
Dietary Regulation of the Crosstalk between Gut Microbiome and Immune Response in Inflammatory Bowel Disease

 ,
, 

Abstract
:1. Introduction
2. Intestinal Microbiota and IBD
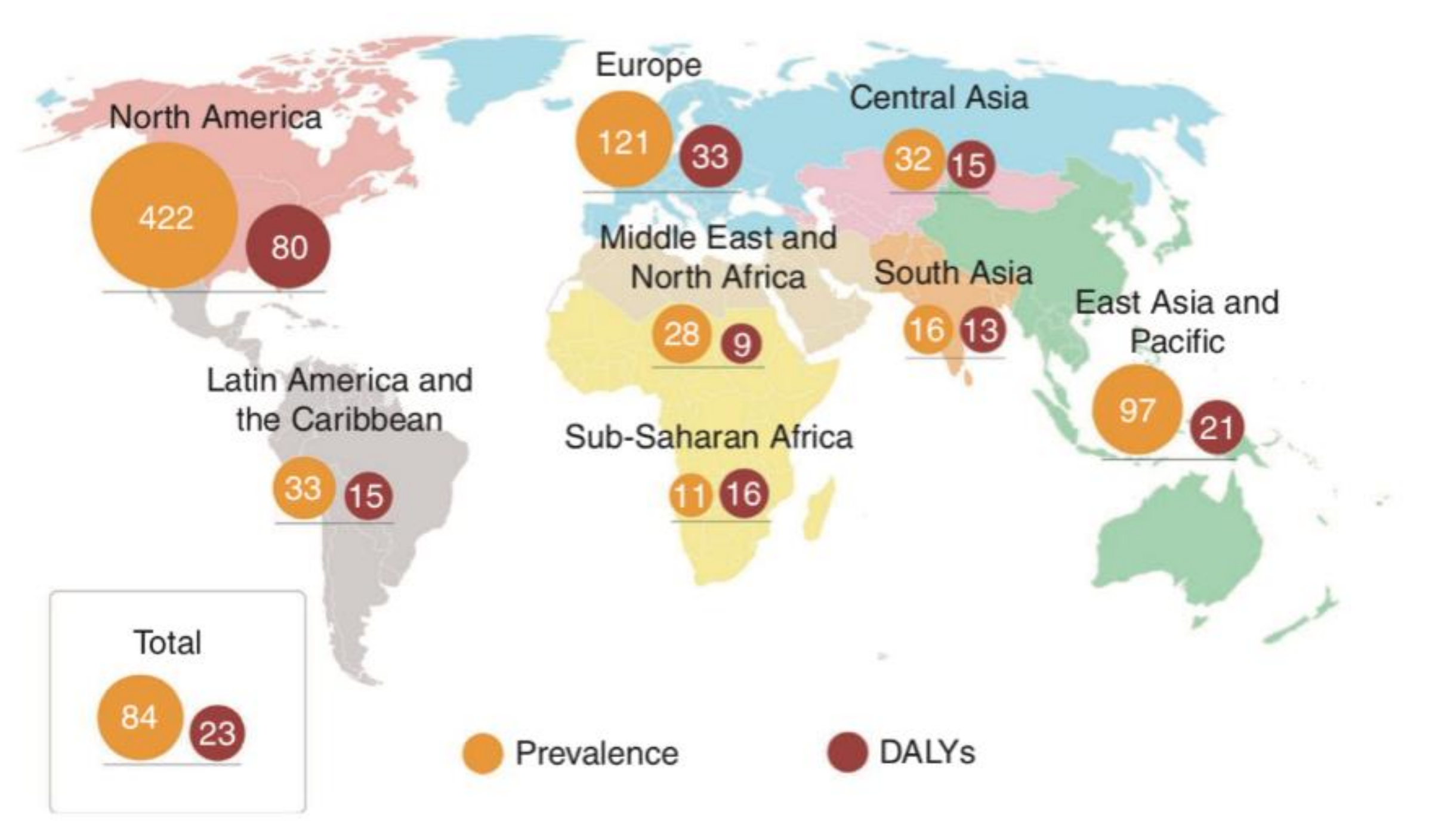
2.1. The Prevalence of IBD Worldwide
2.2. Dietary Nutrients and IBD: A Complex Interaction
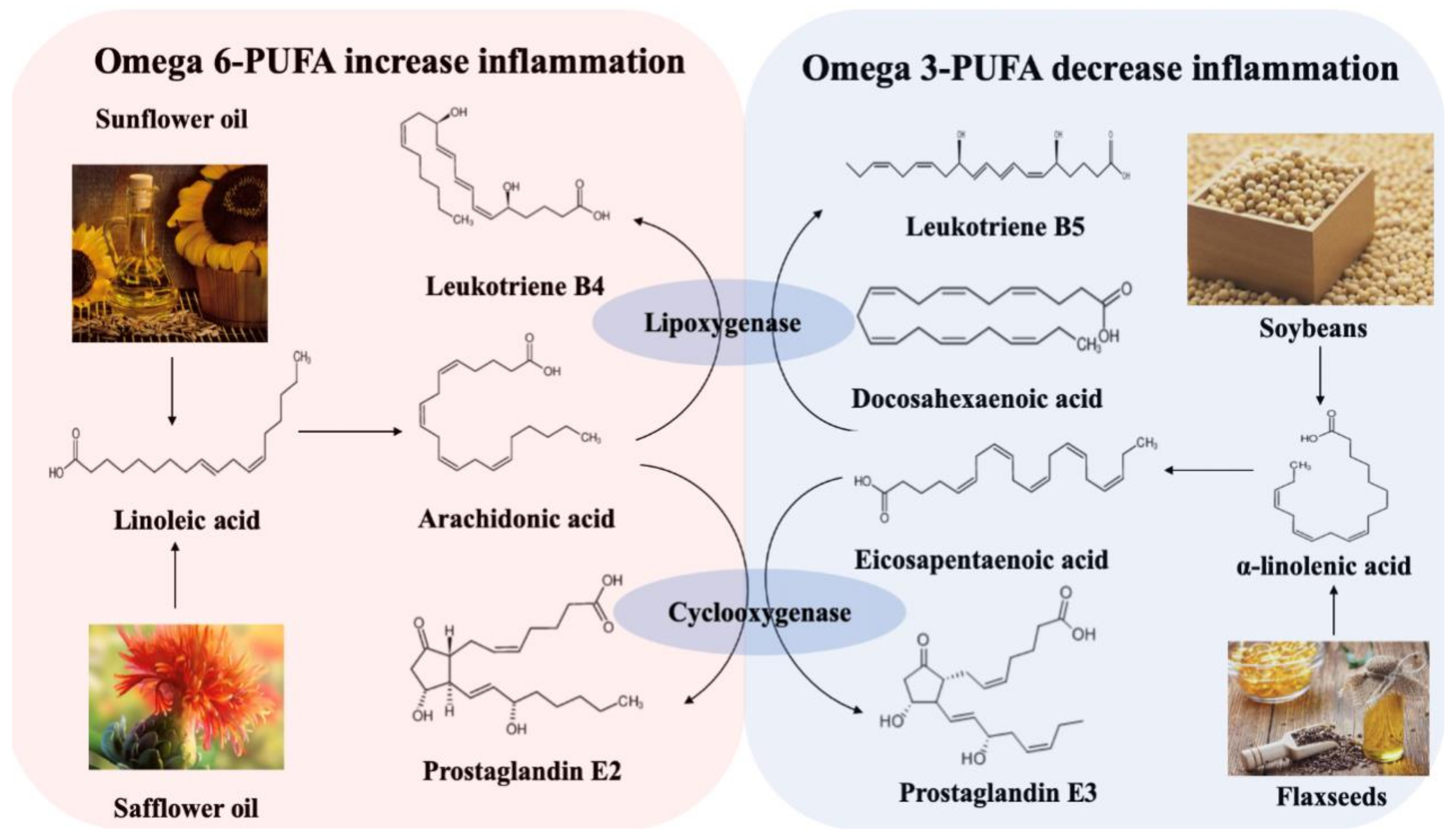
2.3. Dietary Fat
2.4. Dietary Fiber
2.5. Protein
2.6. Vitamin D
2.7. Other Nutrients
2.8. Dietary Nutrients and Microbiota
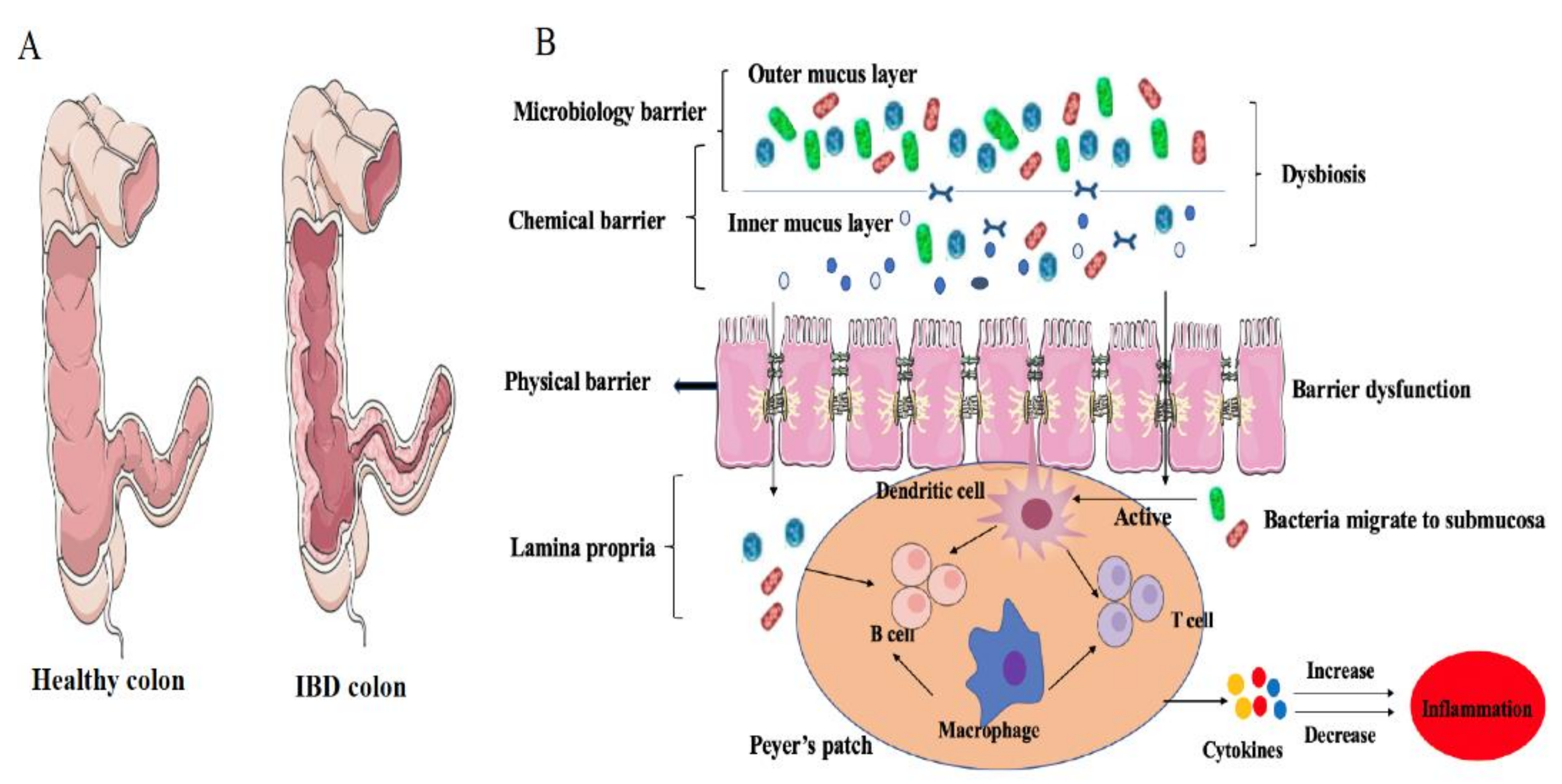
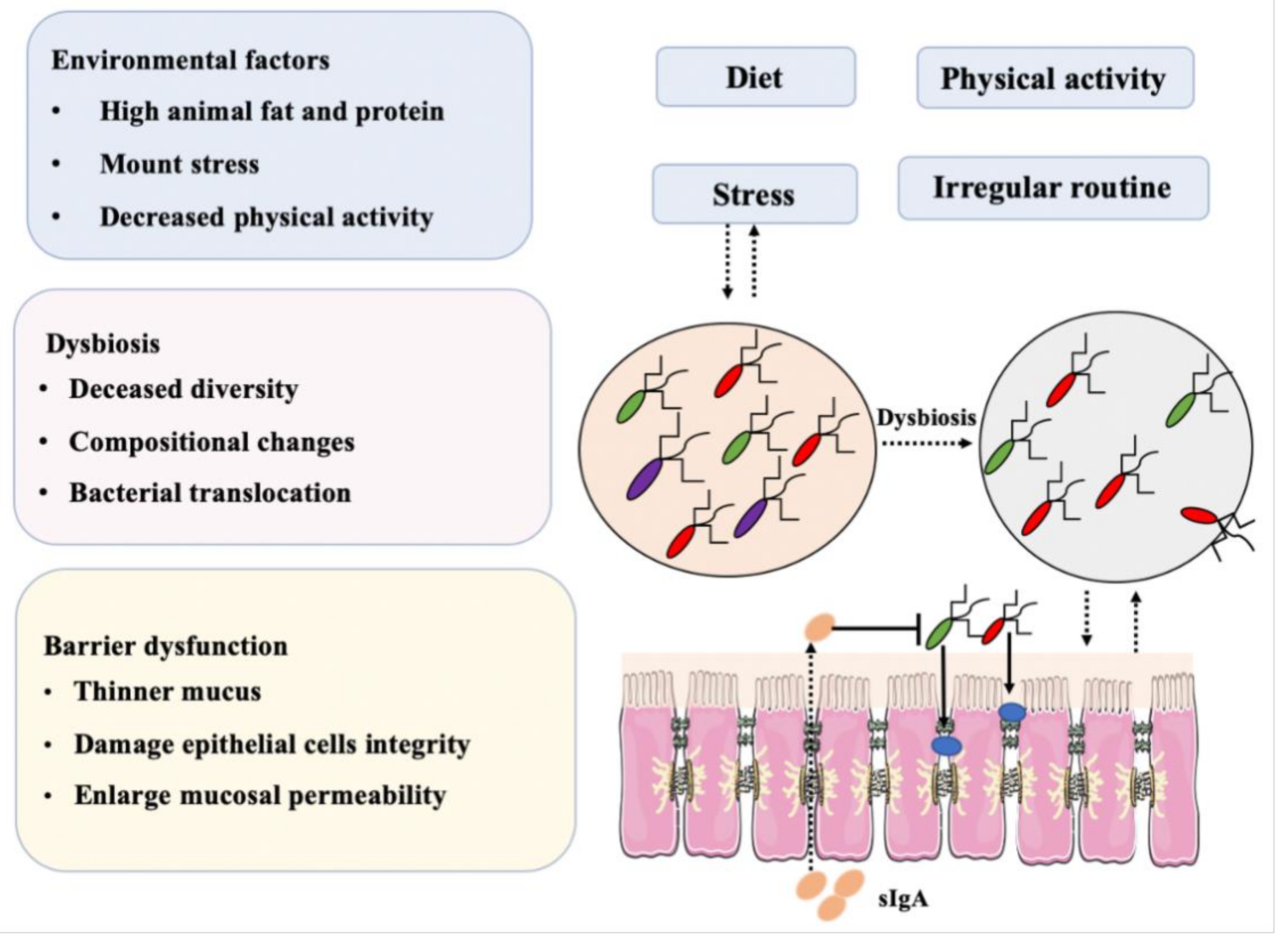
2.9. Gut Microbiota and IBD: Dysbiosis Is a Typical Feature in IBD
2.10. Gut Microbiome Dysbiosis Induces IBD: Role of Immunological Barrier
2.11. Microbiota and Intestinal Epithelial Cells
2.12. Migration of Mucosal Bacteria in IBD
3. GALT Dysfunction during IBD
4. Microbiome and GALT
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Sender, R.; Fuchs, S.; Milo, R. Are we really vastly outnumbered? Revisiting the ratio of bacterial to host cells in humans. Cell 2016, 164, 337–340. [Google Scholar] [CrossRef] [Green Version]
- Pistoli, S.; Smejkal, C.; McCartney, A.; Gibson, G.R. Differences in the fecal flora of healthy individuals and patients with irritable bowel syndrome, and in vitro effects of a symbiotic upon gut flora composition. Clin. Nutr. 2003, 22, S60. [Google Scholar] [CrossRef]
- Khalif, I.L.; Quigley, E.M.M.; Konovitch, E.A.; Maximova, I.D. Alterations in the colonic flora and intestinal permeability and evidence of immune activation in chronic constipation. Digest. Liver Dis. 2005, 37, 838–849. [Google Scholar] [CrossRef] [PubMed]
- Lochs, H. Interaction between nutrition, intestinal flora and the gastrointestinal immune system. Home Care Enter. Feed. 2005, 10, 179–188. [Google Scholar]
- Koutsos, E.A.; Arias, V.J. Intestinal ecology: Interactions among the gastrointestinal tract, nutrition, and the microflora. J. Appl. Poultry Res. 2006, 15, 161–173. [Google Scholar] [CrossRef]
- Ijssennagger, N.; Belzer, C.; Hooiveld, G.J.; Dekker, J.; van Mil, S.W.C.; Müller, M.; Kleerebezem, M.; van der Meer, R. Gut microbiota facilitates dietary heme-induced epithelial hyperproliferation by opening the mucus barrier in colon. Proc. Natl Acad. Sci. USA 2015, 112, 10038–10043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reinhardt, C.; Bergentall, M.; Greiner, T.U.; Schaffner, F.; Ostergren-Lundén, G.; Petersen, L.C.; Ruf, W.; Bäckhed, F. Tissue factor and PAR1 promote microbiota-induced intestinal vascular remodelling. Nature 2012, 483, 627–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischbach, M.A.; Segre, J.A. Signaling in host associated microbial communities. Cell 2016, 164, 1288–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yano, J.M.; Yu, K.; Donaldson, G.P.; Shastri, G.G.; Ann, P.; Ma, L.; Nagler, C.R.; Ismagilov, R.F.; Mazmanian, S.K.; Hsiao, E.Y. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 2015, 161, 264–276. [Google Scholar] [CrossRef] [Green Version]
- Cho, I.; Yamanishi, S.; Cox, L.; Methé, B.A.; Zavadil, J.; Li, K.; Gao, Z.; Mahana, D.; Raju, K.; Teitler, I.; et al. Antibiotics in early life alter the murine colonic microbiome and adiposity. Nature 2012, 488, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Li, C. Relationship among intestinal flora, high-fat diets, and hyperlipidemia. World Chin. J. Dig. 2013, 21, 1273. [Google Scholar] [CrossRef]
- Loftus, E.V., Jr. Clinical epidemiology of inflammatory bowel disease: Incidence, prevalence, and environmental influences. Gastroenterology 2014, 126, 1504–1517. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, A.; Benchimol, E.I.; Bernstein, C.N.; Bitton, A.; Feagan, B.; Panaccione, R.; Glasgow, K.W.; Fernandes, A.; Ghosh, S. Inflammatory bowel disease: A Canadian burden of illness review. Can. J. Gastroenterol. 2012, 26, 811–817. [Google Scholar] [CrossRef]
- Burisch, J.; Jess, T.; Martinato, M.; Lakatos, P.L. The burden of inflammatory bowel disease in Europe. J. Crohns. Colitis 2013, 7, 322–337. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, G.G. The global burden of IBD: From 2015 to 2025. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Piovani, D.; Danese, S.; Peyrin-Biroulet, L.; Bonovas, S. Inflammatory bowel disease: Estimates from the global burden of disease 2017 study. Aliment. Pharmacol. Ther. 2020, 51, 1–10. [Google Scholar] [CrossRef]
- Bilski, J.; Mazur-Bialy, A.; Brzozowski, B.; Magierowski, M.; Zahradnik-Bilska, J.; Wójcik, D.; Magierowska, K.; Kwiecien, S.; Mach, T.; Brzozowski, T. Can exercise affect the course of inflammatory bowel disease? Experimental and clinical evidence. Pharmacol. Rep. 2016, 68, 827–836. [Google Scholar] [CrossRef]
- Probert, C.S.; Jayanthi, V.; Hughes, A.O.; Thompson, J.R.; Wicks, A.C.; Mayberry, J.F. Prevalence and family risk of ulcerative colitis and Crohn’s disease: An epidemiological study among Europeans and south Asians in Leicestershire. Gut 1993, 34, 1547–1551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplan, G.G.; Bernstein, C.N.; Coward, S.; Bitton, A.; Murthy, S.K.; Nguyen, G.C.; Lee, K.; Cooke-Lauder, J.; Benchimol, E.I. The impact of inflammatory bowel disease in Canada 2018: Epidemiology. J. Can. Assoc. Gastroenterol. 2019, 2 (Suppl. 1), 6–16. [Google Scholar] [CrossRef] [Green Version]
