
Synergy of Nitric Oxide and 1-Methylcyclopropene Treatment in Prolong Ripening and Senescence of Peach Fruit

 ,
,
Abstract
:
1. Introduction
2. Material and Methods
2.1. Peach Material and Treatment
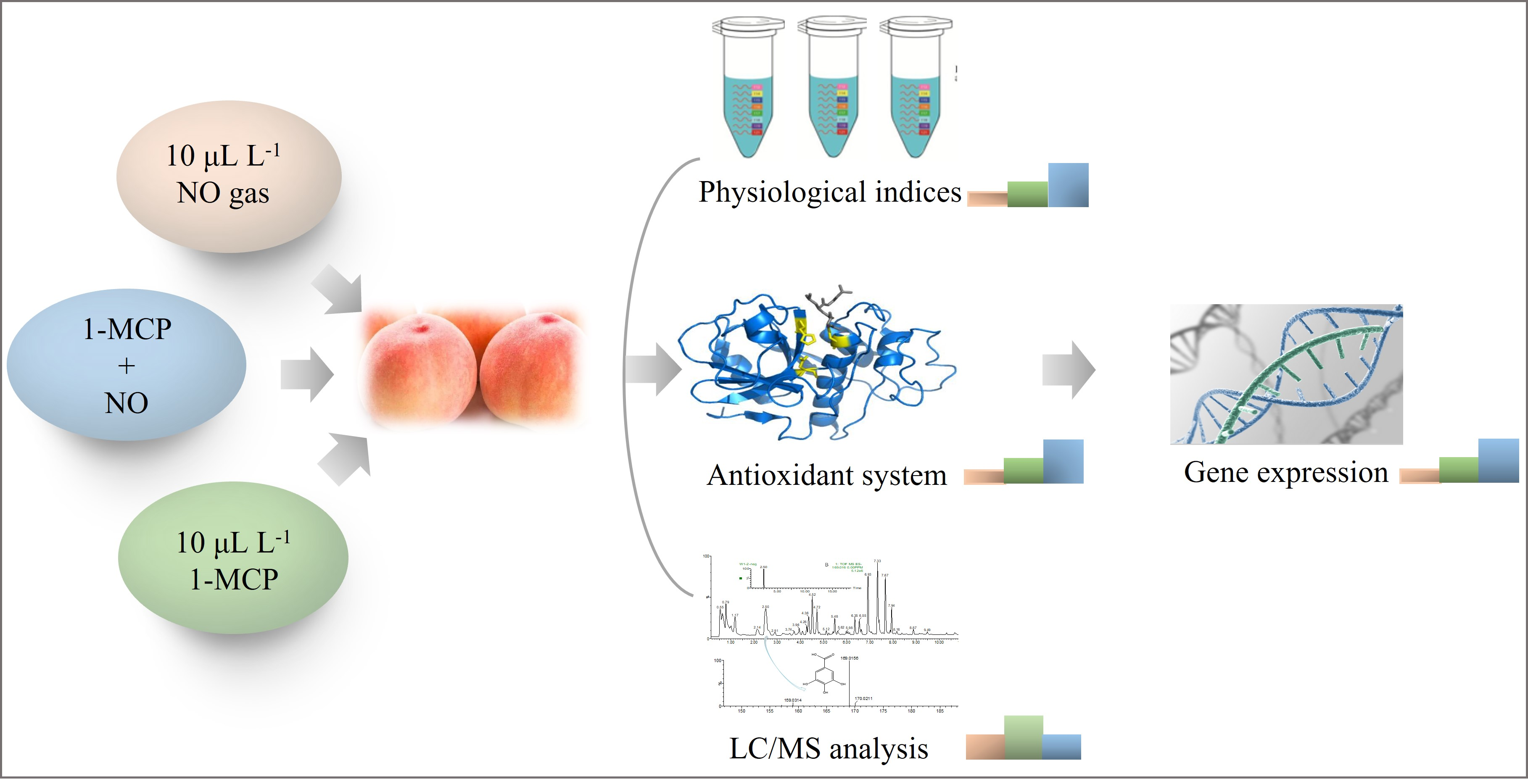
2.2. Respiratory Rate and Ethylene Production
2.3. Firmness, MDA, H2O2 and O2− Detection
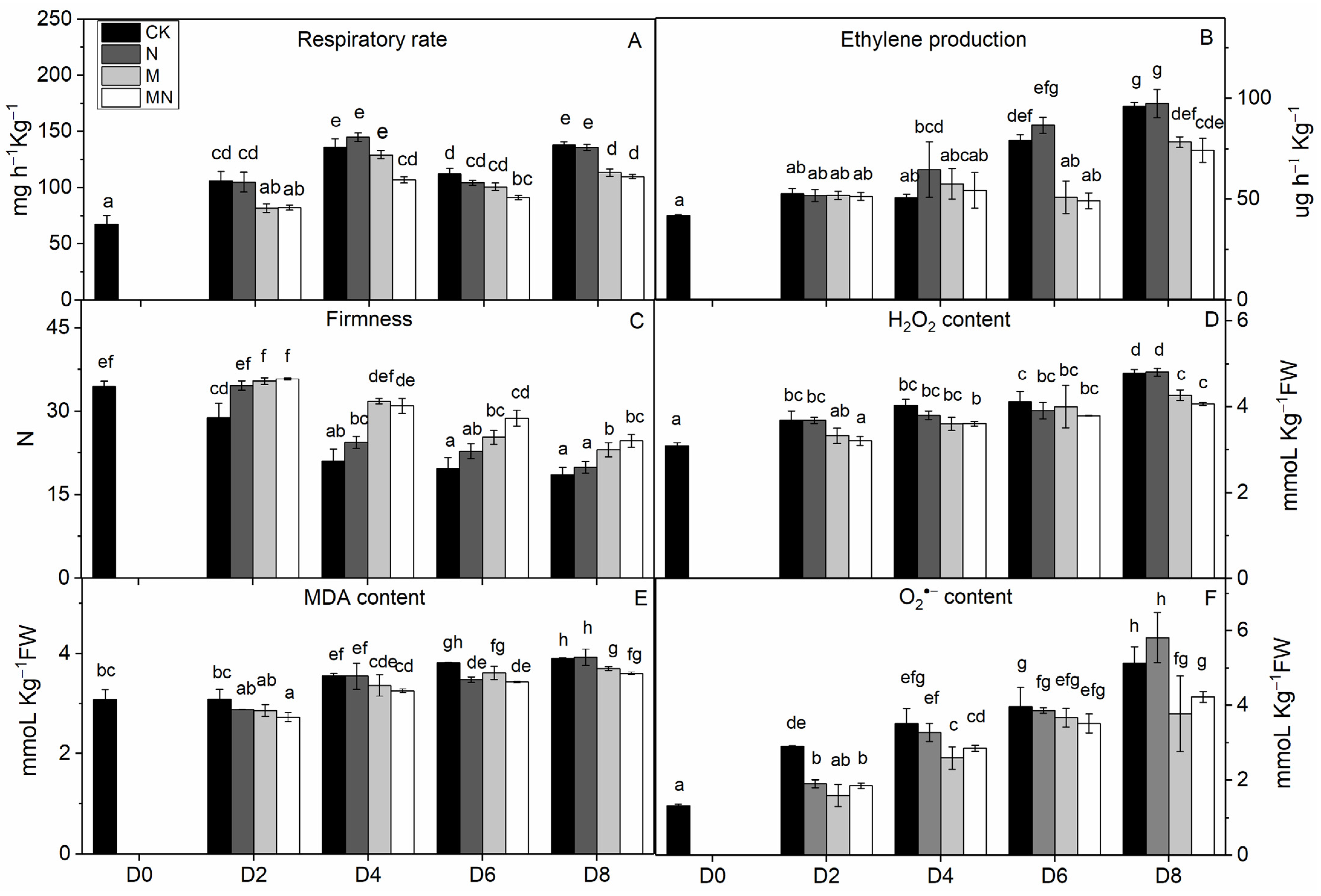
2.4. Enzymatic Assays
2.5. RNA Isolation and Gene Expression Analysis
2.6. LC/MS Analysis of Phenols
2.7. Statistical Analysis
3. Results and Discussion
3.1. Firmness
3.2. Respiration and Ethylene Production
3.3. Reactive Oxygen Species (ROS) Production
3.4. Enzymatic Activity
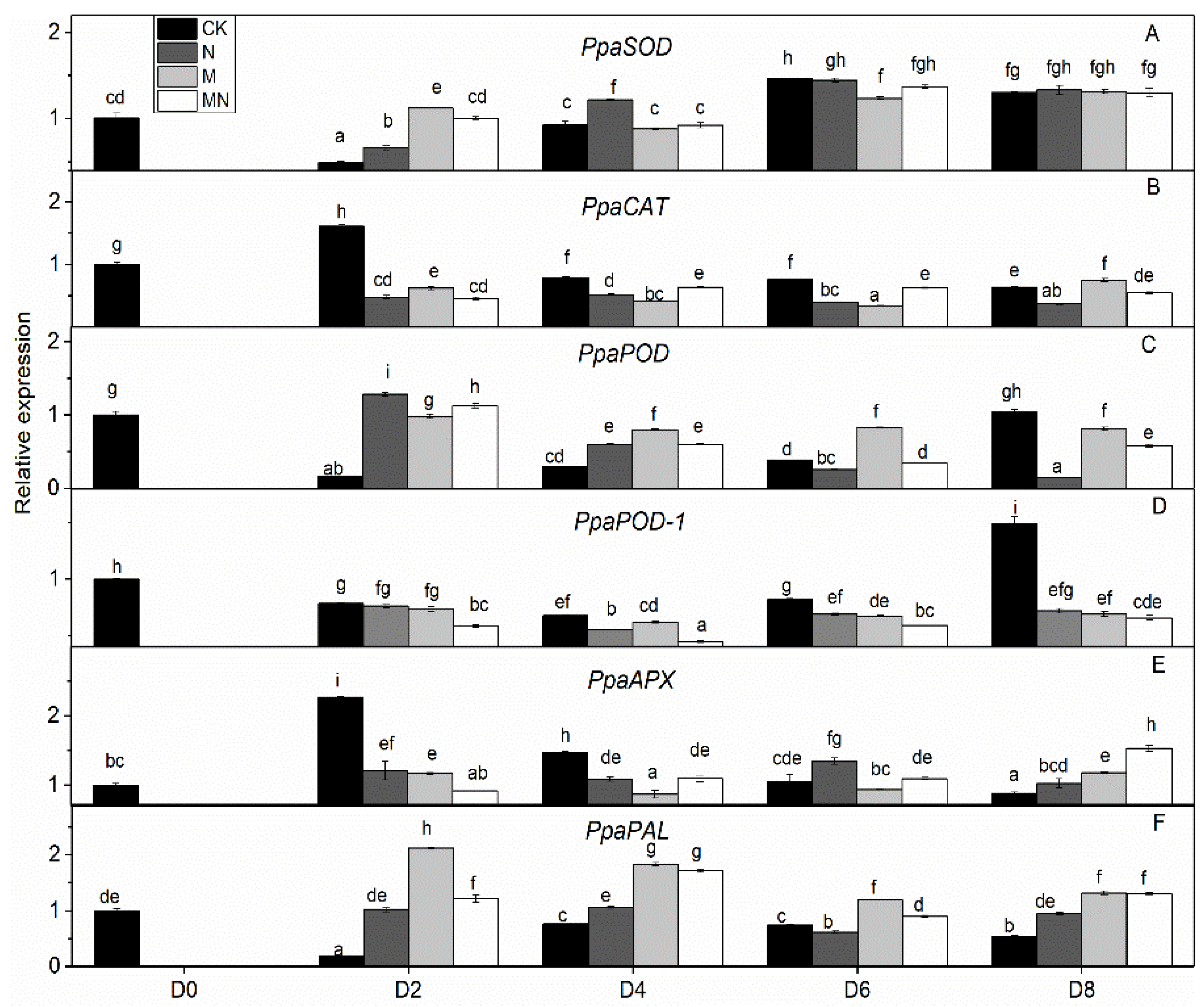
3.5. Gene Expression Analysis
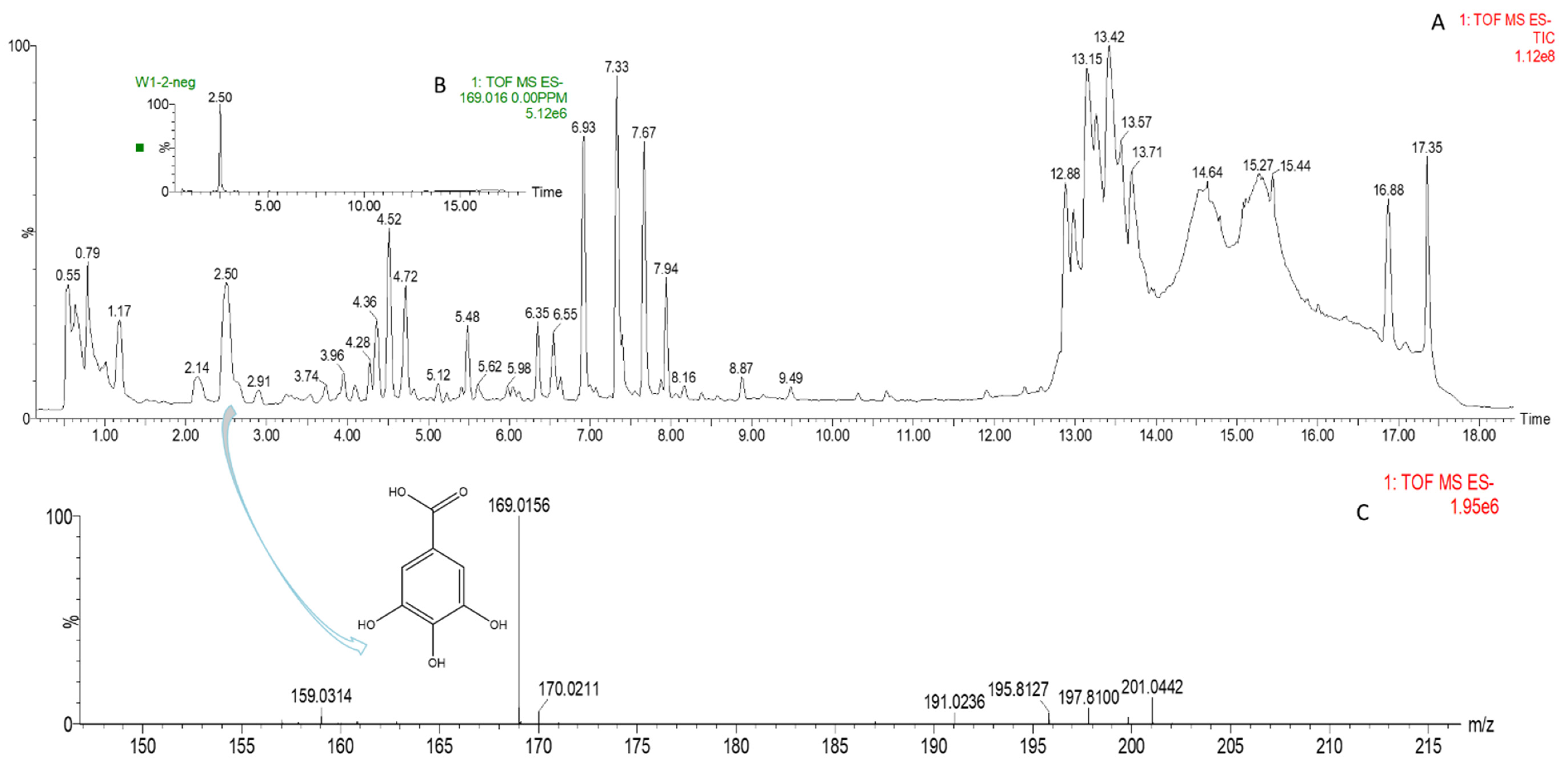
3.6. LC/MS Analysis of Phenols
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jiang, L.; Zhang, L.; Shi, Y.; Lu, Z.X.; Yu, Z.F. Proteomic analysis of peach fruit during ripening upon post-harvest heat combined with 1-MCP treatment. J. Proteom. 2014, 98, 31–43. [Google Scholar] [CrossRef] [PubMed]
- Kang, R.Y.; Zhang, L.; Jiang, L.; Yu, M.L.; Ma, R.J.; Yu, Z.F. Effect of postharvest nitric oxide treatment on the proteome of peach fruit during ripening. Postharvest Biol. Technol. 2016, 112, 277–289. [Google Scholar] [CrossRef]
- Cai, H.F.; An, X.J.; Han, S.; Jiang, L.; Yu, M.L.; Ma, R.J.; Yu, Z.F. Effect of 1-MCP on the production of volatiles and biosynthesis-related gene expression in peach fruit during cold storage. Postharvest Biol. Technol. 2018, 141, 50–57. [Google Scholar] [CrossRef]
- Neill, S.J.; Desikan, R.; Clarke, A.; Hancock, J.T. Nitric oxide is a novel component of abscisic acid signaling in stomatal guard cells. Plant Physiol. 2002, 128, 13–16. [Google Scholar] [CrossRef]
- Han, S.; Cai, H.F.; An, X.J.; Huan, C.; Wu, X.Q.; Jiang, L.; Yu, M.L.; Ma, R.J.; Yu, Z.F. Effect of nitric oxide on sugar metabolism in peach fruit (cv. Xiahui 6) during cold storage. Postharvest Biol. Technol. 2018, 142, 72–80. [Google Scholar] [CrossRef]
