
Relationship between Structure and Biological Activity of Various Vitamin K Forms


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Biological Functions
2.1. Absorption
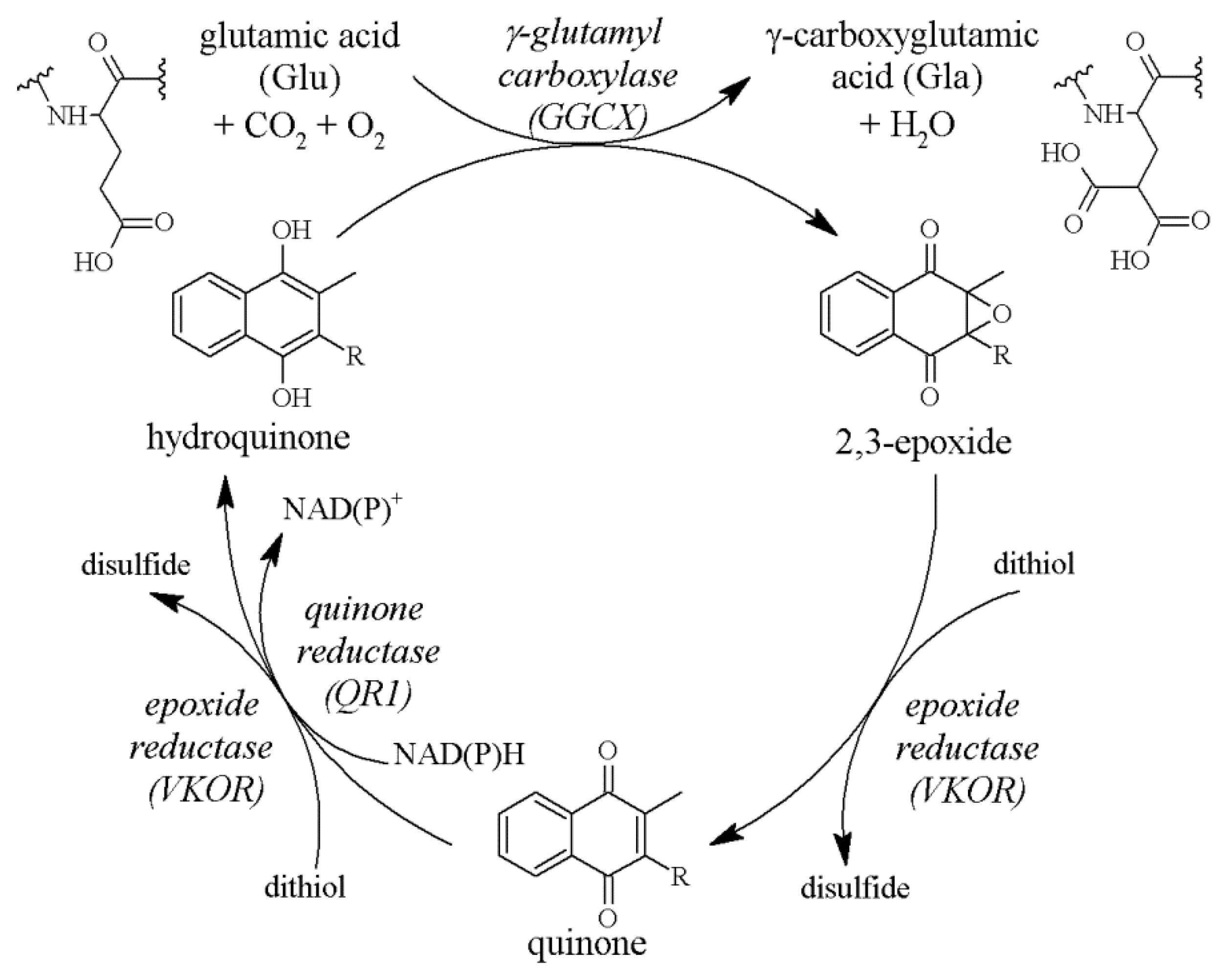
2.2. The Vitamin K Cycle
2.3. The Role of GGCX
2.4. VKDPs
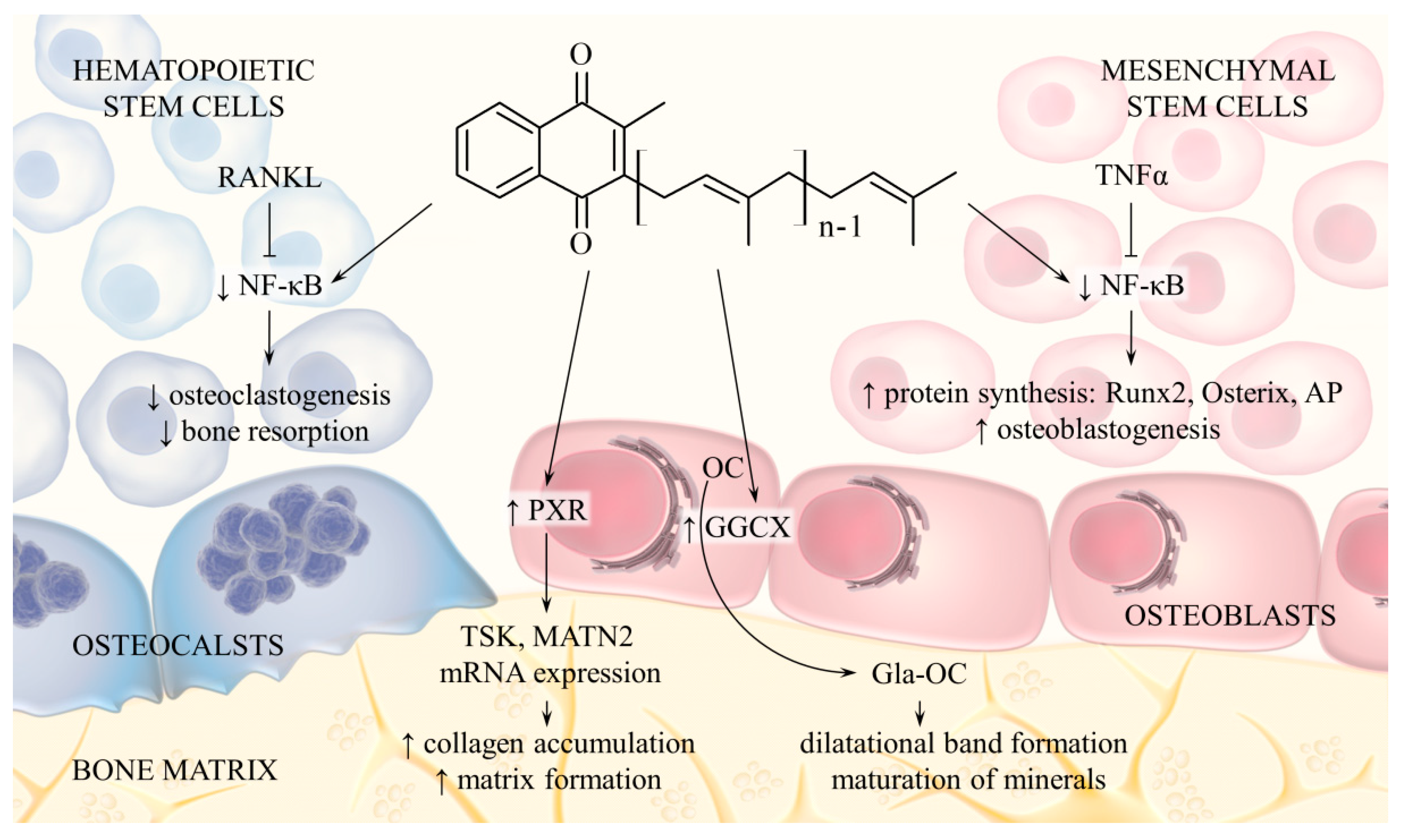
2.5. Effects on Bone Metabolism
2.6. Effects on Vascular Calcification
2.7. Effects on Cell Proliferation
2.8. Effects on Neural Cells
3. Daily Intake and Dietary Sources
4. Differences between Phylloquinone and Menaquinone Biological Activity
4.1. Absorption and Tissue Distribution
4.2. Relationship between Activity and Chain Length
4.3. Relationship between Activity and Double Bond Configuration
4.4. Effect on Blood Coagulation
4.5. Osteoporosis Prevention
4.6. Cardiovascular Disease Prevention
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hamidi, M.S.; Cheung, A.M. Vitamin K and Musculoskeletal Health in Postmenopausal Women. Mol. Nutr. Food Res. 2014, 58, 1647–1657. [Google Scholar] [CrossRef]
- Vetrella, M.; Barthelmai, W. Studies on Drug-Induced Hemolysis: Effects of Menadione and Its Water Soluble Preparations on the Glutathione Peroxidase of Human Erythrocytes. Klin. Wochenschr. 1972, 50, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Marles, R.J.; Roe, A.L.; Oketch-Rabah, H.A. US Pharmacopeial Convention Safety Evaluation of Menaquinone-7, a Form of Vitamin K. Nutr. Rev. 2017, 75, 553–578. [Google Scholar] [CrossRef]
- Di, W.; Khan, M.; Gao, Y.; Cui, J.; Wang, D.; Qu, M.; Feng, L.; Maryam, A.; Gao, H. Vitamin K4 Inhibits the Proliferation and Induces Apoptosis of U2OS Osteosarcoma Cells via Mitochondrial Dysfunction. Mol. Med. Rep. 2017, 15, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Yang, J.; Yang, C.; Meng, F.; Zhou, Y.; Yu, B.; Khan, M.; Yang, H. Vitamin K4 Induces Tumor Cytotoxicity in Human Prostate Carcinoma PC-3 Cells via the Mitochondria-Related Apoptotic Pathway. Pharmazie 2013, 68, 442–448. [Google Scholar] [PubMed]
- Chen, J.; Jiang, Z.; Wang, B.; Wang, Y.; Hu, X. Vitamin K3 and K5 Are Inhibitors of Tumor Pyruvate Kinase M2. Cancer Lett. 2012, 316, 204–210. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Li, Y.; Ahmad, J.; Wang, Y.; Scott, D.E.; Vostal, J.G. Vitamin K5 Is an Efficient Photosensitizer for Ultraviolet A Light Inactivation of Bacteria. FEMS Microbiol. Lett. 2018, 365, fny005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beulens, J.W.J.; Booth, S.L.; van den Heuvel, E.G.H.M.; Stoecklin, E.; Baka, A.; Vermeer, C. The Role of Menaquinones (Vitamin K2) in Human Health. Br. J. Nutr. 2013, 110, 1357–1368. [Google Scholar] [CrossRef] [Green Version]
- McCann, J.C.; Ames, B.N. Vitamin K, an Example of Triage Theory: Is Micronutrient Inadequacy Linked to Diseases of Aging? Am. J. Clin. Nutr. 2009, 90, 889–907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncalves, A.; Margier, M.; Roi, S.; Collet, X.; Niot, I.; Goupy, P.; Caris-Veyrat, C.; Reboul, E. Intestinal Scavenger Receptors Are Involved in Vitamin K1 Absorption*. J. Biol. Chem. 2014, 289, 30743–30752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takada, T.; Yamanashi, Y.; Konishi, K.; Yamamoto, T.; Toyoda, Y.; Masuo, Y.; Yamamoto, H.; Suzuki, H. NPC1L1 Is a Key Regulator of Intestinal Vitamin K Absorption and a Modulator of Warfarin Therapy. Sci. Transl. Med. 2015, 7, 275ra23. [Google Scholar] [CrossRef] [PubMed]
