1. Introduction
2-Arylbenzofurans are uncommon phenolic compounds that have been found only in a limited number of plant families, such as Corsiniaceae, Gnetaceae, Melanthiaceae, Stemonaceae, Moraceae, Fabaceae, and Vitaceae [
1]. They are present in significant amounts in all parts of the mulberry tree (
Morus spp.).
2-Arylbenzofurans have a structure related to stilbenoids and exhibit bioactive effects with potential applications in medicine and human nutrition [
2]. The backbone of their skeleton is based on a benzofuran ring substituted with a phenyl group. Derivatives of benzofurans show potent biological activities including an antimicrobial effect [
3], with some showing similar potency to commercial antibiotics [
4]. They also exhibit antiparasitic [
5], analgesic [
6], antitumor, and kinase inhibitor activity [
7]. Mulberrofuran G (syn. albanol A) showed potent antimicrobial effects [
8] and acts as a COX and LOX inhibitor [
9], which means that it can modulate the inflammatory response. Mulberrofuran Y (
1) and moracin C (
2) demonstrated cytotoxic activity using the THP-1 human monocytic leukemic cell line of 4.8 ± 0.19 and 3.2 ± 0.13 µM, respectively [
10]. These compounds also significantly inhibited the production of nitric oxide in RAW 264.7 macrophage cells [
11]. Other biological activities of moracin C (
2) and mulberrofuran G (
3), such as hypotensive, antitumor preventive, and others, have been summarized in review articles [
12,
13]. These results suggest a potential benefit of 2-arylbenzofurans dietary intake, especially for chronical inflammatory diseases and tumor prevention [
9,
13].
In plants, phenolic compounds are commonly present as part of higher molecular structures. They form polymeric structures with other phenolics, which are bound to other organic substances including dietary fibers, proteins, saccharides, and organic acids. This could lead to a relatively low absorption rate of plant phenolic compounds in the human small intestine, with approximately 5–10% of all of them being digested in the proximal part of the gastrointestinal tract, and the remainder passing to the colon where they can be fermented by bacteria [
14,
15]. Once in the colon, polyphenols are metabolized by the gastrointestinal microbiota to compounds of lower molecular weight, such as phenolic acids, which can be readily absorbed across the intestinal barrier into the bloodstream. In general, for phenolic compounds, there are three well-documented major pathways for most plant metabolites: hydrolysis, cleavage, and reduction [
16].
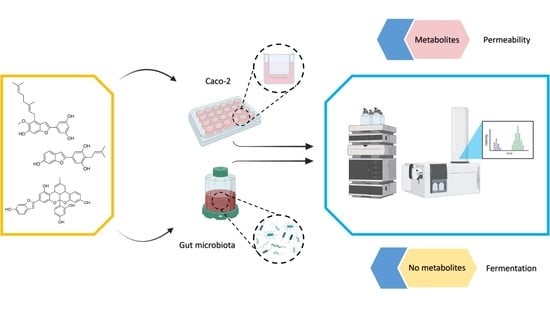
Although information on the metabolism of a number of important polyphenolic compounds by the human gut microbiota is becoming increasingly available, much is still unknown for 2-arylbenzofurans. For this reason, we attempted to investigate the fate of selected 2-arylbenzofurans in the gut based on the hypothesis that these compounds are degraded in the colon by the gut microbiota in a manner similar to that of other structurally or biochemically related phenolic compounds. Because the bioactive compounds including 2-arylbenzofurans can be transformed into metabolites with changed activity or may be retained inside epithelial cells during their transport across the intestinal barrier, it is important to investigate their fate in an in vitro permeability model. This may also indicate the potential site of action of ingested 2-arylbenzofurans. To this end, we used two methods: an in vitro fermentation model to screen for degradation of mulberrofuran Y (1), moracin C (2), and mulberrofuran G (3), and a permeability assay on the Caco-2 cell line in TranswellTM plates to screen for the intestinal metabolism of these compounds.
2. Materials and Methods
2.1. Study Design
Using a batch fermentation model of the human colon, 2-arylbenzofurans were incubated after the addition of human fecal bacteria from three donors. Samples were collected at 0, 2, 4, and 8 h and the effect of microbiota on tested 2-arylbenzofurans was determined by UHPLC/MS HRAM.
2.2. Ethics Statement and Donor Information
All three volunteers (two males and one female aged 23, 28, and 29 years) were asked to donate stool samples free of charge. The ethical agreement for stool collection was obtained by the ethical committee (ZEK/22/09/2017) of the Czech University of Life Sciences in Prague. Body mass index was estimated to be between 23 and 26.5. Because we wanted to limit dietary interference, the donors were forced to follow a low-polyphenol diet for at least 2 days before starting the experiment. No antibiotics were allowed for at least 3 months before the sample collection. They had no history of chronic gastrointestinal disease. The woman was neither breastfeeding nor pregnant. Samples were collected from November to December and analyzed at the Czech University of Life Sciences Prague.