- Schwermer, M.; Fetz, K.; Lngler, A.; Ostermann, T.; Zuzak, T.J. Complementary, alternative, integrative and dietary therapies for children with crohn’s disease--a systematic review. Complement. Ther. Med. 2020, 52, 102493. [Google Scholar] [CrossRef]
- Kathy, V.; Anne, S.L.; Kelcie, W.; Targownik, L.E.; Clove, H.; Graff, L.A.; Sexton, K.A.; Lix, L.M.; Michael, S.; Charles, N.B. Association between change in inflammatory aspects of diet and change in IBD-related inflammation and symptoms over 1 year: The manitoba living with IBD study. Inflamm. Bowel Dis. 2020, 27, 190–202. [Google Scholar]
- Hou, J.K.; Abraham, B.; El-Serag, H. Dietary intake and risk of developing inflammatory bowel disease: A systematic review of the literature. Am. J. Gastroenterol. 2011, 106, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Tjonneland, A.; Overvad, K.; Bergmann, M.M.; Nagel, G.; Linseisen, J.; Hallmans, G.; Palmqvist, R.; Sjodin, H.; Hagglund, G.; Berglund, G.; et al. Linoleic acid, a dietary n-6 polyunsaturated fatty acid, and the aetiology of ulcerative colitis: A nested case-control study within a European prospective cohort study. Gut 2009, 58, 1606–1611. [Google Scholar]
- Goldman, D.W.; Pickett, W.C.; Goetzl, E.J. Human neutrophil chemotactic and degranulating activities of leukotriene B5 (LTB5) derived from eicosapentaenoic acid. Biochem. Biophys. Res. Comm. 1983, 117, 282–288. [Google Scholar] [CrossRef]
- Heller, A.; Koch, T.; Schmeck, J.; van Ackern, K. Lipid mediators in inflammatory disorders. Drugs 1998, 55, 487–496. [Google Scholar] [CrossRef] [PubMed]
- Hou, J.K.; Sellin, J.H. Diet, nutrition and inflammatory bowel disease. Therapy 2010, 7, 179–189. [Google Scholar] [CrossRef]
- Bamba, T.; Shimoyama, T.; Sasaki, M.; Tsujikawa, T.; Fukuda, Y.; Koganei, K.; Hibi, T.; Iwao, Y.; Munakata, A.; Fukuda, S.; et al. Dietary fat attenuates the benefits of an elemental diet in active Crohn’s disease: A randomized, controlled trial. Eur. J. Gastroenterol. Hepatol. 2003, 15, 151–157. [Google Scholar] [CrossRef] [PubMed]
- Thies, F.; Miles, E.A.; Nebevon-Caron, G.; Powell, J.R.; Hurst, T.L.; Newsholme, E.A.; Calder, P.C. Influence of dietary supplementation with long-chain n-3 or n-6 polyunsaturated fatty acids on blood inflammatory cell populations and functions and on plasm soluble adhesion molecules in healthy adults. Lipids 2001, 36, 1183–1193. [Google Scholar] [CrossRef]
- Rodríguez-Cabezas, M.E.; Gálvez, J.; Lorente, M.D.; Concha, A.; Camuesco, D.; Azzouz, S.; Osuna, A.; Redondo, L.; Zarzuelo, A. Dietary fiber down-regulates colonic tumor necrosis factor alpha and nitric oxide production in trinitrobenzenesulfonic acid-induced colitic rats. J. Nutr. 2002, 132, 3263–3271. [Google Scholar] [CrossRef]
- Liu, X.; Wu, Y.; Li, F.; Zhang, D. Dietary fiber intake reduces risk of inflammatory bowel disease: Result from a meta-analysis. Nutr. Res. 2015, 35, 753–758. [Google Scholar] [CrossRef]
- Venkatraman, A.; Ramakrishna, B.S.; Shaji, R.V.; Kumar, N.S.; Pulimood, A.; Patra, S. Amelioration of dextran sulfate colitis by butyrate: Role of heat shock protein 70 and NF-kappaB. Am. J. Physiol. Gastrointest. Liver Physiol. 2003, 285, 177–184. [Google Scholar] [CrossRef] [Green Version]
- Fiocchi, C. IBD: Advances in pathogenesis, complications, diagnosis, and therapy. Curr. Opin. Gastroenterol. 2012, 28, 297–300. [Google Scholar] [CrossRef] [PubMed]
- King, D.E.; Egan, B.M.; Geesey, M.E. Relation of dietary fat and fiber to elevation of C-reactive protein. Am. J. Cardiol. 2003, 92, 1335–1339. [Google Scholar] [CrossRef] [PubMed]
- Deehan, E.C.; Yang, C.; Perez-Munoz, M.E.; Nguyen, N.; Cheng, C.C.; Triador, L.; Zhang, Z.X.; Bakal, J.A.; Walter, J. Precision microbiome modulation with discrete dietary fiber structures directs short-chain fatty acid production. Cell Host Microbe 2020, 27, 389–404. [Google Scholar] [CrossRef]
- Tannock, G.W.; Liu, Y. Guided dietary fibre intake as a means of directing short-chain fatty acid production by the gut microbiota. J. R. Soc. N. Z. 2019, 2, 1–22. [Google Scholar] [CrossRef]
- Reif, S.; Klei, I.; Lubin, F.; Farbstein, M.; Hallak, A.; Gilat, T. Pre-illness dietary factors in inflammatory bowel disease. Gut 1997, 40, 754–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jantchou, P.; Morois, S.; Clavel-Chapelon, F.; Boutron-Ruault, M.C.; Carbonnel, F. Animal protein intake and risk of inflammatory bowel disease: The E3N prospective study. Am. J. Gastroenterol. 2010, 105, 2195–2201. [Google Scholar] [CrossRef] [PubMed]
- Shoda, R.; Matsueda, K.; Yamato, S.; Umeda, N. Epidemiologic analysis of Crohn disease in Japan: Increased dietary intake of n-6 polyunsaturated fatty acids and animal protein relates to the increased incidence of Crohn disease in Japan. Am. J. Clin. Nutr. 1996, 63, 741–745. [Google Scholar] [CrossRef] [PubMed]
- Spooren, C.E.G.M.; Pierik, M.J.; Zeegers, M.P.; Feskens, E.J.M.; Masclee, A.A.M.; Jonkers, D.M.A.E. Review article: The association of diet with onset and relapse in patients with inflammatory bowel disease. Aliment. Pharmacol. Ther. 2013, 38, 1172–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holick, M.F. Vitamin D deficiency. N. Engl. J. Med. 2007, 357, 266–281. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; Zhu, J.G.; DeLuca, H.F. Where is the vitamin D receptor? Arch. Biochem. Biophys. 2012, 523, 123–133. [Google Scholar] [CrossRef] [PubMed]
