
Consumer Perceptions of Meat Redness Were Strongly Influenced by Storage and Display Times

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Sample Collection and Carcass Measures
2.3. Sire Breeding Values
2.4. Apprasel of Meat Colour
- Age group: 18–19 years, 20–25 years, 26–30 years, 31–39 years, 40–60 years, 61–70 years;
- Gender: male, female;
- Primary purchaser for the household: yes, no;
- Consumption frequency of red meat (including sheep meat, beef, and pork): daily, 4–5 times a week, 2–3 times a week, weekly, monthly, never.
2.5. Oxymyoglobin to Metmyoglobin Ratio
2.6. Statistical Analysis
3. Results
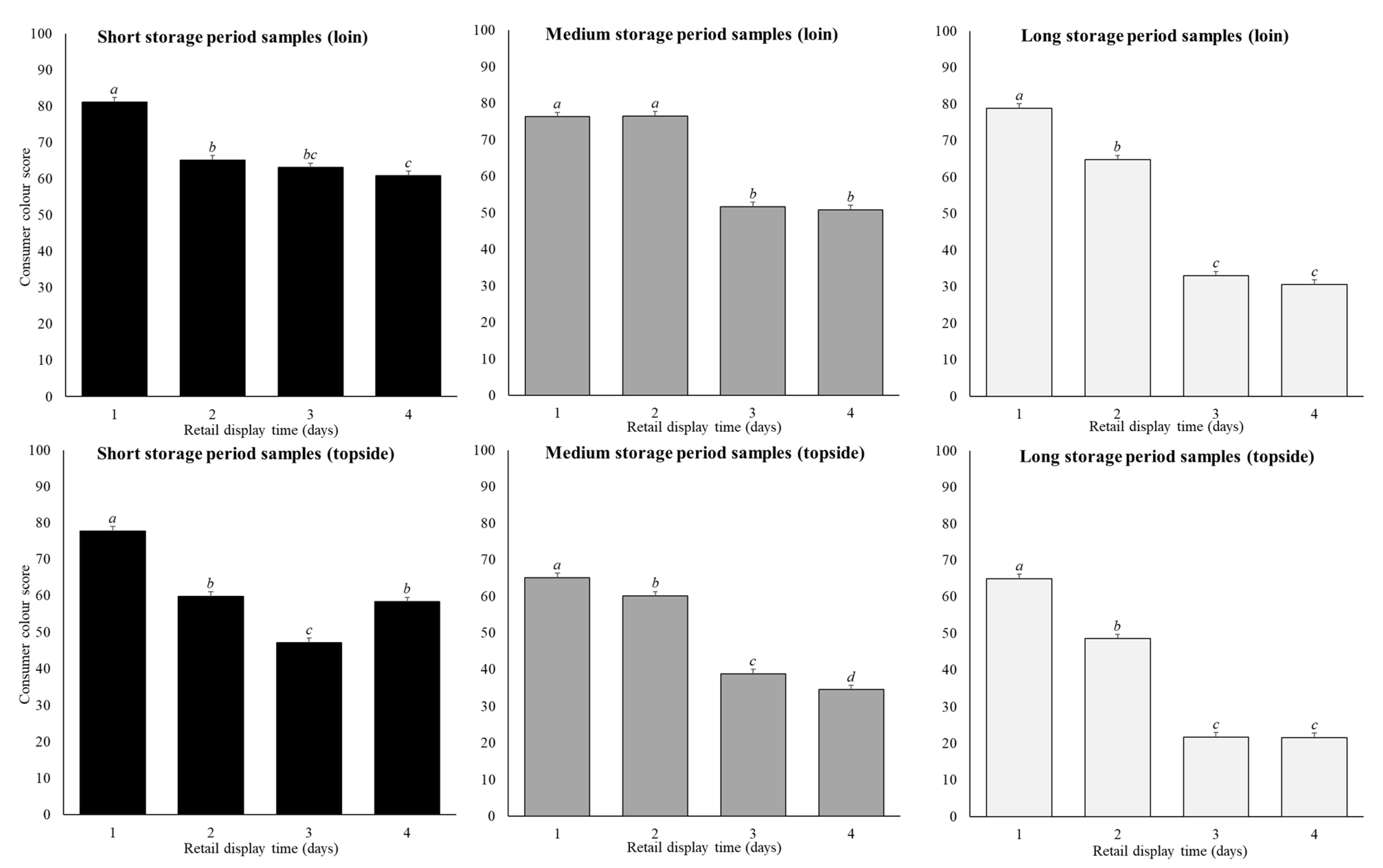
3.1. The Effect of Cut, Storage Period, and Retail Display Time on Consumer Colour Score
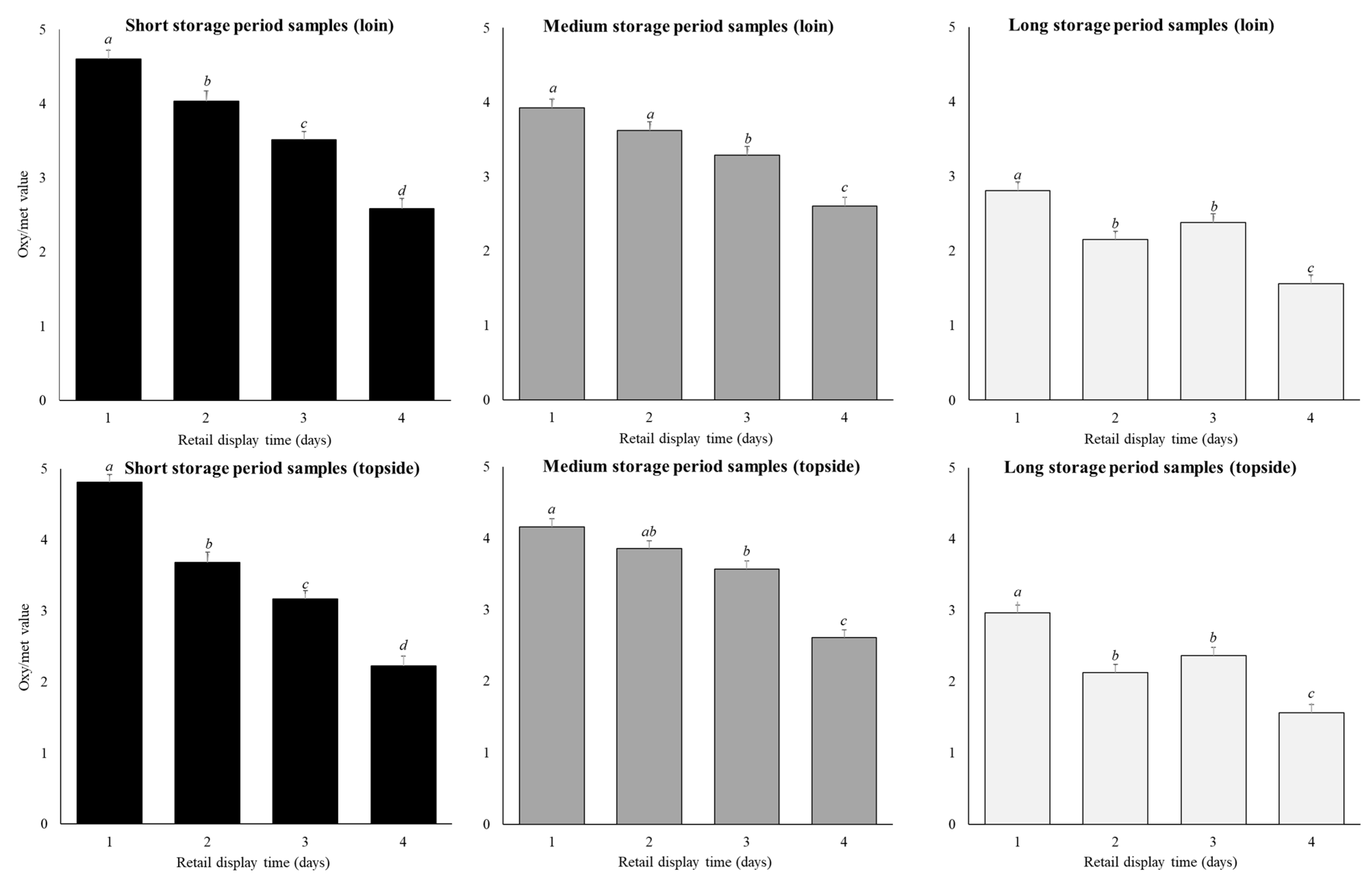
3.2. Oxymyoglobin to Metmyoglobin Ratio
3.3. Association of Carcass Measurements with Consumer Colour Score
3.4. Association of Sire ASBVs on Consumer Colour Score
3.5. Consumer Demographics
4. Discussion
4.1. Effect of Storage Time on Consumer Colour Score
4.2. Effect of Cut on Consumer Colour Score
4.3. Oxymyoglobin to Metmyoglobin Ratio
4.4. Carcass Measurements
4.5. Sire ASBVs
4.6. Industry Relevance
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ASBV | Australian Sheep Breeding Values |
| CCS | consumer colour scores |
| ICDH | isocitrate dehydrogenase |
| IMF | intramuscular fat percentage |
| Oxy/met | reflectance of light at wavelengths 630 nm and 580 nm |
| PEMD | post-weaning eye muscle depth |
| PFAT | post-weaning fat depth over the loin area |
| PWT | post-weaning weight |
References
- Faustman, C.; Cassens, R. The biochemical basis for discoloration in fresh meat: A review. J. Muscle Foods 1990, 1, 217–243. [Google Scholar] [CrossRef]
- Mancini, R.; Hunt, M. Current research in meat color. Meat Sci. 2005, 71, 100–121. [Google Scholar] [CrossRef]