2.3. Reagents
The fermentation medium was prepared from chemicals obtained from Merck (Darmstadt, Germany). Standards: 2-arylbenzofurans mulberrofuran Y (
1), moracin C (
2), and mulberrofuran G (
3) were obtained from ChemFaces (WuHan, China) in 98% purity (
Figure 1). The organic solution, ethyl acetate, and methanol were purchased from the VWR International (Stribrna Skalice, Czech Republic) in the analytical grade. Dimethylsulphoxide was obtained from VWR Chemicals. Formic acid was obtained from Fisher Scientific (Merelbeke, Belgium) in >98% purity. Ultra-pure water was obtained from a Millipore system (Bedford, MA, USA).
2.4. Fermentation Medium
Medium was prepared from 225 mL of distilled water, 112.5 mL of macromineral solution, 56.25 μL of micromineral solution, 562.5 μL of 0.1% resazurin solution, and 1.125 g of tryptone. The macromineral solution was prepared from 7.14 g of Na2HPO4, 6.2 g of KH2PO4, 0.6 g of MgSO4, and up to 1 L of distilled water. The micromineral solution was made of 2.64 g of CaCl2, 2 g of MnCl2, CoCl2, 1.6 g of FeCl3, and up to 20 mL of distilled water. CO3 buffer was created by mixing 1 g of NH4HCO3, 8.75 g of NaHCO3, and distilled water up to 250 mL.
2.5. Phosphate Buffer and Reducing Solution
Phosphate buffer for fecal slurries was created by dissolving 1.77 g of KH2PO4 in 195 mL of distilled water and 3.62 g of Na2HPO4 in 305 mL of distilled water (both 1/15M). The acidity of the mixed phosphate buffer was adjusted to a neutral pH by hydrochloric acid. The reducing solution was created by mixing 125 mg of cysteine hydrochloride, 125 mg of Na2S, 0.8 mL of 1M NaOH, and distilled water up to 20 mL.
2.6. Fermentation with Human Fecal Microbiota
Mulberrofuran Y (1), moracin C (2), and mulberrofuran G (3) were dissolved in DMSO to a final concentration of 10 mg/mL. After boiling the fermentation medium and sodium phosphate buffer, both were cooled to 37 °C. To remove oxygen, the samples were purged with nitrogen gas (approximately 30 min). The pH of the medium was equilibrated to neutral pH using HCl. For each fermentation bottle, 17.18 mL of fermentation medium and 0.8 mL of reducing solution were transferred to create fermentation vials. Fecal slurry was prepared immediately after sampling from each donor by homogenizing the collected feces in a stomacher bag with phosphate buffer to produce a 25% fecal slurry. After filtration through the mesh, 2 mL of the filtered fecal slurry were added to each fermentation bottle. Finally, 20 μL of the tested compound or DMSO as a negative control were added. The bottles were incubated at 37 °C for 8 h with stirring at 100 strokes per minute. Aliquots of fecal suspensions were collected at 0, 2, 4, and 8 h, and stored at −80 °C until further analysis.
2.7. Samples Purification
First, 400 μL sample of supernatant fluid were defrosted and centrifuged (5 min/15,000 RPM) and the pellet was erased. Supernatant was combined with 20 μL of 13C6 trans-resveratrol in methanol (2 μg/mL), which was used as an internal standard. Samples were extracted 3 times with 2.5 mL of ethyl acetate and 2 mL of ultra-pure water. After liquid-liquid extraction, ethyl acetate was evaporated with nitrogen gas and redissolved in 1 mL of methanol (VWR Chemicals, Stribrna Skalice, Czech Republic) in combination with formic acid (Fisher Scientific, Merelbeke, Belgium) (1%).
2.8. Metabolite Analysis
The analysis was performed on an LC/MS system consisting of an Ultimate 3000 UPLC chromatograph Thermo Fisher Scientific (Waltham, MA, USA) coupled to an Impact II ultra-high resolution high mass accuracy mass spectrometer (HRAM) Q-TOF (Bruker Daltonics, Bremen, Germany) equipped with an electrospray ionization (ESI) source.
The separation was carried out on a Phenomenex F5 Kinetex column (1.7 μm 100 Å 100 × 2.1 mm) (Phenomenex, Torrance, CA, USA) by using a mobile phase consisting of 0.1% formic acid (solvent A) and methanol (solvent B). The binary gradient was run at a flow rate of 0.2 mL/min as follows: 0–3 min isocratic at 20% B, 3–6 min from 20% to 50% B, 6–15 min isocratic at 100% B, and 15–20 min isocratic at 20% B. The column oven was adjusted to 35 °C, and the injection volume was 5 μL.