- Mouli, V.P.; Ananthakrishnan, A.N. Vitamin D and inflammatory bowel diseases. Aliment. Pharmacol. Ther. 2014, 39, 125–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ananthakrishnan, A.N.; Cagan, A.; Gainer, V.S.; Cai, T.; Cheng, S.C.; Savova, G.; Chen, P.; Szolovits, P.; Xia, Z.; De Jager, P.L.; et al. Normalization of plasma 25-hydroxy vitamin D is associated with reduced risk of surgery in Crohn’s disease. Inflamm. Bowel Dis. 2013, 19, 1921–1927. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, S.P.; Hvas, C.L.; Agnholt, J.; Christensen, L.A.; Heickendorff, L.; Dahlerup, J.F. Active Crohn’s disease is associated with low vitamin D levels. J. Crohn’s Colitis 2013, 7, 407–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabbani, T.A.; Koutroubakis, I.E.; Schoen, R.E.; Ramos-Rivers, C.; Shah, N.; Swoger, J.; Regueiro, M.; Barrie, A.; Schwartz, M.; Hashash, J.G.; et al. Association of Vitamin D level with clinical status in inflammatory bowel disease: A 5-year longitudinal study. Am. J. Gastroenterol. 2016, 111, 712–719. [Google Scholar] [CrossRef]
- Bartels, L.E.; Jørgensen, S.P.; Bendix, M.; Hvas, C.L.; Agnholt, J.; Agger, R.; Dahlerup, J.F. 25-Hydroxy vitamin D3 modulates dendritic cell phenotype and function in Crohn’s disease. Inflammopharmacology 2013, 21, 177–186. [Google Scholar] [CrossRef] [PubMed]
- Boonstra, A.; Barrat, F.J.; Crain, C.; Heath, V.L.; Savelkoul, H.F.; O’Garra, A. 1alpha,25-Dihydroxyvitamin d3 has a direct effect on naive CD4(+) T cells to enhance the development of Th2 cells. J. Immunol. 2001, 167, 4974–4980. [Google Scholar] [CrossRef] [Green Version]
- Bruce, D.; Yu, S.; Ooi, J.H.; Cantorna, M.T. Converging pathways lead to overproduction of IL-17 in the absence of vitamin D signaling. Int. Immunopharmacol. 2001, 23, 519–528. [Google Scholar] [CrossRef]
- Piechota-Polanczyk, A.; Fichna, J. The role of oxidative stress in pathogenesis and treatment of inflammatory bowel diseases. Naunyn Schmiedeberg’s Arch. Pharmacol. 2014, 387, 605–620. [Google Scholar] [CrossRef] [Green Version]
- Vaghari-Tabari, M.; Moein, S.; Qujeq, D.; Kashifard, M.; Hajian-Tilaki, K. Positive correlation of fecal calprotectin with serum antioxidant enzymes in patients with inflammatory bowel disease: Accidental numerical correlation or a new finding? Am. J. Med. Sci. 2018, 355, 449–455. [Google Scholar] [CrossRef]
- Siva, S.; Rubin, D.T.; Gulotta, G.; Wroblewski, K.; Pekow, J. Zinc deficiency is associated with poor clinical outcomes in patients with inflammatory bowel disease. Inflamm. Bowel Dis. 2016, 23, 152–157. [Google Scholar] [CrossRef] [Green Version]
- Alkhouri, R.H.; Hashmi, H.; Baker, R.D.; Gelfond, D.; Baker, S.S. Vitamin and mineral status in patients with inflammatory bowel disease. J. Pediatr. Gastroenterol. Nutr. 2013, 56, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Vaghari-Tabari, M.; Jafari-Gharabaghlou, D.; Sadeghsoltani, F.; Hassanpour, P.; Qujeq, D.; Rashtchizadeh, N.; Ghorbanihaghjo, A. Zinc and selenium in inflammatory bowel disease: Trace elements with key roles? Biol. Trace Elem. Res. 2020, 1–15. [Google Scholar] [CrossRef]
- Liu, J.; Rozovsky, S. Membrane-bound selenoproteins. Antioxid. Redox Signal. 2015, 23, 795–813. [Google Scholar] [CrossRef] [PubMed]
- Gîlcă-Blanariu, G.E.; Diaconescu, S.; Ciocoiu, M.; Ștefănescu, G. New insights into the role of trace elements in ibd. BioMed Res. Int. 2018, 2018, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Imamura, F.; O’Connor, L.; Ye, Z.; Mursu, J.; Hayashino, Y.; Bhupathiraju, S.N.; Forouhi, N.G. Consumption of sugar sweetened beverages, artificially sweetened beverages, and fruit juice and incidence of type 2 diabetes: Systematic review, meta-analysis, and estimation of population attributable fraction. Br. J. Sports Med. 2016, 50, 496–504. [Google Scholar] [CrossRef] [PubMed]
- Narain, A.; Kwok, C.S.; Mamas, M.A. Soft drinks and sweetened beverages and the risk of cardiovascular disease and mortality: A systematic review and meta-analysis. Int. J. Clin. Pract. 2016, 70, 791–805. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.K.; Curhan, G. Soft drinks, fructose consumption, and the risk of gout in men: Prospective cohort study. BMJ. 2008, 336, 309–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Te Morenga, L.A.; Howatson, A.J.; Jones, R.M.; Mann, J. Dietary sugars and cardiometabolic risk: Systematic review and meta-analyses of randomized controlled trials of the effects on blood pressure and lipids. Am. J. Clin. Nutr. 2014, 100, 65–79. [Google Scholar] [CrossRef] [Green Version]
- Maaz, A.; Anderson, A.J.; Dmitriy, B.; Ramos, R.C.; O’Keefe, S.J.; Regueiro, M.D. The association between added dietary sugars and inflammatory bowel disease severity. Gastroenterology 2018, 154, S622. [Google Scholar]
- De Palma, G.; Nadal, I.; Collado, M.C.; Sanz, Y. Effects of a gluten-free diet on gut microbiota and immune function in healthy adult human subjects. Br. J. Nutr. 2009, 102, 1154–1160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzo Pisarello, M.J.; Vintiñi, E.O.; González, S.N.; Pagani, F.; Medina, M.S. Decrease in lactobacilli in the intestinal microbiota of celiac children with a gluten-free diet, and selection of potentially probiotic strains. Can. J. Microbiol. 2015, 61, 32–37. [Google Scholar] [CrossRef]
