. Standardized methods for the determination of antioxidant capacity and phenolics in foods and dietary supplements. J. Agric. Food Chem. 2005, 53, 4290–4302. [Google Scholar] [CrossRef]
- Bender, C.; Graziano, S.; Zimmerman, B.F.; Weidlich, H.H. Antioxidant potential of aqueous plant extracts assessed by the cellular antioxidant activity assay. Am. J. Biol. Life Sci. 2014, 2, 72–79. [Google Scholar]
- Gruz, J.; Novák, O.; Strnad, M. Rapid analysis of phenolic acids in beverages by UPLC-MS/MS. Food Chem. 2008, 111, 789–794. [Google Scholar] [CrossRef]
- Parejo, I.; Jauregui, O.; Sánchez-Rabaneda, F.; Viladomat, F.; Bastida, J.; Codina, C. Separation and characterization of phenolic compounds in fennel (Foeniculum vulgare) using liquid chromatography-negative electrospray ionization tandem mass spectrometry. J. Agric. Food Chem. 2004, 52, 3679–3687. [Google Scholar] [CrossRef]
- Francišković, M.; Gonzalez-Pérez, R.; Orčić, D.; de Medina, F.S.; Martínez-Augustin, O.; Svirčev, E.; Simin, N.; Mimica-Dukić, N. Chemical Composition and Immuno-Modulatory Effects of Urtica dioica L. (Stinging Nettle) Extracts. Phyther. Res. 2017, 31, 1183–1191. [Google Scholar] [CrossRef]
- Zeković, Z.; Cvetanović, A.; Švarc-Gajić, J.; Gorjanović, S.; Sužnjević, D.; Mašković, P.; Savić, S.; Radojković, M.; Đurović, S. Chemical and biological screening of stinging nettle leaves extracts obtained by modern extraction techniques. Ind. Crop. Prod. 2017, 108, 423–430. [Google Scholar] [CrossRef]
- Chen, Y.; Yu, H.; Wu, H.; Pan, Y.; Wang, K.; Jin, Y.; Zhang, C. Characterization and Quantification by LC-MS/MS of the Chemical Components of the Heating Products of the Flavonoids Extract in Pollen Typhae for Transformation Rule Exploration. Molecules 2015, 20, 18352–18366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bucar, F.; Britzmann, B.; Streit, B.; Weigend, M. LC-PDA-MS-profiles of phenolic compounds in extracts of aerial parts of Urtica species. Planta Med. 2006, 72. [Google Scholar] [CrossRef]
- Pinheiro, P.F.; Justino, G.C. Structural Analysis of Flavonoids and Related Compounds—A Review of Spectroscopic Applications. In Phytochemicals—A Global Perspective of Their Role in Nutrition and Health; Rao, V., Ed.; InTech: Rijeka, Croatia, 2012; pp. 33–56. [Google Scholar]
- Farag, M.A.; Weigend, M.; Luebert, F.; Brokamp, G.; Wessjohann, L.A. Phytochemical, phylogenetic, and anti-inflammatory evaluation of 43 Urtica accessions (stinging nettle) based on UPLC–Q-TOF-MS metabolomic profiles. Phytochemistry 2013, 96, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Proestos, C.; Boziaris, I.S.; Nychas, G.-J.; Komaitis, M. Analysis of flavonoids and phenolic acids in Greek aromatic plants: Investigation of their antioxidant capacity and antimicrobial activity. Food Chem. 2006, 95, 664–671. [Google Scholar] [CrossRef]
