

Combining Celery Oleoresin, Limonene and Rhamnolipid as New Strategy to Control Endospore-Forming Bacillus cereus



Abstract
:1. Introduction
2. Materials and Methods
2.1. Oils, Oleoresins and Biosurfactant
2.2. Mixture Stock Solutions
2.3. Microorganism
2.4. Identification of Celery Oleoresin Components by GC-MS
2.5. Determination of Minimal Inhibitory Concentration (MIC) and Minimal Bactericidal Concentration (MBC)
2.6. Time–Kill Assay
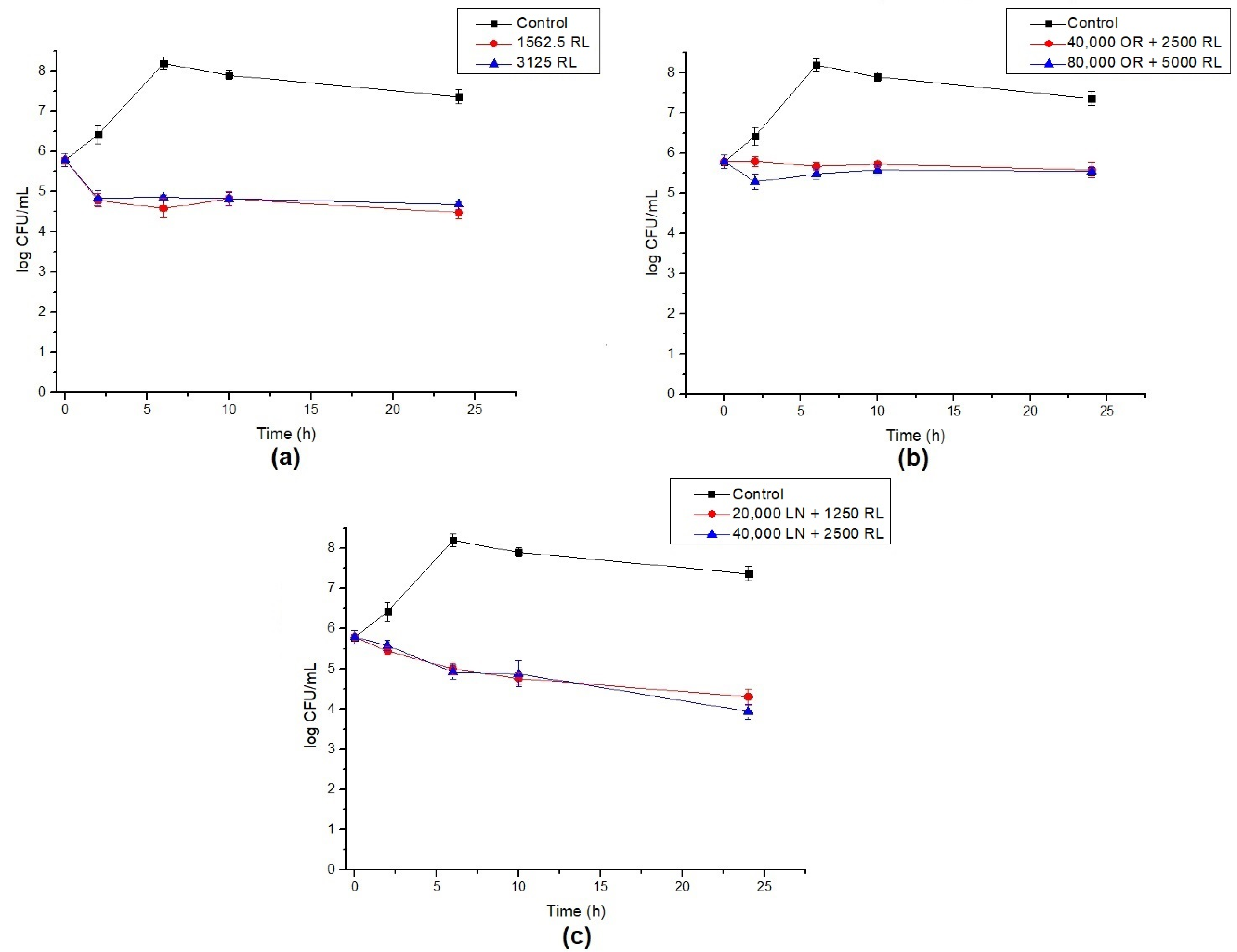
2.7. Endospore Germination Inhibition
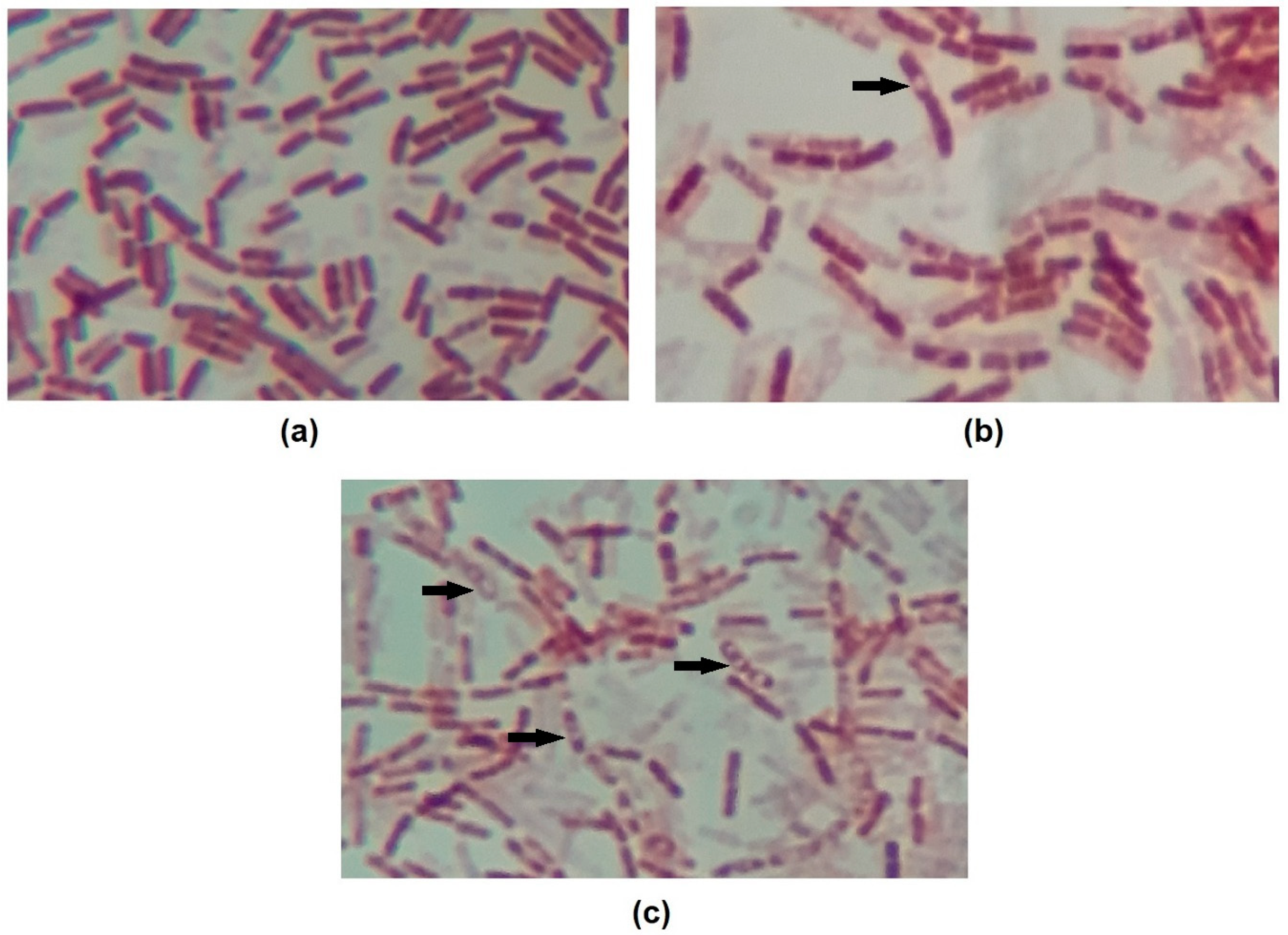
2.8. Endospore Staining
2.9. Statistics
3. Results and Discussion
3.1. Analysis of Celery Oleoresin
3.2. Minimal Inhibitory Concentration (MIC) and Minimal Bactericidal Concentration (MBC)