- Ren, Y.; Xue, Y.; Tian, D.; Zhang, L.; Xiao, G.; He, J. Improvement of Postharvest Anthracnose Resistance in Mango Fruit by Nitric Oxide and the Possible Mechanisms Involved. J. Agric. Food Chem. 2020, 68, 15460–15467. [Google Scholar] [CrossRef]
- Flores, F.B.; Sanchez-Bel, P.; Valdenegro, M.; Romojaro, F.; Martinez-Madrid, M.C.; Egea, M.I. Effects of a pretreatment with nitric oxide on peach (Prunus persica L.) storage at room temperature. Eur. Food Res. Technol. 2008, 227, 1599–1611. [Google Scholar] [CrossRef]
- Huan, C.; Jiang, L.; An, X.J.; Kang, R.Y.; Yu, M.L.; Ma, R.J.; Yu, Z.F. Potential role of glutathione peroxidase gene family in peach fruit ripening under combined postharvest treatment with heat and 1-MCP. Postharvest Biol. Technol. 2016, 111, 175–184. [Google Scholar] [CrossRef]
- Al Ubeed, H.M.S.; Wills, R.B.H.; Bowyer, M.C.; Golding, J.B. Comparison of hydrogen sulphide with 1-methylcyclopropene (1-MCP) to inhibit senescence of the leafy vegetable, pak choy. Postharvest Biol. Technol. 2018, 137, 129–133. [Google Scholar] [CrossRef]
- Arasimowicz, M.; Floryszak-Wieczorek, J. Nitric oxide as a bioactive signalling molecule in plant stress responses. Plant Sci. 2007, 172, 876–887. [Google Scholar] [CrossRef]
- Grozeff, G.E.G.; Alegre, M.L.; Senn, M.E.; Chaves, A.R.; Simontacchi, M.; Bartoli, C.G. Combination of nitric oxide and 1-MCP on postharvest life of the blueberry (Vaccinium spp.) fruit. Postharvest Biol. Technol. 2017, 133, 72–80. [Google Scholar] [CrossRef]
- Chen, H.; An, X.; Yu, M.; Li, J.; Ma, R.; Tu, M.; Yu, Z. Effect of combined heat and 1-MCP treatment on the quality and antioxidant level of peach fruit during storage. Postharvest Biol. Technol. 2018, 145, 193–202. [Google Scholar]
- Wu, X.Q.; Mason, A.M.; Yu, M.L.; Ma, R.J.; Yu, Z.F. Quantitative proteomic analysis of pre- and post-harvest peach fruit ripening based on iTRAQ technique. Acta Physiol. Plant 2017, 39, 181. [Google Scholar] [CrossRef]
- Wu, X.Q.; Jiang, L.; Yu, M.L.; An, X.J.; Ma, R.J.; Yu, Z.F. Proteomic analysis of changes in mitochondrial protein expression during peach fruit ripening and senescence. J. Proteom. 2016, 147, 197–211. [Google Scholar] [CrossRef]