- Nencu, I.; Vlase, L.; Istudor, V.; Duţu, L.E.; Gird, C.E. Preliminary research regarding the therapeutic uses of Urtica dioica L. Note I. The polyphenols evaluation. Farmacia 2012, 60, 493–500. [Google Scholar]
- Wang, F.; Jiang, K.; Li, Z. Purification and Identification of Genistein in Ginkgo biloba Leaf Extract. Chin. J. Chromatogr. 2007, 25, 509–513. [Google Scholar] [CrossRef]
- Mazimba, O. Umbelliferone: Sources, chemistry and bioactivities review. Bull. Fac. Pharm. Cairo Univ. 2017, 55, 223–232. [Google Scholar] [CrossRef]
- Ioana, N.; Viorica, I.; Diana-Carolina, I.; Valeria, R. Preliminary research regarding the therapeutic uses of Urtica dioica l note ii. The dynamics of accumulation of total phenolic compounds and ascorbic acid. Farmacia 2013, 61, 276–283. [Google Scholar]
- Roslon, W.; Weglarz, Z. Polyphenolic acids of female and male forms of Urtica dioica. In Proceedings of the International Conference on Medicinal and Aromatic Plants (Part II), Budapest, Hungary, 8–11 July 2001; pp. 101–104. [Google Scholar]
- Biesiada, A.; Kucharska, A.; Sokół-Łętowska, A.; Kuś, A. Effect of the Age of Plantation and Harvest Term on Chemical Composition and Antioxidant Avctivity of Stinging Nettle (Urtica dioica L.). Ecol. Chem. Eng. A 2010, 17, 1061–1068. [Google Scholar]
- Biesiada, A.; Wołoszczak, E.; Sokόł-Łętowska, A.; Kucharska, A.Z.; Nawirska-Olszańska, A. The effect of nitrogen form and dose on yield, chemical composition and antioxidant activity of stinging nettle (Urtica dioica L.). Herba Pol. 2009, 55, 84–93. [Google Scholar]
- Naser Aldeen, M.G.; Mansoor, R.; AlJoubbeh, M. Fluctuations of phenols and flavonoids in infusion of lemon verbena (Lippia citriodora) dried leaves during growth stages. Nutr. Food Sci. 2015, 45, 766–773. [Google Scholar] [CrossRef]
- Zeipiņa, S.; Alsiņa, I.; Lepse, L. Stinging nettle—The source of biologically active compounds as sustainable daily diet supplement. Res. Rural Dev. 2014, 20, 34–38. [Google Scholar]
- Kong, D.X.; Li, Y.Q.; Wang, M.L.; Bai, M.; Zou, R.; Tang, H.; Wu, H. Effects of light intensity on leaf photosynthetic characteristics, chloroplast structure, and alkaloid content of Mahonia bodinieri (Gagnep.) Laferr. Acta Physiol. Plant. 2016, 38, 120. [Google Scholar] [CrossRef]