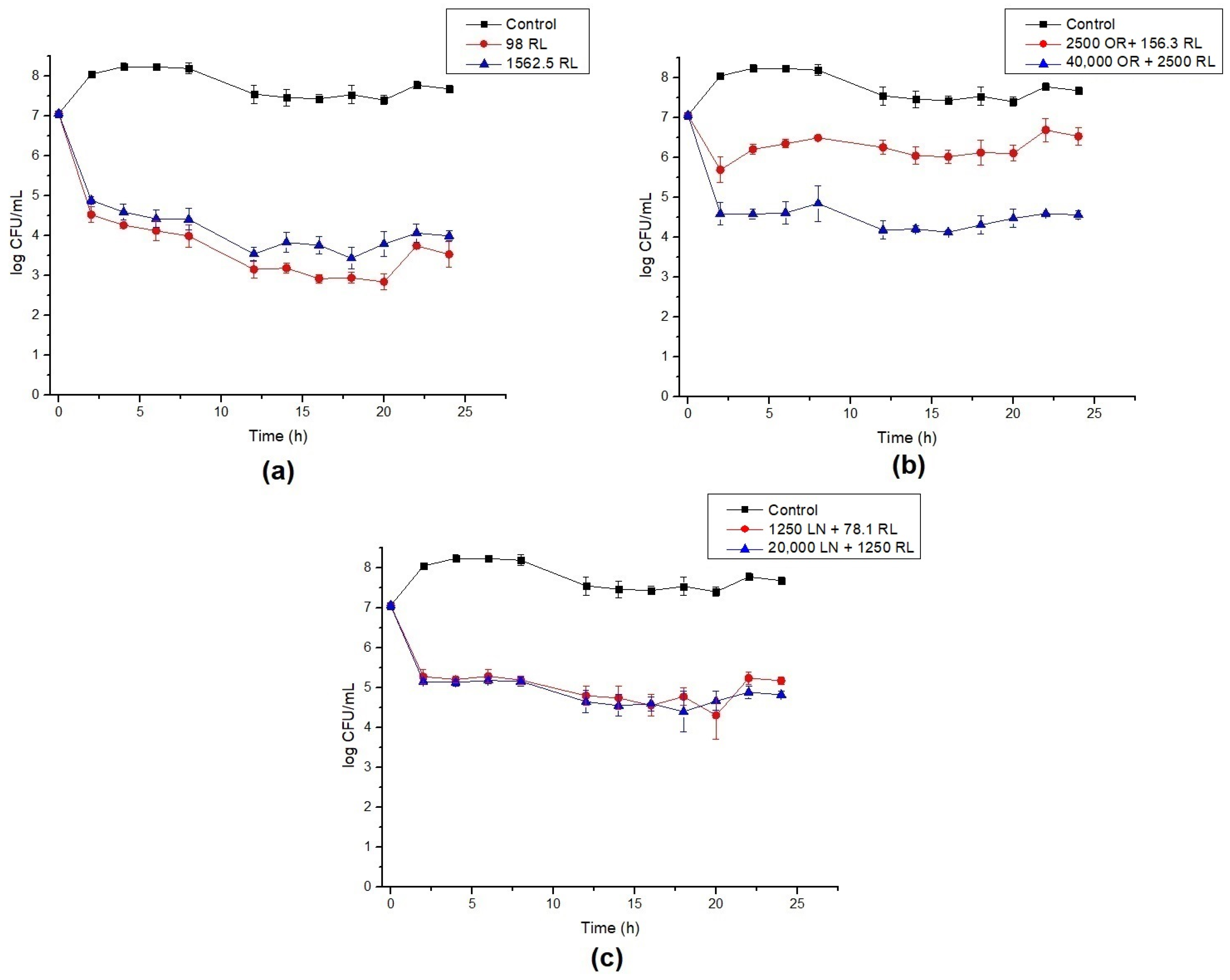
3.3. Time–Kill Assays
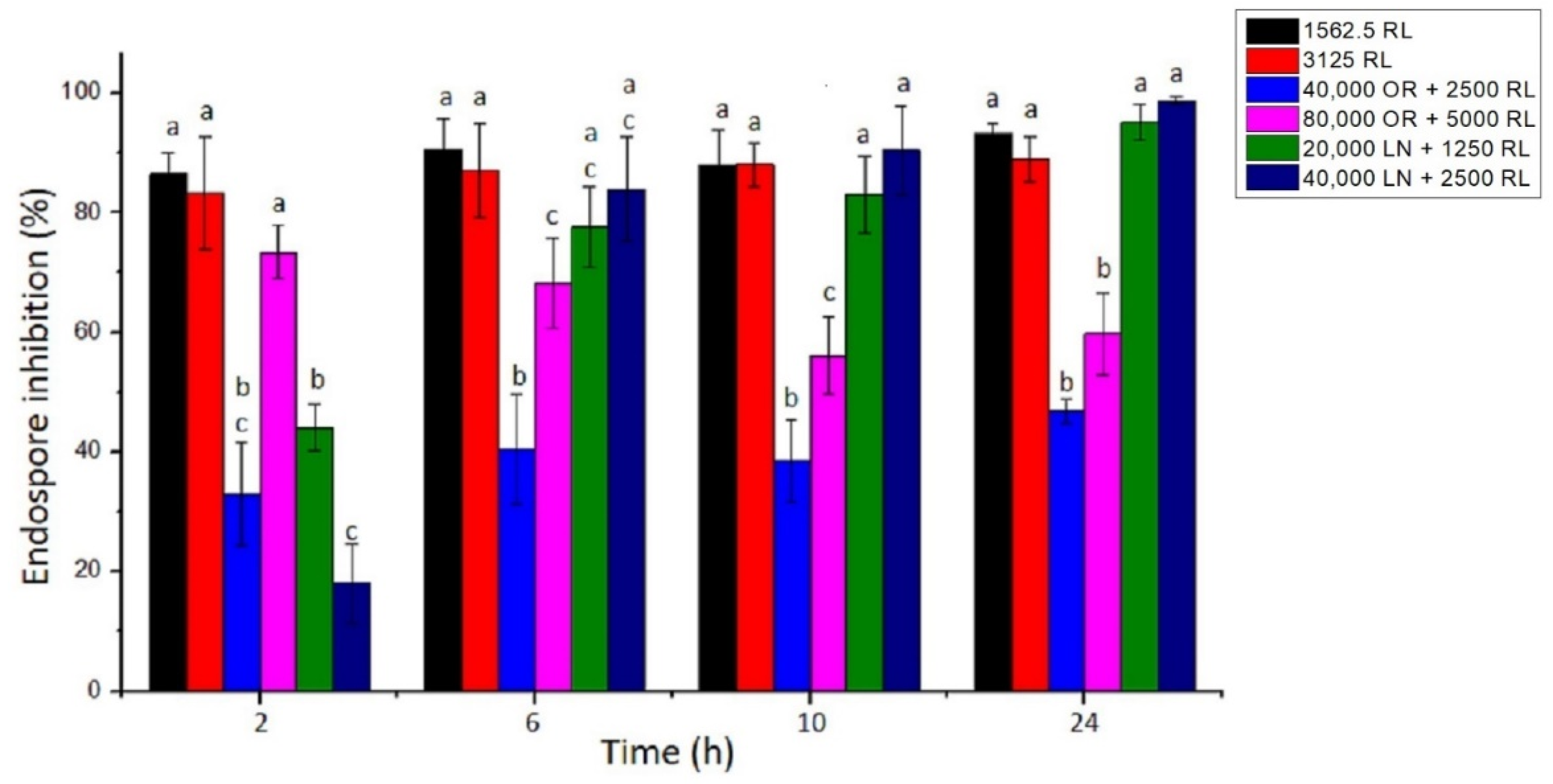
3.4. Endospore Inhibition
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Available online: http://www.who.int/mediacentre/news/releases/2015/foodborne-disease-estimates/en/ (accessed on 5 December 2020).
- World Health Organization. Fact Sheets: Food Safety. Available online: https://www.who.int/news-room/fact-sheets/detail/food-safety (accessed on 3 December 2020).
- Ministério da Saúde. Secretaria de Vigilância em Saúde. Departamento de Vigilância das Doenças Transmissíveis. Surtos de Doenças Transmitidas por Alimentos no Brasil. Available online: http://portalarquivos.saude.gov.br/images/pdf/2017/maio/29/Apresentacao-Surtos-DTA-2017.pdf (accessed on 3 December 2020).