- De Angelis, M.; Vannini, L.; Di Cagno, R.; Cavallo, N.; Minervini, F.; Francavilla, R.; Ercolini, D.; Gobbetti, M. Salivary and fecal microbiota and metabolome of celiac children under gluten-free diet. Int. J. Food Microbiol. 2016, 239, 125–132. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, F.; Pellegrini, N.; Vannini, L.; Jeffery, I.B.; La Storia, A.; Laghi, L.; Serrazanetti, D.I.; Di Cagno, R.; Ferrocino, I.; Lazzi, C.; et al. High-level adherence to a Mediterranean diet beneficially impacts the gut microbiota and associated metabolome. Gut 2016, 65, 1812–1821. [Google Scholar] [CrossRef] [PubMed]
- Chierico, F.; Del Vernocchi, P.; Dallapiccola, B.; Putignani, L. Mediterranean diet and health: Food effects on gut microbiota and disease control. Int. J. Mol. Sci. 2014, 15, 11678–11699. [Google Scholar] [CrossRef]
- Izaskun, G.M.; Marta, S.R.; Cristina, A.; Collado María, C. Shifts on gut microbiota associated to Mediterranean diet adherence and specific dietary intakes on general adult population. Front. Microbiol. 2018, 9, 890. [Google Scholar]
- Myles, I.A. Fast food fever: Reviewing the impacts of the Western diet on immunity. Nutr. J. 2014, 13, 61. [Google Scholar] [CrossRef] [Green Version]
- Zinöcker, M.K.; Lindseth, I.A. The Western diet-microbiome-host interaction and its role in metabolic disease. Nutrients 2018, 10, 365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef]
- West, N.R.; Powrie, F. Immunotherapy not working? Check your microbiota. Cancer Cell 2015, 28, 687–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.J.; Tang, B.; Wang, F.C.; Tang, L.; Lei, Y.Y.; Luo, Y.; Huang, S.J.; Yang, M.; Wu, L.Y.; Wang, W.; et al. Parthenolide ameliorates colon inflammation through regulating Treg/Th17 balance in a gut microbiota-dependent manner. Theranostics 2020, 10, 5225–5241. [Google Scholar] [CrossRef]
- Sivan, A.; Corrales, L.; Hubert, N.; Williams, J.B.; Aquino-Michaels, K.; Earley, Z.M.; Benyamin, F.W.; Lei, Y.M.; Jabri, B.; Alegre, M.L.; et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science 2015, 350, 1084–1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloyd-Price, J.; Arze, C.; Ananthakrishnan, A.N.; Schirmer, M.; Avila-Pacheco, J.; Poon, T.W.; Andrews, E.; Ajami, N.J.; Bonham, K.S.; Brislawn, C.J.; et al. Multi-omics of the gut microbial ecosystem in inflammatory bowel diseases. Nature 2019, 569, 655–662. [Google Scholar] [CrossRef]
- Vich Vila, A.; Imhann, F.; Collij, V.; Jankipersadsing, S.A.; Gurry, T.; Mujagic, Z.; Kurilshikov, A.; Bonder, M.J.; Jiang, X.; Tigchelaar, E.F.; et al. Gut microbiota composition and functional changes in inflammatory bowel disease and irritable bowel syndrome. Sci. Transl. Med. 2018, 10, 8914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, K.M.; Rank, K.M.; Vaughn, B.P.; Khoruts, A. Treatment of recurrent Clostridium difficile infection using fecal microbiota transplantation in patients with inflammatory bowel disease. Gut Microbes 2017, 8, 303–309. [Google Scholar] [CrossRef] [Green Version]
- Quince, C.; Lundin, E.E.; Andreasson, A.N.; Greco, D.; Rafter, J.; Talley, N.J.; Agreus, L.; Andersson, A.F.; Engstrand, L.; D’Amato, M. The impact of Crohn’s disease genes on healthy human gut microbiota: A pilot study. Gut 2013, 62, 952–954. [Google Scholar] [CrossRef]
- Shang, L.J.; Liu, H.B.; Dai, Z.Q.; Li, J.; Chen, M.X.; Zeng, X.F.; Qiao, S.Y. Microbial changes associated with IBD mouse model and microbiota transplantation confers colitis symptom in microbiota deletion mice. Curr. Pharm. Design 2020, 26, 1–30. [Google Scholar]
- Joossens, M.; Huys, G.; Cnockaert, M.; De Preter, V.; Verbeke, K.; Rutgeerts, P.; Vandamme, P.; Vermeire, S. Dysbiosis of the faecal microbiota in patients with Crohn’s disease and their unaffected relatives. Gut 2011, 60, 631–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, D.N.; St Amand, A.L.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-phylogenetic characterization of microbial community imbalances in human inflammatory bowel diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [Green Version]
- Sokol, H.; Lepage, P.; Seksik, P.; Dore, J.; Marteau, P. Temperature gradient gel electrophoresis of fecal 16s rRNA reveals active Escherichia coli in the microbiota of patients with ulcerative colitis. J. Clin. Microbiol. 2006, 44, 3172–3177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andoh, A.; Kuzuoka, H.; Tsujikawa, T.; Nakamura, S.; Hirai, F.; Suzuki, Y.; Matsui, T.; Fujiyama, Y.; Matsumoto, T. Multicenter analysis of fecal microbiota profiles in Japanese patients with Crohn’s disease. J. Gastroenterol. 2012, 47, 1298–1307. [Google Scholar] [CrossRef]
- Seksik, P.; Rigottier-Gois, L.; Gramet, G.; Sutren, M.; Pochart, P.; Marteau, P.; Jian, R.; Dore, J. Alterations of the dominant faecal bacterial groups in patients with Crohn’s disease of the colon. Gut 2003, 52, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Willing, B.; Halfvarson, J.; Dicksved, J.; Rosenquist, M.; Jarnerot, G.; Engstrand, L.; Tysk, C.; Jansson, J.K. Twin studies reveal specific imbalances in the mucosa-associated microbiota of patients with ileal Crohn’s disease. Inflamm. Bowel Dis. 2009, 15, 653–660. [Google Scholar] [CrossRef]
- Nazareth, N.; Magro, F.; Machado, E.; Ribeiro, T.G.; Martinho, A.; Rodrigues, P.; Alves, R.; Macedo, G.N.; Gracio, D.; Coelho, R.; et al. Prevalence of Mycobacterium avium subsp. Paratuberculosis and Escherichia coli in blood samples from patients with inflammatory bowel disease. Med. Microbiol. Immunol. 2015, 204, 681–692. [Google Scholar] [PubMed]
