
Feeding Algae Meal to Feedlot Lambs with Competent Reticular Groove Reflex Increases Omega-3 Fatty Acids in Meat

 , , and
, , and
Abstract
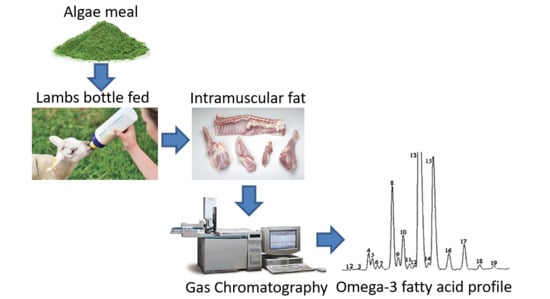
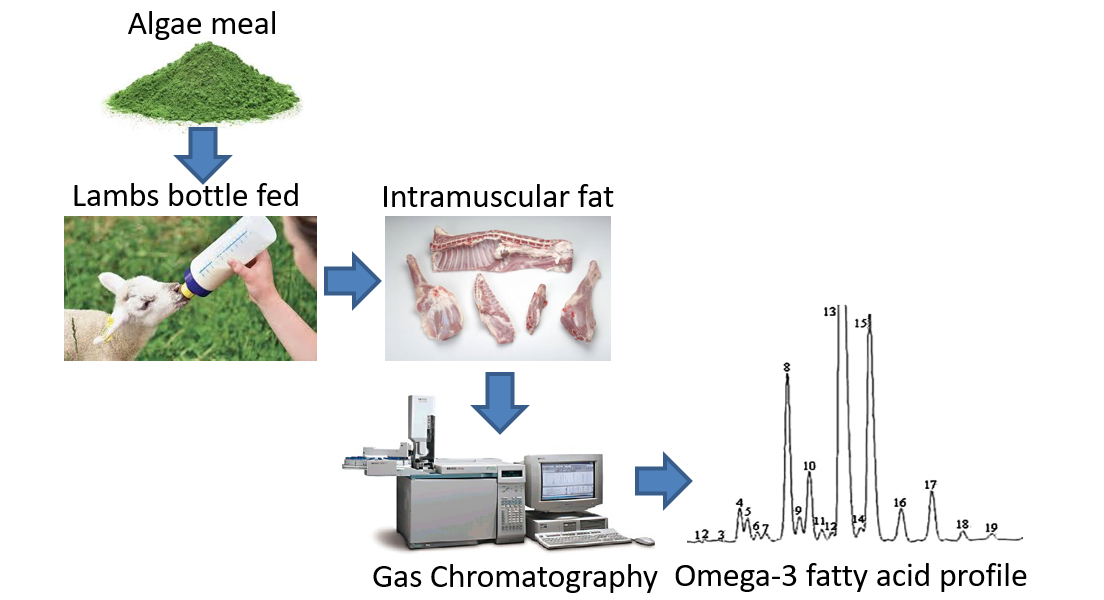
:
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Diets
2.2. Measurements, Sample Collection, and Laboratory Analyses
2.3. Statistical Analysis
3. Results and Discussion
3.1. General Results and Fatty Acid Profile
3.2. Saturated Fatty Acids in Lamb Meat
3.3. Monounsaturated Fatty Acids in Lamb Meat
3.4. Polyunsaturated Fatty Acids in Lamb Meat
3.5. Omega-3 Fatty Acids in Lamb Meat
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Calder, P.C. Very long-chain n-3 fatty acids and human health: Fact, fiction and the future. Proc. Nutr. Soc. 2018, 77, 52–72. [Google Scholar] [CrossRef] [Green Version]
- Shahidi, F.; Ambigaipalan, P. Omega-3 Polyunsaturated Fatty Acids and Their Health Benefits. Annu. Rev. Food Sci. Technol. 2018, 9, 345–381. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.J.; Miles, E.A.; Burdge, G.C.; Yaqoob, P.; Calder, P.C. Metabolism and functional effects of plant-derived omega-3 fatty acids in humans. Prog. Lipid Res. 2016, 64, 30–56. [Google Scholar] [CrossRef]
- Cholewski, M.; Tomczykowa, M.; Tomczyk, M. A comprehensive review of chemistry, sources and bioavailability of omega-3 fatty acids. Nutrients 2018, 10, 1662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chikwanha, O.C.; Vahmani, P.; Muchenje, V.; Dugan, M.E.; Mapiye, C. Nutritional enhancement of sheep meat fatty acid profile for human health and wellbeing. Food Res. Int. 2018, 104, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Kaic, A.; Mioc, B.; Kasap, A.; Potocnik, K. Meta-analysis of intramuscular fatty acid composition of Mediterranean lambs. Arch. Anim. Breed. 2016, 59, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Ponnampalam, E.N.; Sinclair, A.J.; Egan, A.R.; Blakeley, S.J.; Li, D.; Leury, B.J. Effect of dietary modification of muscle long-chain n-3 fatty acid on plasma insulin and lipid metabolites, carcass traits, and fat deposition in lambs. J. Anim. Sci. 2001, 79, 895–903. [Google Scholar] [CrossRef]
- Meale, S.J.; Chaves, A.V.; He, M.L.; McAllister, T.A. Dose-response of supplementing marine algae (Schizochytrium spp.) on production performance, fatty acid profiles, and wool parameters of growing lambs. J. Anim. Sci. 2014, 92, 2202–2213. [Google Scholar] [CrossRef] [PubMed]
- Urrutia, O.; Mendizabal, J.A.; Insausti, K.; Soret, B.; Purroy, A.; Arana, A. Effects of addition of linseed and marine algae to the diet on adipose tissue development, fatty acid profile, lipogenic gene expression, and meat quality in lambs. PLoS ONE 2016, 11, e0156765. [Google Scholar] [CrossRef] [Green Version]