- Carroll, L.M.; Wiedmann, M.; Mukherjee, M.; Nicholas, D.C.; Mingle, L.A.; Dumas, N.B.; Cole, J.A.; Kovac, J. Characterization of Emetic and Diarrheal Bacillus cereus Strains From a 2016 Foodborne Outbreak Using Whole-Genome Sequencing: Addressing the Microbiological, Epidemiological, and Bioinformatic Challenges. Front. Microbiol. 2019, 10, 144. [Google Scholar] [CrossRef]
- Glasset, B.; Herbin, S.; Guillier, L.; Cadel-Six, S.; Vignaud, M.-L.; Grout, J.; Pairaud, S.; Michel, V.; Hennekinne, J.-A.; RamaRao, N.; et al. Bacillus cereus-induced food-borne outbreaks in France, 2007 to 2014: Epidemiology and genetic characterisation. Eurosurveillance 2016, 21. [Google Scholar] [CrossRef]
- Carocho, M.; Morales, P.; Ferreira, I.C. Natural food additives: Quo vadis? Trends Food Sci. Technol. 2015, 45, 284–295. [Google Scholar] [CrossRef]
- Gray, J.A.; Chandry, P.S.; Kaur, M.; Kocharunchitt, C.; Bowman, J.P.; Fox, E.M. Novel Biocontrol Methods for Listeria monocytogenes Biofilms in Food Production Facilities. Front. Microbiol. 2018, 9, 1–12. [Google Scholar] [CrossRef]
- El Asbahani, A.; Miladi, K.; Badri, W.; Sala, M.; Addi, E.H.A.; Casabianca, H.; El Mousadik, A.; Hartmann, D.; Jilale, A.; Renaud, F.N.R.; et al. Essential oils: From extraction to encapsulation. Int. J. Pharm. 2015, 483, 220–243. [Google Scholar] [CrossRef]
- Swamy, M.K.; Akhtar, M.S.; Sinniah, U.R. Antimicrobial Properties of Plant Essential Oils against Human Pathogens and Their Mode of Action: An Updated Review. Evid.-Based Complement. Altern. Med. 2016, 2016, 1–21. [Google Scholar] [CrossRef]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential Oils in Food Preservation: Mode of Action, Synergies, and Interactions with Food Matrix Components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef]
- Okunowo, W.O.; Oyedeji, O.; Afolabi, L.O.; Matanmi, E. Essential Oil of Grape Fruit (Citrus paradisi) Peels and Its Antimicrobial Activities. Am. J. Plant Sci. 2013, 4, 1–9. [Google Scholar] [CrossRef]
- Gomes, M.S.; Cardoso, M.D.G.; Soares, M.J.; Batista, L.R.; Machado, S.M.F.; Andrade, M.A.; De Azeredo, C.M.O.; Resende, J.M.V.; Rodrigues, L.M.A. Use of Essential Oils of the Genus Citrus as Biocidal Agents. Am. J. Plant Sci. 2014, 5, 299–305. [Google Scholar] [CrossRef]
- Andrade, B.F.M.T.; Barbosa, L.N.; Probst, I.D.S.; Júnior, A.F. Antimicrobial activity of essential oils. J. Essent. Oil Res. 2014, 26, 34–40. [Google Scholar] [CrossRef]
- Sowbhagya, H.B. Chemistry, Technology, and Nutraceutical Functions of Celery (Apium graveolens L.): An Overview. Crit. Rev. Food Sci. Nutr. 2013, 54, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Arshad, H.; Ali, T.M.; Abbas, T.; Hasnain, A. Effect of Microencapsulation on Antimicrobial and Antioxidant Activity of Nutmeg Oleoresin Using Mixtures of Gum Arabic, OSA, and Native Sorghum Starch. Starch-Stärke 2018, 70, 1–12. [Google Scholar] [CrossRef]
- King, K. Packaging and storage of herbs and spices. In Handbook of Herbs and Spices; Peter, K.V., Ed.; Woodhead Publishing Series in Food Science, Technology and Nutrition; Woodhead Publishing: Cambridge, UK, 2006; Volume 3, pp. 86–102. ISBN 9781845690175. Available online: http://www.sciencedirect.com/science/article/pii/B9781845690175500051 (accessed on 26 November 2020).