- Mahendran, V.; Riordan, S.M.; Grimm, M.C.; Tran, T.A.; Major, J.; Kaakoush, N.O.; Mitchell, H.; Zhang, L. Prevalence of campylobacter species in adult Crohn’s disease and the preferential colonization sites of campylobacter species in the human intestine. PLoS ONE 2011, 6, e25417. [Google Scholar] [CrossRef] [PubMed]
- Meehan, C.J.; Beiko, R.G. A phylogenomic view of ecological specialization in the Lachnospiraceae, a family of digestive tract-associated bacteria. Genome Biol. Evol. 2014, 6, 703–713. [Google Scholar] [CrossRef] [Green Version]
- Hood, L. Tackling the microbiome. Science 2012, 336, 1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de La Serre, C.B.; Ellis, C.L.; Lee, J.; Hartman, A.L.; Rutledge, J.C.; Raybould, H.E. Propensity to high-fat diet-induced obesity in rats is associated with changes in the gut microbiota and gut inflammation. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 299, 440–448. [Google Scholar] [CrossRef]
- Cani, P.D.; Bibiloni, R.; Knauf, C.; Waget, A.; Neyrinck, A.M.; Delzenne, N.M.; Burcelin, R. Changes in gut microbiota control metabolic endotoxemia-induced inflammation in high-fat diet-induced obesity and diabetes in mice. Diabetes 2008, 57, 1470–1481. [Google Scholar] [CrossRef] [Green Version]
- Baumgart, M.; Dogan, B.; Rishniw, M.; Weitzman, G.; Bosworth, B.; Yantiss, R.; Orsi, R.H.; Wiedmann, M.; McDonough, P.; Kim, S.G.; et al. Culture independent analysis of ileal mucosa reveals a selective increase in invasive Escherichia coli of novel phylogeny relative to depletion of Clostridiales in Crohn’s disease involving the ileum. ISME J. 2007, 1, 403–418. [Google Scholar] [CrossRef] [Green Version]
- Kotlowski, R.; Bernstein, C.N.; Sepehr, S.; Krause, D.O. High prevalence of Escherichia coli belonging to the B2+ D phylogenetic group in inflammatory bowel disease. Gut 2007, 56, 669–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, C.; Antolin, M.; Santos, J.; Torrejon, A.; Casellas, F.; Borruel, N.; Guarner, F.; Malagelada, J.R. Unstable composition of the fecal microbiota in ulcerative colitis during clinical remission. Am. J. Gastroenterol. 2008, 103, 643–648. [Google Scholar] [CrossRef]
- Peterson, D.A.; Frank, D.N.; Pace, N.R.; Gordon, J.I. Metagenomic approaches for defining the pathogenesis of inflammatory bowel diseases. Cell Host Microbe 2008, 3, 417–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, H.M.; Campbell, B.J.; Hart, C.A.; Mpofu, C.; Nayar, M.; Singh, R.; Englyst, H.; Williams, H.F.; Rhodes, J.M. Enhanced Escherichia coli adherence and invasion in Crohn’s disease and colon cancer. Gastroenterology 2004, 127, 80–93. [Google Scholar] [CrossRef]
- Darfeuille-Michaud, A.; Boudeau, J.; Bulois, P.; Neut, C.; Glasser, A.L.; Barnich, N.; Bringer, M.A.; Swidsinski, A.; Beaugerie, L.; Colombel, J.F. High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohn’s disease. Gastroenterology 2004, 127, 412–421. [Google Scholar] [CrossRef]
- Barnich, N.; Darfeuille-Michaud, A. Adherent-invasive Escherichia coli and Crohn’s disease. Curr. Opin. Gastroenterol. 2007, 23, 16–20. [Google Scholar] [CrossRef]
- Gathungu, G.; Zhang, Y.; Rowehl, L.; Frank, D.; Boedeker, E.; Parkinson, J.; Ellen, J. P-217 specific PCR assays using CRISPR genes for detection of AIEC in fecal samples. Inflamma. Bowel Dis. 2014, 20, 113. [Google Scholar]
- O’Brien, C.L.; Bringer, M.A.; Holt, K.E.; Gordon, D.M.; Dubois, A.L.; Barnich, N.; Darfeuille-Michaud, A.; Pavli, P. Comparative genomics of Crohn’s disease-associated adherent invasive Escherichia coli. Gut 2017, 66, 1382–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scanu, A.M.; Bull, T.J.; Cannas, S.; Sanderson, J.D.; Sechi, L.A.; Dettori, G.; Zanetti, S.; Hermon-Taylor, J. Mycobacterium avium subspecies paratuberculosis infection in cases of irritable bowel syndrome and comparison with Crohn’s disease and Johne’s disease: Common neural and immune pathogenicities. J. Clin. Microbiol. 2007, 45, 3883–3890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manning, E.J.; Collins, M.T. Mycobacterium avium subsp. paratuberculosis: Pathogen, pathogenesis and diagnosis. Rev. Sci. Tech. Int. Off. Epizoot. 2001, 20, 133–150. [Google Scholar] [CrossRef] [PubMed]
- Lund, B.M.; Gould, G.W.; Rampling, A.M. Pasteurization of milk and the heat resistance of Mycobacterium avium subsp. paratuberculosis: A critical review of the data. Int. J. Food Microbiol. 2002, 77, 135–145. [Google Scholar] [CrossRef]
- Petersson, J.; Schreiber, O.; Hansson, G.C.; Gendler, S.J.; Velcich, A.; Lundberg, J.O.; Roos, S.; Holm, L.; Phillipson, M. Importance and regulation of the colonic mucus barrier in a mouse model of colitis. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 300, G327–G333. [Google Scholar] [CrossRef] [Green Version]
- Lawley, T.D.; Walker, A.W. Intestinal colonization resistance. Immunology 2013, 138, 1–11. [Google Scholar] [CrossRef]