- Díaz, M.T.; Pérez, C.; Sánchez, C.; Lauzurica, S.; Cañeque, V.; González, C.; De la Fuente, J. Feeding microalgae increases omega 3 fatty acids of fat deposits and muscles in light lambs. J. Food Comp. Anal. 2017, 56, 115–123. [Google Scholar] [CrossRef]
- Andrés, S.; Morán, L.; Aldai, N.; Tejido, M.L.; Prieto, N.; Bodas, R.; Giráldez, F.J. Effects of linseed and quercetin added to the diet of fattening lambs on the fatty acid profile and lipid antioxidant status of meat samples. Meat Sci. 2014, 97, 156–163. [Google Scholar] [CrossRef]
- Nguyen, D.V.; Flakemore, A.R.; Otto, J.; Ives, S.W.; Smith, R.W.; Nichols, P.D.; Malau-Aduli, A.E.O. Nutritional value and sensory characteristics of meat eating quality of Australian prime lambs supplemented with pelleted canola and flaxseed oils: Fatty acid profiles of muscle and adipose tissues. Intern. Med. Rev. 2017, 3, 1–21. [Google Scholar]
- Bessa, R.J.B.; Alves, S.P.; Santos-Silva, J. Constraints and potentials for the nutritional modulation of the fatty acid composition of ruminant meat. Eur. J. Lipid Sci. Technol. 2015, 117, 1325–1344. [Google Scholar] [CrossRef]
- Ørskov, E.R.; Benzie, D. Studies on the oesophageal groove reflex in sheep and on the potential use of the groove to prevent the fermentation of food in the rumen. Br. J. Nutr. 1969, 23, 415–420. [Google Scholar] [CrossRef] [Green Version]
- Dobarganes García, C.; Pérez Hernández, M.; Cantalapiedra, G.; Salas, J.M.; Merino, J.A. Bypassing the rumen in dairy ewes: The reticular groove reflex vs. calcium soap of olive fatty acids. J. Dairy Sci. 2005, 88, 741–747. [Google Scholar] [CrossRef] [Green Version]
- Martínez Marín, A.L.; Gómez-Cortés, P.; Carrión Pardo, D.; Núñez Sánchez, N.; Gómez Castro, G.; Juárez, M.; Pérez Alba, L.; Pérez Hernández, M.; De la Fuente, M.A. Feeding linseed oil to dairy goats with competent reticular groove reflex greatly increases n-3 fatty acids in milk fat. J. Dairy Sci. 2013, 96, 7532–7537. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, M.J.; Hopkins, S.P.; Kealy, J.K. The functioning of the oesophageal groove reflex and comparison of the performance of lambs fed individually or in groups. Br. J. Nutr. 1971, 26, 439–449. [Google Scholar] [CrossRef]
- Saini, R.K.; Keum, Y.S. Omega-3 and omega-6 polyunsaturated fatty acids: Dietary sources, metabolism, and significance—A review. Life Sci. 2018, 203, 255–267. [Google Scholar] [CrossRef]