- Pajevic, S.; Krstic, B.; Katic, S.; Nikolic, N.; Mihailovic, V. Some photosynthetic parameters of alfalfa (Medicago sativa L.) leaves at different phenological stages and in different cuttings. Matica Srp. Proc. Nat. Sci. 1999, 35. Available online: http://www.maticasrpska.org.rs/stariSajt/casopisi/prirodne_nauke_097.pdf#page=38 (accessed on 18 January 2021).
- Cândido, T.L.N.; Silva, M.R.; Agostini-Costa, T.S. Bioactive compounds and antioxidant capacity of buriti (Mauritia flexuosa Lf) from the Cerrado and Amazon biomes. Food Chem. 2015, 177, 313–319. [Google Scholar] [CrossRef]
- Česlová, L.; Šilarová, P.; Fischer, J. Effect of harvesting and processing of stinging nettle on the antioxidant capacity of its infusions. Sci. Pap. Univ. Pardubice Ser. A Fac. Chem. Technol. 2016, 22, 23–33. [Google Scholar]
- Kırca, A.; Arslan, E. Antioxidant capacity and total phenolic content of selected plants from Turkey. Int. J. Food Sci. Technol. 2008, 43, 2038–2046. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Region | Location | Altitude/ Latitude/Longitude | Weather Parameters | Phenological Stage | |||||
|---|---|---|---|---|---|---|---|---|---|
| I Before Flowering | II During Flowering | III After Flowering | |||||||
| April | May | June | July | September | October | ||||
| Continental | Sela Žakanjska | 244 m 45°36′27.80′′N 15°20′38.21′′E | a.d. T (°C) | 11.0 | 13.4 | 22.6 | 22.0 | 16.3 | 12.7 |
| T min (°C) | −0.1 | 0.4 | 11.4 | 9.2 | 2.9 | 0.7 | |||
| T max (°C) | 27.7 | 26.5 | 34.7 | 35.6 | 30.5 | 26.7 | |||
| a.p. (mm) | 143.4 | 170.1 | 73.8 | 85.4 | 101.8 | 55.6 | |||
| Sopčić Vrh | 177 m 45°34′14.88′′N 15°20′24.98′′E | a.d. T (°C) | 11.0 | 13.4 | 22.6 | 22.0 | 16.3 | 12.7 | |
| T min (°C) | −0.1 | 0.4 | 11.4 | 9.2 | 2.9 | 0.7 | |||
| T max (°C) | 27.7 | 26.5 | 34.7 | 35.6 | 30.5 | 26.7 | |||
| a.p. (mm) | 143.4 | 170.1 | 73.8 | 85.4 | 101.8 | 55.6 | |||
| Žakanje | 178 m 45°36′34.38′′N 15°20′14.96′′E | a.d. T (°C) | 11.0 | 13.4 | 22.6 | 22.0 | 16.3 | 12.7 | |
| T min (°C) | −0.1 | 0.4 | 11.4 | 9.2 | 2.9 | 0.7 | |||
| T max (°C) | 27.7 | 26.5 | 34.7 | 35.6 | 30.5 | 26.7 | |||
| a.p. (mm) | 143.4 | 170.1 | 73.8 | 85.4 | 101.8 | 55.6 | |||
| Zagreb I (Gračani) | 119 m 45°51′31.10′′N 15°58′19.34′′E | a.d. T (°C) | 12.4 | 13.7 | 23.8 | 22.9 | 17.2 | 13.2 | |
| T min (°C) | 1.9 | 2.1 | 13.3 | 10.4 | 4.6 | 2.2 | |||
| T max (°C) | 27.1 | 26.1 | 34.6 | 35.9 | 33.1 | 25.9 | |||
| a.p. (mm) | 81.1 | 147.7 | 70.8 | 76.8 | 150.1 | 42.3 | |||
| Zagreb II (Vrapče) | 119 m 45°49′8.69′′N 15°52′49.84′′E | a.d. T (°C) | 13.6 | 14.3 | 24.8 | 24.1 | 18.4 | 14.8 | |
| T min (°C) | 5.4 | 5.6 | 15.1 | 14.0 | 8.5 | 5.2 | |||
| T max (°C) | 22.9 | 27.1 | 27.3 | 35.5 | 32.7 | 24.6 | |||
| a.p. (mm) | 85.2 | 123.1 | 83.9 | 65.8 | 131.6 | 39.5 | |||
