Emulsion and Surface-Active Properties of Fish Solubles Based on Direct Extraction and after Hydrolysis of Atlantic Cod and Atlantic Salmon Backbones


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Chemical Analyses
2.3. Enzymatic Protein Hydrolysis
2.4. Determination of Critical Micelle Concentration by NMR Spectroscopy
2.5. Emulsion Properties
2.6. Statistics
3. Results and Discussion
3.1. Raw Material and Hydrolysate Composition
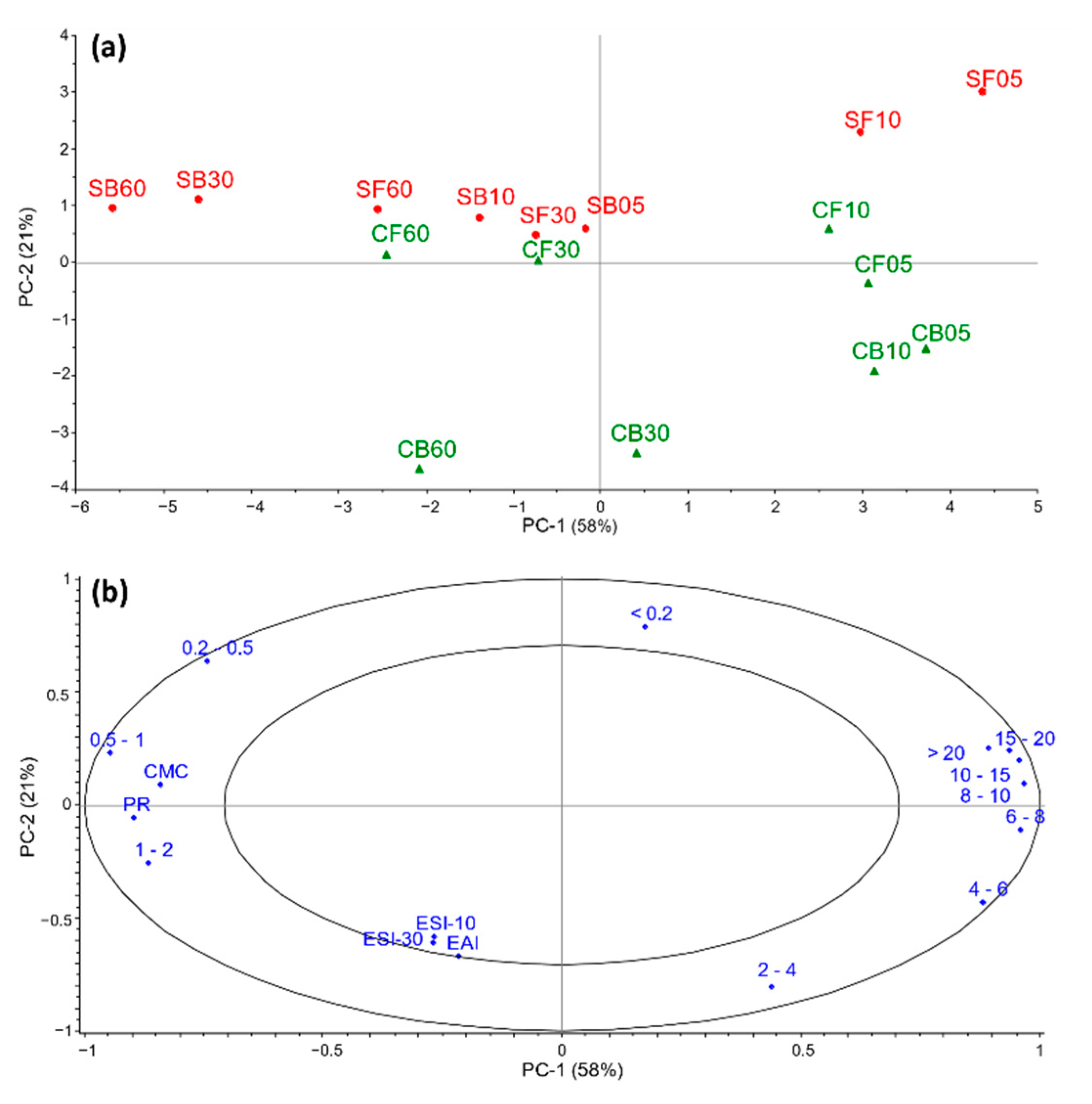
3.2. Associations between MWD and Physicochemical Properties
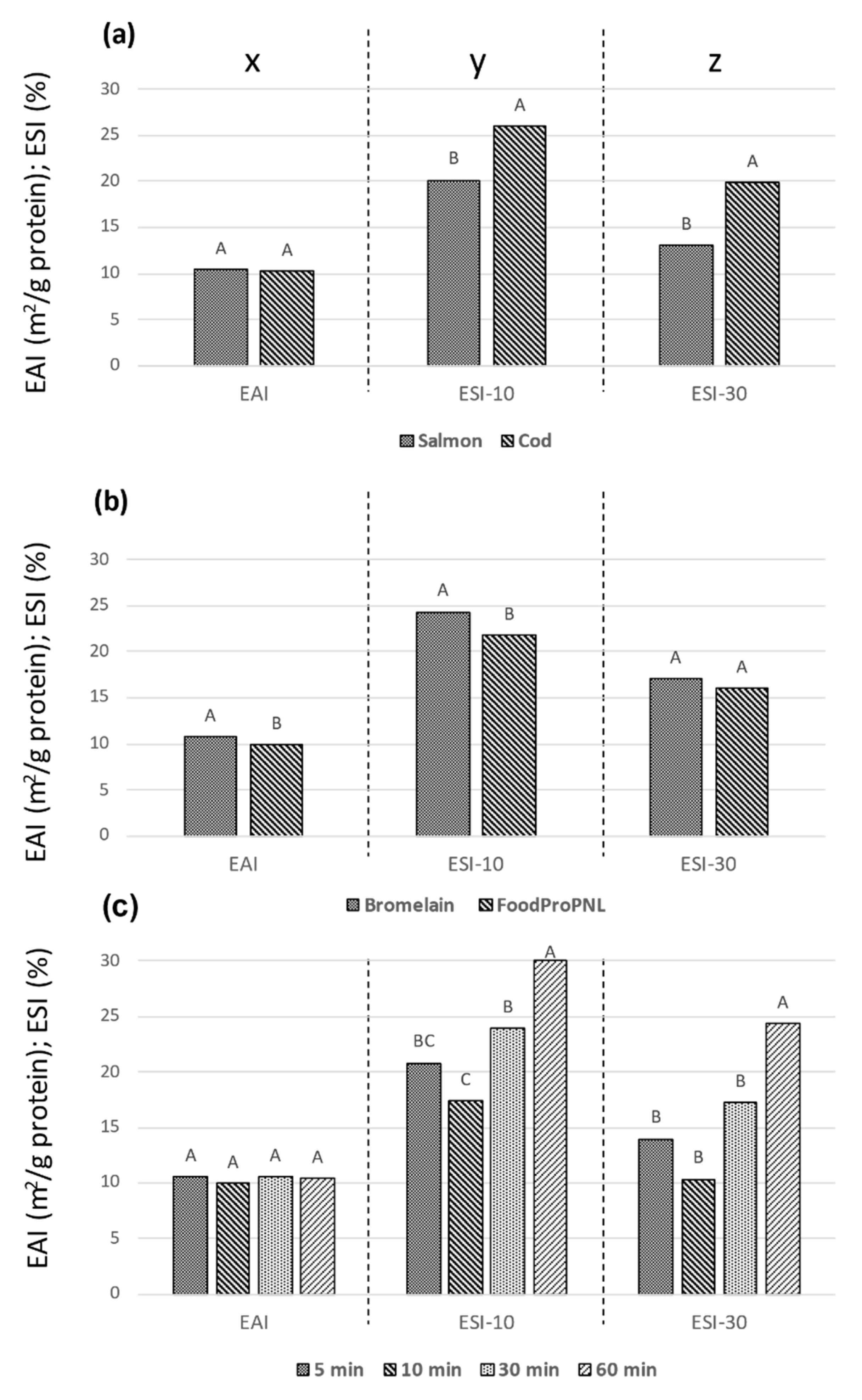
3.3. Effect of Process Parameters on Emulsion Properties
3.4. Critical Micelle Concentration of Fish Protein Hydrolysates
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McClements, D.J. Food Emulsions—Principles, Practices and Techniques, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2016. [Google Scholar]
- Ozturk, B.; McClements, D.J. Progress in natural emulsifiers for utilization in food emulsions. Curr. Opin. Food Sci. 2016, 7, 1–6. [Google Scholar]
- Berton-Carabin, C.; Schroen, K. Towards new food emulsions: Designing the interface and beyond. Curr. Opin. Food Sci. 2019, 27, 74–81. [Google Scholar]
- Kralova, I.; Sjoblom, J. Surfactants Used in Food Industry: A Review. J. Disper. Sci. Technol. 2009, 30, 1363–1383. [Google Scholar]
- Dexter, A.F.; Middelberg, A.P.J. Peptides as functional surfactants. Ind. Eng. Chem. Res. 2008, 47, 6391–6398. [Google Scholar]
- McClements, D.J. Protein-stabilized emulsions. Curr. Opin. Colloid Interface Sci. 2004, 9, 305–313. [Google Scholar]
- Panyam, D.; Kilara, A. Enhancing the functionality of food proteins by enzymatic modification. Trends Food Sci. Technol. 1996, 7, 120–125. [Google Scholar]
- Halim, N.R.A.; Yusof, H.M.; Sarbon, N.M. Functional and bioactive properties of fish protein hydolysates and peptides: A comprehensive review. Trends Food Sci. Technol. 2016, 51, 24–33. [Google Scholar]