- Hopkins, D.L.; Clayton, E.H.; Lamb, T.A.; Van de Ven, R.J.; Refshauge, G.; Kerr, M.J.; Bailes, K.; Lewandoski, P.; Ponnampalam, E.N. The impact of supplementing lambs with algae on growth, meat traits and oxidative status. Meat Sci. 2014, 98, 135–141. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Burnett, V.F.; Norng, S.; Hopkins, D.L.; Plozza, T.; Jacobs, J.L. Muscle antioxidant (vitamin E) and major fatty acid groups, lipid oxidation and retail colour of meat from lambs fed a roughage based diet with flaxseed or algae. Meat Sci. 2016, 111, 154–160. [Google Scholar] [CrossRef]
- Bligh, E.G.; Dyer, W.J. A rapid method of total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Santercole, V.; Mazzette, R.; De Santis, E.P.L.; Banni, S.; Goonewardene, L.; Kramer, J.K.G. Total Lipids of Sarda sheep meat that include the fatty acid and alkenyl composition and the CLA and trans-18:1 isomers. Lipids 2007, 42, 361–382. [Google Scholar] [CrossRef]
- Esquivelzeta, C.; Casellas, J.; Fina, M.; Campo, M.D.M.; Piedrafita, J. Carcass traits and meat fatty acid composition in Mediterranean light lambs. Can. J. Anim. Sci. 2017, 97, 734–741. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Cortés, P.; Galisteo, O.O.; Avilés Ramírez, C.; Peña Blanco, F.; de la Fuente, M.A.; Núñez Sánchez, N.; Martínez Marín, A.L. Intramuscular fatty acid profile of feedlot lambs fed concentrates with alternative ingredients. Anim. Prod. Sci. 2019, 59, 914–920. [Google Scholar] [CrossRef]
- Pérez-Palacios, T.; Ruiz, J.; Martín, D.; Muriel, E.; Antequera, T. Comparison of different methods for total lipid quantification in meat and meat products. Food Chem. 2008, 110, 1025–1029. [Google Scholar] [CrossRef]
- Bond, L.M.; Miyazaki, M.; O’Neill, L.M.; Ding, F.; Ntambi, J.M. Fatty acid desaturation and elongation in mammals. In Biochemistry of Lipids Lipoproteins and Membranes; Ridgway, N.D., McLeod, R.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 185–208. [Google Scholar]
- Ferreira, E.M.; Pires, A.V.; Susin, I.; Gentil, R.S.; Parente, M.O.M.; Nolli, C.P.; Meneghini, R.C.M.; Mendes, C.Q.; Ribeiro, C.V.D.M. Growth, feed intake, carcass characteristics, and meat fatty acid profile of lambs fed soybean oil partially replaced by fish oil blend. Anim. Feed Sci. Technol. 2014, 187, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Cortés, P.; Juárez, M.; de la Fuente, M.A. Milk fatty acids and potential health benefits: An updated vision. Trends Food Sci. Technol. 2018, 81, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.; Alves, S.P.; Francisco, A.; Almeida, J.; Alfaia, C.M.; Martins, S.V.; Prates, J.A.M.; Santos-Silva, J.; Doran, O.; Bessa, R.J.B. The reduction of starch in finishing diets supplemented with oil does not prevent the accumulation of trans-10 18: 1 in lamb meat. J. Anim. Sci. 2017, 95, 3745–3761. [Google Scholar] [CrossRef] [Green Version]