| Koretići | 410 m 45°48′47.23′′N 15°33′36.18′′E | a.d. T (°C) | 9.0 | 9.9 | 20.4 | 19.6 | 14.5 | 13.3 | |
| T min (°C) | −3.0 | −1.8 | 9.8 | 7.2 | 1.5 | 3.5 | |||
| T max (°C) | 22.4 | 21.5 | 31.2 | 30.0 | 28.2 | 22.8 | |||
| a.p. (mm) | 135.7 | 283.7 | 81.4 | 184.8 | 120.2 | 59.5 | |||
| Mountain | Ogulin | 320 m 45°15′47.84′′N 15°13′42.36′′E | a.d. T (°C) | 10.8 | 12.4 | 21.6 | 21.4 | 15.6 | 13.0 |
| T min (°C) | 0.5 | 0.5 | 11.8 | 8.3 | 2.8 | 3.0 | |||
| T max (°C) | 25.5 | 25.1 | 33.4 | 33.0 | 29.3 | 25.9 | |||
| a.p. (mm) | 167.4 | 319.2 | 139.5 | 109.4 | 143.6 | 64.2 | |||
| Čovići I | 456 m 44°49′44.07′′N 15°17′57.29′′E | a.d. T (°C) | 9.4 | 11.1 | 20.1 | 19.7 | 14.0 | 10.6 | |
| T min (°C) | −2.1 | −1.3 | 7.6 | 5.5 | −2.0 | 1.2 | |||
| T max (°C) | 24.8 | 25.0 | 34.1 | 34.5 | 29.6 | 25.1 | |||
| a.p. (mm) | 138.6 | 189.3 | 25.1 | 106.2 | 106.9 | 31.8 | |||
| Čovići II | 456 m 44°49′50.05′′N 15°17′57.18′′E | a.d. T (°C) | 9.4 | 11.1 | 20.1 | 19.7 | 14.0 | 10.6 | |
| T min (°C) | −2.1 | −1.3 | 7.6 | 5.5 | −2.0 | 1.2 | |||
| T max (°C) | 24.8 | 25.0 | 34.1 | 34.5 | 29.6 | 25.1 | |||
| a.p. (mm) | 138.6 | 189.3 | 25.1 | 106.2 | 106.9 | 31.8 | |||
| Seaside | Poreč | 0.34 m 45°13′37.03′′N 13°35′39.64′′E | a.d. T (°C) | 13.0 | 14.5 | 24.3 | 24.9 | 19.4 | 15.7 |
| T min (°C) | 3.9 | 6.0 | 13.2 | 13.4 | 7.3 | 6.3 | |||
| T max (°C) | 23.5 | 22.7 | 33.6 | 33.6 | 30.9 | 25.7 | |||
| a.p. (mm) | 116.1 | 210.0 | 7.3 | 58.7 | 143.2 | 38.6 | |||
| Limski zaljev | 17 m 45°7′56.45′′N 13°39′13.78′′E | a.d. T (°C) | 13.0 | 14.5 | 24.3 | 24.9 | 19.4 | 15.7 | |
| T min (°C) | 3.9 | 6.0 | 13.2 | 13.4 | 7.3 | 6.3 | |||
| T max (°C) | 23.5 | 22.7 | 33.6 | 33.6 | 30.9 | 25.7 | |||
| a.p. (mm) | 116.1 | 210.0 | 7.3 | 58.7 | 143.2 | 38.6 | |||
| Bale | 129 m 45°2′25.93′′N 13°47′8.88′′E | a.d. T (°C) | 13.4 | 14.4 | 23.9 | 24.5 | 19.8 | 15.5 | |
| T min (°C) | 4.9 | 4.5 | 13.8 | 13.6 | 7.5 | 5.8 | |||
| T max (°C) | 23.7 | 24.5 | 34.0 | 34.3 | 33.0 | 25.5 | |||
| a.p. (mm) | 129.5 | 264.7 | 37.4 | 71.5 | 91.1 | 42.0 | |||
| Vodnjan | 141 m 44°57′28.79′′N 13°51′6.10′′E | a.d. T (°C) | 13.4 | 14.4 | 23.9 | 24.5 | 19.8 | 15.5 | |
| T min (°C) | 4.9 | 4.5 | 13.8 | 13.6 | 7.5 | 5.8 | |||
| T max (°C) | 23.7 | 24.5 | 34.0 | 34.3 | 33.0 | 25.5 | |||
| a.p. (mm) | 129.5 | 264.7 | 37.4 | 71.5 | 91.1 | 42.0 | |||
| Muntrilj | 342 m 45°14′30.84′′N 13°48′38.44′′E | a.d. T (°C) | 11.1 | 12.5 | 22.2 | 22.3 | 16.4 | 13.1 | |
| T min (°C) | 0.5 | 1.1 | 11.2 | 9.5 | 2.3 | 2.5 | |||
| T max (°C) | 23.3 | 23.5 | 35.8 | 36.1 | 31.7 | 25.2 | |||
| a.p. (mm) | 135.1 | 295.1 | 26.0 | 72.6 | 90.5 | 26.4 | |||
