
Rice Bran Oil Attenuates Chronic Inflammation by Inducing M2 Macrophage Switching in High-Fat Diet-Fed Obese Mice


Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Dietary Interventions
2.2. RNA Extraction and Quantitative Reverse-Transcription PCR
2.3. Culture and Polarization of Bone Marrow-Derived Macrophages (BMDMs)
2.4. Assessment of Macrophage Activation Markers
2.5. Cytokine Quantification
2.6. Statistical Analysis
2.7. Ethics Statement
3. Results
3.1. Body Weight Gain Was Suppressed by RBO in High-Fat Diet-Fed C57BL/6 Mice
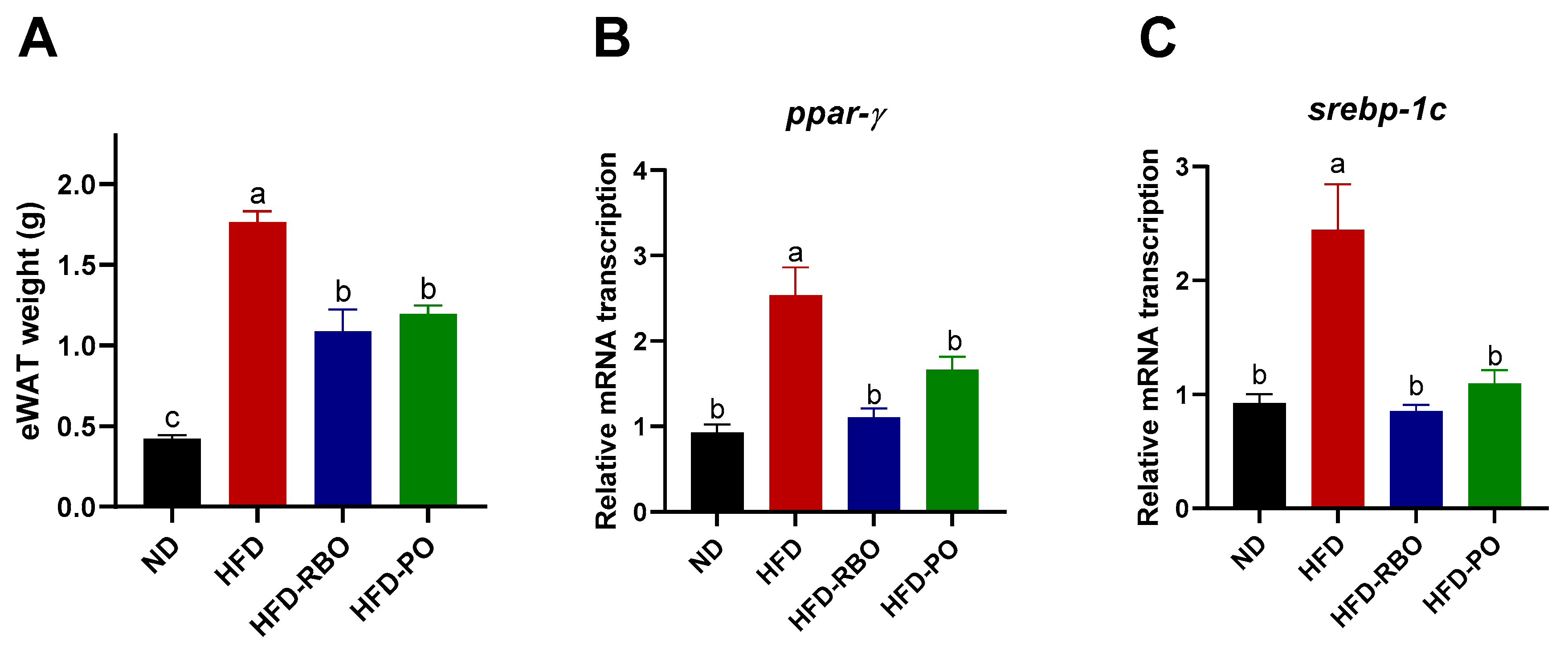
3.2. RBO and PO Were Associated with Downregulation of Adipogenesis in Mice Epididymal White Adipose Tissue (eWAT)
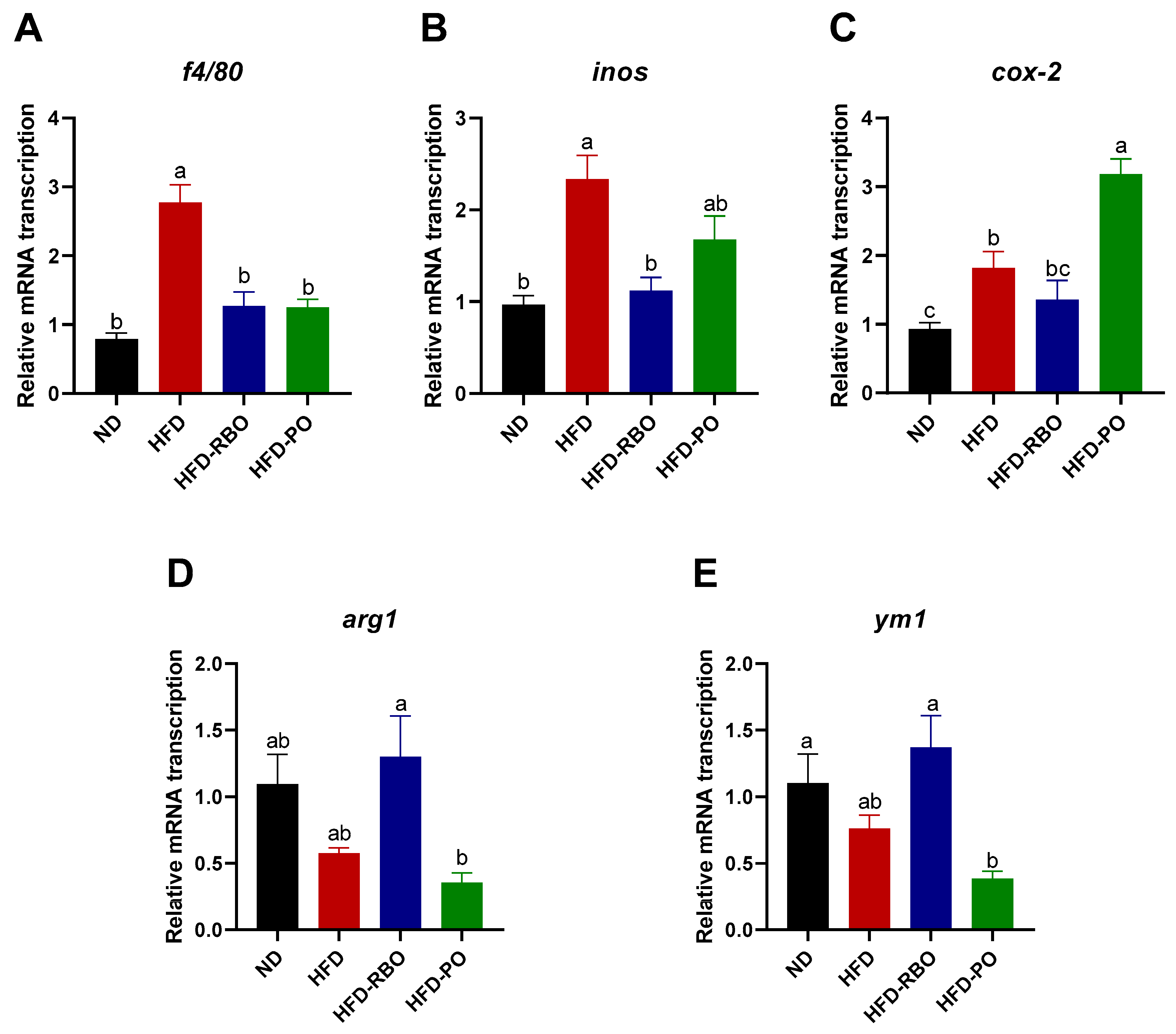
3.3. Rice Bran Oil Induces the Expression of M2-Macrophage Markers in the eWAT of HFD-Fed Mice
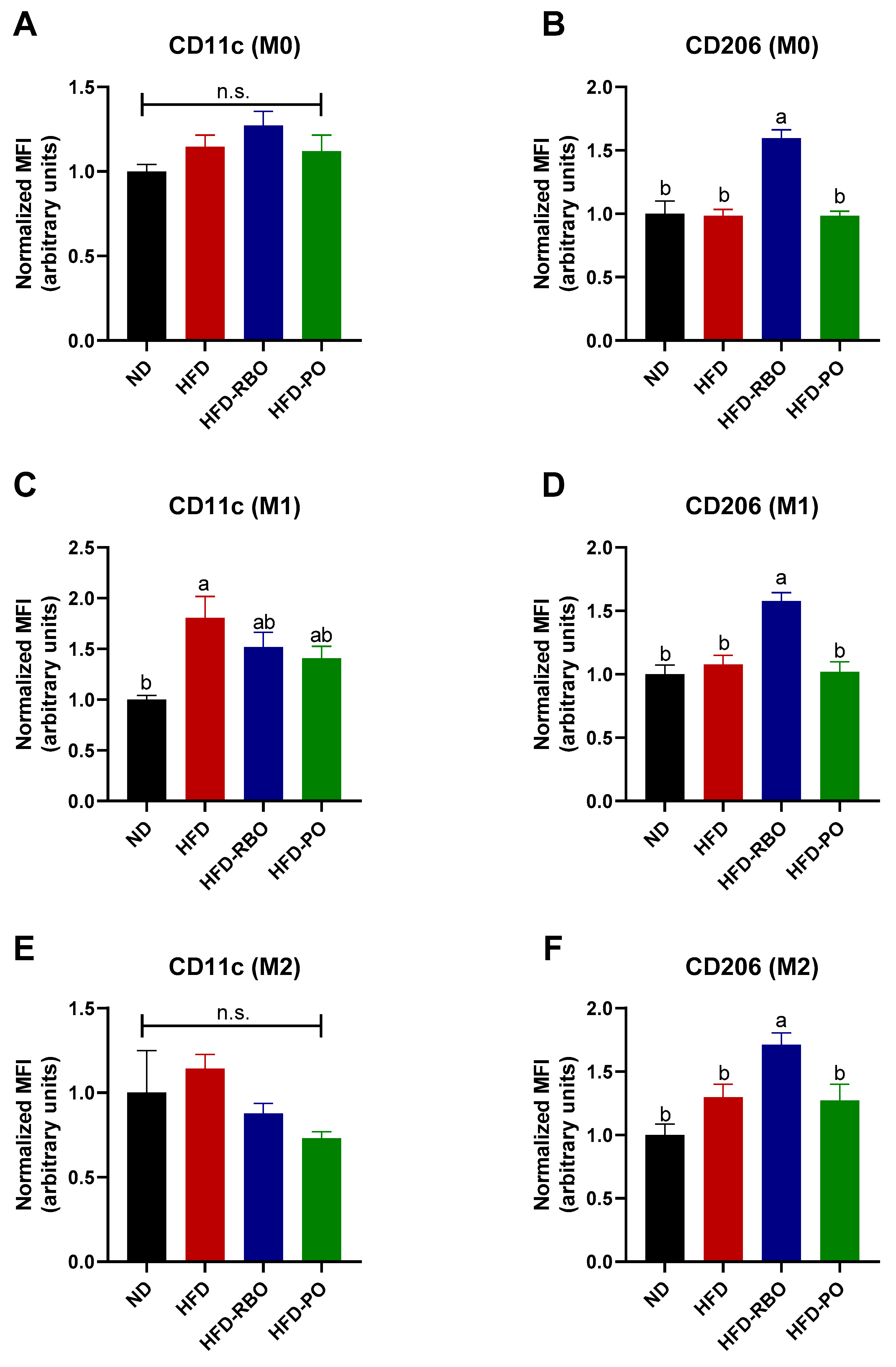
3.4. Rice Bran Oil Increases Surface M2 Marker Expression in Mouse BMDMs
3.5. Rice Bran Oil Regulates Cytokine Production in M1 and M2 BMDMs
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Nathan, C. Points of Control in Inflammation. Nature 2002, 420, 846–852. [Google Scholar] [CrossRef]
- Oishi, Y.; Manabe, I. Macrophages in Age-related Chronic Inflammatory Diseases. NPJ Aging Mech. Dis. 2016, 2, 16018. [Google Scholar] [CrossRef] [Green Version]
- Coppack, S.W. Pro-inflammatory Cytokines and Adipose Tissue. Proc. Nutr. Soc. 2001, 60, 349–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic Adipose Tissue Inflammation Linking Obesity to Insulin Resistance and Type 2 Diabetes. Front. Physiol. 2020, 10, 1607. [Google Scholar] [CrossRef] [PubMed]
- Kosteli, A.; Sugaru, E.; Haemmerle, G.; Martin, J.F.; Lei, J.; Zechner, R.; Ferrante, A.W. Weight Loss and Lipolysis Promote a Dynamic Immune Response in Murine Adipose Tissue. J. Clin. Investig. 2010, 120, 3466–3479. [Google Scholar] [CrossRef] [Green Version]