- Zhang, Z.Q.; Pang, X.Q.; Duan, X.W.; Ji, Z.L.; Jiang, Y.M. Role of peroxidase in anthocyanin degradation in litchi fruit pericarp. Food Chem. 2005, 90, 47–52. [Google Scholar] [CrossRef]
- Yingsanga, P.; Srilaong, V.; Kanlayanarat, S.; Noichinda, S.; McGlasson, W.B. Relationship between browning and related enzymes (PAL, PPO and POD) in rambutan fruit (Nephelium lappaceum Linn.) cvs. Rongrien and See-Chompoo. Postharvest Biol. Technol. 2008, 50, 164–168. [Google Scholar] [CrossRef]
- Song, H.W.; Yuan, W.M.; Jin, P.; Wang, W.; Wang, X.F.; Yang, L.M.; Zhang, Y.F. Effects of chitosan/nano-silica on postharvest quality and antioxidant capacity of loquat fruit during cold storage. Postharvest Biol. Technol. 2016, 119, 41–48. [Google Scholar] [CrossRef]
- Assis, J.S.; Maldonado, R.; Munoz, T.; Escribano, M.I.; Merodio, C. Effect of high carbon dioxide concentration on PAL activity and phenolic contents in ripening cherimoya fruit. Postharvest Biol. Technol. 2001, 23, 33–39. [Google Scholar] [CrossRef] [Green Version]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Huan, C.; Han, S.; Jiang, L.; An, X.J.; Yu, M.L.; Xu, Y.; Ma, R.J.; Yu, Z.F. Postharvest hot air and hot water treatments affect the antioxidant system in peach fruit during refrigerated storage. Postharvest Biol. Technol. 2017, 126, 1–14. [Google Scholar] [CrossRef]
- Tong, Z.; Gao, Z.; Wang, F.; Zhou, J.; Zhang, Z. Selection of reliable reference genes for gene expression studies in peach using real-time PCR. BMC Mol. Biol. 2009, 10, 71. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; An, X.; Yu, M.; Ma, R.; Yu, Z. 1-Methylcyclopropene Treatment on Phenolics and the Antioxidant System in Postharvest Peach Combined with the Liquid Chromatography/Mass Spectrometry Technique. J. Agric. Food Chem. 2018, 66, 6364–6372. [Google Scholar] [CrossRef] [PubMed]
- Sisler, E.C.; Dupille, E.; Serek, M. Effect of 1-methylcyclopropene and methylenecyclopropane on ethylene binding and ethylene action on cut carnations. Plant Growth Regul. 1996, 18, 79–86. [Google Scholar] [CrossRef]
- Crisosto, C.H.; Day, K.R.; Crisosto, G.M.; Garner, D. Quality attributes of white flesh peaches and nectarines grown under California conditions. J. Amer. Pomol. Soc. 2001, 55, 45–51. [Google Scholar]