- Lucera, A.; Costa, C.; Conte, A.; Del Nobile, M.A. Food applications of natural antimicrobial compounds. Front. Microbiol. 2012, 3, 287. [Google Scholar] [CrossRef] [PubMed]
- Nitschke, M.; E Silva, S.S. Recent food applications of microbial surfactants. Crit. Rev. Food Sci. Nutr. 2018, 58, 631–638. [Google Scholar] [CrossRef]
- Magalhães, L.; Nitschke, M. Antimicrobial activity of rhamnolipids against Listeria monocytogenes and their synergistic interaction with nisin. Food Control 2013, 29, 138–142. [Google Scholar] [CrossRef]
- De Araujo, L.V.; Guimarães, C.R.; Marquita, R.L.D.S.; Santiago, V.M.; De Souza, M.P.; Nitschke, M.; Freire, D.M.G. Rhamnolipid and surfactin: Anti-adhesion/antibiofilm and antimicrobial effects. Food Control 2016, 63, 171–178. [Google Scholar] [CrossRef]
- E Silva, S.; Carvalho, J.; Aires, C.; Nitschke, M. Disruption of Staphylococcus aureus biofilms using rhamnolipid biosurfactants. J. Dairy Sci. 2017, 100, 7864–7873. [Google Scholar] [CrossRef]
- Clements, T.; Ndlovu, T.; Khan, S.; Khan, W. Biosurfactants produced by Serratia species: Classification, biosynthesis, production and application. Appl. Microbiol. Biotechnol. 2019, 103, 589–602. [Google Scholar] [CrossRef]
- Marangon, C.A.; Martins, V.C.; Leite, P.M.; Santos, D.A.; Nitschke, M.; Plepis, A.M.G. Chitosan/gelatin/copaiba oil emulsion formulation and its potential on controlling the growth of pathogenic bacteria. Ind. Crop. Prod. 2017, 99, 163–171. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007; ISBN 978-1-932633-21-4. [Google Scholar]
- CLSI. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically; Approved Standard, 9th ed.; CLSI Document M07-A9; CLSI: Wayne, PA, USA, 2012; ISBN 1-56238-784-7. [Google Scholar]
- Verma, P. Methods for determining bactericidal activity and antimicrobial interactions: Synergy testing, time-kill curves, and population analysis. In Antimicrobial Susceptibility Testing Protocols; Schwalbe, R., Steele-Moore, L., Goodwin, A.C., Eds.; Taylor & Francis: Boca Raton, FL, USA, 2007; pp. 275–299. ISBN 9780429132346. [Google Scholar]
- Miles, A.A.; Misra, S.S.; Irwin, J.O. The estimation of the bactericidal power of the blood. Epidemiol. Infect. 1938, 38, 732–749. [Google Scholar] [CrossRef]
- Ali, B.; Lee, L.H.; Laskar, N.; Shaikh, N.; Tahir, H.; Hsu, S.D.; Newby, R.; Valsechi-Diaz, J.; Chu, T. Modified Green Tea Polyphenols, EGCG-S and LTP, Inhibit Endospore in Three Bacillus spp. Adv. Microbiol. 2017, 7, 175–187. [Google Scholar] [CrossRef]
- Schaeffer, A.B.; Fulton, M.D. A simplifified method of staining endospores. Science 1933, 77, 194. [Google Scholar] [CrossRef] [PubMed]
- Hamouda, T.; Shih, A.Y.; Baker, J.R., Jr. A rapid staining technique for the detection of the initiation of germination of bac-terial spores. Lett. Appl. Microbiol. 2002, 34, 86–90. [Google Scholar] [CrossRef]
- Ferreira, J.D.F.; Vieira, E.A.; Nitschke, M. The antibacterial activity of rhamnolipid biosurfactant is pH dependent. Food Res. Int. 2019, 116, 737–744. [Google Scholar] [CrossRef] [PubMed]
- Hassanen, N.H.M.; Eissa, A.M.F.; Hafez, S.A.M. Antioxidant and antimicrobial activity of celery (Apium graveolens) and coriander (Coriandrum sativum) herb and seed essential oils. Int. J. Curr. Microbiol. Aplp. Sci. 2015, 4, 284–296. [Google Scholar]
- Dąbrowska, J.A.; Kunicka-Styczyńska, A.; Śmigielski, K.B. Biological, chemical, and aroma profiles of essential oil from waste celery seeds (Apium graveolens L.). J. Essent. Oil Res. 2020, 32, 308–315. [Google Scholar] [CrossRef]
