
Control of Foodborne Staphylococcus aureus by Shikonin, a Natural Extract

Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Strains
2.3. MICs and MBCs of Shikonin against S. aureus
2.4. NICs of Shikonin against S. aureus
2.5. The Effect of Shikonin on the Membrane Potential of S. aureus
2.6. The Effect of Shikonin on the Intracellular ATP Concentration of S. aureus
2.7. The Effect of Shikonin on the Membrane Integrity of S. aureus
2.8. The Effect of Shikonin on the Cell Morphology of S. aureus
2.9. The Effect of Shikonin on the Biofilm of S. aureus
2.10. RT-qPCR of S. aureus Biofilm Formation Key Genes (icaA, cidA, agrA, sarA) and Virulence Genes (Hla, Sea)
2.11. Colony Count
2.12. Hemolytic Ability
2.13. Western Blotting
2.14. Thermal Stability and Safety Assessment of Shikonin
2.15. Application of Shikonin in Food Model
2.16. Statistical Analysis
3. Result
3.1. MICs and MBCs
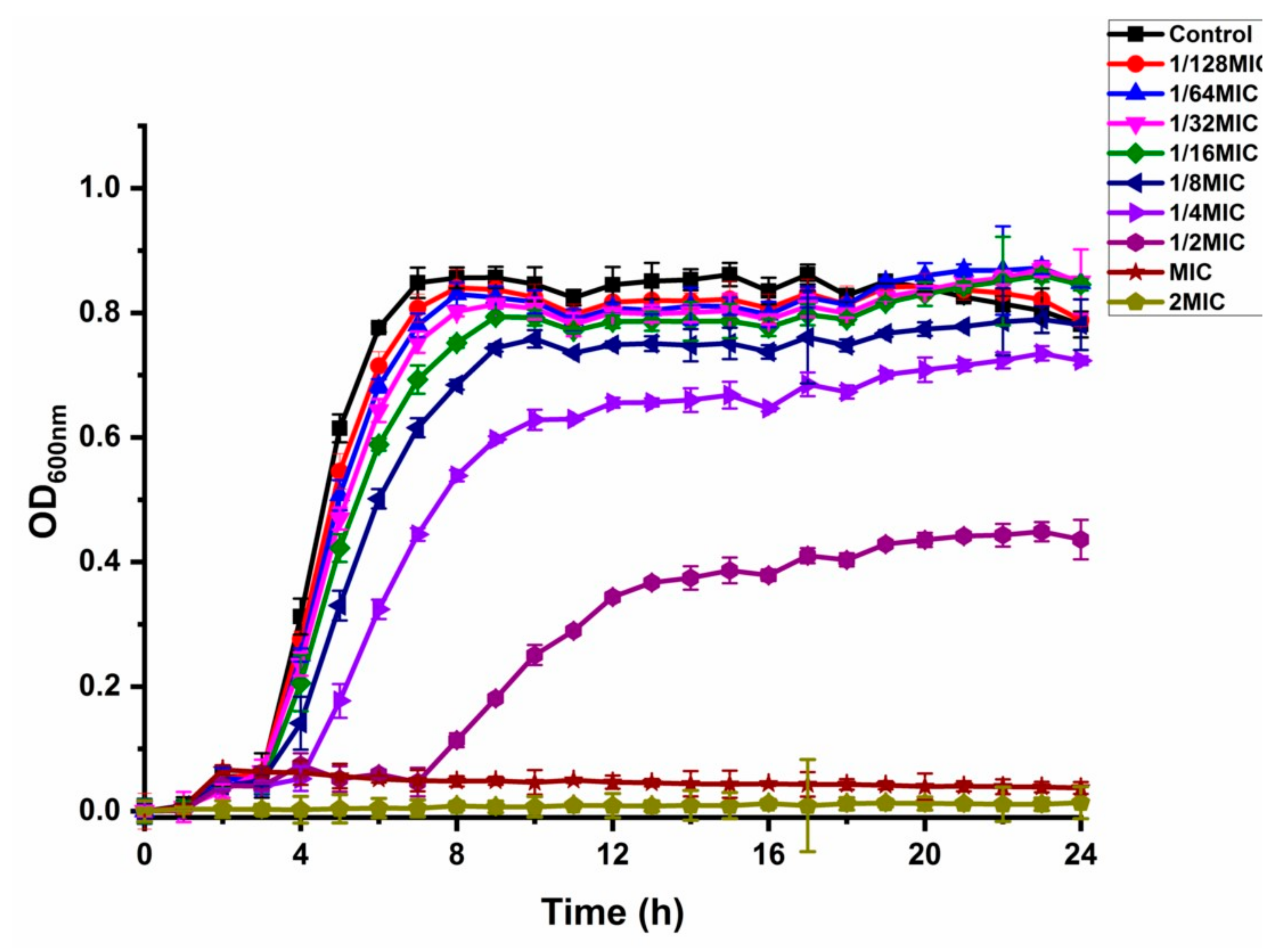
3.2. Growth Curve and NICs
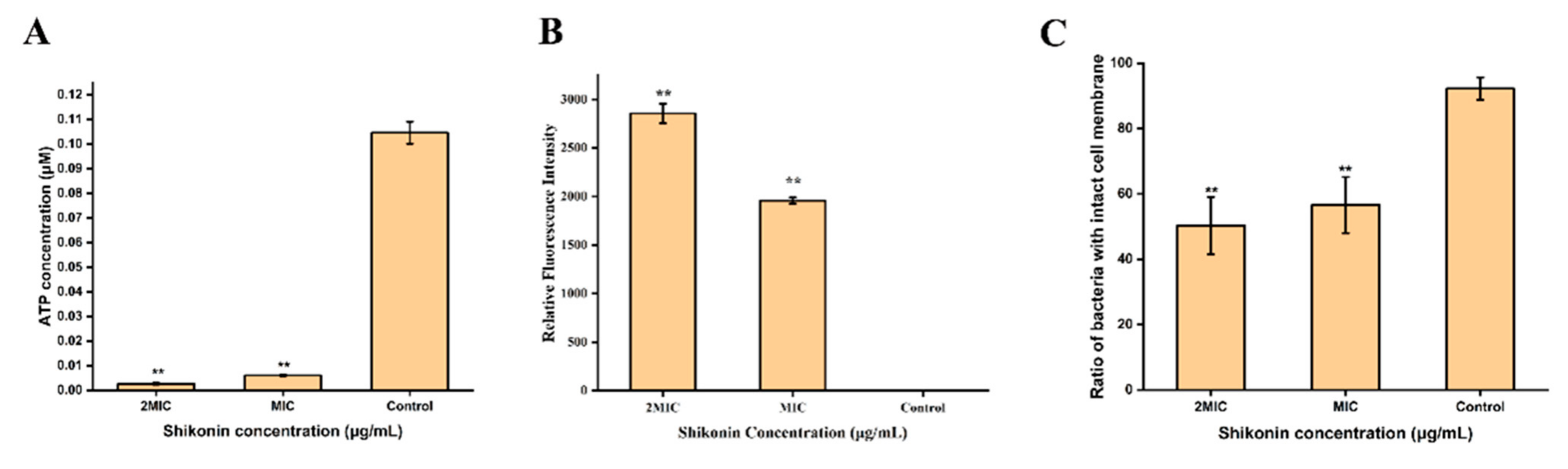
3.3. The Intracellular ATP Concentration of S. aureus
3.4. The Cell Membrane Potential of S. aureus
3.5. The Integrity of S. aureus Cell Membrane
3.6. The Cell Morphology of S. aureus
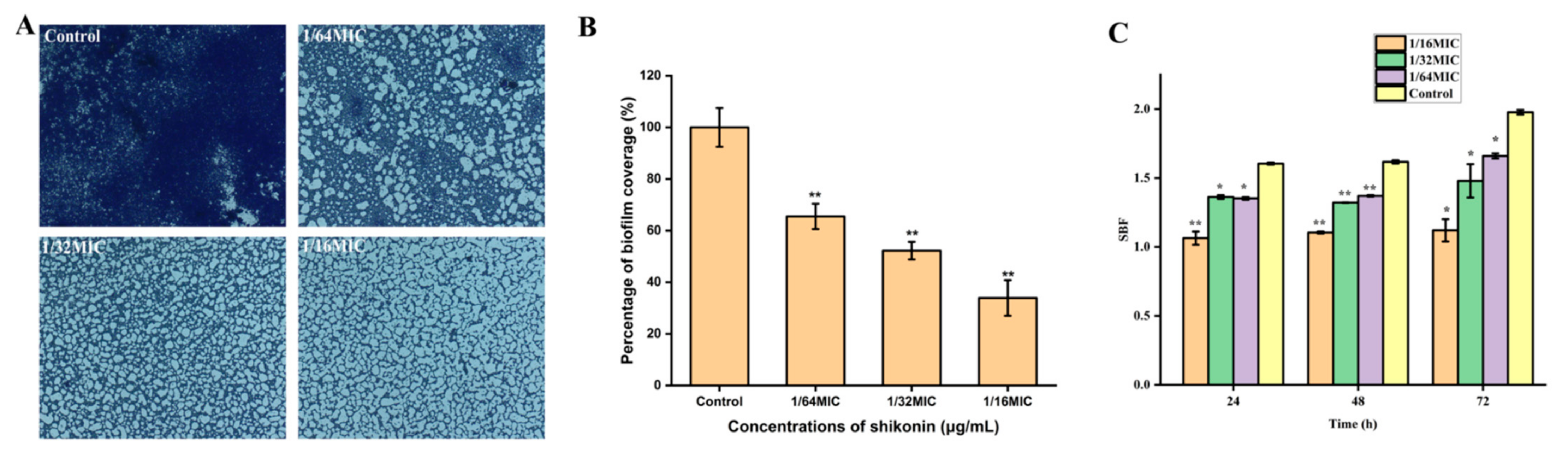
3.7. The Biofilm Formation of S. aureus
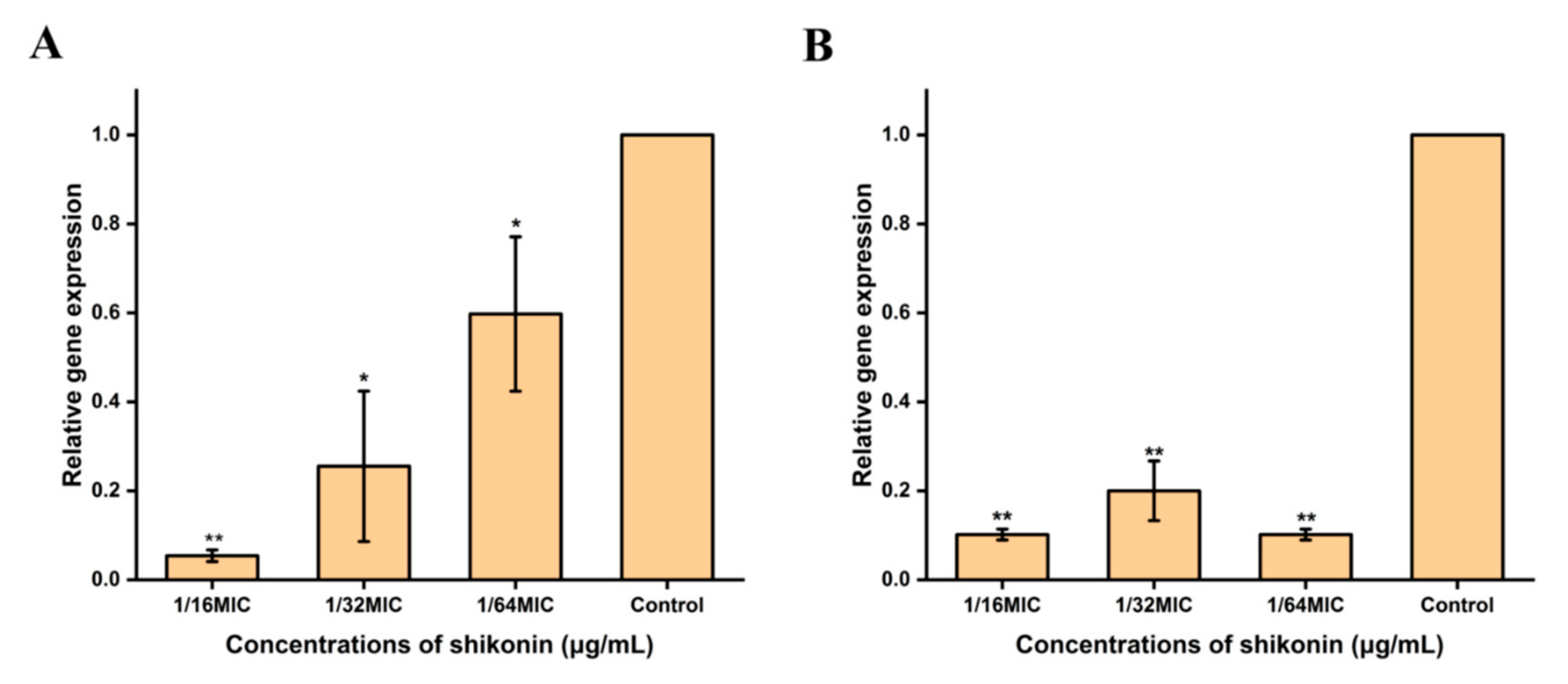
3.8. Shikonin Inhibited the Transcription of Cida and sarA Genes
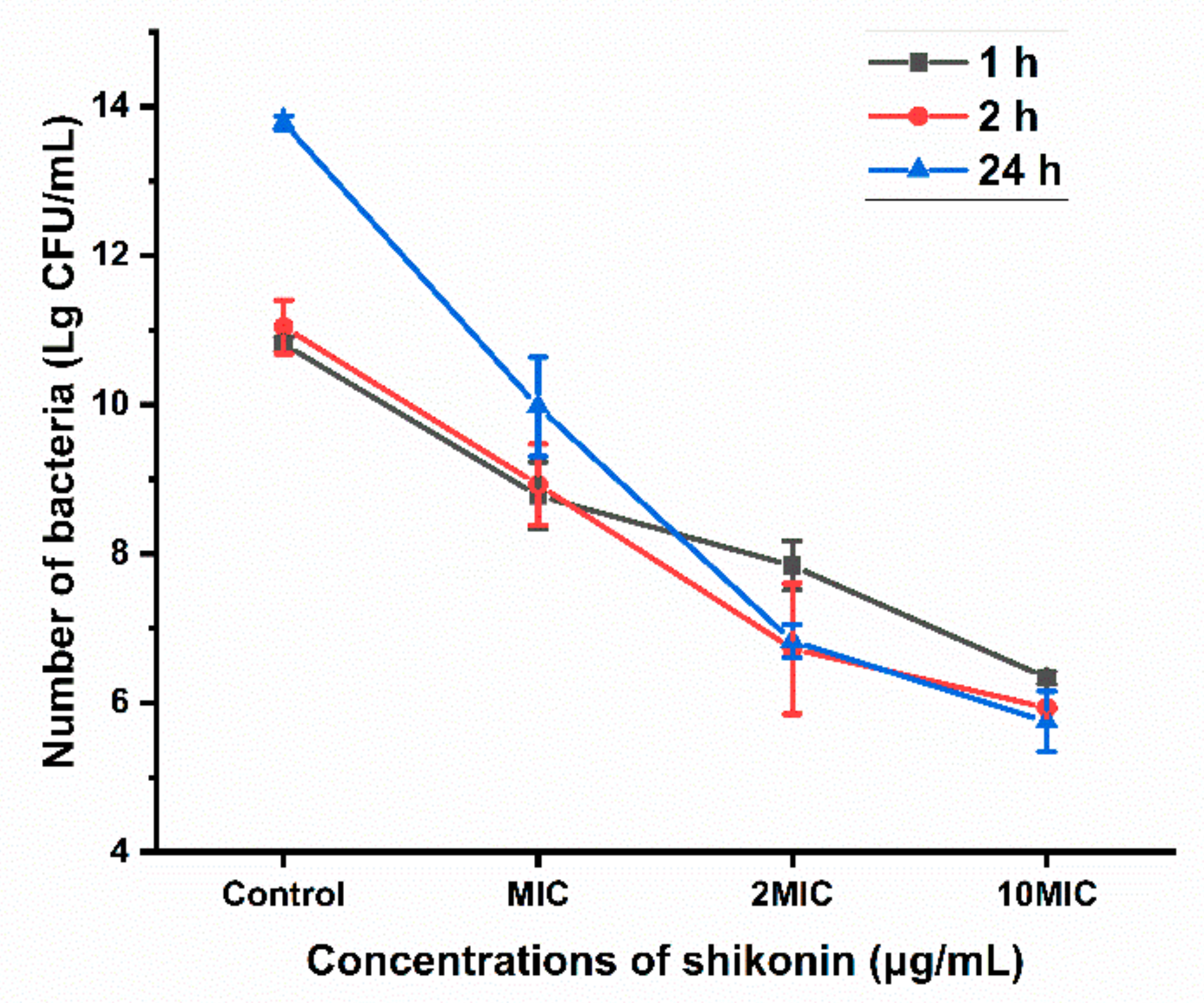
3.9. Antibacterial Activity of Shikonin on S. aureus Bacteria in Biofilm
3.10. Shikonin Inhibited the Hemolytic Ability of S. aureus
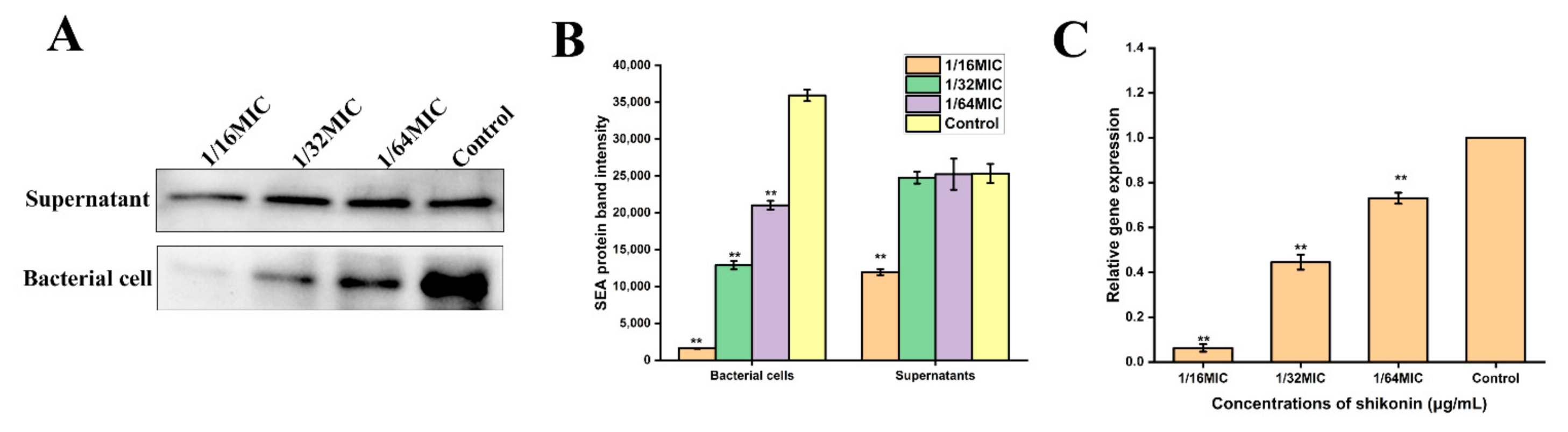
3.11. Shikonin Inhibited Sea Gene Transcription and Expression
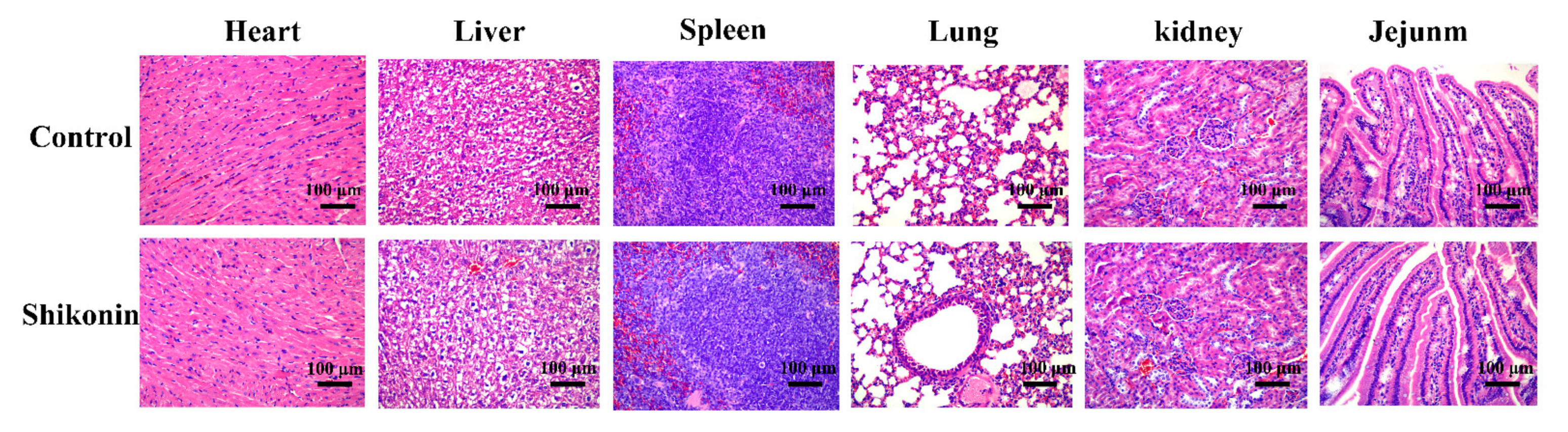
3.12. The Thermal Stability and the Safety of Shikonin
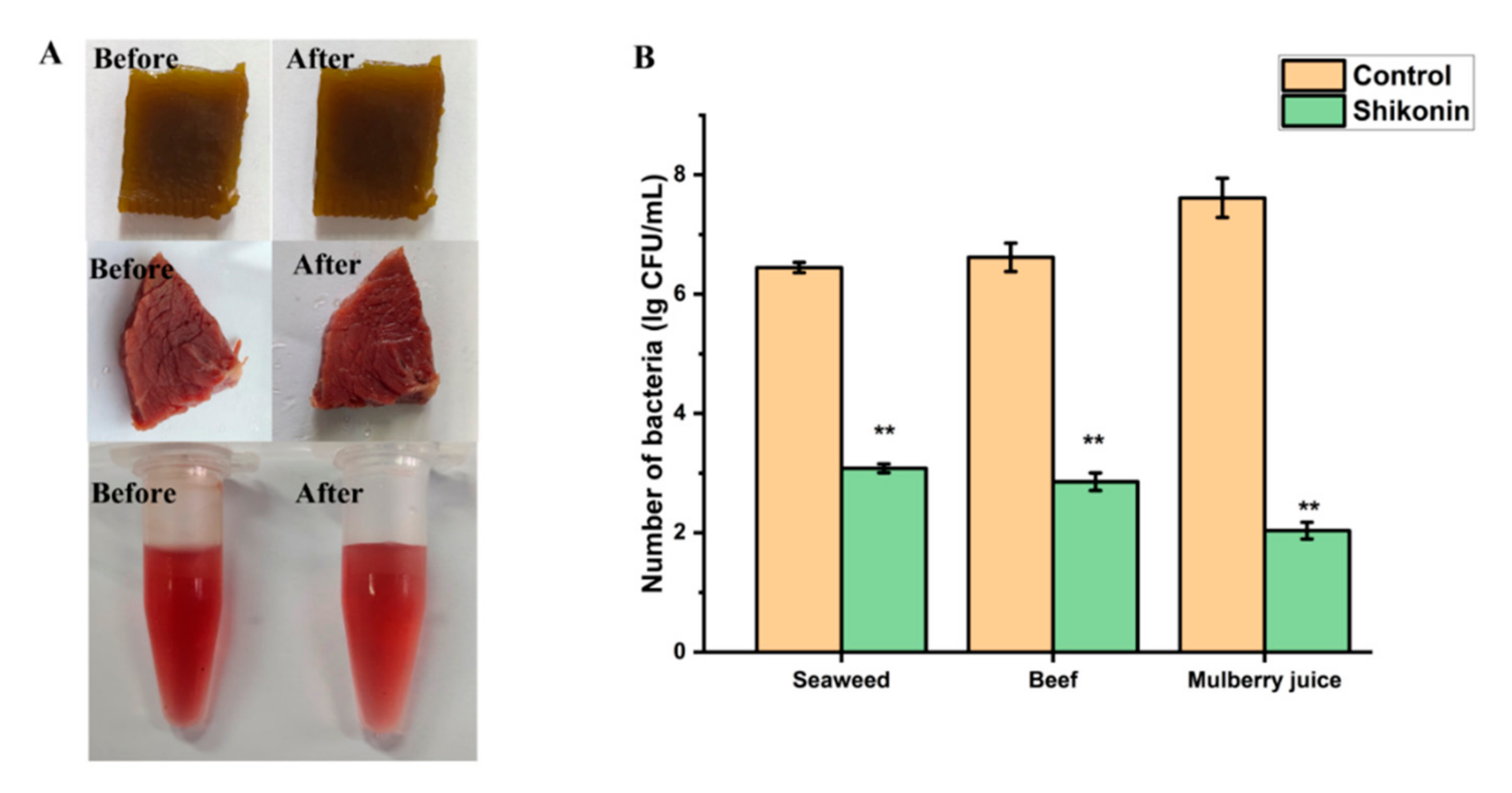
3.13. Application of Shikonin in Food Model
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Argudín, M.Á.; Mendoza, M.C.; Rodicio, M.R. Food Poisoning and Staphylococcus aureus Enterotoxins. Toxins 2010, 2, 1751–1773. [Google Scholar] [CrossRef] [PubMed]
- Wardenburg, J.B.; Bae, T.; Otto, M.; DeLeo, F.; Schneewind, O. Poring over pores: α-hemolysin and Panton-Valentine leukocidin in Staphylococcus aureus pneumonia. Nat. Med. 2007, 13, 1405–1406. [Google Scholar] [CrossRef]