- Forsythe, L.K.; Wallace, J.M.W.; Livingstone, M.B.E. Obesity and Inflammation: The Effects of Weight Loss. Nutr. Res. Rev. 2008, 21, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Oishi, Y.; Manabe, I. Macrophages in Inflammation, Repair and Regeneration. Int. Immunol. 2018, 30, 511–528. [Google Scholar] [CrossRef]
- Liu, Y.-C.; Zou, X.-B.; Chai, Y.-F.; Yao, Y.-M. Macrophage Polarization in Inflammatory Diseases. Int. J. Biol. Sci. 2014, 10, 520–529. [Google Scholar] [CrossRef]
- Martinez, F.O.; Sica, A.; Mantovani, A.; Locati, M. Macrophage Activation and Polarization. Front. Biosci. 2008, 13, 453–461. [Google Scholar] [CrossRef] [Green Version]
- Lavin, Y.; Mortha, A.; Rahman, A.; Merad, M. Regulation of Macrophage Development and Function in Peripheral Tissues. Nat. Rev. Immunol. 2015, 15, 731–744. [Google Scholar] [CrossRef] [Green Version]
- Lumeng, C.N.; Bodzin, J.L.; Saltiel, A.R. Obesity Induces a Phenotypic Switch in Adipose Tissue Macrophage Polarization. J. Clin. Investig. 2007, 117, 175–184. [Google Scholar] [CrossRef] [Green Version]
- Russo, L.; Lumeng, C.N. Properties and Functions of Adipose Tissue Macrophages in Obesity. Immunology 2018, 155, 407–417. [Google Scholar] [CrossRef]
- Romeo, G.R.; Lee, J.; Shoelson, S.E. Metabolic Syndrome, Insulin Resistance, and Roles of Inflammation- Mechanisms and Therapeutic Targets. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 1771–1776. [Google Scholar] [CrossRef] [Green Version]
- Galván-Peña, S.; O’Neill, L.A.J. Metabolic Reprogramming in Macrophage Polarization. Front. Immunol. 2014, 5, 420. [Google Scholar] [CrossRef] [Green Version]