- Hollander, D.; Rim, E.; Ruble, P.E. Vitamin K2 Colonic and Ileal in Vivo Absorption: Bile, Fatty Acids, and PH Effects on Transport. Am. J. Physiol. 1977, 233, E124-9. [Google Scholar] [CrossRef]
- Gardill, S.L.; Suttie, J.W. Vitamin K Epoxide and Quinone Reductase Activities: Evidence for Reduction by a Common Enzyme. Biochem. Pharmacol. 1990, 40, 1055–1061. [Google Scholar] [CrossRef]
- Jin, D.-Y.; Tie, J.-K.; Stafford, D.W. The Conversion of Vitamin K Epoxide to Vitamin K Quinone and Vitamin K Quinone to Vitamin K Hydroquinone Uses the Same Active Site Cysteines. Biochemistry 2007, 46, 7279–7283. [Google Scholar] [CrossRef]
- Berkner, K.L. Vitamin K-Dependent Carboxylation. Vitam. Horm. 2008, 78, 131–156. [Google Scholar] [PubMed]
- Stafford, D.W. The Vitamin K Cycle. J. Thromb. Haemost. 2005, 3, 1873–1878. [Google Scholar] [CrossRef] [PubMed]
- Oldenburg, J.; Marinova, M.; Müller-Reible, C.; Watzka, M. The Vitamin K Cycle. Vitam. Horm. 2008, 78, 35–62. [Google Scholar] [PubMed]
- Bianchet, M.A.; Erdemli, S.B.; Amzel, L.M. Structure, Function, and Mechanism of Cytosolic Quinone Reductases. Vitam. Horm. 2008, 78, 63–84. [Google Scholar] [PubMed]
- Michaux, A.; Matagrin, B.; Debaux, J.-V.; Schurgers, L.J.; Benoit, E.; Lattard, V. Missense Mutation of VKORC1 Leads to Medial Arterial Calcification in Rats. Sci. Rep. 2018, 8, 13733. [Google Scholar] [CrossRef]
- Tie, J.-K.; Stafford, D.W. Functional Study of the Vitamin K Cycle Enzymes in Live Cells. Methods Enzymol. 2017, 584, 349–394. [Google Scholar] [PubMed] [Green Version]
- Watzka, M.; Geisen, C.; Scheer, M.; Wieland, R.; Wiegering, V.; Dörner, T.; Laws, H.-J.; Gümrük, F.; Hanalioglu, S.; Ünal, S.; et al. Bleeding and Non-Bleeding Phenotypes in Patients with GGCX Gene Mutations. Thromb. Res. 2014, 134, 856–865. [Google Scholar] [CrossRef] [PubMed]
- Tie, J.-K.; Carneiro, J.D.A.; Jin, D.-Y.; Martinhago, C.D.; Vermeer, C.; Stafford, D.W. Characterization of Vitamin K-Dependent Carboxylase Mutations That Cause Bleeding and Nonbleeding Disorders. Blood 2016, 127, 1847–1855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanakker, O.M.; Martin, L.; Gheduzzi, D.; Leroy, B.P.; Loeys, B.L.; Guerci, V.I.; Matthys, D.; Terry, S.F.; Coucke, P.J.; Pasquali-Ronchetti, I.; et al. Pseudoxanthoma Elasticum-Like Phenotype with Cutis Laxa and Multiple Coagulation Factor Deficiency Represents a Separate Genetic Entity. J. Investig. Dermatol. 2007, 127, 581–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villa, J.K.D.; Diaz, M.A.N.; Pizziolo, V.R.; Martino, H.S.D. Effect of Vitamin K in Bone Metabolism and Vascular Calcification: A Review of Mechanisms of Action and Evidences. Crit. Rev. Food Sci. Nutr. 2017, 57, 3959–3970. [Google Scholar] [CrossRef] [PubMed]