- Da Silva Bambirra Alves, F.E.; Carpiné, D.; Teixeira, G.L.; Goedert, A.C.; de Paula Scheer, A.; Ribani, R.H. Valorization of an Abundant Slaughterhouse By-product as a Source of Highly Technofunctional and Antioxidant Protein Hydrolysates. Waste Biomass Valorization 2020. [Google Scholar] [CrossRef]
- Aspevik, T.; Totland, C.; Lea, P.; Oterhals, A. Sensory and surface-active properties of protein hydrolysates based on Atlantic salmon (Salmo salar) by-products. Process Biochem. 2016, 51, 1006–1014. [Google Scholar]
- Chobert, J.M.; Bertrandharb, C.; Nicolas, M.G. Solubility and Emulsifying Properties of Caseins and Whey Proteins Modified Enzymatically by Trypsin. J. Agric. Food Chem. 1988, 36, 883–892. [Google Scholar]
- Elavarasan, K.; Kumar, V.N.; Shamasundar, B.A. Antioxidant and Functional Properties of Fish Protein Hydrolysates from Fresh Water Carp (Catla catla) as Influenced by the Nature of Enzyme. J. Food Process. Pres. 2014, 38, 1207–1214. [Google Scholar]
- Gbogouri, G.A.; Linder, M.; Fanni, J.; Parmentier, M. Influence of Hydrolysis Degree on the Functional Properties of Salmon Byproducts Hydrolysates. J. Food Sci. 2004, 69, C615–C622. [Google Scholar]
- Klompong, V.; Benjakul, S.; Kantachote, D.; Shahidi, F. Antioxidative activity and functional properties of protein hydrolysate of yellow stripe trevally (Selaroides leptolepis) as influenced by the degree of hydrolysis and enzyme type. Food Chem. 2007, 102, 1317–1327. [Google Scholar]
- Kristinsson, H.G.; Rasco, B.A. Biochemical and functional properties of Atlantic salmon (Salmo salar) muscle proteins hydrolyzed with various alkaline proteases. J. Agric. Food Chem. 2000, 48, 657–666. [Google Scholar] [PubMed]
- Lee, S.W.; Shimizu, M.; Kaminogawa, S.; Yamauchi, K. Emulsifying Properties of Peptides Obtained from the Hydrolyzates of Beta-Casein. Agric. Biol. Chem. 1987, 51, 161–166. [Google Scholar]