- Kelly, B.; O’Neill, L.A.J. Metabolic Reprogramming in Macrophages and Dendritic Cells in Innate Immunity. Cell Res. 2015, 25, 771–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van den Bossche, J.; Baardman, J.; Otto, N.A.; van der Velden, S.; Neele, A.E.; van den Berg, S.M.; Luque-Martin, R.; Chen, H.J.; Boshuizen, M.C.S.; Ahmed, M.; et al. Mitochondrial Dysfunction Prevents Repolarization of Inflammatory Macrophages. Cell Rep. 2016, 17, 684–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Q.; Wang, H.; Mao, C.; Sun, M.; Dominah, G.; Chen, L.; Zhuang, Z. Fatty Acid Oxidation Contributes to IL-1β Secretion in M2 Macrophages and Promotes Macrophage-mediated Tumor Cell Migration. Mol. Immunol. 2018, 94, 27–35. [Google Scholar] [CrossRef]
- Yu, S.; Go, G.W.; Kim, W. Medium Chain Triglyceride (MCT) Oil Affects the Immunophenotype via Reprogramming of Mitochondrial Respiration in Murine Macrophages. Foods 2019, 8, 553. [Google Scholar] [CrossRef] [Green Version]
- Ohue-Kitano, R.; Yasuoka, Y.; Goto, T.; Kitamura, N.; Park, S.B.; Kishino, S.; Kimura, I.; Kasubuchi, M.; Takahashi, H.; Li, Y.; et al. α-Linolenic acid–derived Metabolites from Gut Lactic Acid Bacteria Induce Differentiation of Anti-inflammatory M2 Macrophages through G Protein-coupled Receptor 40. FASEB J. 2018, 32, 304–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Boer, A.A.; Monk, J.M.; Liddle, D.M.; Power, K.A.; Ma, D.W.L.; Robinson, L.E. Fish Oil-derived Long-chain n-3 Polyunsaturated Fatty Acids Reduce Expression of M1-associated Macrophage Markers in an Ex Vivo Adipose Tissue Culture Model, in part through Adiponectin. Front. Nutr. 2015, 2, 31. [Google Scholar] [CrossRef] [Green Version]
- Friedman, M. Rice Brans, Rice Bran Oils, and Rice Hulls: Composition, Food and Industrial Uses, and Bioactivities in Humans, Animals, and Cells. J. Agric. Food Chem. 2013, 61, 10626–10641. [Google Scholar] [CrossRef]
- Soares, J.F.; dal Prá, V.; de Souza, M.; Lunelli, F.C.; Abaide, E.; da Silva, J.R.F.; Kuhn, R.C.; Martínez, J.; Mazutti, M.A. Extraction of Rice Bran Oil Using Supercritical CO2 and Compressed Liquefied Petroleum Gas. J. Food Eng. 2016, 170, 58–63. [Google Scholar] [CrossRef] [Green Version]