- Omoe, K.; Ishikawa, M.; Shimoda, Y.; Hu, D.-L.; Ueda, S.; Shinagawa, K. Detection of seg, seh, and sei genes in Staphylococcus aureus Isolates and Determination of the Enterotoxin Productivities of S. aureus Isolates Harboring seg, seh, or sei Genes. J. Clin. Microbiol. 2002, 40, 857–862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, Y.; Ono, H.K.; Shimojima, Y.; Kubota, H.; Kato, R.; Kakuda, T.; Hirose, S.; Hu, D.-L.; Nakane, A.; Takai, S.; et al. A novel staphylococcal enterotoxin SE02 involved in a staphylococcal food poisoning outbreak that occurred in Tokyo in 2004. Food Microbiol. 2020, 92, 103588. [Google Scholar] [CrossRef] [PubMed]
- Pinchuk, I.V.; Beswick, E.J.; Reyes, V.E. Staphylococcal Enterotoxins. Toxins 2010, 2, 2177–2197. [Google Scholar] [CrossRef] [Green Version]
- Hall-Stoodley, L.; Costerton, J.W.; Stoodley, P. Bacterial biofilms: From the Natural environment to infectious diseases. Nat. Rev. Genet. 2004, 2, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Archer, N.; Mazaitis, M.J.; Costerton, J.W.; Leid, J.G.; Powers, M.E.; Shirtliff, M.E. Staphylococcus aureus biofilms: Properties, regulation, and roles in human disease. Virulence 2011, 2, 445–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Assimopoulu, A.N.; Boskou, D.; Papageorgiou, V.P. Antioxidant activities of alkannin, shikonin and Alkanna tinctoria root extracts in oil substrates. Food Chem. 2004, 87, 433–438. [Google Scholar] [CrossRef]
- Ezati, P.; Bang, Y.-J.; Rhim, J.-W. Preparation of a shikonin-based pH-sensitive color indicator for monitoring the freshness of fish and pork. Food Chem. 2021, 337, 127995. [Google Scholar] [CrossRef] [PubMed]
- Han, C.-T.; Kim, M.-J.; Moon, S.-H.; Jeon, Y.-R.; Hwang, J.-S.; Nam, C.; Park, C.-W.; Lee, S.-H.; Na, J.-B.; Park, C.-S.; et al. Acute and 28-Day Subacute Toxicity Studies of Hexane Extracts of the Roots of Lithospermum erythrorhizon in Sprague-Dawley Rats. Toxicol. Res. 2015, 31, 403–414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, L.; Liu, L.; Wang, Y.; Yan, G.; Zhang, Y. Long-term systemic toxicity of shikonin derivatives in Wistar rats. Pharm. Biol. 2013, 52, 486–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhou, S.; Yu, J.; Cai, W.; Yang, Y.; Kuang, X.; Liu, H.; He, Z.; Wang, Y. Low dose shikonin and anthracyclines coloaded liposomes induce robust immunogenetic cell death for synergistic chemo-immunotherapy. J. Control. Release 2021, 335, 306–319. [Google Scholar] [CrossRef]
