
Characterization of Vegetative Bacillus cereus and Bacillus subtilis Strains Isolated from Processed Cheese Products in an Italian Dairy Plant

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
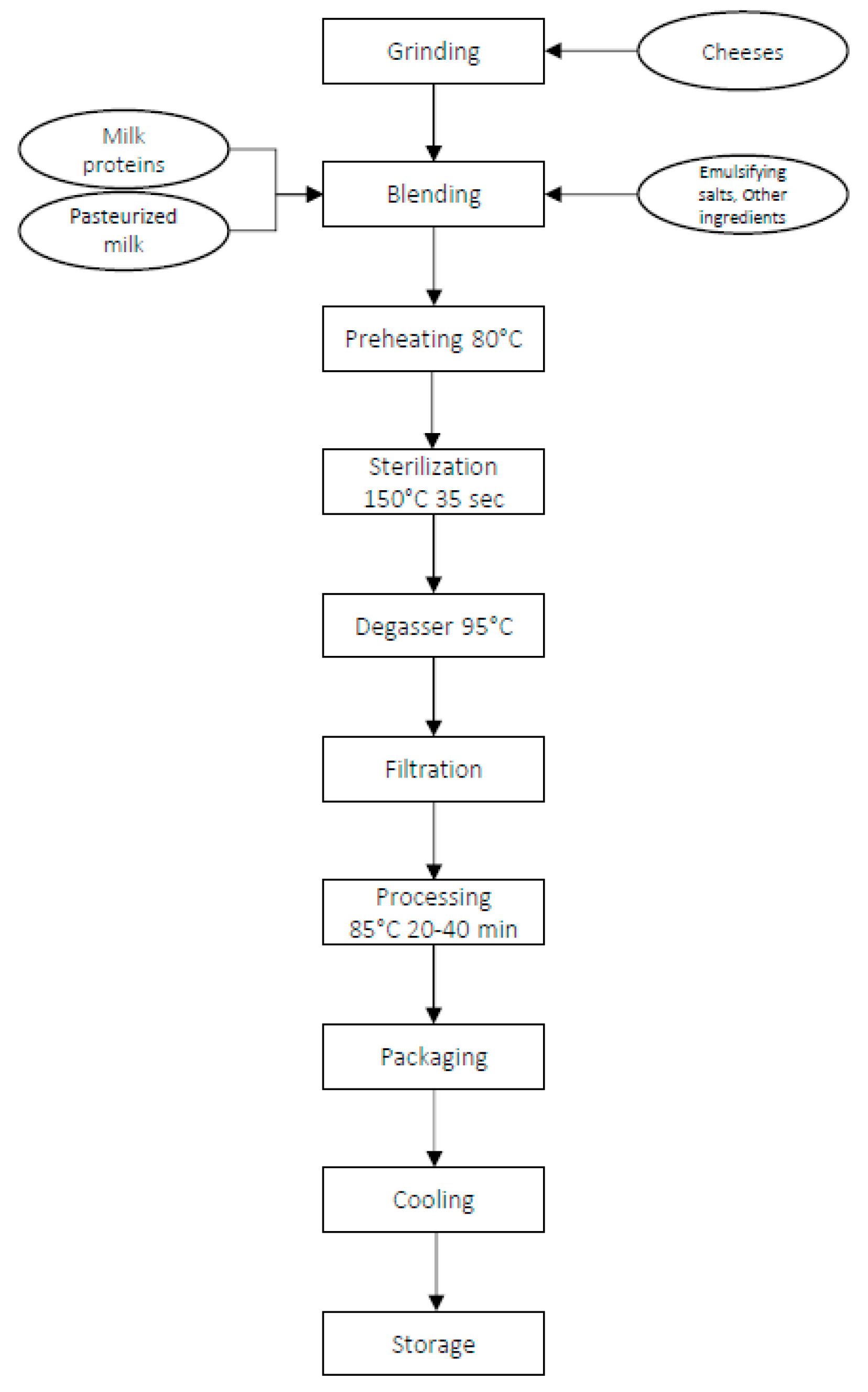
2.1. Experimental Design and Sample Collection
2.2. Isolation and Enumeration of Bacterial Colonies
2.3. DNA Extraction
2.4. Isolated Colonies Identification by Matrix Assisted Laser Desorption Ionization-Time of Flight Mass Spectrometry (MALDI-TOF MS) and 16S rRNA Sequencing
2.5. Characterization by Means of MLST Analysis
2.6. Analysis of Toxin Genes
2.7. Screening of Biofilm Forming Strains Using Micro-Method Assay
2.8. Statistical Analysis
3. Results
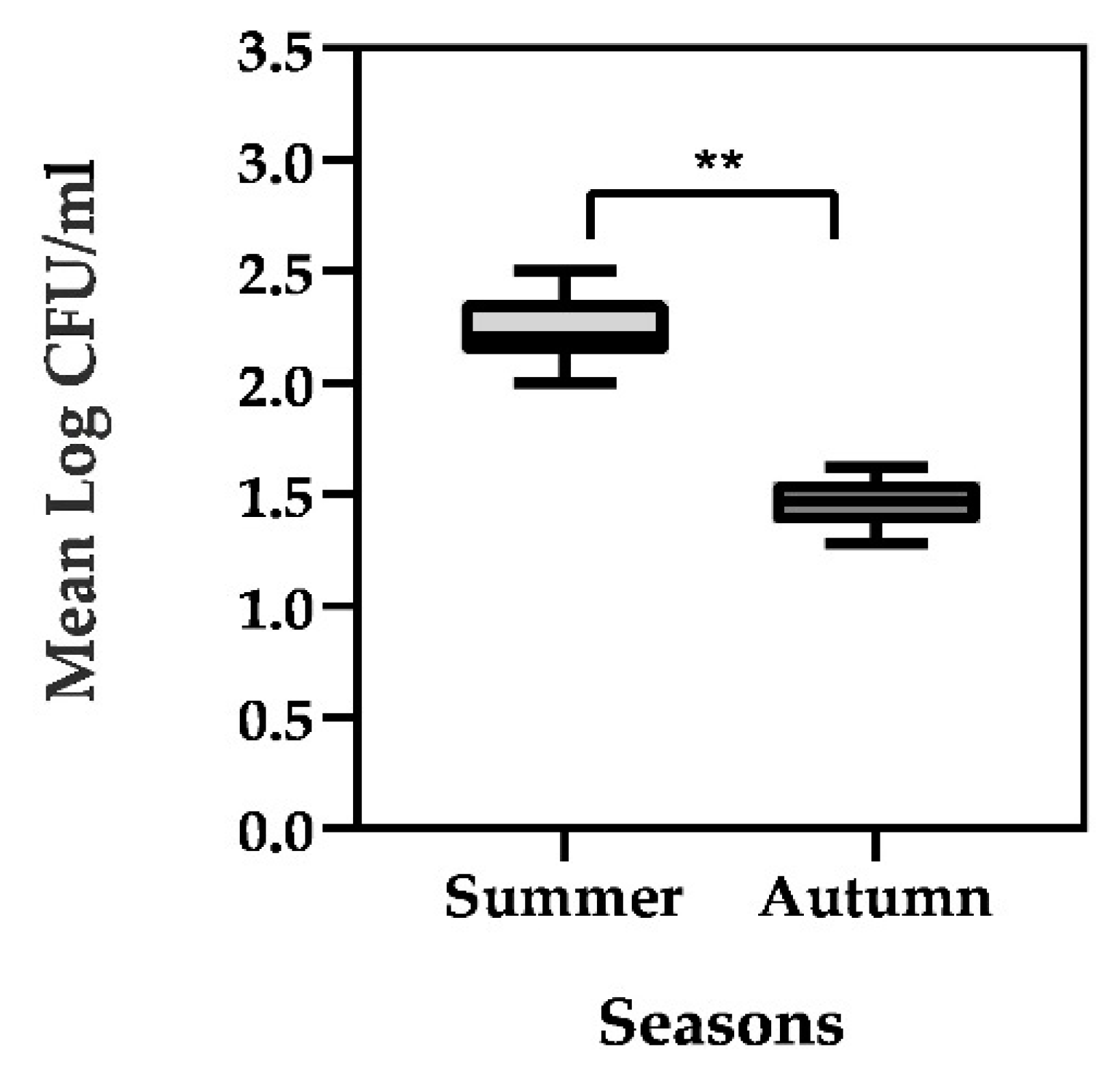