- Swann, J.; Wang, Y.; Abecia, L.; Costabile, A.; Tuohy, K.; Gibson, G.; Roberts, D.; Sidaway, J.; Jones, H.; Wilson, I.D.; et al. Gut microbiome modulates the toxicity of hydrazine: A metabonomic study. Mol. Biosyst. 2009, 5, 351–355. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Wang, Y.; Wang, P.; Wang, F. Intestinal barrier dysfunction in severe burn injury. Burns Trauma 2019, 7, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- König, J.; Wells, J.; Cani, P.D.; García-Ródenas, C.L.; MacDonald, T.; Mercenier, A.; Whyte, J.; Troost, F.; Brummer, R.J. Human intestinal barrier function in health and disease. Clin. Transl. Gastroen. 2016, 7, e196. [Google Scholar] [CrossRef] [PubMed]
- Sharma, U.; Olson, R.K.; Erhart, F.N.; Zhang, L.; Meng, J.; Segura, B.; Banerjee, S.; Sharma, M.; Saluja, A.K.; Ramakrishnan, S.; et al. Prescription opioids induce gut dysbiosis and exacerbate colitis in a murine model of inflammatory bowel disease. J. Crohn’s Colitis 2020, 14, 801–817. [Google Scholar] [CrossRef] [PubMed]
- Curciarello, R.; Canziani, K.E.; Docena, G.H.; Muglia, C.I. Contribution of non-immune cells to activation and modulation of the intestinal inflammation. Front. Immunol. 2019, 10, 647. [Google Scholar] [CrossRef] [Green Version]
- Rubio, C.A.; Schmidt, P.T. Severe defects in the macrophage barrier to gut microflora in inflammatory bowel disease and Colon Cancer. Anticancer Res. 2018, 38, 3811–3815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buttó, L.F.; Haller, D. Dysbiosis in intestinal inflammation: Cause or consequence. Int. J. Med. Microbiol. 2016, 306, 302–309. [Google Scholar] [CrossRef]
- Mcilroy, J.; Ianiro, G.; Mukhopadhya, I.; Hansen, R.; Hold, G.L. Review article: The gut microbiome in inflammatory bowel disease-avenues for microbial management. Aliment. Pharm. Ther. 2018, 47, 26–42. [Google Scholar] [CrossRef] [PubMed]
- Quevrain, E.; Maubert, M.A.; Michon, C.; Chain, F.; Marquant, R.; Tailhades, J.; Miquel, S.; Carlier, L.; Bermúdez-Humarán, L.G.; Pigneur, B.; et al. Identification of an anti-inflammatory protein from Faecalibacterium prausnitzii, a commensal bacterium deficient in Crohn’s disease. Gut 2015, 65, 415–425. [Google Scholar] [CrossRef] [Green Version]
- Coyne, M.J.; Reinap, B.; Lee, M.M.; Comstock, L.E. Human symbionts use a host-like pathway for surface fucosylation. Science 2005, 307, 1778–1781. [Google Scholar] [CrossRef]
- Blacher, E.; Levy, M.; Tatirovsky, E.; Elinav, E. Microbiome-modulated metabolites at the interface of host immunity. J. Immunol. 2017, 198, 572–580. [Google Scholar] [CrossRef] [Green Version]
- Levy, M.; Blacher, E.; Elinav, E. Microbiome, metabolites and host immunity. Curr. Opin. Microbiol. 2017, 35, 8–15. [Google Scholar] [CrossRef]
- Fenton, T.M.; Jørgensen, P.B.; Niss, K.; Rubin, S.; Mörbe, U.M.; Riis, L.B.; Da Silva, C.; Plumb, A.; Vandamme, J.; Jakobsen, H.L.; et al. Immune profiling of human gut-associated lymphoid tissue identifies a role for isolated lymphoid follicles in priming of region-specific immunity. Immunity 2020, 52, 557–570. [Google Scholar] [CrossRef] [PubMed]
- Desai, M.S.; Seekatz, A.M.; Koropatkin, N.M.; Kamada, N.; Hickey, C.A.; Wolter, M.; Pudlo, N.A.; Kitamoto, S.; Terrapon, N.; Muller, A.; et al. A dietary fiber-deprived gut microbiota degrades the colonic mucus barrier and enhances pathogen susceptibility. Cell 2016, 167, 1339–1353. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wrzosek, L.; Miquel, S.; Noordine, M.L.; Bouet, S.; Joncquel Chevalier-Curt, M.; Robert, V.; Philippe, C.; Bridonneau, C.; Cherbuy, C.; Robbe-Masselot, C.; et al. Bacteroides thetaiotaomicron and Faecalibacterium prausnitzii influence the production of mucus glycans and the development of goblet cells in the colonic epithelium of a gnotobiotic model rodent. BMC Biol. 2013, 11, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birchenough, G.M.; Nystrom, E.E.; Johansson, M.E.; Hansson, G.C. A sentinel goblet cell guards the colonic crypt by triggering Nlrp6-dependent Muc2 secretion. Science 2016, 352, 1535–1542. [Google Scholar] [CrossRef] [Green Version]
- Farrell, R.J.; La Mont, J.T. Microbial factors in inflammatory bowel disease. Gastroenterol. Clin. North Am. 2002, 31, 41–62. [Google Scholar]
- Swidsinski, A.; Ladhoff, A.; Pernthaler, A.; Swidsinski, S.; Loening-Baucke, V.; Ortner, M.; Weber, J.; Hoffmann, U.; Schreiber, S.; Dietel, M.; et al. Mucosal flora in inflammatory bowel disease. Gastroenterology 2002, 122, 44–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verma, R.; Verma, A.K.; Ahuja, V.; Paul, J. Real-Time analysis of mucosal flora in patients with inflammatory bowel disease in India. J. Clin. Microbiol. 2010, 48, 4279–4282. [Google Scholar] [CrossRef] [Green Version]
- Wine, E.; Griffiths, A.; Sherman, P. A model for bacterial-mediated changes of the mucosal barrier in IBDs: Campylobacter jejuni infection impairs the integrity of polarized epithelial cells. J. Pediatr. Gastr. Nutr. 2006, 43, 8. [Google Scholar] [CrossRef]
- Häsler, R.; Sheibani-Tezerji, R.; Sinha, A.; Barann, M.; Rehman, A.; Esser, D.; Aden, K.; Knecht, C.; Brandt, B.; Nikolaus, S.; et al. Uncoupling of mucosal gene regulation, mRNA splicing and adherent microbiota signatures in inflammatory bowel disease. Gut 2007, 66, 2087–2097. [Google Scholar] [CrossRef]
- Swidsinski, A.; Loening-Baucke, V.; Theissig, F.; Engelhardt, H.; Bengmark, S.; Koch, S.; Lochs, H.; Dörffel, Y. Comparative study of the intestinal mucus barrier in normal and inflamed colon. Gut 2007, 56, 343–350. [Google Scholar] [CrossRef]