- Buchanan, G.S.; Melvin, T.; Merritt, B.; Bishop, C.; Shuler, F.D. Vitamin K2 (Menaquinone) Supplementation and Its Benefits in Cardiovascular Disease, Osteoporosis, and Cancer. Marshall J. Med. 2016, 2, 8. [Google Scholar] [CrossRef] [Green Version]
- Rani, S.; Barbe, M.F.; Barr, A.E.; Litvin, J. Periostin-like-Factor and Periostin in an Animal Model of Work-Related Musculoskeletal Disorder. Bone 2009, 44, 502–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conway, S.J.; Izuhara, K.; Kudo, Y.; Litvin, J.; Markwald, R.; Ouyang, G.; Arron, J.R.; Holweg, C.T.J.; Kudo, A. The Role of Periostin in Tissue Remodeling across Health and Disease. Cell. Mol. Life Sci. 2014, 71, 1279–1288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, M.; Yokota, K.; Saito, T.; Kobayakawa, K.; Kijima, K.; Yoshizaki, S.; Okazaki, K.; Yoshida, S.; Matsumoto, Y.; Harimaya, K.; et al. Periostin Promotes Fibroblast Migration and Inhibits Muscle Repair After Skeletal Muscle Injury. J. Bone Jt. Surg. 2018, 100, e108. [Google Scholar] [CrossRef] [PubMed]
- Kulman, J.D.; Harris, J.E.; Xie, L.; Davie, E.W. Identification of Two Novel Transmembrane γ-Carboxyglutamic Acid Proteins Expressed Broadly in Fetal and Adult Tissues. Proc. Natl. Acad. Sci. USA 2001, 98, 1370. [Google Scholar] [CrossRef] [PubMed]
- Viegas, C.S.B.; Simes, D.C.; Laizé, V.; Williamson, M.K.; Price, P.A.; Cancela, M.L. Gla-Rich Protein (GRP), A New Vitamin K-Dependent Protein Identified from Sturgeon Cartilage and Highly Conserved in Vertebrates. J. Biol. Chem. 2008, 283, 36655–36664. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vermeer, C.V. Vitamin K: The Effect on Health beyond Coagulation—An Overview. Food Nutr. Res. 2012, 56, 5329. [Google Scholar] [CrossRef] [PubMed]
- van Summeren, M.J.H.; Braam, L.A.J.L.M.; Lilien, M.R.; Schurgers, L.J.; Kuis, W.; Vermeer, C. The Effect of Menaquinone-7 (Vitamin K2) Supplementation on Osteocalcin Carboxylation in Healthy Prepubertal Children. Br. J. Nutr. 2009, 102, 1171–1178. [Google Scholar] [CrossRef] [Green Version]
- Theuwissen, E.; Cranenburg, E.C.; Knapen, M.H.; Magdeleyns, E.J.; Teunissen, K.J.; Schurgers, L.J.; Smit, E.; Vermeer, C. Low-Dose Menaquinone-7 Supplementation Improved Extra-Hepatic Vitamin K Status, but Had No Effect on Thrombin Generation in Healthy Subjects. Br. J. Nutr. 2012, 108, 1652–1657. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Ren, R.; Xu, D. Vitamin K2 Supplementation Does Not Affect Vitamin K-Dependent Coagulation Factors Activity in Healthy Subjects: A Pilot Study. Curr. Dev. Nutr. 2020, 4, 1843. [Google Scholar] [CrossRef]
- Ren, R.; Liu, J.; Cheng, G.; Tan, J. Vitamin K2 (Menaquinone-7) Supplementation Does Not Affect Vitamin K-Dependent Coagulation Factors Activity in Healthy Individuals. Medicine 2021, 100, e26221. [Google Scholar] [CrossRef]