3.1. Enumeration of Vegetative Bacterial Contamination and Identification of Isolates
3.2. Molecular Characterization by MLST Analysis
3.3. Prevalence of Toxigenic Determinants in Bacillus cereus Isolates
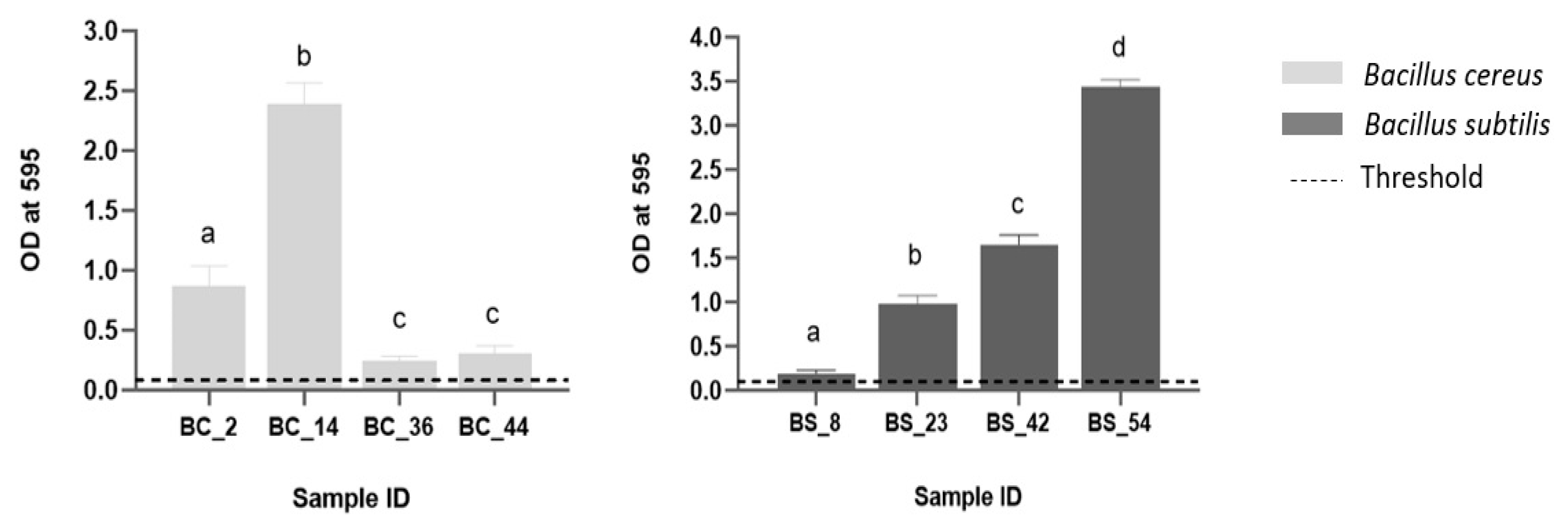
3.4. Biofilm Forming Potential
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Guinee, T.P. Pasteurized processed and imitation cheese products. In Cheese Chemistry, Physics & Microbiology, 4th ed.; Academic Press: Cambridge, MA, USA, 2017; Chapter 46; pp. 1133–1184. [Google Scholar] [CrossRef]
- Tamime, A.Y. Processed cheese and analogues: An overview. In Processed Cheese and Analogues, 1st ed.; Tamime, A., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2011; pp. 1–24. [Google Scholar] [CrossRef]
- Oliveira, R.B.A.; Margalho, L.P.; Nascimento, J.S.; Costa, L.E.O.; Portela, J.B.; Cruz, A.G.; Sant’Ana, A.S. Processed cheese contamination by spore-forming bacteria: A review of sources, routes, fate during processing and control. Trends Food Sci. Technol. 2016, 57, 11–19. [Google Scholar] [CrossRef]
- Postollec, F.; Mathot, A.G.; Bernard, M.; Divanac’h, M.L.; Pavan, S.; Sohier, D. Tracking spore-forming bacteria in food: From natural biodiversity to selection by processes. Int. J. Food Microbiol. 2012, 158, 1–8. [Google Scholar] [CrossRef]
- Glass, K.; Doyle, M.E. Safety of processed cheese. Food Research Institute, 2005. Available online: https://fri.wisc.edu/files/Briefs_File/ProcCheese_May2005_v2.pdf (accessed on 5 May 2021).