- Minhajuddin, M.; Beg, Z.H.; Iqbal, J. Hypolipidemic and Antioxidant Properties of Tocotrienol Rich Fraction Isolated from Rice Bran Oil in Experimentally Induced Hyperlipidemic Rats. Food Chem. Toxicol. 2005, 43, 747–753. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.-W.; Cheng, H.-H. A Rice Bran Oil Diet Increases LDL-Receptor and HMG-CoA Reductase mRNA Expressions and Insulin Sensitivity in Rats with Streptozotocin/Nicotinamide-induced Type 2 Diabetes. J. Nutr. 2006, 136, 1472–1476. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.A.; Mohamed, M.A.; Rashed, L.A.; Abd Elbast, S.A.; Ahmed, E.A. Rice Bran Oil Improves Insulin Resistance by Affecting the Expression of Antioxidants and Lipid-regulatory Genes. Lipids 2018, 53, 505–515. [Google Scholar] [CrossRef]
- Rao, Y.P.C.; Sugasini, D.; Lokesh, B.R. Dietary Gamma Oryzanol Plays a Significant Role in the Anti-inflammatory Activity of Rice Bran Oil by Decreasing Pro-inflammatory Mediators Secreted by Peritoneal Macrophages of Rats. Biochem. Biophys. Res. Commun. 2016, 479, 747–752. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Yu, S.; Park, H.J.; Jung, J.; Go, G.W.; Kim, W. Rice Bran Oil Ameliorates Inflammatory Responses by Enhancing Mitochondrial Respiration in Murine Macrophages. PLoS ONE 2019, 14, 222857. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.Y.; Kim, G.R.; Chae, J.S.; Kan, H.; Kim, S.S.; Hwang, K.S.; Lee, B.H.; Yu, S.; Moon, S.; Park, B.; et al. Antioxidant and Anti-inflammatory Effects of an Ethanol Fraction from the Schisandra Chinensis Baillon Hot Water Extract Fermented Using Lactobacilius Paracasei Subsp. Tolerans. Food Sci. Biotechnol. 2019, 28, 1759–1767. [Google Scholar] [CrossRef] [PubMed]