- Lu, G.H.; Li, C.J.; Lu, Z.H. Wound-Induced Respiration in Thin Slice of Chinese Jujube Fruit. J. Plant Physiol. 1993, 141, 115–119. [Google Scholar] [CrossRef]
- Dal Cin, V.; Rizzini, F.M.; Botton, A.; Tonutti, P. The ethylene biosynthetic and signal transduction pathways are differently affected by 1-MCP in apple and peach fruit. Postharvest Biol. Technol. 2006, 42, 125–133. [Google Scholar] [CrossRef]
- Zhu, S.H.; Liu, M.C.; Zhou, J. Inhibition by nitric oxide of ethylene biosynthesis and lipoxygenase activity in peach fruit during storage. Postharvest Biol. Technol. 2006, 42, 41–48. [Google Scholar] [CrossRef]
- Sabban-Amin, R.; Feygenberg, O.; Belausov, E.; Pesis, E. Low oxygen and 1-MCP pretreatments delay superficial scald development by reducing reactive oxygen species (ROS) accumulation in stored ‘Granny Smith’ apples. Postharvest Biol. Technol. 2011, 62, 295–304. [Google Scholar] [CrossRef]
- Xu, X.; Lei, H.; Ma, X.; Lai, T.; Song, H.; Shi, X.; Li, J. Antifungal activity of 1-methylcyclopropene (1-MCP) against anthracnose (Colletotrichum gloeosporioides) in postharvest mango fruit and its possible mechanisms of action. Int. J. Food Microbiol. 2017, 241, 1–6. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhu, X.; Hou, Y.; Wang, X.; Li, X. Postharvest nitric oxide treatment delays the senescence of winter jujube (Zizyphus jujuba Mill. cv. Dongzao) fruit during cold storage by regulating reactive oxygen species metabolism. Sci. Hortic. 2020, 261, 109009. [Google Scholar] [CrossRef]
- Lin, Y.; Chen, G.; Lin, H.; Lin, M.; Wang, H.; Lin, Y. Chitosan postharvest treatment suppresses the pulp breakdown development of longan fruit through regulating ROS metabolism—ScienceDirect. Int. J. Biol. Macromol. 2020, 165, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Mittler, R. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci. 2002, 7, 405–410. [Google Scholar] [CrossRef]
- Zhang, D.; Xu, X.; Zhang, Z.; Jiang, G.; Feng, L.; Duan, X.; Jiang, Y. 6-Benzylaminopurine improves the quality of harvested litchi fruit. Postharvest Biol. Technol. 2018, 143, 137–142. [Google Scholar] [CrossRef]