For metabolite screening, the ESI source was set in both positive and negative mode. Semiquantitative analysis was performed in negative mode with the parameters listed in the
Appendix A. Each compound was confirmed by MS/MS fragmentation with a collision energy voltage (20, 30, and 50 eV). Data acquisition was performed using HyStar 3.2 SR4, Otof series 4.0 (Bruker Daltonics, Bremen, Germany) and Chromeleon Xpress (Thermo Fisher Scientific, Waltham, MA, USA) software and the data obtained were analyzed by using DataAnalysis 4.3. (Bruker Daltonics, Bremen, Germany). Commercially available standards of mulberrofuran Y (
1), moracin C (
2), and mulberrofuran G (
3), each at 9 concentration levels ranging from 0.5–500 ng/mL, were used for calibration. A list of the 2-arylbenzofurans monitored in the samples is given in the
Appendix B.
2.9. Permeability Experiment
The Caco-2 cell lines were purchased from the American Type Tissue Collection (Rockville, MD, USA). Caco-2 cells in Transwells at passage 25 were used. More information on cell culture and passaging is described in [
17].
2.10. Preparation of Inserts
Cells were cultured in DMEM-F12 (Dulbecco’s modified Eagles medium—Nutrient Mixture F-12) supplemented with 10% FBS (fetal bovine serum), 1% nonessential amino acids, 1% penicillin, and streptomycin, all obtained from Sigma-Aldrich (Prague, Czech Republic) in humidified air containing 5% CO2. Cells were seeded at a density of 2.6 × 105 cells/cm in 24-wells. The inserts were pre-filled with 50 µL of medium before seeding the cells. After seeding, the basolateral side of the chamber was filled with 1 mL of DMEM, and cells were grown in a humidified atmosphere of 5% CO2 at 37 °C. Non-adherent cells were removed by removing the medium after 6 h of incubation and replaced with 0.5 mL of fresh DMEM. The culture medium was replaced seven times a week for 21 to 25 days prior to the transport experiment.
2.11. Measuring of the Monolayer Integrity
The filter inserts containing a monolayer of the Caco-2 cell line were washed three times with HBBS (Hanks’ Balanced Salt solution) heated to 37 °C and pH 7.4. The integrity of the cell monolayer was confirmed by measuring the transepithelial electrical resistance (TEER), which had to be at least 600 Ω. After adding 25 µM lucifer yellow, the plates were incubated at 37 °C in 5% CO2 atmosphere for 1 h with shaking (150 rpm). Using a Tecan Infinite M200 reader (Excitation/Emission wavelength 480 nm/530 nm), the plates were measured. Only plates with an integrity greater than 95% were used in the following experiment.
2.12. Metabolism and Absorption of 2-Arylbenzofurans
Initially, 500 µL of 20 µM test substance solutions were added to the apical side of each insert and 1000 µL of HBSS were added to the basolateral side. At the same time, 50 µL were taken immediately from the apical side at the time point of 0 h. For future analysis, basolateral sampling times of 0.5, 1, 1.5, 2, 3, and 4 h were chosen while placing the plates in the incubator and using an orbital shaker (150 rpm). In this experiment, 500 µL of sample were always taken from the basolateral side and replaced with an equal amount of pure HBSS. After 4 h, the apical side was simultaneously removed, and the inserts were washed three times with HBSS. The TEER method was again used to verify that the experiment did not compromise cellular integrity. Finally, cell integrity was disrupted by adding 100% methanol and the cell contents with the other samples were stored at −80 °C until the analysis.
2.13. Statistical Evaluation
Due to the design of the experiment and the decreasing concentration of substances on the basolateral side, it was necessary to recalculate each time point by the following equation:
where
CA is the actual concentration at the time point,
CP are the previous concentrations, and
CM is the concentration measured at the time point. Three biological repetitions were performed for all compounds. Samples were measured by LC/MS in triplicates. Results are presented as a mean ± standard error. Quantitative data were normalized to 20 µM to correct the minor dilution errors. Graph creation and basic statistical analysis were carried out via Microsoft Excel and SPSS version 25 (IBM Corp., Armonk, NY, USA).
4. Discussion
In the literature, substances like 2-arylbenzofurans are reported to have promising bioactive effects with potential for applications in medicine and human nutrition. This study attempted to add to the unknown information on the biotransformation of mulberrofuran Y (1), moracin C (2), and mulberrofuran G (3) by human microbiota and cellular uptake through an intestinal in vitro model. Bacteria did not produce any metabolites of the three compounds investigated. Moreover, the results clearly indicate a difference in metabolite sulfation and glucuronidation processes when investigating the permeability by intestinal cells.