| Compound | Rt (min) | Cone Voltage (V) | Collision Energy (V) | Ionization Mode | Precursor Ion (m/z) | Fragment Ions (m/z) | Tentative Identification |
|---|---|---|---|---|---|---|---|
| Benzoic acids | |||||||
| 2 | 0.828 | 105 | 9 | - | 153 | 109 | Protocatechuic acid (3,4-dihydoxybenzoic acid) |
| 14 | 0.992 | 100 | 9 | - | 153 | 109 | Gentisic acid (2,5-dihydroxybenzoic acid) |
| 31 | 8.837 | 90 | 7 | - | 197 | 182 | Syringic acid |
| 34 | 11.358 | 80 | 10 | - | 137 | 93 | p-hydroxybenzoic acid |
| 35 | 11.375 | 100 | 10 | - | 169 | 125 | Gallic acid * |
| Cinnamic acids | |||||||
| 12 | 0.975 | 80 | 10 | - | 179 | 135 | Caffeic acid * |
| 15 | 1.254 | 80 | 10 | - | 353 | 191 | Chlorogenic acid * |
| 19 | 3.332 | 80 | 10 | - | 163 | 119 | p-coumaric acid * |
| 21 | 4.490 | 100 | 5 | - | 147 | 103 | Cinnamic acid |
| 25 | 6.158 | 80 | 5 | - | 193 | 178 | Ferulic acid * |
| 32 | 11.012 | 100 | 17 | - | 223 | 193 | Sinapic acid * |
| Other phenolic acids | |||||||
| 16 | 1.620 | 150 | 20 | - | 191 | 85 | Quinic acid * |
| Flavonols | |||||||
| 3 | 0.842 | 120 | 15 | + | 625 | 317 | Isorhamnetin rutinoside |
| 4 | 0.856 | 120 | 15 | + | 595 | 287 | Kaempferol-3-rutinoside * |
| 5 | 0.880 | 100 | 5 | + | 449 | 303 | Quercetin rhamnoside |
| 6 | 0.880 | 30 | 5 | + | 449 | 287 | Kaempferol hexoside |
| 8 | 0.907 | 140 | 25 | + | 319 | 273 | Myricetin * |
| 10 | 0.938 | 130 | 15 | - | 301 | 151 | Quercetin |
| 17 | 1.855 | 100 | 5 | + | 465 | 303 | Quercetin-3-glucoside * |
| 18 | 2.461 | 120 | 5 | + | 611 | 303 | Quercetin-3-rutinoside * |
| 24 | 5.963 | 160 | 21 | - | 315 | 300 | Isorhamnetin |
| 27 | 7.106 | 100 | 5 | + | 419 | 287 | Kaempferol pentoside |
| 28 | 7.256 | 100 | 5 | + | 435 | 303 | Quercetin pentoside |
| 29 | 7.930 | 100 | 5 | + | 433 | 287 | Kaempferol rhamnoside |
| 30 | 8.242 | 100 | 10 | + | 507 | 303 | Quercetin acetylhexoside |
| 33 | 11.232 | 100 | 15 | + | 653 | 303 | Quercetin acetylrutinoside |
| 36 | 11.391 | 100 | 15 | + | 597 | 303 | Quercetin pentosylhexoside |
| 39 | 11.758 | 120 | 15 | + | 581 | 287 | Kaempferol pentosylhexoside |
| 41 | 11.822 | 130 | 0 | - | 285 | 285 | Kaempferol |
| Flavan-3-ols | |||||||
| 23 | 4.728 | 100 | 5, 15 | + | 459 | 289, 139 | Epigallocatechin gallate * |
| 37 | 11.615 | 100 | 10 | + | 291 | 139 | Epicatechin * |
| 38 | 11.621 | 100 | 5 | + | 291 | 165 | Catechin * |
| 40 | 11.792 | 100 | 5 | + | 443 | 291 | Epicatechin gallate * |
| Flavones | |||||||
| 7 | 0.890 | 135 | 5 | + | 433 | 271 | Apigenin hexoside |
| 9 | 0.924 | 140 | 35 | + | 287 | 153 | Luteolin * |
| 22 | 4.615 | 80 | 30 | + | 271 | 153 | Apigenin * |