- Espina, L.; Gelaw, T.K.; De Lamo-Castellví, S.; Pagán, R.; García-Gonzalo, D. Mechanism of Bacterial Inactivation by (+)-Limonene and Its Potential Use in Food Preservation Combined Processes. PLoS ONE 2013, 8, e56769. [Google Scholar] [CrossRef]
- Ndlovu, T.; Rautenbach, M.; Vosloo, J.A.; Khan, S.; Khan, W. Characterisation and antimicrobial activity of biosurfactant extracts produced by Bacillus amyloliquefaciens and Pseudomonas aeruginosa isolated from a wastewater treatment plant. AMB Express 2017, 7, 1–19. [Google Scholar] [CrossRef]
- De Martino, L.; De Feo, V.; Fratianni, F.; Nazzaro, F. Chemistry, Antioxidant, Antibacterial and Antifungal Activities of Volatile Oils and their Components. Nat. Prod. Commun. 2009, 4, 1741–1750. [Google Scholar] [CrossRef] [PubMed]
- Guimarães, A.C.; Meireles, L.M.; Lemos, M.F.; Guimarães, M.C.C.; Endringer, D.C.; Fronza, M.; Scherer, R. Antibacterial Activity of Terpenes and Terpenoids Present in Essential Oils. Molecules 2019, 24, 2471. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, A.; Teruel, J.A.; Espuny, M.J.; Marqués, A.; Manresa, Á.; Aranda, F.J. Effects of dirhamnolipid on the structural properties of phosphatidylcholine membranes. Int. J. Pharm. 2006, 325, 99–107. [Google Scholar] [CrossRef]
- Sotirova, A.V.; Spasova, D.I.; Galabova, D.N.; Karpenko, E.; Shulga, A. Rhamnolipid–Biosurfactant Permeabilizing Effects on Gram-Positive and Gram-Negative Bacterial Strains. Curr. Microbiol. 2008, 56, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-H.; Cai, M.; Liu, Y.-S.; Sun, P.; Luo, S.-L. Antibacterial Activity and Mechanisms of Essential Oil from Citrus medica L. var. sarcodactylis. Molecules 2019, 24, 1577. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Lu, Z.; Bie, X.; Lü, F.; Zhao, H.; Yang, S. Optimization of inactivation of endospores of Bacillus cereus by antimicrobial lipopeptides from Bacillus subtilis fmbj strains using a response surface method. Appl. Microbiol. Biotechnol. 2006, 74, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Van Melis, C.; Almeida, C.B.; Kort, R.; Groot, M.N.; Abee, T. Germination inhibition of Bacillus cereus spores: Impact of the lipophilic character of inhibiting compounds. Int. J. Food Microbiol. 2012, 160, 124–130. [Google Scholar] [CrossRef]
- Cho, W.-I.; Cheigh, C.-I.; Hwang, H.-J.; Chung, M.-S. Sporicidal Activities of Various Surfactant Components against Bacillus subtilis Spores. J. Food Prot. 2015, 78, 1221–1225. [Google Scholar] [CrossRef]
- Cho, W.-I.; Chung, M.-S. Bacillus spores: A review of their properties and inactivation processing technologies. Food Sci. Biotechnol. 2020, 29, 1447–1461. [Google Scholar] [CrossRef]
- De Rienzo, M.D.; Stevenson, P.; Marchant, R.; Banat, I. Pseudomonas aeruginosa biofilm disruption using microbial surfactants. J. Appl. Microbiol. 2016, 120, 868–876. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Li, Y.; Ye, R. Effect of Alcohols on the Phase Behavior of Microemulsions Formed by a Biosurfactant—Rhamnolipid. J. Dispers. Sci. Technol. 2005, 26, 455–461. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| RT (min) 1 | Compound | % GC-MS | Exp RRI 2 | Lit RRI 3 | |
|---|---|---|---|---|---|
| 1 | 9.965 | limonene | 9.48 | 1039 | 1024 |
| 3 | 13.415 | pentyl ciclohexa-1,3-diene | 0.36 | 1158 | 1156 |