- Santos-Silva, J.; Francisco, A.; Alves, S.P.; Portugal, P.; Dentinho, T.; Almeida, J.; Soldado, D.; Jerónimo, E.; Bessa, R.J.B. Effect of dietary neutral detergent fibre source on lambs growth, meat quality and biohydrogenation intermediates. Meat Sci. 2019, 147, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Bessa, R.J.B.; Alves, S.P.; Jerónimo, E.; Alfaia, C.M.; Prates, J.A.; Santos-Silva, J. Effect of lipid supplements on ruminal biohydrogenation intermediates and muscle fatty acids in lambs. Eur. J. Lipid Sci. Technol. 2007, 109, 868–878. [Google Scholar] [CrossRef]
- Altomonte, I.; Salaria, F.; Licitra, R.; Martini, M. Use of microalgae in ruminant nutrition and implications on milk quality-A review. Livest. Sci. 2018, 214, 25–35. [Google Scholar] [CrossRef]
- Cooper, S.L.; Sinclair, L.A.; Wilkinson, R.G.; Hallett, K.G.; Enser, M.; Wood, J.D. Manipulation of the n-3 polyunsaturated fatty acid content of muscle and adipose tissue in lambs. J. Anim. Sci. 2004, 82, 1461–1470. [Google Scholar] [CrossRef]
- Alvarenga, T.I.R.C.; Chen, Y.Z.; Lewandowski, P.; Ponnampalam, E.N.; Sadiq, S.; Clayton, E.H.; Van de Ven, R.J.; Perez, J.R.O.; Hopkins, D.L. The expression of genes encoding enzymes regulating fat metabolism is affected by maternal nutrition when lambs are fed algae high in omega-3. Livest. Sci. 2016, 187, 53–60. [Google Scholar] [CrossRef]
- EFSA. Scientific opinion on the tolerable upper intake level of eicosapentaenoic acid (EPA), docosahexaenoic acid (DHA) and docosapentaenoic acid (DPA). EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). EFSA J. 2012, 10, 2815. [Google Scholar]
- EU. Amending Regulation (EC) No 1924/2006 of the European Parliament and of the Council with regard to the list of nutrition claims. Commission Regulation (EU) No 116/2010 of 10 February 2010. Off. J. Eur. Union 2010, 404, L37/16–L37/19. [Google Scholar]
- Alvarenga, T.I.R.C.; Chen, Y.; Furusho-Garcia, I.F.; Pérez, J.R.O.; Hopkins, D.L. Manipulation of omega-3 PUFAs in lamb: Phenotypic and genotypic views. Compr. Rev. Food Sci. Food Saf. 2015, 14, 189–204. [Google Scholar] [CrossRef]
- Drouin, G.; Rioux, V.; Legrand, P. The n-3 docosapentaenoic acid (DPA): A new player in the n-3 long chain polyunsaturated fatty acid family. Biochimie 2019, 159, 36–48. [Google Scholar] [CrossRef]