- Renerre, M. Biochemical basis of fresh meat colour. In Proceedings of the International Congress of Meat Science and Technology, Yokohoma, Japan, 1–6 August 1999; pp. 344–353. [Google Scholar]
- Calnan, H.; Jacob, R.; Pethick, D.; Gardner, G. Factors affecting the colour of lamb meat from the longissimus muscle during display: The influence of muscle weight and muscle oxidative capacity. Meat Sci. 2014, 96, 1049–1057. [Google Scholar] [CrossRef]
- Channon, H.; Baud, S.; Walker, P. Modified atmosphere packaging improves retail display life of lamb cuts with variation between loin and knuckle. Anim. Prod. Sci. 2005, 45, 585–592. [Google Scholar] [CrossRef]
- Greenwood, P.; Gardner, G.; Hegarty, R. Lamb myofibre characteristics are influenced by sire estimated breeding values and pastoral nutritional system. Aust. J. Agric. Res. 2006, 57, 627–639. [Google Scholar] [CrossRef]
- Jose, C.; Pethick, D.; Gardner, G.; Jacob, R. The colour stability of aged lamb benefits from Vitamin E supplementation. In Proceedings of the 54th International Congress of Meat Science and Technology, Cape Town, South Africa, 10–15 August 2008; pp. 1–3. [Google Scholar]
- Hopkins, D.; Lamb, T.; Kerr, M.; Van de Ven, R.; Ponnampalam, E. Examination of the effect of ageing and temperature at rigor on colour stability of lamb meat. Meat Sci. 2013, 95, 311–316. [Google Scholar] [CrossRef] [PubMed]
- Moore, V.J.; Young, O.A. The effects of electrical stimulation, thawing, ageing and packaging on the colour and display life of lamb chops. Meat Sci. 1991, 131–145. [Google Scholar] [CrossRef]
- Jacob, R.; D’Antuono, M.; Smith, G.; Pethick, D.; Warner, R. Effect of lamb age and electrical stimulation on the colour stability of fresh lamb meat. Crop Pasture Sci. 2007, 58, 374–382. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Plozza, T.; Kerr, M.G.; Linden, N.; Mitchell, M.; Bekhit, A.E.-D.A.; Jacobs, J.L.; Hopkins, D.L. Interaction of diet and long ageing period on lipid oxidation and colour stability of lamb meat. Meat Sci. 2017, 129, 43–49. [Google Scholar] [CrossRef] [PubMed]