- Hu, H.; Liu, Y.; Shi, G.L.; Liu, Y.P.; Wu, R.J.; Yang, A.Z.; Wang, Y.M.; Hua, B.G.; Wang, Y.N. Proteomic analysis of peach endocarp and mesocarp during early fruit development. Physiol. Plant 2011, 142, 390–406. [Google Scholar] [CrossRef] [PubMed]
- Pandey, V.P.; Singh, S.; Jaiswal, N.; Awasthi, M.; Pandey, B.; Dwivedi, U.N. Papaya fruit ripening: ROS metabolism, gene cloning, characterization and molecular docking of peroxidase. J. Mol. Catal. B Enzym. 2013, 98, 98–105. [Google Scholar] [CrossRef]
- Tang, R.X.; Yong-Hong, G.E.; Can-Ying, L.I.; Sun, R.H. Effect of Brassinolide Treatment on the Active Oxygen Metabolism of Postharvest ‘Okubao’ Peach. Storage Process 2016, 16, 5–9. [Google Scholar]
- Shi, T.; Li, Z.; Zhang, Z.; Zhang, C.; Gao, Z. Effect of 1-methylcyclopropene (1- MCP) treatment on antioxidant enzymes of postharvest Japanese apricot. Afr. J. Biotechnol. 2013, 12, 689–694. [Google Scholar]
- Clark, D.; Durner, J.; Navarre, D.A.; Klessig, D.F. Nitric oxide inhibition of tobacco catalase and ascorbate peroxidase. Mol. Plant-Microbe Interact. 2000, 13, 1380–1384. [Google Scholar] [CrossRef] [Green Version]
- Zhu, S.H.; Sun, L.; Liu, M.C.; Zhou, J. Effect of nitric oxide on reactive oxygen species and antioxidant enzymes in kiwifruit during storage. J. Sci. Food Agric. 2008, 88, 2324–2331. [Google Scholar] [CrossRef]
- Huan, C.; Jiang, L.; An, X.J.; Yu, M.L.; Xu, Y.; Ma, R.J.; Yu, Z.F. Potential role of reactive oxygen species and antioxidant genes in the regulation of peach fruit development and ripening. Plant Physiol. Biochem. 2016, 104, 294–303. [Google Scholar] [CrossRef]
- Yang, N.; Wang, C.; Chen, X.; Yishen, L.I.; Zhang, X. Effect of Drought Stress on Antioxidant System and Reactive Oxygen in Chorispora bungeana Plantlets in vitro. Acta Bot. Boreali Occident. Sin. 2014, 34, 2483–2490. [Google Scholar]
- Dixon, R.A.; Paiva, N.L. Stress-Induced Phenylpropanoid Metabolism. Plant Cell 1995, 7, 1085–1097. [Google Scholar] [CrossRef]