| Isoflavones | |||||||
| 20 | 4.468 | 145 | 32 | - | 269 | 133 | Genistein |
| Flavanones | |||||||
| 11 | 0.945 | 130 | 16 | - | 271 | 151 | Naringenin * |
| Coumarins | |||||||
| 1 | 0.821 | 120 | 19 | - | 161 | 133 | Umbelliferone (7-hydroxycoumarin) |
| 13 | 0.979 | 105 | 15 | - | 177 | 133 | Esculetin * |
| 26 | 6.333 | 80 | 8 | - | 191 | 176 | Scopoletin * |
| Source of Variation | Benzoic Acids | Cinnamic Acids | Other Acids | Flavonols | Flavan-3-ols | Flavones | Isoflavones | Flavanones | Coumarins | Total Polyphenols | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Plant part | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | |
| leaves | 12.55 ± 0.04b | 209.46 ± 0.26b | 4.30 ± 0.03b | 160.26 ± 0.14b | 25.99 ± 0.04b | 29.28 ± 0.05b | 3.37 ± 0.02b | 0.40 ± 0.01b | 6.53 ± 0.01b | 452.14 ± 0.39b | |
| stalks | 7.86 ± 0.04a | 148.98 ± 0.26a | 1.45 ± 0.03a | 108.94 ± 0.14a | 15.42 ± 0.04a | 19.84 ± 0.05a | 2.81 ± 0.02a | 0.29 ± 0.01a | 4.09 ± 0.01a | 309.67 ± 0.39a | |
| Phenological stage | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | |
| 1st | 12.65 ± 0.05c | 223.32 ± 0.32c | 3.66 ± 0.03c | 169.53 ± 0.17c | 22.23 ± 0.05b | 31.89 ± 0.06c | 3.70 ± 0.02c | 0.48 ± 0.01c | 7.28 ± 0.02c | 474.75 ± 0.48c | |
| 2nd | 11.55 ± 0.05b | 202.70 ± 0.32b | 3.18 ± 0.03b | 141.72 ± 0.17b | 23.88 ± 0.05c | 22.28 ± 0.06b | 2.99 ± 0.02b | 0.33 ± 0.01b | 5.11 ± 0.02b | 413.75 ± 0.48b | |
| 3rd | 6.42 ± 0.05a | 111.63 ± 0.32a | 1.78 ± 0.03a | 92.54 ± 0.17a | 16.00 ± 0.05a | 19.50 ± 0.06a | 2.58 ± 0.02a | 0.22 ± 0.01a | 3.53 ± 0.02a | 254.21 ± 0.48a | |
| Region/Habitat | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | |
| C | Sela Žakanjska | 12.06 ± 0.10g | 200.25 ± 0.70h | 2.10 ± 0.07b | 134.87 ± 0.38g | 16.68 ± 0.11b | 24.18 ± 0.12g | 2.92 ± 0.05d | 0.31 ± 0.02bcd | 4.41 ± 0.04a | 397.78 ± 1.03g |
| Sopčić Vrh | 9.11 ± 0.10cd | 215.63 ± 0.70i | 4.12 ± 0.07f | 150.83 ± 0.38i | 26.97 ± 0.11i | 26.69 ± 0.12h | 3.25 ± 0.05e | 0.49 ± 0.02fg | 5.20 ± 0.04d | 442.29 ± 1.03j | |
| Žakanje | 19.39 ± 0.10i | 227.10 ± 0.70j | 2.56 ± 0.07c | 177.87 ± 0.38j | 32.13 ± 0.11j | 42.44 ± 0.12j | 5.29 ± 0.05i | 0.41 ± 0.02def | 5.92 ± 0.04f | 513.12 ± 1.03l | |
| Zagreb I | 10.21 ± 0.10e | 172.62 ± 0.70e | 4.22 ± 0.07fg | 130.21 ± 0.38f | 15.63 ± 0.11a | 23.33 ± 0.12f | 1.95 ± 0.05a | 0.33 ± 0.02cde | 6.76 ± 0.04h | 365.26 ± 1.03e | |
| Zagreb II | 11.06 ± 0.10f | 185.09 ± 0.70f | 4.21 ± 0.07fg | 125.81 ± 0.38d | 18.80 ± 0.11cd | 21.49 ± 0.12d | 3.47 ± 0.05ef | 0.31 ± 0.02bcd | 6.51 ± 0.04g | 376.74 ± 1.03f | |