- Holman, B.W.B.; van de Ven, R.J.; Mao, Y.; Coombs, C.E.O.; Hopkins, D.L. Using instrumental (CIE and reflectance) measures to predict consumers’ acceptance of beef colour. Meat Sci. 2017, 127, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Khliji, S.; van de Ven, R.; Lamb, T.A.; Lanza, M.; Hopkins, D.L. Relationship between consumer ranking of lamb colour and objective measures of colour. Meat Sci. 2010, 85, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Anderson, F.; Pethick, D.; Gardner, G. The impact of genetics on retail meat value in Australian lamb. Meat Sci. 2016, 117, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Pethick, D.W.; Ball, A.J.; Banks, R.G.; Hocquette, J.F. Current and future issues facing red meat quality in a competitive market and how to manage continuous improvement. Anim. Prod. Sci. 2011, 51, 13–18. [Google Scholar] [CrossRef]
- Hopkins, D.; Hegarty, R.; Walker, P.; Pethick, D. Relationship between animal age, intramuscular fat, cooking loss, pH, shear force and eating quality of aged meat from sheep. Aust. J. Exp. Agric. 2006, 46, 879–884. [Google Scholar] [CrossRef]
- Pannier, L.; Gardner, G.; Pearce, K.; McDonagh, M.; Ball, A.; Jacob, R.; Pethick, D. Associations of sire estimated breeding values and objective meat quality measurements with sensory scores in Australian lamb. Meat Sci. 2014, 96, 1076–1087. [Google Scholar] [CrossRef]
- Kelman, K.; Pannier, L.; Pethick, D.; Gardner, G. Selection for lean meat yield in lambs reduces indicators of oxidative metabolism in the longissimus muscle. Meat Sci. 2014, 96, 1058–1067. [Google Scholar] [CrossRef] [PubMed]
- Gardner, G.; Pethick, D.; Hopkins, D.; Hegarty, R.; Cake, M.; Boyce, M.; Allingham, P. The impact of carcase estimated breeding values on yield and quality of sheep meat. Int. J. Sheep Wool Sci. 2006, 54, 33–39. [Google Scholar]