| 4 | 18.980 | 1-phenyl-1-pentanone | 0.57 | 1358 | 1364 |
| 5 | 20.705 | β-kariophylene | 0.23 | 1423 | 1417 |
| 6 | 22.455 | β-selinene | 3.11 | 1494 | 1489 |
| 7 | 22.615 | α-selinene | 0.77 | 1501 | 1498 |
| 8 | 23.190 | methyl dodecanoate | 0.06 | 1524 | -- |
| 11 | 24.725 | kariophylene oxide | 0.27 | 1586 | 1582 |
| 12 | 26.295 | 3-butylphthalide | 5.67 | 1655 | 1647 |
| 13 | 26.415 | β-eudesmol | 0.86 | 1661 | 1649 |
| 14 | 26.730 | 3Z-butylidenephthalide | 0.20 | 1675 | 1671 |
| 16 | 27.765 | sedanenolide | 7.41 | 1722 | 1719 |
| 17 | 27.915 | neocnidilide | 1.75 | 1728 | 1722 |
| 18 | 28.040 | Z-ligustilide | 0.34 | 1734 | 1734 |
| 19 | 29.980 | methyl pentadecanoate | 0.27 | 1815 | -- |
| 23 | 31.435 | methyl 7,10,13-hexadecatrienoate | 0.28 | 1859 | -- |
| 24 | 31.535 | methyl 7-hexadecenoate | 0.37 | 1862 | -- |
| 25 | 31.600 | methyl 9-hexadecenoate | 0.87 | 1865 | -- |
| 27 | 32.0.45 | methyl hexadecanoate | 8.23 | 1878 | -- |
| 28 | 32.745 | hexadecanoic acid | 0.08 | 1900 | -- |
| 31 | 33.985 | methyl heptadecanoate | 0.26 | 1947 | -- |
| 35 | 35.280 | methyl 9,12-octadecadienoate | 11.68 | 1996 | -- |
| 36 | 35.465 | methyl 9-octadecenoate | 30.50 | 2003 | -- |
| 37 | 35.875 | methyl octadecanoate | 3.58 | 2019 | -- |
| 38 | 36.115 | 9-octadecenoic acid | 1.03 | 2028 | -- |
| 40 | 38.880 | methyl 11-eicosenoate | 0.23 | 2134 | -- |
| 42 | 39.375 | methyl eicosanoate | 0.66 | 2152 | -- |
| 44 | 41.030 | methyl henicosanoate | 0.08 | 2216 | -- |
| 47 | 42.620 | methyl docosanoate | 0.56 | 2276 | -- |
| 48 | 44.155 | methyl tricosanoate | 0.18 | 2334 | -- |
| 52 | 45.630 | methyl tetracosanoate | 0.58 | 2391 | -- |
| 53 | 46.785 | trans-squalene | 0.14 | 2435 | -- |
| 54 | 47.065 | methyl pentacosanoate | 0.08 | 2445 | -- |
| 57 | 48.440 | methyl hexadocosanoate | 0.40 | 2498 | -- |
| 60 | 51.070 | methyl octacosanoate | 0.49 | 2598 | -- |
| 61 | 52.705 | β-stigmasterol | 0.51 | 2660 | -- |
| 63 | 54.045 | methyl triacontanoate | 0.16 | 2711 | -- |
| Identified | 92.30 | ||||
| Non-identified | 7.70 | ||||
| Fatty esters | 60.63 | ||||
| Total | 100.00 | ||||
| Compound | MIC (μg/mL) | MBC 24h (μg/mL) | MBC 48h (μg/mL) |
|---|---|---|---|
| RL | 98 | 1562.5 | 1562.5 |
| OR | 40,000 | >40,000 | >40,000 |
| OR + RL | 2500 OR + 156.3 RL | 20,000 OR + 1250 RL | >40,000 OR + 2500 RL |
| LN | 2500 | >40,000 | >40,000 |
| LN + RL | 1250 LN + 78.1 RL | 20,000 LN + 1250 RL | 20,000 LN + 1250 RL |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bertuso, P.d.C.; Mayer, D.M.D.; Nitschke, M. Combining Celery Oleoresin, Limonene and Rhamnolipid as New Strategy to Control Endospore-Forming Bacillus cereus. Foods 2021, 10, 455. https://doi.org/10.3390/foods10020455
Bertuso PdC, Mayer DMD, Nitschke M. Combining Celery Oleoresin, Limonene and Rhamnolipid as New Strategy to Control Endospore-Forming Bacillus cereus. Foods. 2021; 10(2):455. https://doi.org/10.3390/foods10020455
Chicago/Turabian StyleBertuso, Paula de Camargo, Débora M. Drappé Mayer, and Marcia Nitschke. 2021. "Combining Celery Oleoresin, Limonene and Rhamnolipid as New Strategy to Control Endospore-Forming Bacillus cereus" Foods 10, no. 2: 455. https://doi.org/10.3390/foods10020455
APA StyleBertuso, P. d. C., Mayer, D. M. D., & Nitschke, M. (2021). Combining Celery Oleoresin, Limonene and Rhamnolipid as New Strategy to Control Endospore-Forming Bacillus cereus. Foods, 10(2), 455. https://doi.org/10.3390/foods10020455