- Watterson, M.J.; Kent, D.J.; Boor, K.J.; Wiedemann, M.; Martin, N.H. Evaluation of dairy powder products implicates thermophilic spore formers as the primary organisms of interest. J. Dairy Sci. 2014, 97, 2487–2497. [Google Scholar] [CrossRef] [Green Version]
- Scott, S.A.; Brooks, J.D.; Rakonjac, J.; Walker, K.M.R.; Flint, S.H. The formation of thermophilic spores during the manufacture of whole milk powder. Int. J. Dairy Technol. 2007, 60, 109–117. [Google Scholar] [CrossRef]
- Luu, S.; Cruz-Mora, J.; Setlow, B.; Feeherry, F.E.; Doona, C.J.; Setlow, P. The effects of heat activation on Bacillus spore germination, with nutrients or under high pressure, with or without various germination proteins. Appl. Environ. Microbiol. 2015, 81, 2927–2938. [Google Scholar] [CrossRef] [Green Version]
- Setlow, P. Germination of spores of Bacillus species: What we know and do not know. J. Bacteriol. 2014, 196, 1297–1305. [Google Scholar] [CrossRef] [Green Version]
- Checinska, A.; Paszczynski, A.; Burbank, M. Bacillus and other spore-forming genera: Variations in responses and mechanisms for survival. Ann. Rev. Food Sci. Technol. 2015, 6, 351–369. [Google Scholar] [CrossRef]
- Stecchini, M.L.; Del Torre, M.; Polese, P. Survival strategies of Bacillus spores in food. Indian J. Exp. Biol. 2013, 51, 905–909. Available online: http://nopr.niscair.res.in/bitstream/123456789/23451/1/IJEB%2051%2811%29%20905-909.pdf (accessed on 5 May 2021). [PubMed]
- Bermúdez, J.; González, M.J.; Olivera, J.A.; Burgueño, J.A.; Juliano, P.; Fox, E.M.; Reginensi, S.M. Seasonal occurrence and molecular diversity of clostridia species spores along cheesemaking streams of 5 commercial dairy plants. J. Dairy Sci. 2016, 99, 3358–3366. [Google Scholar] [CrossRef] [Green Version]
- Montanhini, M.T.M.; Montanhini, R.N.; Pinto, J.P.N.; Bersot, L.S. Effect of temperature on the lipolytic and proteolytic activity of Bacillus cereus isolated from dairy products. Int. Food Res. J. 2013, 20, 1417–1420. Available online: http://www.ifrj.upm.edu.my/20%20(03)%202013/56%20IFRJ%2020%20(03)%202013%20Montanhini%20(423).pdf (accessed on 5 May 2021).