- Hasturk, H.; Kantarci, A.; van Dyke, T.E. Oral Inflammatory Diseases and Systemic Inflammation: Role of the Macrophage. Front. Immunol. 2012, 3, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, S.; Choi, J.H.; Kim, H.J.; Park, S.H.; Go, G.W.; Kim, W. In Vitro Evidence of Anti-inflammatory and Anti-obesity Effects of Medium-Chain Fatty Acid-Diacylglycerols. J. Microbiol. Biotechnol. 2017, 27, 1617–1627. [Google Scholar] [CrossRef] [Green Version]
- Song, I.B.; Han, H.J.; Kwon, J. Immune-enhancing Effects of Gamma-irradiated Sericin. Food Sci. Biotechnol. 2020, 29, 969–976. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, S.; Wu, H.; Rong, X.; Guo, J. M2b Macrophage Polarization and Its Roles in Diseases. J. Leukoc. Biol. 2019, 106, 345–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castoldi, A.; de Souza, C.N.; Saraiva Camara, N.O.; Moraes-Vieira, P.M. The Macrophage Switch in Obesity Development. Front. Immunol. 2016, 6, 637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalmas, E.; Clément, K.; Guerre-Millo, M. Defining Macrophage Phenotype and Function in Adipose Tissue. Trends Immunol. 2011, 32, 307–314. [Google Scholar] [CrossRef]
- Lumeng, C.N.; de Young, S.M.; Bodzin, J.L.; Saltiel, A.R. Increased Inflammatory Properties of Adipose Tissue Macrophages Recruited during Diet-induced Obesity. Diabetes 2007, 56, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Gu, M.; Cai, W.; Yu, L.; Feng, L.; Zhang, L.; Zang, Q.; Wang, Y.; Wang, D.; Chen, H.; et al. Dietary Component Isorhamnetin Is a PPARγ Antagonist and Ameliorates Metabolic Disorders Induced by Diet or Leptin Deficiency. Sci. Rep. 2016, 6, 19288. [Google Scholar] [CrossRef] [PubMed]
- Jemai, R.; Drira, R.; Makni, M.; Fetoui, H.; Sakamoto, K. Colocynth (Citrullus Colocynthis) Seed Extracts Attenuate Adipogenesis by Down-regulating PPARγ/SREBP-1c and C/EBPα in 3T3-L1 Cells. Food Biosci. 2020, 33, 100491. [Google Scholar] [CrossRef]