- Nalinanon, S.; Benjakul, S.; Kishimura, H.; Shahidi, F. Functionalities and antioxidant properties of protein hydrolysates from the muscle of ornate threadfin bream treated with pepsin from skipjack tuna. Food Chem. 2011, 124, 1354–1362. [Google Scholar]
- Pacheco-Aguilar, R.; Mazorra-Manzano, M.A.; Ramirez-Suarez, J.C. Functional properties of fish protein hydrolysates from Pacific whiting (Merluccius productus) muscle produced by a commercial protease. Food Chem. 2008, 109, 782–789. [Google Scholar]
- Quaglia, G.B.; Orban, E. Influence of Enzymatic-Hydrolysis on Structure and Emulsifying Properties of Sardine (Sardina-Pilchardus) Protein Hydrolysates. J. Food Sci. 1990, 55, 1571–1573. [Google Scholar]
- Schroder, A.; Berton-Carabin, C.; Venema, P.; Cornacchia, L. Interfacial properties of whey protein and whey protein hydrolysates and their influence on O/W emulsion stability. Food Hydrocoll. 2017, 73, 129–140. [Google Scholar]
- Tan, Y.; Chang, S.K.C.; Meng, S. Comparing the kinetics of the hydrolysis of by-product from channel catfish (Ictalurus punctatus) fillet processing by eight proteases. LWT 2019, 111, 809–820. [Google Scholar]
- Aspevik, T.; Oterhals, Å.; Ronning, S.B.; Altintzoglou, T.; Wubshet, S.G.; Gildberg, A.; Afseth, N.K.; Whitaker, R.D.; Lindberg, D. Valorization of Proteins from Co- and By-Products from the Fish and Meat Industry. Topics Curr. Chem. 2017, 375. [Google Scholar] [CrossRef] [Green Version]
- Liceaga-Gesualdo, A.M.; Li-Chan, E.C.Y. Functional properties of fish protein hydrolysate from herring (Clupea harengus). J. Food Sci. 1999, 64, 1000–1004. [Google Scholar] [CrossRef]
- Van der Ven, C.; Gruppen, H.; de Bont, D.B.; Voragen, A.G. Emulsion properties of casein and whey protein hydrolysates and the relation with other hydrolysate characteristics. J. Agric. Food Chem. 2001, 49, 5005–5012. [Google Scholar] [CrossRef] [PubMed]
- Pearce, K.N.; Kinsella, J.E. Emulsifying properties of proteins—Evaluation of a turbidimetric technique. J. Agric. Food Chem. 1978, 26, 716–723. [Google Scholar] [CrossRef]