- Nguyen, D.V.; Malau-Aduli, B.S.; Cavalieri, J.; Nichols, P.D.; Malau-Aduli, A.E.O. Supplementation with plant-derived oils rich in omega-3 polyunsaturated fatty acids for lamb production. Vet. Anim. Sci. 2018, 6, 29–40. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Evolutionary aspects of the dietary omega-6/omega-3 fatty acid ratio: Medical implications. In Evolutionary Thinking in Medicine. Advances in the Evolutionary Analysis of Human Behaviour; Alvergne, A., Jenkinson, C., Faurie, C., Eds.; Springer: Berlin/Heidelberg, Germany, 2016; pp. 119–134. [Google Scholar]
- Chen, J.; Liu, H. Nutritional indices for assessing fatty acids: A Mini-Review. Int. J. Mol. Sci. 2020, 21, 5695. [Google Scholar] [CrossRef]

{kind=link}
| Fatty Acid | Control Concentrate | Algae Meal | Milk Replacer |
|---|---|---|---|
| 8:0 | 1.36 | ||
| 10:0 | 1.34 | ||
| 12:0 | 16.23 | ||
| 14:0 | 0.26 | 7.17 | 5.84 |
| 15:0 | 1.52 | ||
| 16:0 | 15.33 | 58.80 | 27.98 |
| 16:1 | 0.26 | ||
| 17:0 | 0.38 | ||
| 18:0 | 2.22 | 1.38 | 3.82 |
| c9 18:1 | 22.91 | 30.48 | |
| 18:2 omega-6 LA | 51.64 | 11.12 | |
| 18:3 omega-3 ALA | 4.18 | ||
| 20:0 | 0.44 | ||
| 20:4 omega-6 AA | 0.98 | ||
| 20:5 omega-3 EPA | 0.36 | ||
| 22:0 | 0.23 | ||
| 22:5 omega-6 | 3.24 | ||
| 22:5 omega-3 DPA | 0.32 | ||
| 22:6 omega-3 DHA | 22.66 | ||
| TOTAL SFA | 18.48 | 69.51 | 56.57 |
| TOTAL MUFA | 22.91 | 30.48 | |
| TOTAL PUFA | 55.82 | 27.56 | 11.12 |
| TOTAL omega-3 | 4.18 | 23.34 | |
| TOTAL omega-6 | 51.64 | 4.22 | 11.12 |
| Fatty Acids | NOALG | ALGCON | ALGMILK | SEM | p |
|---|---|---|---|---|---|
| 10:0 | 0.11 b | 0.15 a | 0.14 ab | 0.006 | <0.05 |
| 12:0 | 0.08 b | 0.15 a | 0.17 a | 0.012 | <0.01 |
| 13:0 | 0.02 | 0.02 | 0.02 | 0.001 | 0.87 |
| 14:0 | 1.31 b | 1.91 a | 1.81 a | 0.083 | <0.01 |
| 15:0 | 0.42 | 0.36 | 0.37 | 0.019 | 0.36 |
| 16:0 | 16.51 b | 22.04 a | 20.87 a | 0.463 | <0.001 |
| 17:0 | 2.41 a | 1.69 b | 1.71 b | 0.116 | <0.05 |
| 18:0 | 13.88 | 12.32 | 13.36 | 0.266 | 0.06 |
| 20:0 | 0.08 | 0.07 | 0.08 | 0.002 | 0.20 |
| 21:0 | 0.05 a | 0.02 b | 0.02 b | 0.003 | <0.001 |
| 22:0 | 0.11 a | 0.06 b | 0.06 b | 0.005 | <0.001 |
| 23:0 | 0.04 | 0.03 | 0.04 | 0.006 | 0.48 |
| 24:0 | 0.05 c | 0.09 b | 0.11 a | 0.006 | <0.001 |
| TOTAL non-branched SFA | 35.09 b | 38.89 a | 38.76 a | 0.490 | <0.001 |
| iso 15:0 | 0.05 | 0.05 | 0.05 | 0.002 | 0.69 |
| anteiso 15:0 | 0.12 | 0.10 | 0.10 | 0.003 | 0.09 |
| Other branched 15:0 | 0.12 a | 0.06 b | 0.06 b | 0.011 | <0.05 |
| iso 16:0 | 0.10 | 0.09 | 0.08 | 0.003 | 0.09 |
| Other branched 16:0 | 0.47 a | 0.30 b | 0.31 b | 0.026 | <0.05 |
| iso 17:0 | 0.16 ab | 0.13 b | 0.17 a | 0.005 | <0.01 |
| anteiso 17:0 | 0.67 a | 0.51 b | 0.51 b | 0.017 | <0.001 |
| iso 18:0 | 0.16 | 0.14 | 0.13 | 0.007 | 0.17 |