- Binkley, N.C.; Krueger, D.C.; Engelke, J.A.; Foley, A.L.; Suttie, J.W. Vitamin K Supplementation Reduces Serum Concentrations of Under-γ-Carboxylated Osteocalcin in Healthy Young and Elderly Adults. Am. J. Clin. Nutr. 2000, 72, 1523–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westenfeld, R.; Krueger, T.; Schlieper, G.; Cranenburg, E.C.M.; Magdeleyns, E.J.; Heidenreich, S.; Holzmann, S.; Vermeer, C.; Jahnen-Dechent, W.; Ketteler, M.; et al. Effect of Vitamin K2 Supplementation on Functional Vitamin K Deficiency in Hemodialysis Patients: A Randomized Trial. Am. J. Kidney Dis. 2012, 59, 186–195. [Google Scholar] [CrossRef] [PubMed]
- Neve, A.; Corrado, A.; Cantatore, F.P. Osteocalcin: Skeletal and Extra-Skeletal Effects. J. Cell. Physiol. 2013, 228, 1149–1153. [Google Scholar] [CrossRef]
- Poundarik, A.A.; Diab, T.; Sroga, G.E.; Ural, A.; Boskey, A.L.; Gundberg, C.M.; Vashishth, D. Dilatational Band Formation in Bone. Proc. Natl. Acad. Sci. USA 2012, 109, 19178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikel, O.; Laurencin, D.; McCallum, S.A.; Gundberg, C.M.; Vashishth, D. NMR Investigation of the Role of Osteocalcin and Osteopontin at the Organic–Inorganic Interface in Bone. Langmuir 2013, 29, 13873–13882. [Google Scholar] [CrossRef] [Green Version]
- Boskey, A.L.; Gadaleta, S.; Gundberg, C.; Doty, S.B.; Ducy, P.; Karsenty, G. Fourier Transform Infrared Microspectroscopic Analysis of Bones of Osteocalcin-Deficient Mice Provides Insight into the Function of Osteocalcin. Bone 1998, 23, 187–196. [Google Scholar] [CrossRef]
- Tsao, Y.-T.; Huang, Y.-J.; Wu, H.-H.; Liu, Y.-A.; Liu, Y.-S.; Lee, O. Osteocalcin Mediates Biomineralization during Osteogenic Maturation in Human Mesenchymal Stromal Cells. Int. J. Mol. Sci. 2017, 18, 159. [Google Scholar] [CrossRef] [PubMed]
- Patti, A.; Gennari, L.; Merlotti, D.; Dotta, F.; Nuti, R. Endocrine Actions of Osteocalcin. Int. J. Endocrinol. 2013, 2013, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, X.; Brennan-Speranza, T.C.; Levinger, I.; Yeap, B.B. Undercarboxylated Osteocalcin: Experimental and Human Evidence for a Role in Glucose Homeostasis and Muscle Regulation of Insulin Sensitivity. Nutrients 2018, 10, 847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delmas, P.D. Clinical Use of Biochemical Markers of Bone Remodeling in Osteoporosis. Bone 1992, 13, S17–S21. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Weitzmann, M.N. Vitamin K2 Stimulates Osteoblastogenesis and Suppresses Osteoclastogenesis by Suppressing NF-ΚB Activation. Int. J. Mol. Med. 2011, 27, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Uchiyama, S.; Yamaguchi, M. Anabolic Effect of β-Cryptoxanthin in Osteoblastic MC3T3-E1 Cells Is Enhanced with 17β-Estradiol, Genistein, or Zinc Sulfate in Vitro: The Unique Effect with Zinc on Runx2 and A1(I) Collagen MRNA Expressions. Mol. Cell. Biochem. 2007, 307, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Ouchi, Y.; Inoue, S. Vitamin K: Novel Molecular Mechanisms of Action and Its Roles in Osteoporosis: Molecular Mechanism of Vitamin K. Geriatr. Gerontol. Int. 2014, 14, 1–7. [Google Scholar] [CrossRef]