- Yuan, D.-P.; Gu, L.; Long, J.; Chen, J.; Ni, J.; Qian, N.; Shi, Y.-L. Shikonin reduces endometriosis by inhibiting RANTES secretion and mononuclear macrophage chemotaxis. Exp. Ther. Med. 2014, 7, 685–690. [Google Scholar] [CrossRef] [Green Version]
- Vinaik, R.; Barayan, D.; Auger, C.; Abdullahi, A.; Jeschke, M.G. Regulation of glycolysis and the Warburg effect in wound healing. JCI Insight 2020, 5. [Google Scholar] [CrossRef]
- Vegara, S.; Funes, L.; Martí, N.; Saura, D.; Micol, V.; Valero, M. Bactericidal activities against pathogenic bacteria by selected constituents of plant extracts in carrot broth. Food Chem. 2011, 128, 872–877. [Google Scholar] [CrossRef]
- Zhang, Y.; Han, H.; Sun, L.; Qiu, H.; Lin, H.; Yu, L.; Zhu, W.; Qi, J.; Yang, R.; Pang, Y.; et al. Antiviral activity of shikonin ester derivative PMM-034 against enterovirus 71 in vitro. Braz. J. Med Biol. Res. 2017, 50, 6586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, J.-R.; Zhong, K.; Wu, Y.-P.; Elena, G.; Gao, H. Antibiofilm activity of shikimic acid against Staphylococcus aureus. Food Control 2019, 95, 327–333. [Google Scholar] [CrossRef]
- Guo, D.; Wang, S.; Li, J.; Bai, F.; Yang, Y.; Xu, Y.; Liang, S.; Xia, X.; Wang, X.; Shi, C. The antimicrobial activity of coenzyme Q against planktonic and biofilm forms of Cronobacter sakazakii. Food Microbiol. 2020, 86, 103337. [Google Scholar] [CrossRef]
- Shi, C.; Sun, Y.; Liu, Z.; Guo, D.; Sun, H.; Sun, Z.; Chen, S.; Zhang, W.; Wen, Q.; Peng, X.; et al. Inhibition of Cronobacter sakazakii Virulence Factors by Citral. Sci. Rep. 2017, 7, 43243. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Xu, Y.; Dong, M.; Hang, B.; Sun, Y.; Wang, L.; Wang, Y.; Hu, J.; Zhang, W. HJH-1, a Broad-Spectrum Antimicrobial Activity and Low Cytotoxicity Antimicrobial Peptide. Molecules 2018, 23, 2026. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Li, S.; Li, H.; Guo, X.; Guo, D.; Yang, Y.; Wang, X.; Zhang, C.; Shan, Z.; Xia, X.; et al. Antibiofilm activity of shikonin against Listeria monocytogenes and inhibition of key virulence factors. Food Control 2021, 120, 107558. [Google Scholar] [CrossRef]
- Amalaradjou, M.A.R.; Venkitanarayanan, K. Effect of trans-Cinnamaldehyde on Inhibition and Inactivation of Cronobacter sakazakii Biofilm on Abiotic Surfaces. J. Food Prot. 2011, 74, 200–208. [Google Scholar] [CrossRef]