| TOTAL branched SFA | 1.85 a | 1.39 b | 1.42 b | 0.056 | <0.01 |
| TOTAL SFA | 36.94 b | 40.28 a | 40.18 a | 0.490 | <0.01 |
| 14:0 desaturation index 1 | 0.04 a | 0.03 b | 0.03 b | 0.001 | <0.001 |
| 16:0 desaturation index 2 | 0.06 a | 0.04 b | 0.04 b | 0.002 | <0.001 |
| 18:0 desaturation index 3 | 0.65 | 0.68 | 0.66 | 0.005 | 0.07 |
| 16:0 elongation index 4 | 0.69 a | 0.62 b | 0.64 b | 0.007 | <0.001 |
| Fatty Acids | NOALG | ALGCON | ALGMILK | SEM | p |
|---|---|---|---|---|---|
| c9 14:1 | 0.05 | 0.05 | 0.05 | 0.002 | 0.60 |
| c7 16:1 | 0.18 a | 0.15 b | 0.16 ab | 0.005 | <0.05 |
| c9 16:1 | 1.00 | 0.98 | 0.84 | 0.033 | 0.10 |
| c9 18:1 + (c10+t13+t14) 18:1 | 25.68 | 25.82 | 26.21 | 0.484 | 0.86 |
| c11 18:1 + t15 18:1 | 3.20 a | 2.58 b | 2.66 b | 0.065 | <0.001 |
| c12 18:1 | 0.25 ab | 0.18 b | 0.30 a | 0.015 | <0.01 |
| c13 18:1 | 0.14 a | 0.06 b | 0.04 b | 0.007 | <0.001 |
| c16 18:1 | 0.02 | 0.02 | 0.02 | 0.002 | 0.60 |
| 20:1 | 0.03 a | 0.03 a | 0.02 b | 0.001 | <0.001 |
| c8 20:1 | 0.02 a | 0.01 b | 0.01 b | 0.001 | <0.001 |
| c11 20:1 | 0.13 | 0.12 | 0.10 | 0.006 | 0.08 |
| 22:1 | 0.04 c | 0.08 b | 0.11 a | 0.006 | <0.001 |
| c15 24:1 | 0.02 | 0.02 | 0.02 | 0.003 | 0.84 |
| TOTAL cis MUFA | 30.73 | 30.11 | 30.52 | 0.484 | 0.86 |
| t8 16:1 | 0.08 a | 0.09 a | 0.06 b | 0.004 | <0.001 |
| t9 16:1 | 0.12 | 0.13 | 0.14 | 0.006 | 0.73 |
| t5 18:1 | 0.02 | 0.02 | 0.02 | 0.001 | 0.16 |
| t(6+7+8) 18:1 | 0.15 a | 0.05 b | 0.13 a | 0.011 | <0.001 |
| t10 18:1 | 3.94 b | 6.02 a | 2.71 b | 0.488 | <0.001 |
| t11 18:1 | 2.25 | 1.95 | 2.06 | 0.222 | 0.70 |
| TOTAL trans MUFA | 7.42 ab | 7.75 a | 6.08 b | 0.266 | <0.05 |
| TOTAL MUFA | 38.15 | 37.86 | 36.60 | 0.592 | 0.50 |
| Fatty Acids | NOALG | ALGCON | ALGMILK | SEM | p |
|---|---|---|---|---|---|
| t9 t12 18:2 | 0.02 b | 0.03 a | 0.01c | 0.001 | <0.001 |
| c9 t13 + t8 c12 18:2 | 0.06 | 0.05 | 0.07 | 0.004 | 0.14 |
| c9 t12 + t8 c13 18:2 | 0.04 | 0.03 | 0.05 | 0.003 | 0.13 |
| t9 c12 18:2 | 0.02 | 0.02 | 0.02 | 0.001 | 0.21 |
| t11 c15 18:2 | 0.16 ab | 0.20 a | 0.12 b | 0.010 | <0.01 |
| c9 c12 18:2 LA | 10.64 | 8.80 | 8.83 | 0.344 | 0.06 |
| c9 c15 18:2 | 0.28 a | 0.19 b | 0.18 b | 0.010 | <0.001 |
| TOTAL non-conjugated 18:2 | 11.22 | 9.34 | 9.27 | 0.353 | 0.06 |
| c9 t11 18:2 RA | 0.09 | 0.09 | 0.10 | 0.022 | 0.93 |
| t9 c11 18:2 | 0.09 a | 0.06 b | 0.05 b | 0.004 | <0.001 |
| t10 c12 18:2 | 0.06 a | 0.03 b | 0.02 b | 0.003 | <0.001 |
| TOTAL conjugated 18:2 | 0.24 | 0.17 | 0.27 | 0.022 | 0.11 |
| TOTAL 18:2 | 11.46 | 8.93 | 9.54 | 0.402 | 0.06 |
| 18:3 omega-6 | 0.12 a | 0.06 b | 0.08 b | 0.005 | <0.001 |