- Ichikawa, T.; Horie-Inoue, K.; Ikeda, K.; Blumberg, B.; Inoue, S. Steroid and Xenobiotic Receptor SXR Mediates Vitamin K 2 -Activated Transcription of Extracellular Matrix-Related Genes and Collagen Accumulation in Osteoblastic Cells. J. Biol. Chem. 2006, 281, 16927–16934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cranenburg, E.C.M.; Vermeer, C.; Koos, R.; Boumans, M.-L.; Hackeng, T.M.; Bouwman, F.G.; Kwaijtaal, M.; Brandenburg, V.M.; Ketteler, M.; Schurgers, L.J. The Circulating Inactive Form of Matrix Gla Protein (UcMGP) as a Biomarker for Cardiovascular Calcification. J. Vasc. Res. 2008, 45, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Mayer, O.; Seidlerová, J.; Bruthans, J.; Filipovský, J.; Timoracká, K.; Vaněk, J.; Černá, L.; Wohlfahrt, P.; Cífková, R.; Theuwissen, E.; et al. Desphospho-Uncarboxylated Matrix Gla-Protein Is Associated with Mortality Risk in Patients with Chronic Stable Vascular Disease. Atherosclerosis 2014, 235, 162–168. [Google Scholar] [CrossRef]
- Yao, Y.; Bennett, B.J.; Wang, X.; Rosenfeld, M.E.; Giachelli, C.; Lusis, A.J.; Bostrom, K.I. Inhibition of Bone Morphogenetic Proteins Protects Against Atherosclerosis and Vascular Calcification. Circ. Res. 2010, 107, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Delanaye, P.; Krzesinski, J.-M.; Warling, X.; Moonen, M.; Smelten, N.; Médart, L.; Pottel, H.; Cavalier, E. Dephosphorylated-Uncarboxylated Matrix Gla Protein Concentration Is Predictive of Vitamin K Status and Is Correlated with Vascular Calcification in a Cohort of Hemodialysis Patients. BMC Nephrol. 2014, 15, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farzaneh-Far, A.; Weissberg, P.L.; Proudfoot, D.; Shanahan, C.M. Transcriptional Regulation of Matrix Gla Protein. Z. Für Kardiol. 2001, 90, 38–42. [Google Scholar] [CrossRef] [PubMed]
- Osako, M.K.; Nakagami, H.; Koibuchi, N.; Shimizu, H.; Nakagami, F.; Koriyama, H.; Shimamura, M.; Miyake, T.; Rakugi, H.; Morishita, R. Estrogen Inhibits Vascular Calcification via Vascular RANKL System: Common Mechanism of Osteoporosis and Vascular Calcification. Circ. Res. 2010, 107, 466–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dihingia, A.; Kalita, J.; Manna, P. Implication of a Novel Gla-Containing Protein, Gas6 in the Pathogenesis of Insulin Resistance, Impaired Glucose Homeostasis, and Inflammation: A Review. Diabetes Res. Clin. Pract. 2017, 128, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.-S.; Hung, Y.-J.; Lee, C.-H.; Hsiao, F.-C.; Hsieh, P.-S. The Involvement of GAS6 Signaling in the Development of Obesity and Associated Inflammation. Int. J. Endocrinol. 2015, 2015, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakaue, M.; Mori, N.; Okazaki, M.; Kadowaki, E.; Kaneko, T.; Hemmi, N.; Sekiguchi, H.; Maki, T.; Ozawa, A.; Hara, S.; et al. Vitamin K Has the Potential to Protect Neurons from Methylmercury-Induced Cell Death in Vitro. J. Neurosci. Res. 2011, 89, 1052–1058. [Google Scholar] [CrossRef] [PubMed]
- Hadipour, E.; Tayarani-Najaran, Z.; Fereidoni, M. Vitamin K2 Protects PC12 Cells against Aβ (1-42) and H2O2-Induced Apoptosis via P38 MAP Kinase Pathway. Nutr. Neurosci. 2020, 23, 343–352. [Google Scholar] [CrossRef] [PubMed]