- Costa, E.; Silva, S.; Tavaria, F.; Pintado, M.M. Insights into chitosan antibiofilm activity against methicillin-resistant Staphylococcus aureus. J. Appl. Microbiol. 2017, 122, 1547–1557. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Meng, R.; Zhao, X.; Shi, C.; Zhang, X.; Zhang, Y.; Guo, N. Inhibition effect of tea tree oil on Listeria monocytogenes growth and exotoxin proteins listeriolysin O and p60 secretion. Lett. Appl. Microbiol. 2016, 63, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Teng, Z.; Shi, D.; Liu, H.; Shen, Z.; Zha, Y.; Li, W.; Deng, X.; Wang, J. Lysionotin attenuates Staphylococcus aureus pathogenicity by inhibiting α-toxin expression. Appl. Microbiol. Biotechnol. 2017, 101, 6697–6703. [Google Scholar] [CrossRef] [PubMed]
- National Standard of the People’s Republic of China. National Food Safety Standard Food Microbiological Examination: Staphylococcus Aureus; GB 4789.10-2016; Standard Press of China: Beijing, China, 2016. [Google Scholar]
- Hennekinne, J.-A.; De Buyser, M.-L.; Dragacci, S. Staphylococcus aureus and its food poisoning toxins: Characterization and outbreak investigation. FEMS Microbiol. Rev. 2012, 36, 815–836. [Google Scholar] [CrossRef] [Green Version]
- Qiu, J.; Wang, D.; Xiang, H.; Feng, H.; Jiang, Y.; Xia, L.; Dong, J.; Lu, J.; Yu, L.; Deng, X. Subinhibitory Concentrations of Thymol Reduce Enterotoxins A and B and α-Hemolysin Production in Staphylococcus aureus Isolates. PLoS ONE 2010, 5, e9736. [Google Scholar] [CrossRef] [Green Version]
- Liao, P.-L.; Lin, C.-H.; Li, C.-H.; Tsai, C.-H.; Ho, J.-D.; Chiou, G.C.Y.; Kang, J.-J.; Cheng, Y.-W. Anti-inflammatory properties of shikonin contribute to improved early-stage diabetic retinopathy. Sci. Rep. 2017, 7, 44985. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; He, C.; Lu, S.; Wang, X.; Wang, C.; Wang, L.; Zhang, J.; Piao, M.; Chi, G.; Luo, Y.; et al. MLKL contributes to shikonin-induced glioma cell necroptosis via promotion of chromatinolysis. Cancer Lett. 2019, 467, 58–71. [Google Scholar] [CrossRef]