| Koretići | 10.87 ± 0.10f | 195.03 ± 0.70g | 4.53 ± 0.07gh | 144.36 ± 0.38h | 22.09 ± 0.11g | 18.87 ± 0.12b | 3.77 ± 0.05g | 0.45 ± 0.02efg | 5.06 ± 0.04d | 405.02 ± 1.03h | |
| M | Ogulin | 13.18 ± 0.10h | 212.80 ± 0.70i | 4.62 ± 0.07h | 182.65 ± 0.38k | 20.88 ± 0.11f | 36.09 ± 0.12i | 3.51 ± 0.05f | 0.56 ± 0.02g | 6.77 ± 0.04h | 481.06 ± 1.03k |
| Čovići I | 9.44 ± 0.10cd | 203.51 ± 0.70h | 3.73 ± 0.07e | 152.62 ± 0.38i | 19.69 ± 0.11e | 23.92 ± 0.12fg | 4.47 ± 0.05h | 0.44 ± 0.02efg | 5.63 ± 0.04e | 423.46 ± 1.03i | |
| Čovići II | 8.96 ± 0.10c | 194.18 ± 0.70g | 3.08 ± 0.07d | 127.91 ± 0.38e | 21.62 ± 0.11g | 17.14 ± 0.12a | 2.91 ± 0.05d | 0.53 ± 0.02g | 4.70 ± 0.04bc | 381.01 ± 1.03f | |
| S | Poreč | 9.37 ± 0.10cd | 130.43 ± 0.70a | 1.31 ± 0.07a | 107.51 ± 0.38a | 16.43 ± 0.11b | 20.64 ± 0.12c | 1.87 ± 0.05a | 0.16 ± 0.02a | 4.40 ± 0.04a | 292.12 ± 1.03a |
| Limski zaljev | 9.53 ± 0.10d | 141.67 ± 0.70c | 1.07 ± 0.07a | 111.21 ± 0.38b | 18.32 ± 0.11c | 23.43 ± 0.12f | 2.36 ± 0.05b | 0.23 ± 0.02abc | 4.80 ± 0.04c | 312.61 ± 1.03c | |
| Bale | 7.14 ± 0.10b | 143.55 ± 0.70c | 1.82 ± 0.07b | 114.39 ± 0.38c | 19.17 ± 0.11de | 22.41 ± 0.12e | 2.68 ± 0.05cd | 0.16 ± 0.02a | 4.49 ± 0.04a | 315.81 ± 1.03cd | |
| Vodnjan | 6.14 ± 0.10a | 134.44 ± 0.70b | 1.03 ± 0.07a | 112.89 ± 0.38bc | 22.74 ± 0.11h | 22.13 ± 0.12e | 2.35 ± 0.05b | 0.19 ± 0.02ab | 5.15 ± 0.04d | 307.05 ± 1.03b | |
| Muntrilj | 6.40 ± 0.10a | 152.75 ± 0.70d | 1.87 ± 0.07b | 111.27 ± 0.38b | 18.72 ± 0.11cd | 21.06 ± 0.12cd | 2.46 ± 0.05bc | 0.22 ± 0.02abc | 4.55 ± 0.04ab | 319.29 ± 1.03d | |
| Grand mean | 10.20 | 179.22 | 2.88 | 134.60 | 20.70 | 24.56 | 3.09 | 0.34 | 5.31 | 380.90 | |
| Source of Variation | Carotenoids | Chlorophylls | Total Pigments | ORAC | |
|---|---|---|---|---|---|
| Plant Part | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | |
| leaves | 61.46 ± 0.08b | 1126.94 ± 0.66b | 1188.40 ± 0.71b | 11.96 ± 0.02b | |
| stalks | 4.60 ± 0.08a | 95.45 ± 0.66a | 100.05 ± 0.71a | 7.37 ± 0.02a | |
| Phenological Stage | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | |
| 1st | 32.49 ± 0.10b | 589.07 ± 0.81b | 621.56 ± 0.86b | 11.26 ± 0.04b | |
| 2nd | 36.97 ± 0.10c | 691.46 ± 0.81c | 728.44 ± 0.86c | 12.10 ± 0.04c | |
| 3rd | 29.64 ± 0.10a | 553.04 ± 0.81a | 582.67 ± 0.86a | 5.63 ± 0.04a | |
| Region/Habitat | p < 0.01 * | p < 0.01 * | p < 0.01 * | p < 0.01 * | |
| C | Sela Žakanjska | 37.29 ± 0.21h | 701.89 ± 1.76g | 739.18 ± 1.87h | 11.76 ± 0.06i |
| Sopčić Vrh | 31.95 ± 0.21d | 558.11 ± 1.76c | 590.06 ± 1.87d | 11.78 ± 0.06j | |
| Žakanje | 27.94 ± 0.21c | 466.84 ± 1.76a | 494.78 ± 1.87a | 12.25 ± 0.06m | |