- Baek, J.H.; Kim, S.J.; Kang, H.G.; Lee, H.W.; Kim, J.H.; Hwang, K.A.; Song, J.; Chun, K.H. Galectin-3 Activates PPARγ and Supports White Adipose Tissue Formation and High-Fat Diet-induced Obesity. Endocrinology 2015, 156, 147–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Li, Y.; Wang, Q.; Yang, Y. Crude Extracts from Lycium Barbarum Suppress SREBP-1c Expression and Prevent Diet-induced Fatty Liver through AMPK Activation. BioMed Res. Int. 2014, 2014, 196198. [Google Scholar] [CrossRef]
- Zainal, Z.; Rahim, A.A.; Khaza’ai, H.; Chang, S.K. Effects of Palm Oil Tocotrienol-rich Fraction (TRF) and Carotenes in Ovalbumin (OVA)-Challenged Asthmatic Brown Norway Rats. Int. J. Mol. Sci. 2019, 20, 1764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zainal, Z.; Longman, A.J.; Hurst, S.; Duggan, K.; Hughes, C.E.; Caterson, B.; Harwood, J.L. Modification of Palm Oil for Anti-inflammatory Nutraceutical Properties. Lipids 2009, 44, 581–592. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.J.; Liu, P.L.; Ng, L.T. Tocotrienol-rich Fraction of Palm Oil Exhibits Anti-inflammatory Property by Suppressing the Expression of Inflammatory Mediators in Human Monocytic Cells. Mol. Nutr. Food Res. 2008, 52, 921–929. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.Y.; Kamisah, Y.; Faizah, O.; Jubri, Z.; Qodriyah, H.M.S.; Jaarin, K. Involvement of Inflammation and Adverse Vascular Remodelling in the Blood Pressure Raising Effect of Repeatedly Heated Palm Oil in Rats. Int. J. Vasc. Med. 2012, 2012, 404025. [Google Scholar] [CrossRef] [PubMed]
- Nie, Y.; Luo, F.; Wang, L.; Yang, T.; Shi, L.; Li, X.; Shen, J.; Xu, W.; Guo, T.; Lin, Q. Anti-hyperlipidemic Effect of Rice Bran Polysaccharide and Its Potential Mechanism in High-Fat Diet Mice. Food Funct. 2017, 8, 4028–4041. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient (g/kg) | Normal Diet | High-Fat Diet | High-At Diet-RBO | High-Fat Diet-PO |
|---|---|---|---|---|
| Casein | 200 | 200 | 200 | 200 |
| DL-methionine | 3 | 3 | 3 | 3 |