- Al-Soufi, W.; Piñeiro, L.; Novo, M. A model for monomer and micellar concentration in surfactant solutions: Application to conductivity, NMR, diffusion, and surface tension data. J. Colloid Interface Sci. 2012, 370, 102–110. [Google Scholar] [CrossRef]
- Liu, X.; Yu, Q.; Song, A.; Dong, S.; Hao, J. Progress in nuclear magnetic resonance studies of surfactant systems. Curr. Opin. Colloid Interface Sci. 2020, 45, 14–27. [Google Scholar] [CrossRef]
- ISO 5983-2:2009. Animal Feeding Stuffs—Determination of Nitrogen Content and Calculation of Crude Protein Content; Part 2: Block Digestion and Steam Distillation Method; International Organization for Standardization (ISO): Geneva, Switzerland, 2009. [Google Scholar]
- Sriperm, N.; Pesti, G.M.; Tillman, P.B. Evaluation of the fixed nitrogen-to-protein (N:P) conversion factor (6.25) versus ingredient specific N:P conversion factors in feedstuffs. J. Sci. Food Agric. 2011, 91, 1182–1186. [Google Scholar] [CrossRef]
- Cohen, S.A.; Michaud, D.P. Synthesis of a fluorescent derivatization reagent, 6-aminoquinolyl-N-hydroxysuccinimidyl carbamate, and its application for the analysis of hydrolysate amino-acids via high-performance liquid-chromatography. Anal. Biochem. 1993, 211, 279–287. [Google Scholar] [CrossRef]
- Aspevik, T.; Egede-Nissen, H.; Oterhals, Å. A Systematic Approach to the Comparison of Cost Efficiency of Endopeptidases for the Hydrolysis of Atlantic Salmon (Salmo salar) By-Products. Food Technol. Biotech. 2016, 54, 421–431. [Google Scholar] [CrossRef]
- Conway, E.J.; Byrne, A. An absorption apparatus for the micro-determination of certain volatile substances: The micro-determination of ammonia. Biochem. J. 1933, 27, 419–429. [Google Scholar]
- Wang-Andersen, J.; Haugsgjerd, B.O. Improved Analysis for Peptide Size Distribution in Marine Hydrolysates (In Norwegian); RUBIN: Trondheim, Norway, 2011; ISBN 978-82-7251-926-0. [Google Scholar]