- Lee, Y.-S.; Lee, D.-Y.; Kim, Y.B.; Lee, S.-W.; Cha, S.-W.; Park, H.-W.; Kim, G.-S.; Kwon, D.-Y.; Lee, M.-H.; Han, S.-H. The Mechanism Underlying the Antibacterial Activity of Shikonin against Methicillin-Resistant Staphylococcus aureus. Evid.-Based Complement. Altern. Med. 2015, 2015, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Shi, C.; Wu, Q.; Zheng, Z.; Liu, P.; Li, G.; Peng, X.; Xia, X. Antimicrobial Activity of Punicalagin Against Staphylococcus aureus and Its Effect on Biofilm Formation. Foodborne Pathog. Dis. 2017, 14, 282–287. [Google Scholar] [CrossRef]
- Li, G.; Qiao, M.; Guo, Y.; Wang, X.; Xu, Y.; Xia, X. Effect of Subinhibitory Concentrations of Chlorogenic Acid on Reducing the Virulence Factor Production by Staphylococcus aureus. Foodborne Pathog. Dis. 2014, 11, 677–683. [Google Scholar] [CrossRef]
- Duan, J.; Li, M.; Hao, Z.; Shen, X.; Liu, L.; Jin, Y.; Wang, S.; Guo, Y.; Yang, L.; Wang, L.; et al. Subinhibitory concentrations of resveratrol reduce alpha-hemolysin production in Staphylococcus aureus isolates by downregulating saeRS. Emerg. Microbes Infect. 2018, 7, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleem, M.; Nazir, M.; Ali, M.S.; Hussain, H.; Lee, Y.S.; Riaz, N.; Jabbar, A. Antimicrobial natural products: An update on future antibioticdrug candidates. Nat. Prod. Rep. 2010, 27, 238–254. [Google Scholar] [CrossRef] [PubMed]
- Steinmann, M.E.; González-Salgado, A.; Bütikofer, P.; Mäser, P.; Sigel, E. A heteromeric potassium channel involved in the modulation of the plasma membrane potential is essential for the survival of African trypanosomes. FASEB J. 2015, 29, 3228–3237. [Google Scholar] [CrossRef] [Green Version]
- Yun, D.G.; Lee, D.G. Silymarin exerts antifungal effects via membrane-targeted mode of action by increasing permeability and inducing oxidative stress. Biochim. Biophys. Acta (BBA) Biomembr. 2017, 1859, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Ji, Y.R.; Ryoo, Z.Y.; Choi, M.-S.; Woo, E.-R.; Lee, D.G. Antibacterial Mechanism of (−)-Nortrachelogenin in Escherichia coli O157. Curr. Microbiol. 2015, 72, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Zhang, X.; Sun, Y.; Yang, M.; Song, K.; Zheng, Z.; Chen, Y.; Liu, X.; Jia, Z.; Dong, R.; et al. Antimicrobial Activity of Ferulic Acid Against Cronobacter sakazakiiand Possible Mechanism of Action. Foodborne Pathog. Dis. 2016, 13, 196–204. [Google Scholar] [CrossRef]
- Song, J.; Chen, H.; Wei, Y.; Liu, J. Synthesis of carboxymethylated β-glucan from naked barley bran and its antibacterial activity and mechanism against Staphylococcus aureus. Carbohydr. Polym. 2020, 242, 116418. [Google Scholar] [CrossRef]