To find out how and whether these substances can be metabolized by bacterial communities in the human gut, the tested 2-arylbenzofurans were fermented by human fecal microbiota. No metabolites were found. It has been reported in the literature that benzofuran can be cleaved at the oxygen bond in its heterocycle [
18,
19]. Oberoi et al. described the aerobic bacterial decomposition of benzofuran to carbon dioxide and water, with either catechol or salicylic acid as possible intermediates [
18]. These products have also been observed after metabolism in flavonol compounds [
20]. Similar results were described for heterocyclic O-bound cleavage of dibenzofuran when 2-hydroxybenzoic and 2,5-dihydroxybenzoic acid were formed as the major products on the O-heterocycle [
19]. In this study, no common microbiota degradation products, such as 4-hydroxybenzoic acid, 3,5-dihydroxybenzoic acid, protocatechuic acid, resorcinol, or phloroglucinol, were found. This suggests that the metabolism of these compounds occurs in an unusual way or not at all. There is an assumption that the decreasing concentrations are due to the intestinal microbiota, so differences among donors could be caused by differences in the microbiota composition [
21]. In our study, 2-arylbenzofurans were gradually catabolized, and their final concentration, after 8 h of fermentation, ranged from 87 ± 10% (D2) to 66 ± 3% (D1) for moracin C (
2), 69 ± 8% (D2) to 25 ± 0% (D1) for mulberrofuran Y (
1), and 102 ± 13% (D2) to 48 ± 0% (D1). These results clearly indicate that microbiota composition is an important aspect affecting the stability of 2-arylbenzofurans.
In contrast to our findings in bacterial exposure, human colonocytes have the ability to transform 2-arylbenzofurans. Metabolism by enterocytes is an important step affecting their bioactivity. We observed major differences among each of these compounds.
The main amount of mulberrofuran Y (
1) (85.3 ± 19.9%) was detected to be unchanged in the cells after 4 h of incubation. This indicates that the intestinal wall is the main target site of this compound. After 4 h of incubation, 8.9 ± 5.9% of unchanged mulberrofuran Y (
1) was found in the medium in the apical chamber. Interestingly, no amount of this compound was found on the basolateral side. However, its glucuronic conjugate was detected predominantly inside the cells and on the apical side, and only a small amount of this metabolite was detected on the basolateral side. Intestinal cells have an enzymatic system that allows them to produce two types of metabolites. Metabolites formed by conjugation with glucuronic acid or with a sulfonic acid functional group are produced by the action of the enzymes UDP-glucuronosyltransferases (UGTs) and sulfotransferases (SULTs) [
22,
23]. UGTs are the primary phase II enzymes catalyzing the conjugation of glucuronic acid to xenobiotics with polar groups to facilitate their clearance [
23]. Our data suggest that UGTs are probably the main enzyme system included in the metabolism of mulberrofuran Y. In addition, the metabolites probably leave the cells via ABC transporters, such as multidrug resistance protein (MRP2-apical side, MRP3-basolateral side) and breast cancer resistance protein (BRCP), similar to other phenolics [
24,
25].
The metabolism of mulberrofuran G (3) differed significantly from mulberrofuran Y (1). Only 14.0 ± 0.6% of the parental compound was found in the cells, and 2.7 ± 0.5% on the apical side at the end of the experiment. This means that the majority of compound (83.3 ± 1.1%) was changed and nothing of the original molecule was able to reach the basolateral side. Only a small volume of glucuronic metabolite was detectable in the cells after 4 h of incubation. These results point to a significant lowering of the original molecule’s bioavailability. However, since we did not find all the metabolites, we cannot decipher the main site of action for these substances.
After application of moracin C (
2) on Caco-2 cells, the vast majority, 91.1 ± 0.7%, were transformed. Interestingly, two types of glucuronic metabolites were detected in the cells, on the apical and basolateral sides. While in the case of mulberrofuran Y (
1), we were able to find the metabolite mostly in the cellular region, in the case of moracin C (
2), both metabolites favored the apical side, and then the basolateral over the cellular region. The two detected types of glucuronic metabolites can be explained by the composition of UGT (UGT1A1, 1A4, 2B7, 2B15, and others are abundant in the gastrointestinal tract) [
23]. These metabolites also differ from each other in the use of ABC transporters. The result shows that the first metabolite passed to the apical side at a lower percentage (32.4 ± 2.7%) than to the basolateral side (64.1 ± 6.7%). The second metabolite was accumulated preferentially at the apical side (67.1 ± 0.2%), followed by the basolateral side (17.3 ± 2.7%). This could be due to a preference for the BRCP or MRP2. Finally, tracking the timeline of metabolite passage to the basolateral side shows that in the first 30 min of the experiment, the first metabolite can be detected. In contrast, the second metabolite began to be released after 1 h.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}