- Oterhals, A.; Samuelsen, T.A. Plasticization effect of solubles in fishmeal. Food Res. Int. 2015, 69, 313–321. [Google Scholar] [CrossRef]
- Steinsholm, S.; Oterhals, Å.; Underhaug, J.; Måge, I.; Malmendal, A.; Aspevik, T. Sensory Assessment of Fish and Chicken Protein Hydrolysates. Evaluation of NMR Metabolomics Profiling as a New Prediction Tool. J. Agric. Food Chem. 2020, 68, 3881–3890. [Google Scholar] [CrossRef] [PubMed]
- BRENDA. The Comprehensive Enzyme Information System. Available online: http://www.brenda-enzymes.org/ (accessed on 16 September 2019).
- Kousoulaki, K.; Olsen, H.J.; Albrektsen, S.; Langmyhr, E.; Mjos, S.A.; Campbell, P.; Aksnes, A. High growth rates in Atlantic salmon (Salmo salar L.) fed 7.5% fish meal in the diet. Micro-, ultra- and nano-filtration of stickwater and effects of different fractions and compounds on pellet quality and fish performance. Aquaculture 2012, 338, 134–146. [Google Scholar] [CrossRef]
- Wouters, A.G.B.; Rombouts, I.; Fierens, E.; Brijs, K.; Delcour, J.A. Relevance of the Functional Properties of Enzymatic Plant Protein Hydrolysates in Food Systems. Compr. Rev. Food Sci. Food Saf. 2016, 15, 786–800. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| AA (g/kg) ** | Cod Backbone | Salmon Backbone |
|---|---|---|
| Alanine | 11.0 | 9.9 |
| Arginine | 11.0 | 9.0 |
| Asparagine * | 7.1 | 6.3 |
| Aspartate | 8.9 | 7.7 |
| Glutamate | 16.1 | 12.0 |
| Glutamine * | 7.9 | 7.0 |
| Glycine | 17.0 | 14.0 |
| Histidine | 3.3 | 3.6 |
| Isoleucine | 6.4 | 5.7 |
| Leucine | 12.0 | 9.6 |
| Lysine | 13.0 | 11.0 |
| Methionine | 5.4 | 4.6 |
| Phenylalanine | 5.9 | 5.2 |
| Proline | 8.8 | 7.8 |
| Serine | 8.6 | 6.5 |
| Threonine | 6.9 | 6.2 |
| Tyrosine | 6.9 | 4.0 |
| Valine | 7.4 | 6.7 |
| Hydroxyproline | 3.8 | 3.5 |
| Nitrogen | 27 | 25 |
| NH3 (acid digest) | 1.8 | 1.6 |
| Total AA | 167.4 | 140.3 |
| Total n | 27.0 | 25.0 |
| fN *** | 5.5 | 5.2 |