- Bajpai, V.K.; Sharma, A.; Baek, K.-H. Antibacterial mode of action of seed essential oil of Eleutherococcus senticosusagainst foodborne pathogens. Int. J. Food Sci. Technol. 2013, 48, 2300–2305. [Google Scholar] [CrossRef]
- Mempin, R.; Tran, H.; Chen, C.; Gong, H.; Ho, K.K.; Lu, S. Release of extracellular ATP by bacteria during growth. BMC Microbiol. 2013, 13, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.-H.; Zhou, T.-T.; Wei, C.-H.; Lan, W.-Q.; Zhao, Y.; Pan, Y.-J.; Wu, V.C. Antibacterial effect and mechanism of anthocyanin rich Chinese wild blueberry extract on various foodborne pathogens. Food Control 2018, 94, 155–161. [Google Scholar] [CrossRef]
- Jia, M.; Mai, B.; Liu, S.; Li, Z.; Liu, Q.; Wang, P. Antibacterial effect of S-Porphin sodium photodynamic therapy on Staphylococcus aureus and multiple drug resistance Staphylococcus aureus. Photodiagnosis Photodyn. Ther. 2019, 28, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, A.; Wu, V.C.; White, J.; Tadepalli, S.; Andre, E.E. The antimicrobial properties of the lowbush blueberry (Vaccinium angustifolium) fractional components against foodborne pathogens and the conservation of probiotic Lactobacillus rhamnosus. Food Microbiol. 2012, 30, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Bajpai, V.K.; Sharma, A.; Baek, K.-H. Antibacterial mode of action of Cudrania tricuspidata fruit essential oil, affecting membrane permeability and surface characteristics of food-borne pathogens. Food Control 2013, 32, 582–590. [Google Scholar] [CrossRef]
- Lal, S.; Pearce, M.; Achilles-Day, U.E.M.; Day, J.G.; Morton, L.H.G.; Crean, S.J.; Singhrao, S.K. Developing an ecologically relevant heterogeneous biofilm model for dental-unit waterlines. Biofouling 2017, 33, 75–87. [Google Scholar] [CrossRef] [Green Version]
- Oxaran, V.; Dittmann, K.K.; Lee, S.H.I.; Chaul, L.T.; de Oliveira, C.A.F.; Corassin, C.H.; Alves, V.F.; De Martinis, E.C.P.; Gram, L. Behavior of Foodborne Pathogens Listeria monocytogenes and Staphylococcus aureus in Mixed-Species Biofilms Exposed to Biocides. Appl. Environ. Microbiol. 2018, 84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Shen, X.; Yu, J.; Cao, X.; Zhan, Q.; Guo, Y.; Yu, F. Subinhibitory Concentrations of Fusidic Acid May Reduce the Virulence of S. aureus by Down-Regulating sarA and saeRS to Reduce Biofilm Formation and α-Toxin Expression. Front. Microbiol. 2020, 11, 25. [Google Scholar] [CrossRef]