- Calnan, H.; Jacob, R.; Pethick, D.; Gardner, G. Selection for intramuscular fat and lean meat yield will improve the bloomed colour of Australian lamb loin meat. Meat Sci. 2017, 131, 187–195. [Google Scholar] [CrossRef] [Green Version]
- Gardner, G.; Hopkins, D.; Greenwood, P.; Cake, M.; Boyce, M.; Pethick, D. Sheep genotype, age and muscle type affect the expression of metabolic enzyme markers. Aust. J. Exp. Agric. 2007, 47, 1180–1189. [Google Scholar] [CrossRef]
- Ponnampalam, E.N.; Butler, K.L.; Jacob, R.H.; Pethick, D.W.; Ball, A.J.; Edwards, J.E.H.; Geesink, G.; Hopkins, D.L. Health beneficial long chain omega-3 fatty acid levels in Australian lamb managed under extensive finishing systems. Meat Sci. 2014, 96, 1104–1110. [Google Scholar] [CrossRef]
- Van der Werf, J.; Kinghorn, B.; Banks, R. Design and role of an information nucleus in sheep breeding programs. Anim. Prod. Sci. 2010, 50, 998–1003. [Google Scholar] [CrossRef]
- Pearce, K.; Van De Ven, R.; Mudford, C.; Warner, R.; Hocking-Edwards, J.; Jacob, R.; Pethick, D.; Hopkins, D. Case studies demonstrating the benefits on pH and temperature decline of optimising medium-voltage electrical stimulation of lamb carcasses. Anim. Prod. Sci. 2010, 50, 1107–1114. [Google Scholar] [CrossRef]
- AUS-MEAT. Handbook of Australian Meat; AUS-MEAT: Sydney, Australia, 2005. [Google Scholar]
- Perry, D.; Shorthose, W.; Ferguson, D.; Thompson, J. Methods used in the CRC program for the determination of carcass yield and beef quality. Aust. J. Exp. Agric. 2001, 41, 953–957. [Google Scholar] [CrossRef] [Green Version]
- MLA. Australian Lamb Product Guide. Meat and Livestock Australia. North Sydney, Australia. Available online: https://www.trueaussiebeefandlamb.com.au/globalassets/true-aussie---template/protein/lamb/lamb-product-guide-global.pdf (accessed on 3 February 2021).