- Vos, M.; Esposito, G.; Edirisinghe, J.N.; Vilain, S.; Haddad, D.M.; Slabbaert, J.R.; Van Meensel, S.; Schaap, O.; De Strooper, B.; Meganathan, R.; et al. Vitamin K2 Is a Mitochondrial Electron Carrier That Rescues Pink1 Deficiency. Science 2012, 336, 1306–1310. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Li, Y.; Gao, F.; Hu, Q.; Zhang, Y.; Chen, D.; Wang, G. Vitamin K2 Suppresses Rotenone-Induced Microglial Activation in Vitro. Acta Pharmacol. Sin. 2016, 37, 1178–1189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nimptsch, K.; Rohrmann, S.; Linseisen, J. Dietary Intake of Vitamin K and Risk of Prostate Cancer in the Heidelberg Cohort of the European Prospective Investigation into Cancer and Nutrition (EPIC-Heidelberg). Am. J. Clin. Nutr. 2008, 87, 985–992. [Google Scholar] [CrossRef] [PubMed]
- Gast, G.C.M.; de Roos, N.M.; Sluijs, I.; Bots, M.L.; Beulens, J.W.J.; Geleijnse, J.M.; Witteman, J.C.; Grobbee, D.E.; Peeters, P.H.M.; van der Schouw, Y.T. A High Menaquinone Intake Reduces the Incidence of Coronary Heart Disease. Nutr. Metab. Cardiovasc. Dis. 2009, 19, 504–510. [Google Scholar] [CrossRef] [PubMed]
- Apalset, E.M.; Gjesdal, C.G.; Eide, G.E.; Tell, G.S. Intake of Vitamin K1 and K2 and Risk of Hip Fractures: The Hordaland Health Study. Bone 2011, 49, 990–995. [Google Scholar] [CrossRef] [PubMed]
- Vissers, L.E.T.; Dalmeijer, G.W.; Boer, J.M.A.; Monique Verschuren, W.M.; van der Schouw, Y.T.; Beulens, J.W.J. Intake of Dietary Phylloquinone and Menaquinones and Risk of Stroke. J. Am. Heart Assoc. 2013, 2, e000455. [Google Scholar] [CrossRef] [Green Version]
- Turck, D.; Bresson, J.-L.; Burlingame, B.; Dean, T.; Fairweather-Tait, S.; Heinonen, M.; Hirsch-Ernst, K.I.; Mangelsdorf, I.; McArdle, H.J.; Naska, A.; et al. Dietary Reference Values for Vitamin K. EFSA J. 2017, 15, e04780. [Google Scholar] [CrossRef] [PubMed]
- Forli, L.; Bollerslev, J.; Simonsen, S.; Isaksen, G.A.; Kvamsdal, K.E.; Godang, K.; Gadeholt, G.; Pripp, A.H.; Bjortuft, O. Dietary Vitamin K2 Supplement Improves Bone Status after Lung and Heart Transplantation. Transplantation 2010, 89, 458–464. [Google Scholar] [CrossRef] [PubMed]
- Knapen, M.; Drummen, N.; Smit, E.; Vermeer, C.; Theuwissen, E. Three-Year Low-Dose Menaquinone-7 Supplementation Helps Decrease Bone Loss in Healthy Postmenopausal Women. Osteoporos. Int. 2013, 24, 2499–2507. [Google Scholar] [CrossRef] [PubMed]
- Booth, S.L. Roles for Vitamin K Beyond Coagulation. Annu. Rev. Nutr. 2009, 29, 89–110. [Google Scholar] [CrossRef] [PubMed]
- Shearer, M.J.; Fu, X.; Booth, S.L. Vitamin K Nutrition, Metabolism, and Requirements: Current Concepts and Future Research. Adv. Nutr. 2012, 3, 182–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walther, B.; Karl, J.P.; Booth, S.L.; Boyaval, P. Menaquinones, Bacteria, and the Food Supply: The Relevance of Dairy and Fermented Food Products to Vitamin K Requirements. Adv. Nutr. 2013, 4, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Kamao, M.; Suhara, Y.; Tsugawa, N.; Uwano, M.; Yamaguchi, N.; Uenishi, K.; Ishida, H.; Sasaki, S.; Okano, T. Vitamin K Content of Foods and Dietary Vitamin K Intake in Japanese Young Women. J. Nutr. Sci. Vitaminol. 2007, 53, 464–470. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Yamada, Y.; Ohtani, Y.; Mitsui, N.; Murasawa, H.; Araki, S. Production of Menaquinone (Vitamin K2)-7 by Bacillus Subtilis. J. Biosci. Bioeng. 2001, 91, 16–20. [Google Scholar] [CrossRef]