- Karaka, O.B.; Güven, M. Effects of Proteolytic and Lipolytic Enzyme Supplementations on Lipolysis and Proteolysis Characteristics of White Cheeses. Foods 2018, 7, 125. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Daniel, R.M.; Coolbear, T. Detection and impact of protease and lipase activities in milk and milk powder. Int. Dairy J. 2003, 13, 255–275. [Google Scholar] [CrossRef]
- Kumari, S.; Sarkar, P.K. Prevalence and characterization of Bacillus cereus group from various marketed dairy products in India. Dairy Sci. Technol. 2014, 94, 483–497. [Google Scholar] [CrossRef] [Green Version]
- Stenfors Arnesen, L.P.; Fagerlund, A.; Granum, P.E. From soil to gut: Bacillus cereus and its food poisoning toxins. FEMS Microbiol. Rev. 2008, 32, 579–606. [Google Scholar] [CrossRef] [Green Version]
- Salkinoja-Salonen, M.S.; Vuorio, R.; Andersson, M.A.; Kämpfer, P.; Andersson, M.C.; Honkanen-Buzalski, T.; Scoging, A.C. Toxigenic strains of Bacillus licheniformis related to food poisoning. Appl. Environ. Microbiol. 1999, 65, 4637–4645. [Google Scholar] [CrossRef] [Green Version]
- Suominen, I.; Andersson, M.A.; Andersson, M.C.; Hallaksela, A.M.; Kampfer, P.; Rainey, F.A.; Salkinoja-Salonen, M. Toxic Bacillus pumilus from indoor air, recycled paper pulp, Norway spruce, food poisoning outbreaks and clinical samples. Syst. Appl. Microbiol. 2001, 24, 267–276. [Google Scholar] [CrossRef]
- Mikkola, R.; Andersson, M.A.; Grigoriev, P.; Teplova, V.V.; Saris, N.E.; Rainey, F.A.; Salkinoja-Salonen, M.S. Bacillus amyloliquefaciens strains isolated from moisture-damaged buildings produced surfactin and a substance toxic to mammalian cells. Arch. Microbiol. 2004, 181, 314–323. [Google Scholar] [CrossRef]
- Gopal, N.; Hill, C.; Ross, P.R.; Beresford, T.P.; Fenelon, M.A.; Cotter, P.D. The Prevalence and Control of Bacillus and related spore-forming bacteria in the dairy industry. Front. Microbiol. 2015, 21, 1418. [Google Scholar] [CrossRef]
- Shemesh, M.; Ostrov, I. Role of Bacillus species in biofilm persistence and emerging antibiofilm strategies in the dairy industry. J. Sci. Food Agric. 2020, 100, 2327–2336. [Google Scholar] [CrossRef]
- Croxatto, A.; Prod’hom, G.; Greub, G. Applications of MALDI-TOF mass spectrometry in clinical diagnostic microbiology. FEMS Microbiol. Rev. 2012, 36, 380–407. [Google Scholar] [CrossRef]
- Huse, S.M.; Dethlefsen, L.; Huber, J.A.; Welch, D.M.; Relman, D.A.; Sogin, M.L. Exploring microbial diversity and taxonomy using SSU rRNA hypervariable tag sequencing. Plos Genet. 2008, 4, e1000255. [Google Scholar] [CrossRef]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Maiden, M.C.J. Multilocus sequence typing of bacteria. Annu. Rev. Microbiol. 2006, 60, 561–588. [Google Scholar] [CrossRef]
- Guinebretière, M.H.; Broussolle, V.; Nguyen-The, C. Enterotoxigenic profiles of food-poisoning and food-borne Bacillus cereus strains. J. Clin. Microbiol. 2002, 40, 3053–3056. [Google Scholar] [CrossRef] [Green Version]
- Yang, I.C.; Yang-Chih Shih, D.; Huang, T.P.; Huang, Y.P.; Wang, J.Y.; Pan, T.M. Establishment of a novel multiplex PCR assay and detection of toxigenic strains of the species in the Bacillus cereus group. J. Food Prot. 2005, 68, 2123–2130. [Google Scholar] [CrossRef]
- Stepanovic, S.; Vukovic, D.; Dakic, I.; Savic, B.; Svabic-Vlahovic, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Fernàndez-No, I.C.; Böhme, K.; Dìaz-Bao, M.; Cepeda, A.; Barros-Velàzquez, J.; Calo-Mata, P. Characterization and profiling of Bacillus subtilis, Bacillus cereus and Bacillus licheniformis by MALDI-TOF mass fingerprinting. Food Microbiol. 2013, 33, 235–242. [Google Scholar] [CrossRef]