- Zheng, X.L.; Hu, B.; Song, L.J.; Pan, J.; Liu, M.M. Changes in quality and defense resistance of kiwifruit in response to nitric oxide treatment during storage at room temperature. Sci. Hortic. 2017, 222, 187–192. [Google Scholar] [CrossRef]
- Kovacik, J.; Klejdus, B.; Backor, M. Nitric oxide signals ROS scavenger-mediated enhancement of PAL activity in nitrogen-deficient Matricaria chamomilla roots: Side effects of scavengers. Free Radic. Biol. Med. 2009, 46, 1686–1693. [Google Scholar] [CrossRef]
- Lai, T.F.; Wang, Y.Y.; Li, B.Q.; Qin, G.Z.; Tian, S.P. Defense responses of tomato fruit to exogenous nitric oxide during postharvest storage. Postharvest Biol. Technol. 2011, 62, 127–132. [Google Scholar] [CrossRef]
- Shao, X.F.; Wang, H.F.; Xu, F.; Cheng, S. Effects and possible mechanisms of tea tree oil vapor treatment on the main disease in postharvest strawberry fruit. Postharvest Biol. Technol. 2013, 77, 94–101. [Google Scholar] [CrossRef]
- Cao, S.F.; Zheng, Y.H.; Wang, K.T.; Rui, H.J.; Shang, H.T.; Tang, S.S. The effects of 1-methylcyclopropene on chilling and cell wall metabolism in loquat fruit. J. Hortic. Sci. Biotechnol. 2010, 85, 147–153. [Google Scholar] [CrossRef]
- Ozkaya, O.; Yildirim, D.; Dundar, O.; Tukel, S.S. Effects of 1-methylcyclopropene (1-MCP) and modified atmosphere packaging on postharvest storage quality of nectarine fruit. Sci. Hortic. 2016, 198, 454–461. [Google Scholar] [CrossRef]
- Constabel, C.P.; Bergey, D.R.; Ryan, C.A. Systemin activates synthesis of wound-inducible tomato leaf polyphenol oxidase via the octadecanoid defense signaling pathway. Proc. Natl. Acad. Sci. USA 1995, 92, 407–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Ballesta, M.T.; Lafuente, M.T.; Zacarias, L.; Granell, A. Involvement of phenylalanine ammonia-lyase in the response of Fortune mandarin fruits to cold temperature. Physiol. Plant 2000, 108, 382–389. [Google Scholar] [CrossRef]
- Liu, X.; Cui, X.; Ji, D.; Zhang, Z.; Tian, S. Luteolin-induced activation of the phenylpropanoid metabolic pathway contributes to quality maintenance and disease resistance of sweet cherry. Food Chem. 2020, 342, 128309. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Peng, W.; Zhang, Y.; Yuanyuan, L.I.; Wang, L.; Shan, T.; Jin, P.; Zheng, Y. Effects of Nitric Oxide Treatment on Quality and Phenolic Metabolism in Strawberry Fruit. J. Nucl. Agric. Sci. 2016, 30, 1959–1966. [Google Scholar]