- Snyder, C.D.; Rapoport, H. Synthesis of Menaquinones. J. Am. Chem. Soc. 1974, 96, 8046–8054. [Google Scholar] [CrossRef] [PubMed]
- Daines, A.M.; Payne, R.J.; Humphries, M.E.; Abell, A.D. The Synthesis of Naturally Occurring Vitamin K and Vitamin K Analogues. Curr. Org. Chem. 2003, 7, 1625–1634. [Google Scholar] [CrossRef] [Green Version]
- Baj, A.; Wałejko, P.; Kutner, A.; Kaczmarek, Ł.; Morzycki, J.W.; Witkowski, S. Convergent Synthesis of Menaquinone-7 (MK-7). Org. Process. Res. Dev. 2016, 20, 1026–1033. [Google Scholar] [CrossRef]
- Knauer, T.E.; Siegfried, C.; Willingham, A.K.; Matschiner, J.T. Metabolism and Biological Activity of Cis- and Trans-Phylloquinone in the Rat. J. Nutr. 1975, 105, 1519–1524. [Google Scholar] [CrossRef]
- Lowenthal, J.; Rivera, G.M.V. Comparison of the Activity of the Cis and Trans Isomer of Vitamin K1 in Vitamin K-Deficient and Coumarin Anticoagulant-Pretreated Rats. J. Pharmacol. Exp. Ther. 1979, 209, 330–333. [Google Scholar] [PubMed]
- Matschiner, J.T.; Bell, R.G. Metabolism and Vitamin K Activity of Cis Phylloquinone in Rats. J. Nutr. 1972, 102, 625–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thijssen, H.H.; Drittij-Reijnders, M.J. Vitamin K Status in Human Tissues: Tissue-Specific Accumulation of Phylloquinone and Menaquinone-4. Br. J. Nutr. 1996, 75, 121–127. [Google Scholar] [CrossRef] [PubMed]
- Schurgers, L.J.; Teunissen, K.J.; Hamulyák, K.; Knapen, M.H.; Vik, H.; Vermeer, C. Vitamin K–Containing Dietary Supplements: Comparison of Synthetic Vitamin K1 and Natto-Derived Menaquinone-7. Blood 2007, 109, 3279–3283. [Google Scholar] [CrossRef] [Green Version]
- Sato, T.; Schurgers, L.J.; Uenishi, K. Comparison of Menaquinone-4 and Menaquinone-7 Bioavailability in Healthy Women. Nutr. J. 2012, 11, 93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garber, A.K.; Binkley, N.C.; Krueger, D.C.; Suttie, J.W. Comparison of Phylloquinone Bioavailability from Food Sources or a Supplement in Human Subjects. J. Nutr. 1999, 129, 1201–1203. [Google Scholar] [CrossRef] [PubMed]
- Schurgers, L.J.; Vermeer, C. Determination of Phylloquinone and Menaquinones in Food. Haemostasis 2000, 30, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Schurgers, L.J.; Vermeer, C. Differential Lipoprotein Transport Pathways of K-Vitamins in Healthy Subjects. Biochim. Biophys. Acta 2002, 1570, 27–32. [Google Scholar] [CrossRef]
- Buitenhuis, H.; Soute, B.; Vermeer, C. Comparison of the Vitamins K1, K2 and K3 as Cofactors for the Hepatic Vitamin K-Dependent Carboxylase. Biochim. Et Biophys. Acta (BBA)-Gen. Subj. 1990, 1034, 170–175. [Google Scholar] [CrossRef]