- Chiodini, R.J.; Dowd, S.E.; Barron, J.N.; Galandiuk, S.; Davis, B.; Glassing, A. Transitional and temporal changes in the mucosal and submucosal intestinal microbiota in advanced Crohn’s disease of the terminal ileum. J. Med. Microbiol. 2018, 67, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Chiodini, R.J.; Dowd, S.E.; Chamberlin, W.M.; Galandiuk, S.; Davis, B.; Glassing, A. Microbial population differentials between mucosal and submucosal intestinal tissues in advanced Crohn’s disease of the ileum. PLoS ONE 2015, 10, e0134382. [Google Scholar] [CrossRef] [PubMed]
- Fyderek, K.; Strus, M.; Kowalska-Duplaga, K.; Gosiewski, T.; Wedrychowicz, A.; Jedynak-Wasowicz, U.; Sładek, M.; Pieczarkowski, S.; Adamski, P.; Kochan, P.; et al. Mucosal bacterial microflora and mucus layer thickness in adolescents with inflammatory bowel disease. World J. Gastroenterol. 2009, 15, 5287–5294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, K.; Kawamoto, S.; Maruya, M.; Fagarasan, S. GALT: Organization and dynamics leading to IgA synthesis. Adv. Immunol. 2010, 107, 153–185. [Google Scholar]
- Eberl, G.; Colonna, M.; Di Santo, J.P.; McKenzie, A.N. Innate lymphoid cells. Innate lymphoid cells: A new paradigm in immunology. Science 2015, 348, 6566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tum, A.P.; Hubbard, A.L. Transcytosis: Crossing cellular barriers. Physiol. Rev. 2003, 83, 871–932. [Google Scholar] [CrossRef]
- Lewkowicz, N.; Klink, M.; Mycko, M.P.; Lewkowicz, P. Neutrophil—CD4+CD25+ T regulatory cell interactions: A possible new mechanism of infectious tolerance. Immunobiology 2013, 218, 455–464. [Google Scholar] [CrossRef]
- Ise, W.; Nakamura, K.; Shimizu, N.; Goto, H.; Fujimoto, K.; Kaminogawa, S.; Hachimura, S. Orally tolerized T cells can form conjugates with APCs but are defective in immunological synapse formation. J. Immunol. 2005, 175, 829–838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezende, R.M.; Weiner, H.L. History and mechanisms of oral tolerance. Semin. Immunol. 2017, 30, 3–11. [Google Scholar] [CrossRef]
- Tangye, S.G.; Ma, C.S.; Brink, R.; Deenick, E.K. The good, the bad and the ugly-TFH cells in human health and disease. Nat. Rev. Immunol. 2013, 13, 412–426. [Google Scholar] [CrossRef]
- Qi, H. T follicular helper cells in space-time. Nat. Rev. Immunol. 2016, 16, 612–625. [Google Scholar] [CrossRef]
- Walker, J.A.; Barlow, J.L.; McKenzie, A.N. Innate lymphoid cells—How did we miss them? Nat. Rev. Immunol. 2013, 13, 75–87. [Google Scholar] [CrossRef]
- Round, J.L.; Mazmanian, S.K. Inducible Foxp3+ regulatory T cell development by a commensal bacterium of the intestinal microbiota. Proc. Natl Acad. Sci. USA 2010, 107, 12204–12209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atarashi, K.; Tanoue, T.; Ando, M.; Kamada, N.; Nagano, Y.; Narushima, S.; Suda, W.; Imaoka, A.; Setoyama, H.; Nagamori, T.; et al. Th17 cell induction by adhesion of microbes to intestinal epithelial cells. Cell 2015, 163, 367–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xavier, R.J.; Podolsky, D.K. Unravelling the pathogenesis of inflammatory bowel diseas. Nature 2007, 448, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Gaboriau-Routhiau, V.; Rakotobe, S.; Lécuyer, E.; Mulder, I.; Lan, A.; Bridonneau, C.; Rochet, V.; Pisi, A.; De Paepe, M.; Brandi, G.; et al. The key role of segmented filamentous bacteria in the coordinated maturation of gut helper T cell responses. Immunity 2009, 31, 677–689. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, I.I.; Atarashi, K.; Manel, N.; Brodie, E.L.; Shima, T.; Karaoz, U.; Wei, D.; Goldfarb, K.C.; Santee, C.A.; Lynch, S.V.; et al. Induction of intestinal Th17 cells by segmented filamentous bacteria. Cell 2009, 139, 485–498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirota, K.; Turner, J.E.; Villa, M.; Duarte, J.H.; Demengeot, J.; Steinmetz, O.M.; Stockinger, B. Plasticity of Th17 cells in Peyer’s patches is responsible for the induction of T cell-dependent IgA responses. Nat. Immunol. 2013, 14, 372–379. [Google Scholar] [CrossRef]
- Savino, W.; Dardenne, M.; Velloso, L.A.; Dayse Silva-Barbosa, S. The thymus is a common target in malnutrition and infection. Br. J. Nutr. 2007, 98, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Palm, N.W.; de Zoete, M.R.; Cullen, T.W.; Barry, N.A.; Stefanowski, J.; Hao, L.; Degnan, P.H.; Hu, J.; Peter, I.; Zhang, W.; et al. Immunoglobulin A coating identifies colitogenic bacteria in inflammatory bowel disease. Cell 2014, 158, 1000–1010. [Google Scholar] [CrossRef] [Green Version]
- Gaffen, S.L.; Hajishengallis, G. A new inflammatory cytokine on the block: Rethinking periodontal disease and the Th1/Th2 paradigm in the context of Th17 cells and IL-17. J. Dent. Res. 2008, 87, 817–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazmanian, S.K.; Liu, C.H.; Tzianabos, A.O.; Dennis, L.K. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell 2005, 122, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Chappert, P.; Bouladoux, N.; Naik, S.; Schwartz, R.H. Specific gut commensal flora locally alters T cell tuning to endogenous ligands. Immunity 2013, 38, 1198–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazmanian, S.K.; Round, J.L.; Kasper, D.L. A microbial symbiosis factor prevents intestinal inflammatory disease. Nature 2008, 453, 620–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atarashi, K.; Tanoue, T.; Oshima, K.; Suda, W.; Nagano, Y.; Nishikawa, H.; Fukuda, S.; Saito, T.; Narushima, S.; Hase, K.; et al. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature 2013, 500, 232–236. [Google Scholar] [CrossRef]