- Zhou, S.J. Effects of nitric oxide fumigation on phenolic metabolism of postharvest Chinese winter jujube (Zizyphus jujuba Mill. cv. Dongzao) in relation to fruit quality. LWT—Food Sci. Technol. 2009, 42, 1009–1014. [Google Scholar] [CrossRef]
- Dong, J.; Ming, Z.; Lu, L.; Sun, U.; Xu, M. Nitric oxide fumigation stimulates flavonoid and phenolic accumulation and enhances antioxidant activity of mushroom. Food Chem. 2012, 135, 1220–1225. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| PpaSOD/SOD | PpaCAT/CAT | PpaPOD/POD | PpaPOD-1/POD | PpaAPX/APX | PpaPAL/PAL | |
|---|---|---|---|---|---|---|
| Pearson correlation | −0.053 | −0.533 ** | 0.112 | −0.406 | 0.016 | 0.588 ** |
| Peak | Proposed Compounds | Catagory | RT (min) | Neutral Mass (Da) | a [M+H]/b [M-H] (m/z) | Mass Error (Ppm) | Formula | Fragment Number | Relative Amount of Phenolic Compounds c |
|---|---|---|---|---|---|---|---|---|---|
| 1 | Pigment A | Anthocyanins | 4.09 | 609.1615 | 610.1688 a | 0.1 | C31H29O13 | 22 |  |
| 2 | Peonidin 3-O-(6″-p-coumaroyl-glucoside) | Anthocyanins | 4.82 | 609.1622 | 610.1695 a | −0.2 | C31H29O13 | 37 |  |
| 3 | Cyanidin 3-O-xylosyl-rutinoside | Anthocyanins | 9.64 | 727.2090 | 728.2163 a | 0.5 | C32H39O19 | 7 |  |
| 4 | Pelargonidin 3-O-rutinoside | Anthocyanins | 9.70 | 579.1731 | 580.1804 a | 1.2 | C27H31O14 | 6 |  |
| 5 | Naringenin 4′-O-glucuronide | Flavanones | 3.94 | 448.1003 | 449.1076 a | 0 | C21H20O11 | 27 |  |
| 6 | Poncirin | Flavanones | 9.57 | 594.1942 | 595.2014 a | −0.3 | C28H34O14 | 19 |  |
| 7 | (+)-Gallocatechin | Flavanols | 4.23 | 306.0725 | 307.0797 a | 0.1 | C15H14O7 | 6 |  |
| 8 | (+)-Catechin | Flavanols | 3.46 | 290.0788 | 291.0861 a | −0.2 | C15H14O6 | 3 |  |
| 9 | Luteolin 7-O-glucuronide | Flavones | 0.92 | 462.0811 | 463.0883 a | 1.4 | C21H18O12 | 17 |  |
| 10 | Kaempferol | Flavones | 3.94 | 286.0479 | 287.0738 a | 0 | C15H10O6 | 5 |  |