- Zhao, S.; Chen, J.; Fei, P.; Feng, H.; Wang, Y.; Ali, M.A.; Li, S.; Jing, H.; Yang, W. Prevalence, molecular characterization, and antibiotic susceptibility of Bacillus cereus isolated from dairy products in China. J. Dairy Sci. 2020, 103, 3994–4001. [Google Scholar] [CrossRef]
- Hanson, M.L.; Wendor, W.L.; Houck, K.B. Effect of heat treatment of milk on activation of Bacillus spores. J. Food Prot. 2005, 68, 1484–1486. [Google Scholar] [CrossRef]
- Owusu-Kwarteng, J.; Wuni, A.; Akabanda, F.; Tano-Debrah, K.; Jespersen, L. Prevalence, virulence factor genes and antibiotic resistance of Bacillus cereus sensu lato isolated from dairy farms and traditional dairy products. BMC Microbiol. 2017, 17, 65. [Google Scholar] [CrossRef] [Green Version]
- Lücking, G.; Stoeckel, M.; Atamer, Z.; Hinrichs, J.; Ehling-Schulz, M. Characterization of aerobic spore-forming bacteria associated with industrial dairy processing environments and product spoilage. Int. J. Food Microbiol. 2013, 166, 270–279. [Google Scholar] [CrossRef]
- Larsen, H.D.; Jørgensen, K. The occurrence of Bacillus cereus in Danish pasteurized milk. Int. J. Food Microbiol. 1997, 34, 179–186. [Google Scholar] [CrossRef]
- Adame-Gómez, R.; Muñoz-Barrios, S.; Castro-Alarcón, N.; Leyva-Vázquez, M.A.; Toribio-Jiménez, J.; Ramírez-Peralta, A. Prevalence of the Strains of Bacillus cereus Group in Artisanal Mexican Cheese. Foodborne Pathog. Dis. 2020, 17, 8–14. [Google Scholar] [CrossRef]
- Tirloni, E.; Stella, S.; Bernardi, C.; Mazzantini, D.; Celandroni, F.; Ghelardi, E. Identification and pathogenic potential of Bacillus cereus strains isolated from a dairy processing plant producing PDO taleggio cheese. Microorganisms 2020, 8, 949. [Google Scholar] [CrossRef]
- Liu, X.Y.; Hu, Q.; Xu, F.; Ding, S.Y.; Zhu, K. Characterization of Bacillus cereus in Dairy Products in China. Toxins 2020, 12, 454. [Google Scholar] [CrossRef] [PubMed]
- Linsday, D.; Robertson, R.; Fraser, R.; Engstrom, S.; Jordan, K. Heat induced inactivation of microorganism in milk and dairy products. Int. Dairy J. 2021, 121, 105096. [Google Scholar] [CrossRef]
- Ivy, R.A.; Ranieri, M.L.; Martin, N.H.; den Bakker, H.C.; Xavier, B.M.; Wiedmann, M.; Boor, K.J. Identification and characterization of psychrotolerant spore-formers associated with fluid milk production and processing. Appl. Environ. Microbiol. 2012, 78, 1853–1864. [Google Scholar] [CrossRef] [Green Version]
- Kumari, S.; Sarkar, P.K. Bacillus cereus hazard and control in industrial dairy processing environment. Food Control 2016, 69, 20–29. [Google Scholar] [CrossRef]
- Kwon, G.H.; Lee, H.A.; Park, J.Y.; Kim, J.S.; Lim, J.; Park, C.S.; Kwon, D.Y.; Kim, J.S.; Kim, J.H. Development of a RAPD-PCR method for identification of Bacillus species isolated from Cheonggukjang. Int. J. Food Microbiol. 2009, 129, 282–287. [Google Scholar] [CrossRef]
- Jiyeon, L.; Kwon, G.H.; Park, J.Y.; Park, C.S.; Kwon, D.Y.; Lim, J.; Kim, J.S.; Kim, J.H. A RAPD-PCR Method for the Rapid Detection of Bacillus cereus. J. Microbiol. Biotechnol. 2011, 21, 274–276. [Google Scholar] [CrossRef]
- Guinebretière, M.H.; Thompson, F.L.; Sorokin, A.; Normand, P.; Dawyndt, P.; Ehling-Schulz, M.; Svensson, B.; Sanchis, V.; Nguyen-The, C.; Heyndrickx, M.; et al. Ecological diversification in the Bacillus cereus Group. Environ. Microbiol. 2008, 10, 851–865. [Google Scholar] [CrossRef]