| 18:3 omega-3 ALA | 0.36 | 0.32 | 0.32 | 0.009 | 0.24 |
| 20:2 omega-6 | 0.08 | 0.07 | 0.07 | 0.003 | 0.06 |
| 20:3 omega-3 | 0.33 a | 0.16 b | 0.18 b | 0.016 | <0.001 |
| 20:3 omega-6 | 0.32 a | 0.25 b | 0.29 ab | 0.015 | 0.06 |
| 20:4 omega-6 AA | 4.57 a | 2.82 b | 2.99 b | 0.242 | <0.01 |
| 20:5 omega-3 EPA | 0.20 c | 0.86 b | 1.27 a | 0.085 | <0.001 |
| 22:3 omega-3 | 0.04 | 0.04 | 0.04 | 0.002 | 0.55 |
| 22:4 omega-6 | 0.56 a | 0.16 b | 0.13 b | 0.034 | <0.001 |
| 22:5 omega-6 | 0.17 b | 0.25 a | 0.27 a | 0.014 | <0.01 |
| 22:5 omega-3 DPA | 0.42 b | 0.45 b | 0.62 a | 0.027 | <0.01 |
| 22:6 omega-3 DHA | 0.11c | 2.12 b | 2.54 a | 0.182 | <0.001 |
| TOTAL PUFA | 18.77 | 16.49 | 18.31 | 0.722 | 0.41 |
| TOTAL omega-3 | 1.45c | 3.94 b | 4.96 a | 0.274 | <0.001 |
| TOTAL omega-6 | 16.46 a | 12.41 b | 12.64 b | 0.626 | <0.05 |
| c9 c12 18:2 elongation index 1 | 0.29 a | 0.24 b | 0.25 b | 0.007 | <0.01 |
| Index | NOALG | ALGCON | ALGMILK | SEM | p |
|---|---|---|---|---|---|
| omega-6/omega-3 | 11.27 a | 3.16 b | 2.71 b | 0.581 | <0.001 |
| PUFA/SFA | 0.53 | 0.41 | 0.46 | 0.024 | 0.15 |
| h/H 1 | 2.63 a | 1.88 b | 2.06 b | 0.066 | <0.001 |
| AI 2 | 0.39 b | 0.56 a | 0.52 a | 0.018 | <0.001 |
| TI 3 | 1.00 | 0.98 | 0.92 | 0.023 | 0.22 |
| PI 4 | 41.26 b | 49.83 ab | 57.99 a | 2.441 | <0.01 |
| DFA 5 | 70.70 a | 66.66 b | 68.27 ab | 0.499 | <0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez-Cortés, P.; de la Fuente, M.A.; Peña Blanco, F.; Núñez-Sánchez, N.; Requena Domenech, F.; Martínez Marín, A.L. Feeding Algae Meal to Feedlot Lambs with Competent Reticular Groove Reflex Increases Omega-3 Fatty Acids in Meat. Foods 2021, 10, 366. https://doi.org/10.3390/foods10020366
Gómez-Cortés P, de la Fuente MA, Peña Blanco F, Núñez-Sánchez N, Requena Domenech F, Martínez Marín AL. Feeding Algae Meal to Feedlot Lambs with Competent Reticular Groove Reflex Increases Omega-3 Fatty Acids in Meat. Foods. 2021; 10(2):366. https://doi.org/10.3390/foods10020366
Chicago/Turabian StyleGómez-Cortés, Pilar, Miguel Angel de la Fuente, Francisco Peña Blanco, Nieves Núñez-Sánchez, Francisco Requena Domenech, and Andrés L. Martínez Marín. 2021. "Feeding Algae Meal to Feedlot Lambs with Competent Reticular Groove Reflex Increases Omega-3 Fatty Acids in Meat" Foods 10, no. 2: 366. https://doi.org/10.3390/foods10020366
APA StyleGómez-Cortés, P., de la Fuente, M. A., Peña Blanco, F., Núñez-Sánchez, N., Requena Domenech, F., & Martínez Marín, A. L. (2021). Feeding Algae Meal to Feedlot Lambs with Competent Reticular Groove Reflex Increases Omega-3 Fatty Acids in Meat. Foods, 10(2), 366. https://doi.org/10.3390/foods10020366