| 11 | Kaempferol 3-O-galactoside | Flavones | 3.95 | 448.1006 | 449.1079 a | 0 | C21H20O11 | 23 |  |
| 12 | Apigenin 7-O-glucoside | Flavones | 4.39 | 432.1066 | 433.1139 a | 0.1 | C21H20O10 | 17 |  |
| 13 | Luteolin 7-O-(2-apiosyl-6-malonyl)-glucoside | Flavones | 9.48 | 666.1414 | 667.1486 a | −2.8 | C29H30O18 | 22 |  |
| 14 | 5,3′,4′-Trihydroxy-3-methoxy-6:7-methylenedioxyflavone 4′-O-glucuronide | Flavonols | 1.32 | 520.0844 | 521.0917 a | 0.2 | C23H20O14 | 9 |  |
| 15 | Isorhamnetin 3-O-glucoside 7-O-rhamnoside | Flavonols | 4.84 | 624.1701 | 625.1774 a | 1.2 | C28H32O16 | 40 |  |
| 16 | Gallic acid | Phenolic acids | 2.50 | 170.0210 | 169.0156 b | 0.1 | C7H6O5 | 2 |  |
| 17 | 4-Hydroxybenzoic acid 4-O-glucoside | Phenolic acids | 3.38 | 323.0734 | 299.0841 b | −0.2 | C13H16O8 | 10 |  |
| 18 | 3-Caffeoylquinic acid | Phenolic acids | 3.55 | 354.0945 | 353.1012 b | −0.6 | C16H18O9 | 8 |  |
| 19 | 3-Feruloylquinic acid | Phenolic acids | 4.36 | 368.1102 | 367.1101 b | −0.4 | C17H20O9 | 13 |  |
| 20 | Caffeoyl glucose | Phenolic acids | 7.44 | 342.0935 | 341.0570 b | 1 | C15H18O9 | 7 |  |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Yuan, J.; Wang, X.; Yu, M.; Ma, R.; Yu, Z. Synergy of Nitric Oxide and 1-Methylcyclopropene Treatment in Prolong Ripening and Senescence of Peach Fruit. Foods 2021, 10, 2956. https://doi.org/10.3390/foods10122956
Wu X, Yuan J, Wang X, Yu M, Ma R, Yu Z. Synergy of Nitric Oxide and 1-Methylcyclopropene Treatment in Prolong Ripening and Senescence of Peach Fruit. Foods. 2021; 10(12):2956. https://doi.org/10.3390/foods10122956
Chicago/Turabian StyleWu, Xiaoqin, Jiawei Yuan, Xiaoqing Wang, Mingliang Yu, Ruijuan Ma, and Zhifang Yu. 2021. "Synergy of Nitric Oxide and 1-Methylcyclopropene Treatment in Prolong Ripening and Senescence of Peach Fruit" Foods 10, no. 12: 2956. https://doi.org/10.3390/foods10122956
APA StyleWu, X., Yuan, J., Wang, X., Yu, M., Ma, R., & Yu, Z. (2021). Synergy of Nitric Oxide and 1-Methylcyclopropene Treatment in Prolong Ripening and Senescence of Peach Fruit. Foods, 10(12), 2956. https://doi.org/10.3390/foods10122956