| Zagreb I | 26.70 ± 0.21b | 480.27 ± 1.76b | 506.97 ± 1.87b | 11.89 ± 0.06k | |
| Zagreb II | 32.14 ± 0.21d | 600.73 ± 1.76d | 632.87 ± 1.87e | 9.46 ± 0.06f | |
| Koretići | 31.81 ± 0.21d | 596.83 ± 1.76d | 628.64 ± 1.87e | 11.22 ± 0.06h | |
| M | Ogulin | 33.23 ± 0.21e | 598.67 ± 1.76d | 631.91 ± 1.87e | 12.20 ± 0.06l |
| Čovići I | 25.65 ± 0.21a | 472.09 ± 1.76ab | 497.74 ± 1.87a | 10.59 ± 0.06g | |
| Čovići II | 34.29 ± 0.21f | 650.39 ± 1.76e | 684.68 ± 1.87f | 9.46 ± 0.06f | |
| S | Poreč | 27.40 ± 0.21bc | 552.88 ± 1.76c | 580.28 ± 1.87c | 6.28 ± 0.06b |
| Limski zaljev | 40.21 ± 0.21j | 719.69 ± 1.76h | 759.90 ± 1.87i | 6.26 ± 0.06a | |
| Bale | 38.35 ± 0.21i | 760.95 ± 1.76i | 799.30 ± 1.87j | 6.58 ± 0.06c | |
| Vodnjan | 35.93 ± 0.21g | 678.67 ± 1.76f | 714.60 ± 1.87g | 8.08 ± 0.06e | |
| Muntrilj | 39.55 ± 0.21j | 718.67 ± 1.76h | 758.23 ± 1.87i | 7.52 ± 0.06d | |
| Grand mean | 33.03 | 611.19 | 644.22 | 9.67 | |
| Group of Compounds | ORAC Value |
|---|---|
| Benzoic acids | 0.53 * |
| Cinnamic acids | 0.71 * |
| Other acids | 0.59 * |
| Flavonols | 0.68 * |
| Flavan-3-ols | 0.47 * |
| Flavones | 0.36 * |
| Isoflavones | 0.36 * |
| Flavanones | 0.39 * |
| Coumarins | 0.60 * |
| Total phenols | 0.71 * |
| Carotenoids | 0.46 * |
| Chlorophylls | 0.44 * |
| Total pigments | 0.44 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Repajić, M.; Cegledi, E.; Zorić, Z.; Pedisić, S.; Elez Garofulić, I.; Radman, S.; Palčić, I.; Dragović-Uzelac, V. Bioactive Compounds in Wild Nettle (Urtica dioica L.) Leaves and Stalks: Polyphenols and Pigments upon Seasonal and Habitat Variations. Foods 2021, 10, 190. https://doi.org/10.3390/foods10010190
Repajić M, Cegledi E, Zorić Z, Pedisić S, Elez Garofulić I, Radman S, Palčić I, Dragović-Uzelac V. Bioactive Compounds in Wild Nettle (Urtica dioica L.) Leaves and Stalks: Polyphenols and Pigments upon Seasonal and Habitat Variations. Foods. 2021; 10(1):190. https://doi.org/10.3390/foods10010190
Chicago/Turabian StyleRepajić, Maja, Ena Cegledi, Zoran Zorić, Sandra Pedisić, Ivona Elez Garofulić, Sanja Radman, Igor Palčić, and Verica Dragović-Uzelac. 2021. "Bioactive Compounds in Wild Nettle (Urtica dioica L.) Leaves and Stalks: Polyphenols and Pigments upon Seasonal and Habitat Variations" Foods 10, no. 1: 190. https://doi.org/10.3390/foods10010190
APA StyleRepajić, M., Cegledi, E., Zorić, Z., Pedisić, S., Elez Garofulić, I., Radman, S., Palčić, I., & Dragović-Uzelac, V. (2021). Bioactive Compounds in Wild Nettle (Urtica dioica L.) Leaves and Stalks: Polyphenols and Pigments upon Seasonal and Habitat Variations. Foods, 10(1), 190. https://doi.org/10.3390/foods10010190