| Corn starch | 150 | 111 | 111 | 111 |
| Sucrose | 500 | 370 | 370 | 370 |
| Cellulose | 50 | 50 | 50 | 50 |
| Corn Oil | 50 | 30 | 30 | 30 |
| Lard | - | 170 | - | - |
| Rice bran oil | - | - | 170 | - |
| Palm oil | - | - | - | 170 |
| Mineral mix S10001 | 35 | 42 | 42 | 42 |
| Vitamin mix V1001 | 10 | 12 | 12 | 12 |
| Choline Bitartrate | 2 | 2 | 2 | 2 |
| Gene | Primer Sequence |
|---|---|
| β-actin | Forward, AGGCCCAGAGCAAGAGAG Reverse, GGGTGTTGAAGGTCTCAAAC |
| ppar-γ | Forward, TTTTCAAGGGTGCCAGTTTCAATCC Reverse, AATCCTTGGCCCTCTGAGAT |
| srebp-1c | Forward, AATGGTCCAGGCAAGTTCGT Reverse, TCCCTCTCAGCTGTGGTGGTGAA |
| f4/80 | Forward, AAAGACTGGATTCTGGGAAGTTTGG Reverse, CGAGAGTGTTGTGGCAGGTTG |
| inos | Forward, CAGAGGACCCAGAGACAAGC Reverse, TGCTGAAACATTTCCTGTGC |
| cox-2 | Forward, TTCAAAAGAAGTGCTGGAAAAGGT Reverse, GATCATCTCTACCTGAGTGTCTTT |
| arg1 | Forward, CTGGCAGTTGGAAGCATCTCT Reverse, GTGAGCATCCACCCAAATGAC |
| ym1 | Forward, ATCTATGCCTTTGCTGGAATGC Reverse, TGAATGAATATCTGACGGTTCTGAG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.; Yu, S.; Kim, W. Rice Bran Oil Attenuates Chronic Inflammation by Inducing M2 Macrophage Switching in High-Fat Diet-Fed Obese Mice. Foods 2021, 10, 359. https://doi.org/10.3390/foods10020359
Park H, Yu S, Kim W. Rice Bran Oil Attenuates Chronic Inflammation by Inducing M2 Macrophage Switching in High-Fat Diet-Fed Obese Mice. Foods. 2021; 10(2):359. https://doi.org/10.3390/foods10020359
Chicago/Turabian StylePark, Hyejeong, Seungmin Yu, and Wooki Kim. 2021. "Rice Bran Oil Attenuates Chronic Inflammation by Inducing M2 Macrophage Switching in High-Fat Diet-Fed Obese Mice" Foods 10, no. 2: 359. https://doi.org/10.3390/foods10020359
APA StylePark, H., Yu, S., & Kim, W. (2021). Rice Bran Oil Attenuates Chronic Inflammation by Inducing M2 Macrophage Switching in High-Fat Diet-Fed Obese Mice. Foods, 10(2), 359. https://doi.org/10.3390/foods10020359