| MWD (%) | C-ext | CF05 | CF10 | CF30 | CF60 | CB05 | CB10 | CB30 | CB60 | S-ext | SF05 | SF10 | SF30 | SF60 | SB05 | SB10 | SB30 | SB60 | AA * |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| >20 | 8.9 | 1.3 | 1.2 | 0.4 | 0.2 | 1.3 | 1.4 | 0.8 | 0.3 | 4.9 | 2.6 | 1.5 | 0.4 | 0.1 | 1.1 | 0.8 | 0.1 | <0.1 | >198 |
| 15–20 | 3.9 | 1.1 | 0.9 | 0.3 | 0.1 | 1.0 | 0.9 | 0.5 | 0.2 | 2.2 | 1.7 | 1.1 | 0.4 | 0.1 | 0.6 | 0.4 | 0.1 | <0.1 | 138 |
| 10–15 | 5.7 | 2.9 | 2.7 | 1.1 | 0.5 | 3.0 | 2.6 | 1.3 | 0.5 | 3.1 | 4.1 | 3.1 | 1.3 | 0.4 | 1.2 | 0.8 | 0.2 | <0.1 | 99 |
| 8–10 | 3.4 | 3.0 | 2.9 | 1.5 | 0.8 | 3.5 | 3.2 | 1.8 | 0.8 | 1.7 | 4.0 | 3.4 | 1.9 | 0.9 | 1.0 | 0.7 | 0.2 | 0.1 | 71 |
| 6–8 | 4.0 | 6.0 | 6.1 | 3.9 | 2.5 | 7.8 | 7.4 | 5.1 | 2.8 | 2.2 | 7.0 | 6.5 | 4.5 | 2.7 | 2.5 | 2.0 | 0.6 | 0.3 | 55 |
| 4–6 | 3.8 | 12.0 | 12.6 | 9.7 | 7.2 | 15.9 | 16.0 | 13.7 | 10.0 | 2.4 | 11.6 | 11.2 | 8.9 | 6.7 | 7.6 | 6.6 | 2.9 | 1.7 | 32 |
| 2–4 | 3.7 | 21.4 | 23.0 | 22.8 | 21.2 | 26.4 | 27.9 | 30.0 | 29.3 | 2.5 | 18.6 | 20.0 | 20.0 | 18.2 | 23.3 | 23.0 | 16.0 | 12.0 | 20 |
| 1–2 | 1.9 | 16.1 | 17.2 | 20.2 | 21.4 | 14.6 | 16.2 | 20.7 | 25.0 | 1.3 | 12.3 | 14.0 | 16.9 | 18.4 | 22.2 | 23.7 | 25.7 | 24.5 | 12 |
| 0.5–1 | 1.2 | 10.0 | 11.0 | 14.9 | 17.5 | 6.2 | 6.9 | 9.7 | 13.3 | 0.8 | 7.7 | 9.2 | 13.0 | 16.0 | 12.8 | 14.3 | 21.1 | 24.2 | 5.9 |
| 0.2–0.5 | 3.3 | 6.6 | 7.1 | 10.3 | 13.0 | 3.3 | 3.4 | 4.4 | 6.0 | 18.4 | 10.3 | 11.0 | 13.6 | 16.4 | 10.7 | 11.5 | 16.5 | 20.0 | 2.8 |
| <0.2 | 60.3 | 19.7 | 15.4 | 14.8 | 15.7 | 17.0 | 14.1 | 12.1 | 11.9 | 60.7 | 20.2 | 19.1 | 19.1 | 20.1 | 16.9 | 16.4 | 16.4 | 17.2 | 1.0 |
| EAI (m2/g) | ESI-10 | ESI-30 | CMC (g/l) | PR (%) | |
|---|---|---|---|---|---|
| C-ext | 13 ± 0.9 a | 21 ± 1.9 cde | 16 ± 3.0 de | 1.6 | 6.2 |
| CF05 | 11 ± 1.2 bcd | 33 ± 2.7 a | 25 ± 0.6 ab | 3.7 | 18.1 |
| CF10 | 9.0 ± 0.2 cd | 18 ± 1.9 def | 9 ± 0.6 efg | 4.8 | 23.4 |
| CF30 | 11 ± 0.7 bcd | 14 ± 2.1 ef | 7 ± 1.3 fgh | 6.0 | 31.4 |
| CF60 | 9.0 ± 0.2 d | 27 ± 0.5 abcd | 22 ± 2.9 abc | 6.2 | 35.6 |