- Inaba, N.; Sato, T.; Yamashita, T. Low-Dose Daily Intake of Vitamin K(2) (Menaquinone-7) Improves Osteocalcin γ-Carboxylation: A Double-Blind, Randomized Controlled Trials. J. Nutr. Sci. Vitaminol. 2015, 61, 471–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakamura, E.; Aoki, M.; Watanabe, F.; Kamimura, A. Low-Dose Menaquinone-4 Improves γ-Carboxylation of Osteocalcin in Young Males: A Non-Placebo-Controlled Dose–Response Study. Nutr. J. 2014, 13, 85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzaka, T.; Yoshinaga, M.; Tsuji, Y. Prophylaxis of Intracranial Hemorrhage Due to Vitamin K Deficiency in Infants. Brain Dev. 1987, 9, 305–308. [Google Scholar] [CrossRef]
- Knapen, M.H.J.; Schurgers, L.J.; Vermeer, C. Vitamin K2 Supplementation Improves Hip Bone Geometry and Bone Strength Indices in Postmenopausal Women. Osteoporos. Int. 2007, 18, 963–972. [Google Scholar] [CrossRef] [Green Version]
- Kanellakis, S.; Moschonis, G.; Tenta, R.; Schaafsma, A.; van den Heuvel, E.G.H.M.; Papaioannou, N.; Lyritis, G.; Manios, Y. Changes in Parameters of Bone Metabolism in Postmenopausal Women Following a 12-Month Intervention Period Using Dairy Products Enriched with Calcium, Vitamin D, and Phylloquinone (Vitamin K1) or Menaquinone-7 (Vitamin K2): The Postmenopausal Health Study II. Calcif. Tissue Int. 2012, 90, 251–262. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Wang, H.; Shi, Y.; Shen, Q.; Tsamlag, L.; Wang, Z.; Yu, S.; Shen, T.; Wang, Y.; Cai, Y. Impact of Calcium, Vitamin D, Vitamin K, Oestrogen, Isoflavone and Exercise on Bone Mineral Density for Osteoporosis Prevention in Postmenopausal Women: A Network Meta-Analysis. Br. J. Nutr. 2020, 123, 84–103. [Google Scholar] [CrossRef]
- Geleijnse, J.M.; Vermeer, C.; Grobbee, D.E.; Schurgers, L.J.; Knapen, M.H.J.; van der Meer, I.M.; Hofman, A.; Witteman, J.C.M.; Witteman, J.C.M. Dietary Intake of Menaquinone Is Associated with a Reduced Risk of Coronary Heart Disease: The Rotterdam Study. J. Nutr. 2004, 134, 3100–3105. [Google Scholar] [CrossRef] [PubMed]
- Beulens, J.W.J.; Bots, M.L.; Atsma, F.; Bartelink, M.-L.E.L.; Prokop, M.; Geleijnse, J.M.; Witteman, J.C.M.; Grobbee, D.E.; van der Schouw, Y.T. High Dietary Menaquinone Intake Is Associated with Reduced Coronary Calcification. Atherosclerosis 2009, 203, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Yanagita, M.; Arai, H.; Ishii, K.; Nakano, T.; Ohashi, K.; Mizuno, K.; Varnum, B.; Fukatsu, A.; Doi, T.; Kita, T. Gas6 Regulates Mesangial Cell Proliferation through Axl in Experimental Glomerulonephritis. Am. J. Pathol. 2001, 158, 1423–1432. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bus, K.; Szterk, A. Relationship between Structure and Biological Activity of Various Vitamin K Forms. Foods 2021, 10, 3136. https://doi.org/10.3390/foods10123136
Bus K, Szterk A. Relationship between Structure and Biological Activity of Various Vitamin K Forms. Foods. 2021; 10(12):3136. https://doi.org/10.3390/foods10123136
Chicago/Turabian StyleBus, Katarzyna, and Arkadiusz Szterk. 2021. "Relationship between Structure and Biological Activity of Various Vitamin K Forms" Foods 10, no. 12: 3136. https://doi.org/10.3390/foods10123136
APA StyleBus, K., & Szterk, A. (2021). Relationship between Structure and Biological Activity of Various Vitamin K Forms. Foods, 10(12), 3136. https://doi.org/10.3390/foods10123136