- Beenken, K.E.; Dunman, P.M.; McAleese, F.; Macapagal, D.; Murphy, E.; Projan, S.J.; Blevins, J.S.; Smeltzer, M.S. Global Gene Expression in Staphylococcus aureus Biofilms. J. Bacteriol. 2004, 186, 4665–4684. [Google Scholar] [CrossRef] [Green Version]
- Valle, J.; Toledo-Arana, A.; Berasain, C.; Ghigo, J.-M.; Amorena, B.; Penadés, J.R.; Lasa, I. SarA and not σB is essential for biofilm development by Staphylococcus aureus. Mol. Microbiol. 2003, 48, 1075–1087. [Google Scholar] [CrossRef]
- Nygaard, T.; Pallister, K.B.; Dumont, A.L.; Dewald, M.; Watkins, R.L.; Pallister, E.Q.; Malone, C.; Griffith, S.; Horswill, A.R.; Torres, V.; et al. Alpha-Toxin Induces Programmed Cell Death of Human T cells, B cells, and Monocytes during USA300 Infection. PLoS ONE 2012, 7, e36532. [Google Scholar] [CrossRef] [PubMed]
- Shallcross, L.; Williams, K.; Hopkins, S.; Aldridge, R.; Johnson, A.; Hayward, A. Panton-Valentine leukocidin associated staphylococcal disease: A cross-sectional study at a London hospital, England. Clin. Microbiol. Infect. 2010, 16, 1644–1648. [Google Scholar] [CrossRef] [PubMed]
- Xiang, H.; Qiu, J.-Z.; Wang, D.-C.; Jiang, Y.-S.; Xia, L.-J.; Deng, X.-M. Influence of Magnolol on the Secretion of α-Toxin by Staphylococcus aureus. Molecules 2010, 15, 1679–1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Dong, J.; Qiu, J.-Z.; Wang, J.-F.; Luo, M.-J.; Li, H.-E.; Leng, B.-F.; Ren, W.-Z.; Deng, X.-M. Peppermint Oil Decreases the Production of Virulence-Associated Exoproteins by Staphylococcus aureus. Molecules 2011, 16, 1642–1654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Standard of the People’s Republic of China. National Food Safety Standard: Standards for Uses of Food Additives; GB 2760-2014; Standard Press of China: Beijing, China, 2014. [Google Scholar]
- Ordoudi, S.A.; Tsermentseli, S.K.; Nenadis, N.; Assimopoulou, A.N.; Tsimidou, M.Z.; Papageorgiou, V.P. Structure-radical scavenging activity relationship of alkannin/shikonin derivatives. Food Chem. 2011, 124, 171–176. [Google Scholar] [CrossRef]