- Ki, J.S.; Zhang, W.; Qian, P.Y. Discovery of marine Bacillus species by 16S rRNA and rpoB comparisons and their usefulness for species identification. J. Microbiol. Methods 2009, 77, 48–57. [Google Scholar] [CrossRef]
- Cardazzzo, B.; Negrisolo, E.; Carraro, L.; Alberghini, L.; Patarnello, T.; Giaccone, V. Multiple-locus sequence typing and analysis of toxin genes in Bacillus cereus food-borne isolates. Appl. Environ. Microbiol. 2008, 74, 850–860. [Google Scholar] [CrossRef] [Green Version]
- Priest, F.G.; Barker, M.; Baille, L.W.J.; Holmes, E.C.; Maiden, M.C.J. Population structure and evolution of the Bacillus cereus group. J. Bacteriol. 2004, 186, 7959–7970. [Google Scholar] [CrossRef] [Green Version]
- Hansen, B.M.; Hendriksen, N.B. Detection of enterotoxic Bacillus cereus and Bacillus thuringiensis strains by PCR analysis. Appl. Environ. Microbiol. 2001, 67, 185–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veld, P.H.; Ritmeester, W.S.; Delfgou-van Asch, E.H.M.; Dufrenne, J.B.; Wernars, K.; Smit, E.; van Leusden, F.M. Detection of genes encoding for enterotoxins and determination of the production of enterotoxins by HBL blood plates and immunoassays of psychrotrophic strains of Bacillus cereus isolated from pasteurised milk. Int. J. Food Microbiol. 2001, 64, 63–70. [Google Scholar] [CrossRef]
- Carter, L.; Chase, H.R.; Gieseker, C.M.; Hasbrouck, N.R.; Stine, C.B.; Khan, A.; Ewing-Peeples, L.J.; Tall, B.D.; Gopinath, G.R. Analysis of enterotoxigenic Bacillus cereus strains from dried foods using whole genome sequencing, multi-locus sequence analysis and toxin gene prevalence and distribution using endpoint PCR analysis. Int. J. Food Microbiol. 2018, 284, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Guinebretière, M.H.; Velge, P.; Couvert, O.; Carlin, F.; Debuyser, M.L.; Nguyen-The, C. Ability of Bacillus cereus group strains to cause food poisoning varies according to phylogenetic affiliation (groups I to VII) rather than species affiliation. J. Clin. Microbiol. 2010, 48, 3388–3391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngamwongsatit, P.; Buasri, W.; Pianariyanon, P.; Pulsrikarn, C.; Ohba, M.; Assavanig, A.; Panbangred, W. Broad distribution of enterotoxin genes (hblCDA, nheABC, cytK, and entFM) among Bacillus thuringiensis and Bacillus cereus as shown by novel primers. Int. J. Food Microbiol. 2008, 121, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Gao, T.; Ding, Y.; Wu, Q.; Wang, J.; Zhang, J.; Yu, S.; Yu, P.; Liu, C.; Kong, L.; Feng, Z.; et al. Prevalence, virulence genes, antimicrobial susceptibility, and genetic diversity of Bacillus cereus isolated from pasteurized milk in China. Front. Microbiol. 2018, 9, 533. [Google Scholar] [CrossRef] [Green Version]
- Moravek, M.; Dietrich, R.; Buerk, C.; Broussolle, V.; Guinebretière, M.H.; Granum, P.E.; Nguyen-the, C.; Martlbauer, E. Determination of the toxic potential of Bacillus cereus isolates by quantitative enterotoxin analyses. FEMS Microbiol. Lett. 2006, 257, 293–298. [Google Scholar] [CrossRef] [Green Version]
- Mehta, A.; Tatini, S.R. An evaluation of the microbiological safety of reduced-fat cheddar-like cheese. J. Food Prot. 1994, 57, 776–779. [Google Scholar] [CrossRef] [PubMed]
- Yang, R.; Xie, Y.; Lombardo, S.P.; Tang, J. Oil protects bacteria from humid heat in thermal processing. Food Control 2021, 123, 107690. [Google Scholar] [CrossRef]