- Fogarty, N. Genetic parameters for live weight, fat and muscle measurements, wool production and reproduction in sheep: A review. Anim. Breed. Abstr. 1995, 63, 101–143. [Google Scholar]
- Hall, D.; Gilmour, A.; Fogarty, N.; Holst, P. Growth and carcass composition of second-cross lambs. 2. Relationship between estimated breeding values of sires and their progeny performance under fast and slow growth regimes. Aust. J. Agric. Res. 2002, 53, 1341–1348. [Google Scholar] [CrossRef]
- Hunt, M. Meat Colour Measurements. In Proceedings of the Reciprocal Meat Conference, West Lafayette, IN, USA, 22–25 June 1980; pp. 41–46. [Google Scholar]
- Seyfert, M.; Mancini, R.A.; Hunt, M.C.; Tang, J.; Faustman, C.; Garcia, M. Color stability, reducing activity, and cytochrome c oxidase activity of five bovine muscles. J. Agric. Food Chem. 2006, 54, 8919–8925. [Google Scholar] [CrossRef]
- Wulf, D.; Morgan, J.; Sanders, S.; Tatum, J.; Smith, G.; Williams, S. Effects of dietary supplementation of vitamin E on storage and caselife properties of lamb retail cuts. J. Anim. Sci. 1995, 73, 399–405. [Google Scholar] [CrossRef]
- MacDougall, D.; Allen, R. Meat Chilling—Why and How? In Proceedings of the Meat Research Institute Symposium, Bristol, UK, 1972; p. 8. [Google Scholar]
- Tang, J.; Faustman, C.; Hoagland, T.A.; Mancini, R.A.; Seyfert, M.; Hunt, M.C. Postmortem oxygen consumption by mitochondria and its effects on myoglobin form and stability. J. Agric. Food Chem. 2005, 53, 1223–1230. [Google Scholar] [CrossRef] [PubMed]
- Ponnampalam, E.N.; Butler, K.L.; Burnett, V.F.; McDonagh, M.B.; Jacobs, J.L.; Hopkins, D.L. Aged vacuum packaged lamb cuts are less brown than fresh muscle cuts under simulated retail display. Food Nutr. Sci. 2013, 4, 147. [Google Scholar] [CrossRef] [Green Version]
- MSA. Meat Standards Australia—Sheepmeat. Meat and Livestock Australia. North Sydney, Australia. Available online: https://www.mla.com.au/globalassets/mla-corporate/marketing-beef-and-lamb/documents/meat-standards-australia/msa-sheep-brochure_web.pdf (accessed on 4 March 2021).
- Jose, C.; Jacob, R.; Pethick, D.; Gardner, G. Short term supplementation rates to optimise vitamin E concentration for retail colour stability of Australian lamb meat. Meat Sci. 2016, 111, 101–109. [Google Scholar] [CrossRef] [Green Version]
- Shorthose, W.R.; Harris, P.V.; Hopkins, A.F.; Kingston, O.L. Relationships between some objective properties of beef and consumer perceptions of meat quality. In Proceedings of the 34th International Congress of Meat Science and Technology, Brisbane, Australia, 29 August–2 September 1988; pp. 667–669. [Google Scholar]
- Hopkins, D. Assessment of lamb meat colour. Meat Focus Int. 1996, 5, 400–401. [Google Scholar]
- Morrissey, E.; Jacob, R.; Pluske, J. Perception of red brown colour by consumers. In Proceedings of the 54th International Congress of Meat Science and Technology, Cape Town, South Africa, 10–15 August 2008; pp. 1–3. [Google Scholar]
- Pannier, L.; Pethick, D.; Geesink, G.; Ball, A.; Jacob, R.; Gardner, G. Intramuscular fat in the longissimus muscle is reduced in lambs from sires selected for leanness. Meat Sci. 2014, 96, 1068–1075. [Google Scholar] [CrossRef]
- Pethick, D.; Harper, G.; Oddy, V. Growth, development and nutritional manipulation of marbling in cattle: A review. Aust. J. Exp. Agric. 2004, 44, 705–715. [Google Scholar] [CrossRef]
- Mezgebo, G.; Monahan, F.J.; McGee, M.; O’riordan, E.; Picard, B.; Richardson, R.; Moloney, A. Biochemical and organoleptic characteristics of muscle from early and late maturing bulls in different production systems. Animal 2017, 11, 1636–1644. [Google Scholar] [CrossRef] [Green Version]