| CB05 | 11 ± 0.5 bcd | 31 ± 1.7 abc | 22 ± 2.9 bcd | 5.1 | 24.0 |
| CB10 | 11 ±0.7 bcd | 25 ± 1.6 abc | 17 ± 1.5 cde | 5.4 | 27.0 |
| CB30 | 11 ± 0.4 bcd | 33 ± 1.4 abc | 26 ± 3.1 ab | 6.0 | 31.5 |
| CB60 | 11 ± 0.1 abc | 31 ± 2.2 a | 27 ± 1.6 a | 6.6 | 36.6 |
| S-ext | 12 ± 1.2 ab | 10 ± 1.0 f | 6 ± 1.8 gh | 1.8 | 7.0 |
| SF05 | 10 ± 1.2 bcd | 11 ± 2.4 f | 6 ± 1.0 gh | 5.8 | 26.5 |
| SF10 | 10 ± 0.3 bcd | 12 ± 0.0 f | 6 ± 1.2 gh | 5.3 | 25.8 |
| SF30 | 10 ± 0.4 bcd | 30 ± 2.2 ab | 23 ± 1.6 abc | 6.7 | 31.1 |
| SF60 | 10 ± 0.3 bcd | 32 ± 3.6 a | 25 ± 3.7 ab | 6.8 | 33.7 |
| SB05 | 11 ± 0.3 bcd | 11 ± 3.5 f | 4 ± 0.7 h | 5.3 | 26.1 |
| SB10 | 10 ± 0.5 bcd | 13 ± 0.6 ef | 5 ± 1.0 h | 6.2 | 30.0 |
| SB30 | 10 ± 0.5 bcd | 22 ± 1.7 bcd | 13 ± 0.4 ef | 7.6 | 40.0 |
| SB60 | 11 ± 0.5 bcd | 31 ± 3.3 ab | 22 ± 1.4 abc | 7.6 | 40.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steinsholm, S.; Oterhals, Å.; Underhaug, J.; Aspevik, T. Emulsion and Surface-Active Properties of Fish Solubles Based on Direct Extraction and after Hydrolysis of Atlantic Cod and Atlantic Salmon Backbones. Foods 2021, 10, 38. https://doi.org/10.3390/foods10010038
Steinsholm S, Oterhals Å, Underhaug J, Aspevik T. Emulsion and Surface-Active Properties of Fish Solubles Based on Direct Extraction and after Hydrolysis of Atlantic Cod and Atlantic Salmon Backbones. Foods. 2021; 10(1):38. https://doi.org/10.3390/foods10010038
Chicago/Turabian StyleSteinsholm, Silje, Åge Oterhals, Jarl Underhaug, and Tone Aspevik. 2021. "Emulsion and Surface-Active Properties of Fish Solubles Based on Direct Extraction and after Hydrolysis of Atlantic Cod and Atlantic Salmon Backbones" Foods 10, no. 1: 38. https://doi.org/10.3390/foods10010038
APA StyleSteinsholm, S., Oterhals, Å., Underhaug, J., & Aspevik, T. (2021). Emulsion and Surface-Active Properties of Fish Solubles Based on Direct Extraction and after Hydrolysis of Atlantic Cod and Atlantic Salmon Backbones. Foods, 10(1), 38. https://doi.org/10.3390/foods10010038