- Lightfoot, Y.L.; Selle, K.; Yang, T.; Goh, Y.J.; Sahay, B.; Zadeh, M.; Owen, J.L.; Colliou, N.; Li, E.; Johannssen, T.; et al. SIGNR3-dependent immune regulation by Lactobacillus acidophilus surface layer protein A in colitis. EMBO J. 2015, 34, 881–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dietary Strategy | Characteristics | Affected Species | References |
|---|---|---|---|
| Gluten-free diet | No gluten | Bifidobacterium↓ Lactobacillus↓ Enterobacteriaceae↑ Escherichia coli↑ Prevotella↓ | [61,62,63] |
| Mediterranean diet | High fiber, low red meat | Prevotella ↑ Lachnospira ↑ Bifidobacteria ↑ Lactobacillus ↑ Bacteroides ↑ Clostridium ↓ | [64,65,66] |
| Western diet | High animal fat, high animal protein | Bifidobacteria ↓ Lactobacillus↓ Bacteroides ↑ Enterobacteria ↑ | [67,68] |
| Samples | Type of Disease | Increased | Decreased | References |
|---|---|---|---|---|
| Fecal samples of 68 CD patients, 84 of their unaffected relatives and 55 matched healthy individuals | CD | Ruminococcus gnavus | Dialister invisus, Faecalibacterium prausnitzii, Bifidobacterium adolescentis | [78] |
| 190 tissue colon samples from CD, UC, and non-IBD control | UC, UD | Bacillus, Proteobacteria, Actinobacteria | Firmicutes and Bacteroidetes, Lachnospiraceae | [79] |
| 18 fecal samples from active UC patients and healthy control | UC | Active Escherichia coli | Biodiversity of active bacteria | [80] |
| Fecal samples from CD patients (n = 161) and healthy individuals (n = 121) | CD | Bacteroides, Prevotella, Proteus | Faecalibacterium, Fusobacterium, Eubacterium, Bifidobacterium, | [81] |
| 8 samples from active colonic CD patients and 16 from healthy volunteers | CD | Escherichia coli, microflora diversity | Clostridium coccoides, Bacteroides | [82] |
| Biopsies from 5 different locations between ileum and rectum in 10 twin pairs | CD | Escherichia coli | F. prausnitzii (in ileal CD) | [83] |
| Peripheral blood from 202 IBD patients, 24 non-IBD controls and 29 healthy individuals | IBD | Mycobacterium avium subsp. paratuberculosis, adherent-invasive Escherichia coli | No detection | [84] |
| 301 biopsies from between ileum and rectum of 15 CD, 13 UC and 33 healthy individuals | CD, UC | Campylobacter concisus | No detection | [85] |
| Animal Model/Clinic Data Analysis | Mucosal Bacteria | Outcome | References |
|---|---|---|---|
| 345 colonic biopsies samples from bowel inflammation patients (305) and controls (40) | High concentrations of bacteria attached to mucosa in patients, no translocation | Mucosal bacteria increased with the severity of inflammation | [121] |
| Biopsy specimens from 72 UC, 12 CD patients and 65 healthy controls | Harmful bacterial groups increased, while beneficial bacterial species declined | Components of mucosal flora changed in IBD patients | [122] |
| MDCK1 cells infected with Campylobacter jejuni | — | Monolayer integrity changed, affected tight junction protein ZO-1 distribution | [123] |
| Mucosal biopsies from CD, UC, disease and healthy controls (n = 63) | Active mucosa-attached microbiota changed in IBD patients | Altered inflammation status | [124] |
| 120 biopsies from controls (20), self-limiting colitis (SLC, 20), UC (20) and randomly individual (60) | Bacteria found within mucus in UC, SLC | Bacterial migration, adherence to and invasion of the mucosa | [125] |
| Mucosal and submucosal samples from CD and controls | 4 and 13 bacterial species were found within submucosa at the center and margin of disease | Changed bacteria might drive or inhibit certain organisms in CD | [126] |
| 35 ilea mucosal and submucosal tissues from CD patients (n = 20) and healthy controls (n = 15) | Ruminococcus spp., Oscillospira spp., Pseudobutyrivibrio spp., and Tumebacillus spp increased in subjacent submucosa | Bacterial migration to submucosal resulting from mucosal barrier injury | [127] |
| Intestinal biopsies from IBD patients (inflammation, non-inflammation) and from controls | Streptococcus spp. accounted for 80% in the inflamed mucosa of CD; mucus layer in the inflamed IBD patients was remarkably thinner | Streptococcus spp. increased with the severity of IBD | [128] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yao, Q.; Li, H.; Fan, L.; Zhang, Y.; Zhao, S.; Zheng, N.; Wang, J. Dietary Regulation of the Crosstalk between Gut Microbiome and Immune Response in Inflammatory Bowel Disease. Foods 2021, 10, 368. https://doi.org/10.3390/foods10020368
Yao Q, Li H, Fan L, Zhang Y, Zhao S, Zheng N, Wang J. Dietary Regulation of the Crosstalk between Gut Microbiome and Immune Response in Inflammatory Bowel Disease. Foods. 2021; 10(2):368. https://doi.org/10.3390/foods10020368
Chicago/Turabian StyleYao, Qianqian, Huiying Li, Linlin Fan, Yangdong Zhang, Shengguo Zhao, Nan Zheng, and Jiaqi Wang. 2021. "Dietary Regulation of the Crosstalk between Gut Microbiome and Immune Response in Inflammatory Bowel Disease" Foods 10, no. 2: 368. https://doi.org/10.3390/foods10020368
APA StyleYao, Q., Li, H., Fan, L., Zhang, Y., Zhao, S., Zheng, N., & Wang, J. (2021). Dietary Regulation of the Crosstalk between Gut Microbiome and Immune Response in Inflammatory Bowel Disease. Foods, 10(2), 368. https://doi.org/10.3390/foods10020368