- Ostrov, I.; Harel, A.; Bernstein, S.; Steinberg, D.; Shemesh, M. Development of a method to determine the effective-ness of cleaning agents in removal of biofilm derived spores in milking system. Front. Microbiol. 2016, 7, 1498. [Google Scholar] [CrossRef] [PubMed]
- Parkar, S.G.; Flint, S.H.; Brooks, J.D. Physiology of biofilms of thermophilic bacilli-potential consequences for cleaning. J. Ind. Microbiol. Biotechnol. 2003, 30, 553–560. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Weeks | Season | Total Number of Colonies Identified | Bacillus subtilis Identified | Bacillus cereus Identified |
|---|---|---|---|---|
| I | Summer | 13 | 5 (38.5%) | 8 (61.5%) |
| II | 12 | 10 (83.3%) | 2 (16.7%) | |
| III | 12 | 6 (50.0%) | 6 (50.0%) | |
| IV | 14 | 13 (92.9%) | 1 (7.1%) | |
| V | Autumn | 19 | 15 (78.9%) | 4 (21.1%) |
| VI | 12 | 8 (66.7%) | 4 (33.3%) | |
| VII | 12 | 10 (83.3%) | 2 (16.7%) | |
| VIII | 9 | 5 (55.6%) | 4 (44.4%) |
| (A) | ||||||||
|---|---|---|---|---|---|---|---|---|
| Sequence Type | Allelic Type | Number of Isolates | ||||||
| glpF | gmk | ilvD | pta | purH | pycA | tpiA | ||
| ST-32 | 5 | 4 | 3 | 4 | 15 | 6 | 16 | 1/14 |
| ST-371 | 21 | 2 | 19 | 5 | 32 | 18 | 2 | 13/14 |
| (B) | ||||||||
| Sequence Type | Allelic Type | Number of Isolates | ||||||
| glpF | ilvD | pta | purH | pycA | rpoD | tpiA | ||
| ST-249 | 3 | 89 | 35 | 4 | 108 | 3 | 4 | 21/21 |
| (new) | (new) | |||||||
| Strains | HBL Complex | NHE Complex | Other Enterotoxins | Emetic Strains | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| hblA | hblC | hblD | nheA | nheB | nheC | entFM | bceT | cytK | ces | |
| BC_2 | − | − | − | + | + | + | + | − | + | − |
| BC_3 | − | − | − | + | + | + | + | − | + | − |
| BC_5 | − | − | − | + | + | + | + | − | + | − |
| BC_14 | − | − | − | + | + | + | + | − | + | − |
| BC_19 | − | − | − | + | + | + | + | − | + | − |
| BC_22 | − | − | − | + | + | + | + | − | + | − |
| BC_24 | − | − | − | + | + | + | + | − | + | − |
| BC_26 | − | − | − | + | + | + | + | − | + | − |
| BC_29 | − | − | − | + | + | + | + | − | + | − |
| BC_35 | − | − | − | + | + | + | + | − | + | − |
| BC_36 | − | − | − | + | + | + | + | − | + | − |
| BC_38 | − | − | − | + | + | + | + | − | + | − |
| BC_40 | − | − | − | + | + | + | + | − | + | − |
| BC_44 | − | − | − | + | + | + | + | − | + | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Catania, A.M.; Civera, T.; Di Ciccio, P.A.; Grassi, M.A.; Morra, P.; Dalmasso, A. Characterization of Vegetative Bacillus cereus and Bacillus subtilis Strains Isolated from Processed Cheese Products in an Italian Dairy Plant. Foods 2021, 10, 2876. https://doi.org/10.3390/foods10112876
Catania AM, Civera T, Di Ciccio PA, Grassi MA, Morra P, Dalmasso A. Characterization of Vegetative Bacillus cereus and Bacillus subtilis Strains Isolated from Processed Cheese Products in an Italian Dairy Plant. Foods. 2021; 10(11):2876. https://doi.org/10.3390/foods10112876
Chicago/Turabian StyleCatania, Angela Maria, Tiziana Civera, Pierluigi Aldo Di Ciccio, Maria Ausilia Grassi, Patrizia Morra, and Alessandra Dalmasso. 2021. "Characterization of Vegetative Bacillus cereus and Bacillus subtilis Strains Isolated from Processed Cheese Products in an Italian Dairy Plant" Foods 10, no. 11: 2876. https://doi.org/10.3390/foods10112876
APA StyleCatania, A. M., Civera, T., Di Ciccio, P. A., Grassi, M. A., Morra, P., & Dalmasso, A. (2021). Characterization of Vegetative Bacillus cereus and Bacillus subtilis Strains Isolated from Processed Cheese Products in an Italian Dairy Plant. Foods, 10(11), 2876. https://doi.org/10.3390/foods10112876