- Huisman, A.; Brown, D. Genetic parameters for bodyweight, wool, and disease resistance and reproduction traits in Merino sheep. 2. Genetic relationships between bodyweight traits and other traits. Aust. J. Exp. Agric. 2008, 48, 1186–1193. [Google Scholar] [CrossRef]
- Suzuki, A.; Cassens, R. A histochemical study of myofiber types in the serratus ventralis thoracis muscle of sheep during growth. J. Anim. Sci. 1983, 56, 1447–1458. [Google Scholar] [CrossRef]
- Wegner, J.; Albrecht, E.; Fiedler, I.; Teuscher, F.; Papstein, H.-J.; Ender, K. Growth-and breed-related changes of muscle fiber characteristics in cattle. J. Anim. Sci. 2000, 78, 1485–1496. [Google Scholar] [CrossRef]
- Jacob, R. Implications of the variation in bloom properties of red meat: A review. Meat Sci. 2020, 162, 108040. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Mean | Standard Deviation | Range |
|---|---|---|---|
| Carcass measurements | |||
| Intramuscular fat (%) | 3.65 | 0.738 | 2.45–5.97 |
| Hot carcass weight (kg) | 20.11 | 1.742 | 16.70–25.50 |
| GR tissue depth (mm) | 6.68 | 2.011 | 3.00–11.00 |
| Loin fat weight (g) | 128.56 | 37.575 | 55.00–224.00 |
| Loin weight (g) | 308.37 | 49.078 | 223.00–491.00 |
| Loin eye muscle area (cm2) | 15.15 | 2.401 | 9.74–21.89 |
| Sire Australian Sheep Breeding Values | |||
| Post-weaning weight (kg) | 13.42 | 2.191 | 8.94–17.91 |
| Post-weaning eye muscle depth (mm) | 2.09 | 1.242 | −0.36–5.21 |
| Post-weaning fat depth (mm) | −0.53 | 0.703 | −2.00–1.06 |
| Sire intramuscular fat (%) | −0.61 | 0.483 | −2.01–0.22 |
| Effect | NDF | Consumer Colour Scores F-Value | Oxy/Met F-Value |
|---|---|---|---|
| Storage period | 2 | 224.97 * | 95.34 * |
| Cut | 1 | 175.52 * | ns |
| Retail display time | 3 | 1748.44 * | 511.17 * |
| Storage period * Cut | 2 | 75.65 * | 59.13 * |
| Cut * Retail display time | 3 | 16.86 * | 21.77 * |
| Storage period * Retail display time | 6 | 150.8 * | 43.21 * |
| Storage period * Cut * Retail display time | 6 | 26.91 * | 8.86 * |
| Age | Percentage (and Number) of Consumers |
| 18–19 years | 21.9 (192) |
| 20–25 years | 41.0 (360) |
| 26–30 years | 11.3 (99) |
| 31–39 years | 8.0 (70) |
| 40–60 years | 12.6 (111) |
| 61–70 years | 5.2 (46) |
| Gender | |
| Female | 59.5 (522) |
| Male | 40.5 (356) |
| Primary purchaser for household | |
| Yes | 61.5 (540) |
| No | 38.5 (338) |
| Red meat consumption frequency | |
| Daily | 11.2 (98) |
| 4–5 times a week | 28.0 (246) |
| 2–3 times a week | 36.3 (319) |
| Weekly | 15.0 (132) |
| Monthly | 6.8 (60) |
| Never | 2.6 (23) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corlett, M.T.; Pethick, D.W.; Kelman, K.R.; Jacob, R.H.; Gardner, G.E. Consumer Perceptions of Meat Redness Were Strongly Influenced by Storage and Display Times. Foods 2021, 10, 540. https://doi.org/10.3390/foods10030540
Corlett MT, Pethick DW, Kelman KR, Jacob RH, Gardner GE. Consumer Perceptions of Meat Redness Were Strongly Influenced by Storage and Display Times. Foods. 2021; 10(3):540. https://doi.org/10.3390/foods10030540
Chicago/Turabian StyleCorlett, Maddison T., David W. Pethick, Khama R. Kelman, Robin H. Jacob, and Graham E. Gardner. 2021. "Consumer Perceptions of Meat Redness Were Strongly Influenced by Storage and Display Times" Foods 10, no. 3: 540. https://doi.org/10.3390/foods10030540
APA StyleCorlett, M. T., Pethick, D. W., Kelman, K. R., Jacob, R. H., & Gardner, G. E. (2021). Consumer Perceptions of Meat Redness Were Strongly Influenced by Storage and Display Times. Foods, 10(3), 540. https://doi.org/10.3390/foods10030540