- Vukić, M.; Vukovic, N.L.; Obradović, A.; Popovic, S.; Zaric, M.; Djurdjevic, P.; Marković, S.; Baskic, D.D. Naphthoquinone rich Onosma visianii Clem (Boraginaceae) root extracts induce apoptosis and cell cycle arrest in HCT-116 and MDA-MB-231 cancer cell lines. Nat. Prod. Res. 2017, 32, 2712–2716. [Google Scholar] [CrossRef] [PubMed]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence (5′-3′) |
|---|---|
| icaA-F | GATACTGATATGATTACCGAAGAT |
| icaA-R | GAACCAACATCCAACACAT |
| cidA-F | ATTCATAAGCGTCTACACCTT |
| cidA-R | TTCTTCATACCGTCAGTTGT |
| agrA-F | TGAAATTCGTAAGCATGACCC |
| agrA-R | CATCGCTGCAACTTTGTAGAC |
| sarA-F | TGTTTGCTTCAGTGATTCGTTTA |
| sarA-R | AACCACAAGTTGTTAAAGCAGTTA |
| 16SrRNA-F | CGTGCTACAATGGACAATACAAA |
| 16SrRNA-R | ATCTACGATTACTAGCGATTCCA |
| hla-F | TTGGTGCAAATGTTTC |
| hla-R | TCACTTTCCAGCCTACT |
| sea-F | ATGGTGCTTATTATGGTTATC |
| sea-R | CGTTTCCAAAGGTACTGTATT |
| Strain | MIC (μg/mL) | MBC (μg/mL) | Original Source of Strain |
|---|---|---|---|
| ATCC29213 | 35 | 35 | American Type Culture Collection |
| ATCC25923 | 35 | 35 | American Type Culture Collection |
| A48 | 35 | 35 | Patient anal swab |
| A86 | 35 | 35 | Patient anal swab |
| 13# | 70 | 70 | Grilled Chicken |
| 124 | 70 | 70 | Patient vomit |
| 265 | 70 | 70 | Patient vomit |
| S2 | 70 | 70 | Patient vomit |
| S4 | 70 | 70 | Patient vomit |
| Processing Conditions | MIC (μg/mL) | MBC (μg/mL) | |
|---|---|---|---|
| Temperature (°C) | Time (Min) | ||
| 100 | 1 | 35 | 35 |
| 100 | 3 | 35 | 35 |
| 100 | 5 | 35 | 35 |
| 121 | 0.5 | 35 | 35 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wan, Y.; Wang, X.; Zhang, P.; Zhang, M.; Kou, M.; Shi, C.; Peng, X.; Wang, X. Control of Foodborne Staphylococcus aureus by Shikonin, a Natural Extract. Foods 2021, 10, 2954. https://doi.org/10.3390/foods10122954
Wan Y, Wang X, Zhang P, Zhang M, Kou M, Shi C, Peng X, Wang X. Control of Foodborne Staphylococcus aureus by Shikonin, a Natural Extract. Foods. 2021; 10(12):2954. https://doi.org/10.3390/foods10122954
Chicago/Turabian StyleWan, Yangli, Xiaowen Wang, Pengfei Zhang, Meng Zhang, Mingying Kou, Chao Shi, Xiaoli Peng, and Xin Wang. 2021. "Control of Foodborne Staphylococcus aureus by Shikonin, a Natural Extract" Foods 10, no. 12: 2954. https://doi.org/10.3390/foods10122954
APA StyleWan, Y., Wang, X., Zhang, P., Zhang, M., Kou, M., Shi, C., Peng, X., & Wang, X. (2021). Control of Foodborne Staphylococcus aureus by Shikonin, a Natural Extract. Foods, 10(12), 2954. https://doi.org/10.3390/foods10122954